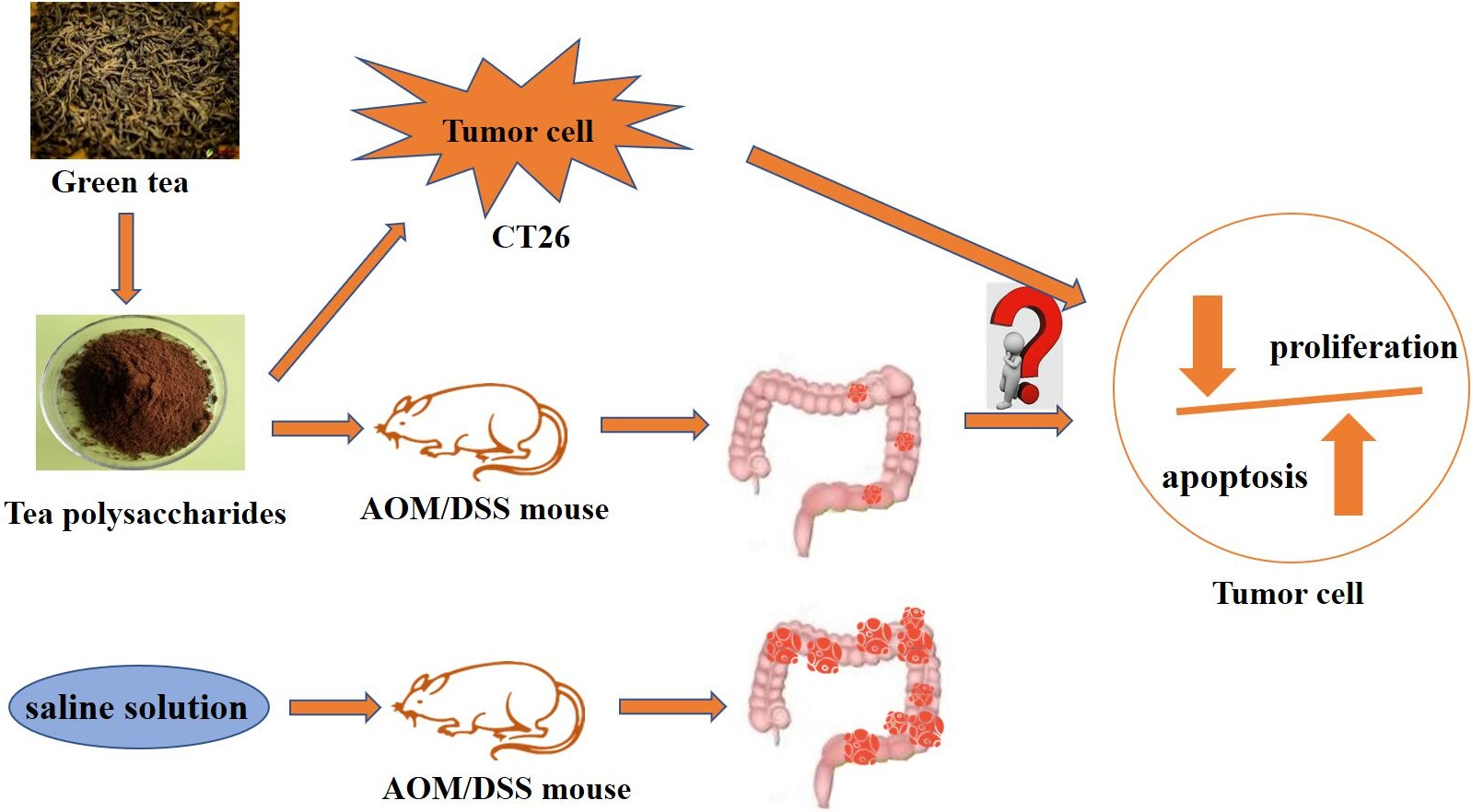
Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells

 ,
,
Abstract
:
1. Introduction
2. Results
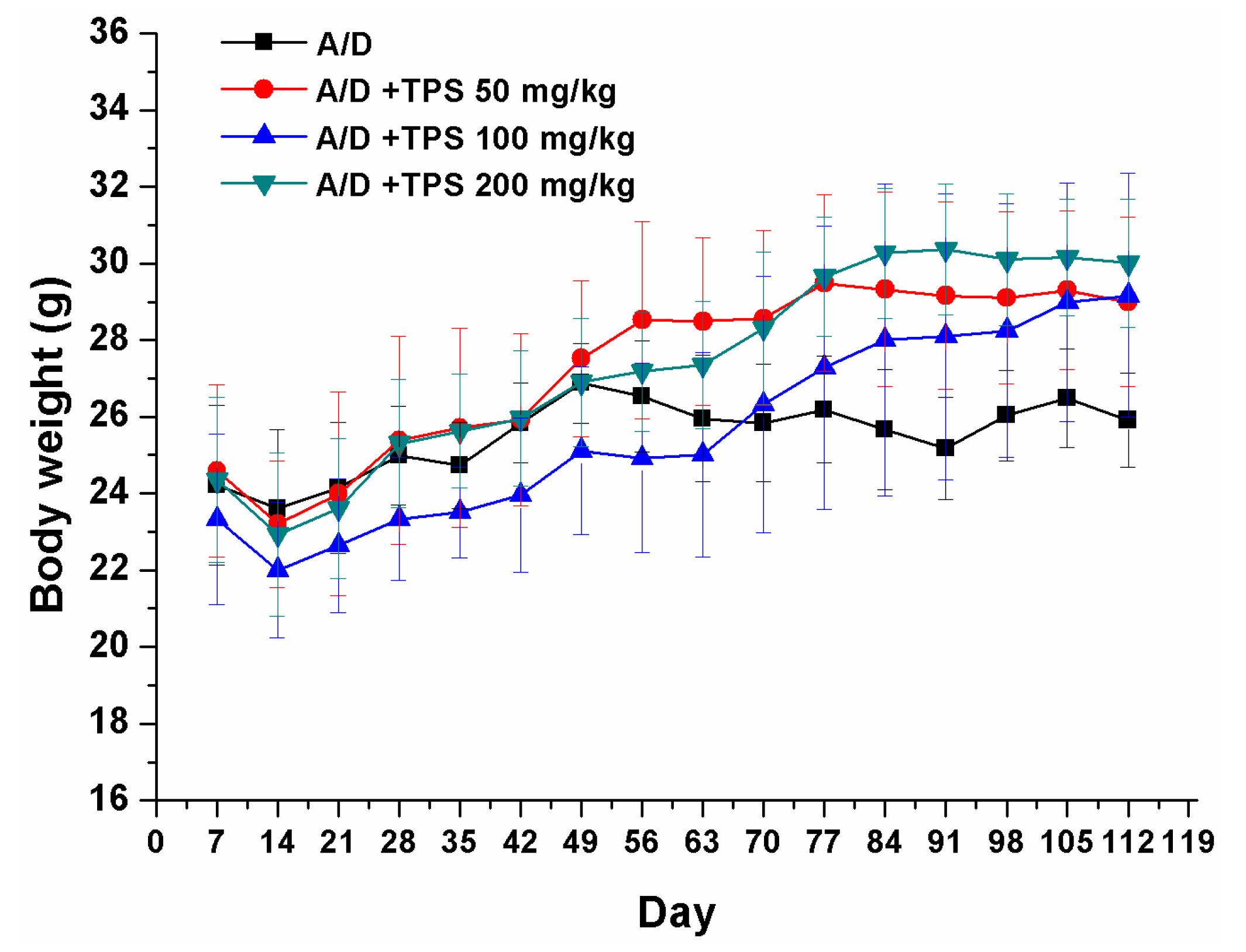
2.1. TPS Inhibited the Progression of Colitis Associated Colorectal (CAC) Cancer
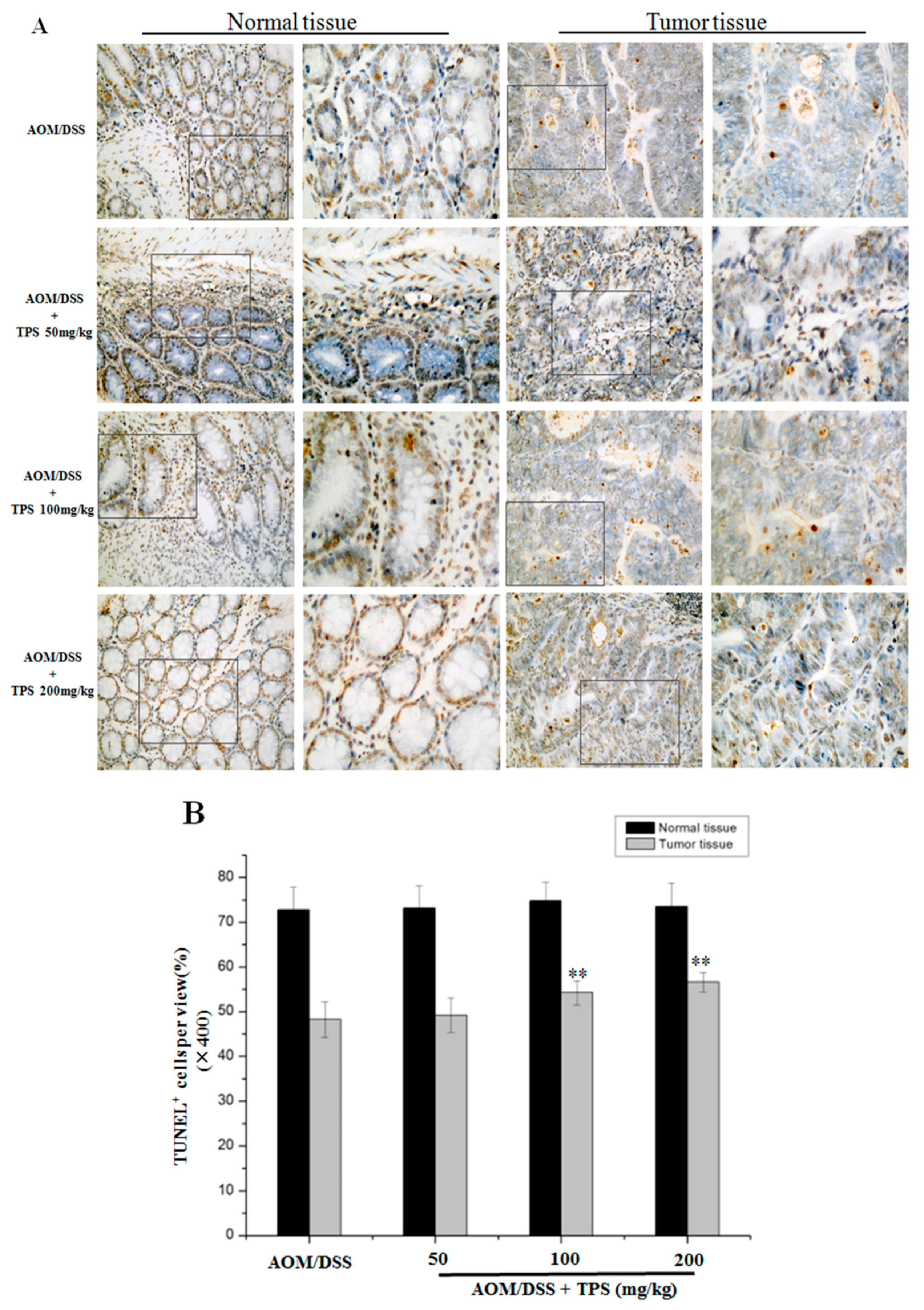
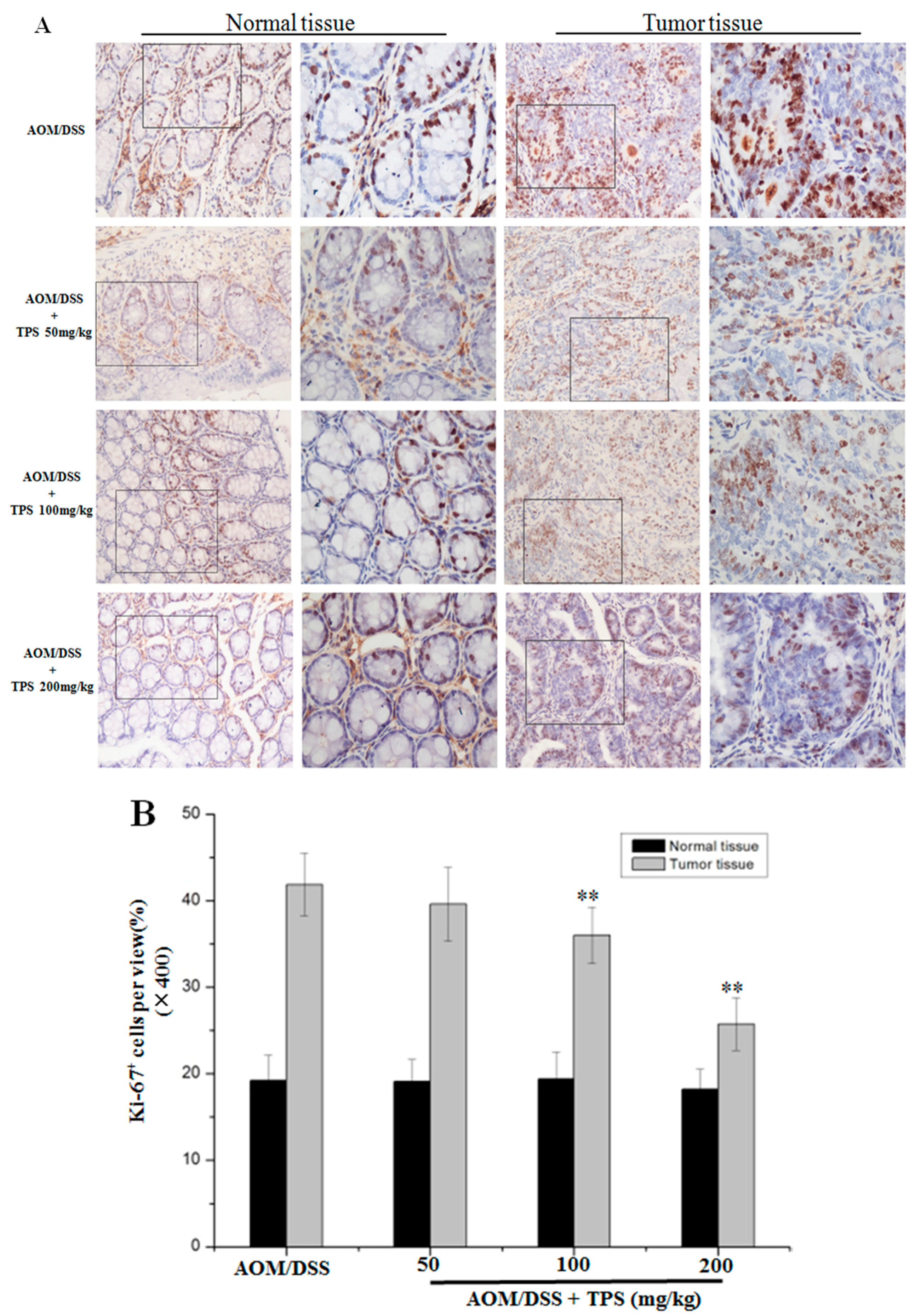
2.2. TPS-Induced Apoptosis and Inhibited Cell Cycle Progression in CAC Mice
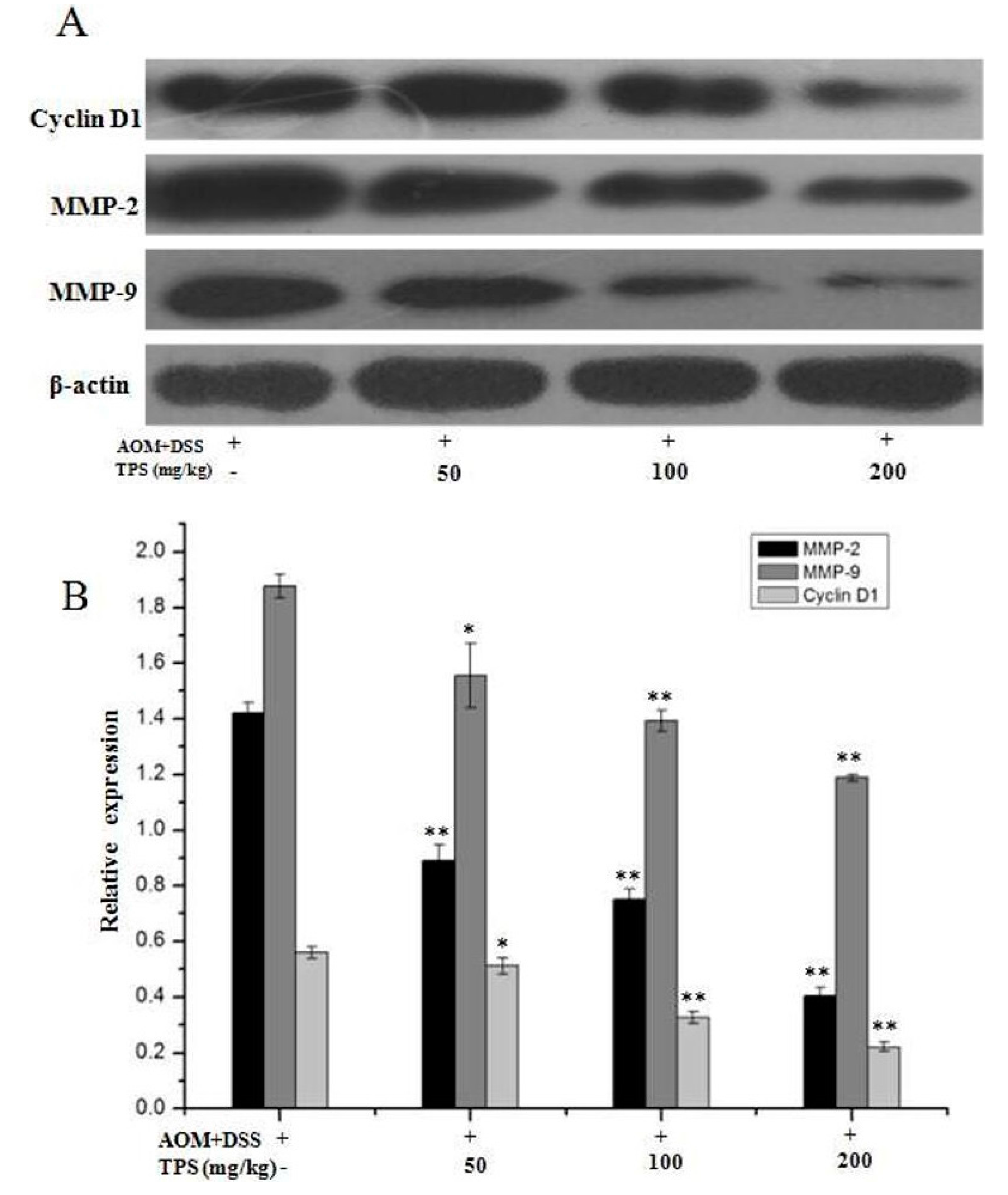
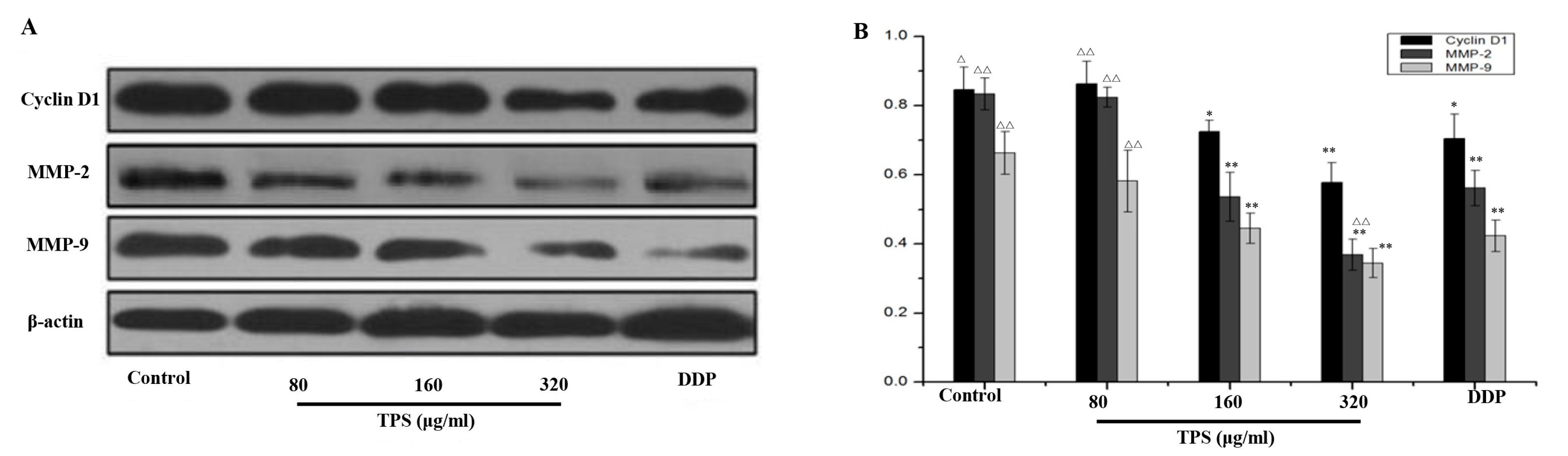
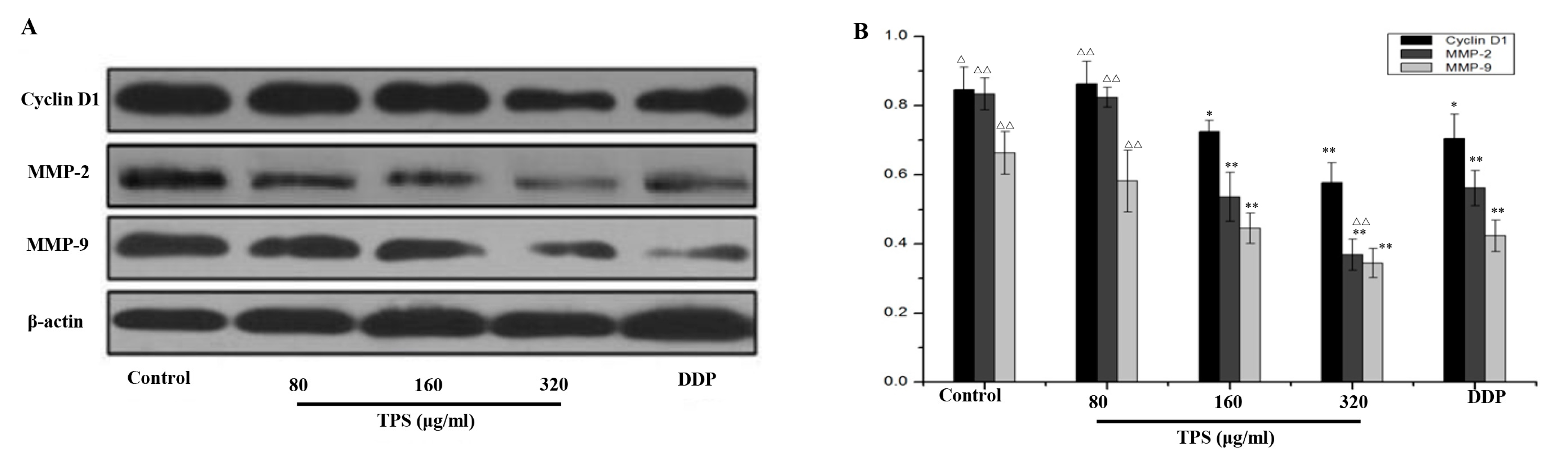
2.3. TPS Inhibited the Expression of Cyclin D1, MMP-2, and MMP-9 in CAC Mice
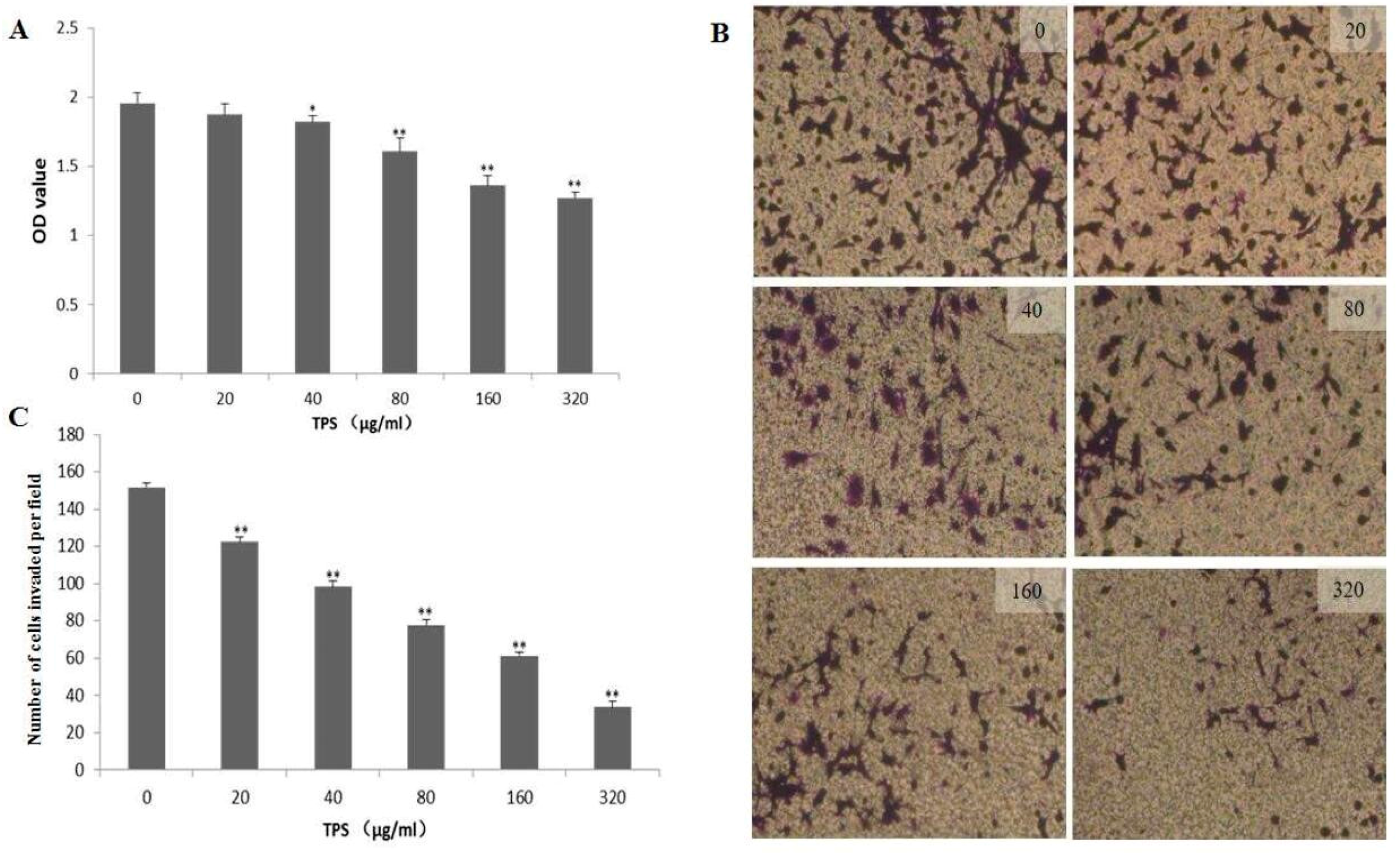
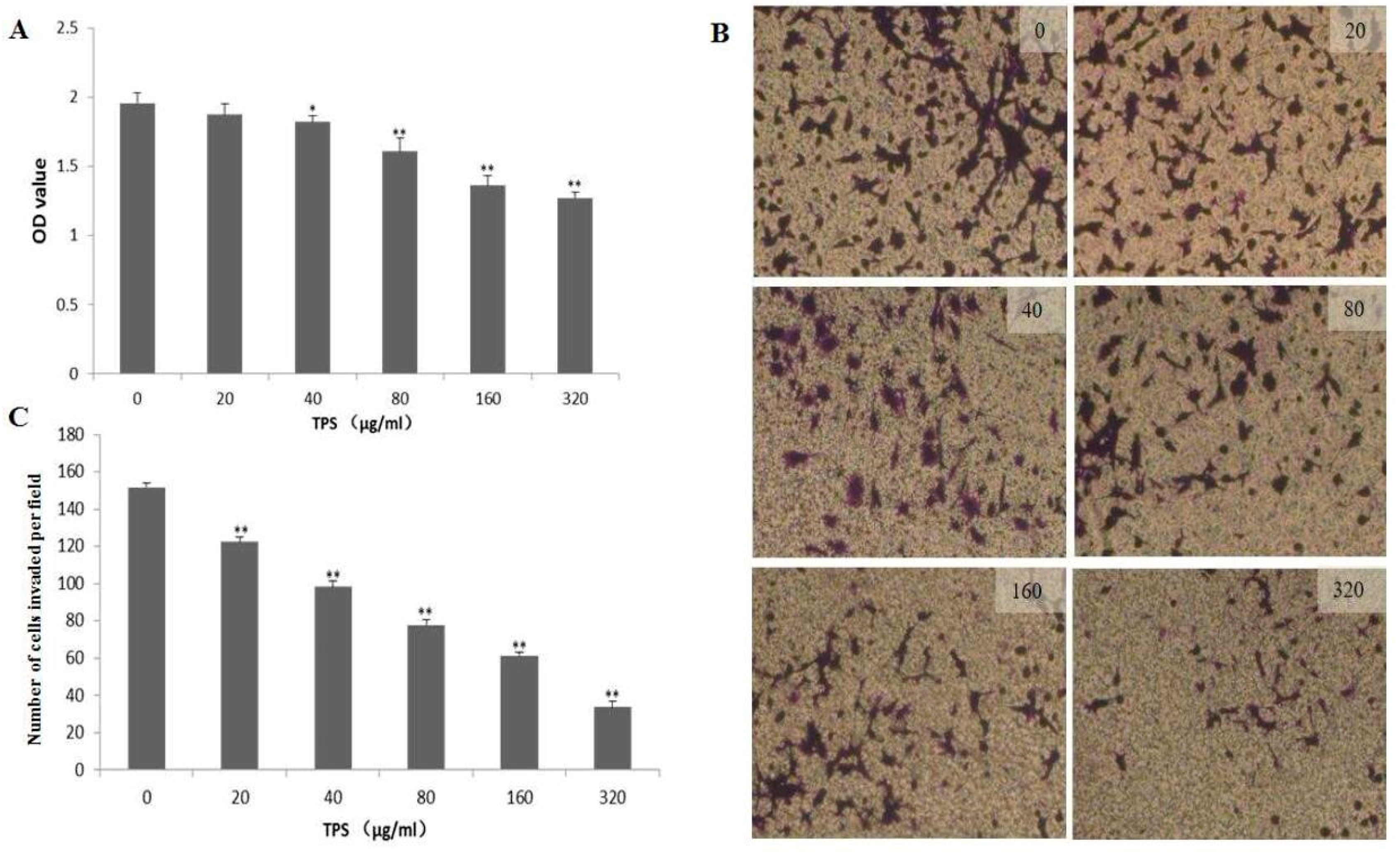
2.4. Effect of TPS on Cell Proliferation and Invasion of CT26 Cells In Vitro
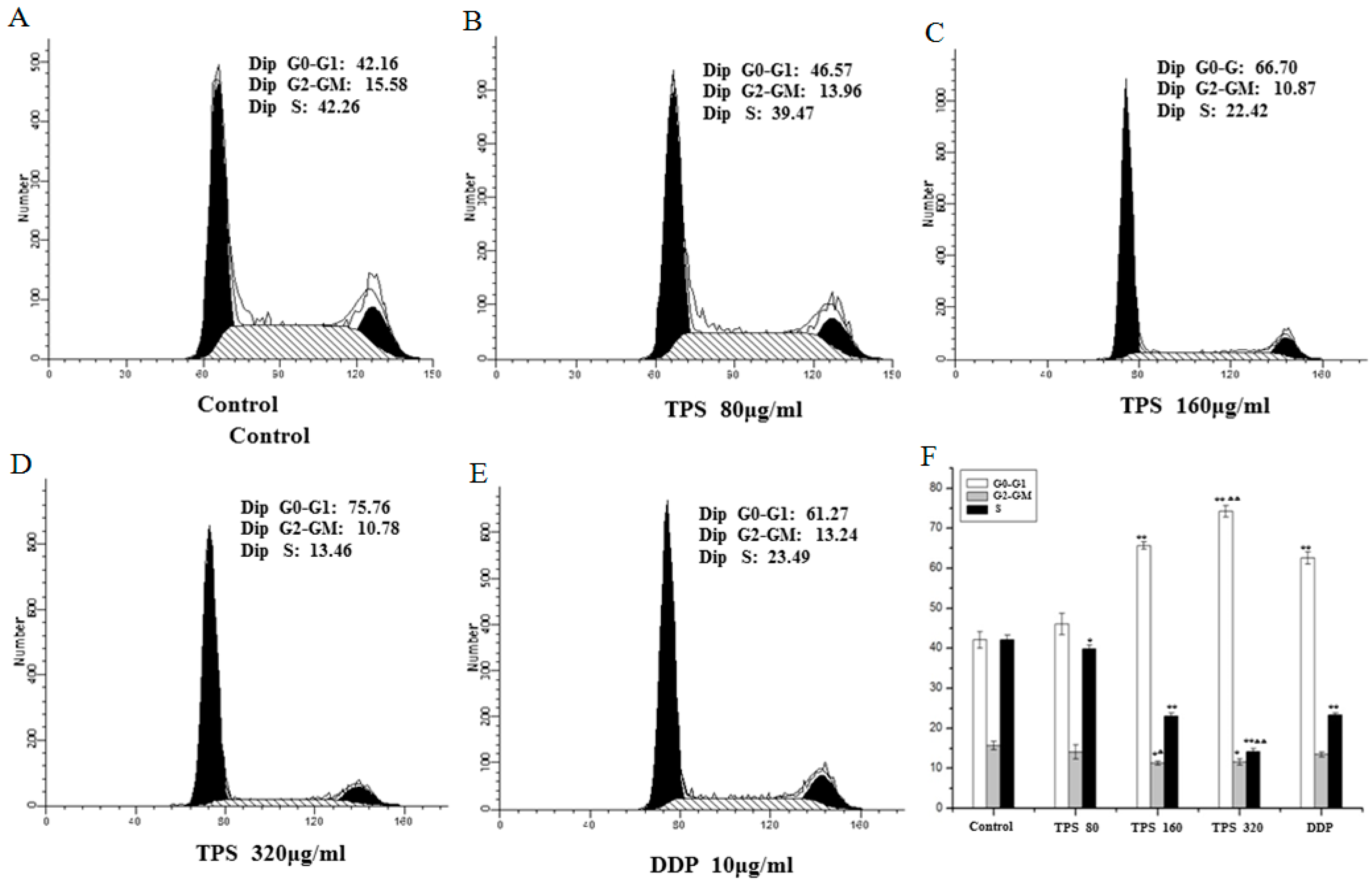
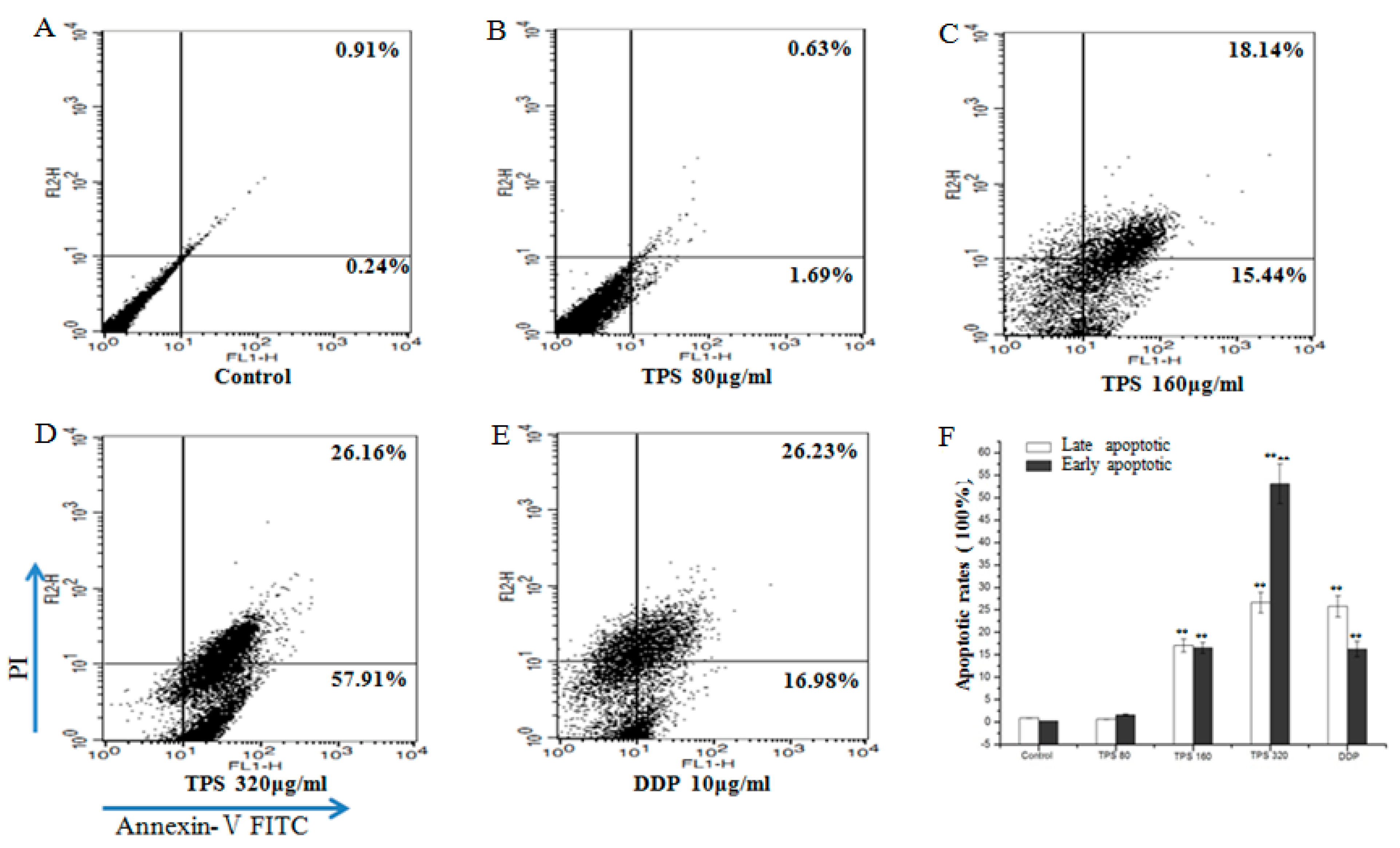
2.5. TPS-Induced Apoptosis and Inhibited Cell Cycle Progression of CT26 Cells In Vitro
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Line and Conditioned Culture
4.3. Cell Viability and Proliferation Assays
4.4. Assessment of Cell Apoptosis by Annexin V-FITC/PI Double-Staining Assay
4.5. Analysis of Cell Cycle
4.6. Cell Matrigel Invasion Assay
4.7. AOM/DSS-Induced Colitis-Associated Colorectal Carcinogenesis and TPS Treatment
4.8. Immunohistochemistry
4.9. Terminal Deoxynucleotide Transferase (TdT) dUTP Nick End Labeling (TUNEL) Assay
4.10. Western Blot Analysis
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, D.; DuBois, R.N. The role of anti-inflammatory drugs in colorectal cancer. Annu. Rev. Med. 2013, 64, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Brückner, M.; Eveslage, M.; Tepasse, P.; Pott, F.; Thoennissen, N.H.; Hengst, K.; Ross, M.; Bettenworth, D. The risk of colorectal cancer in patients with ulcerative colitis. Dig. Dis. Sci. 2015, 60, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Bingham, S.A.; Day, N.E.; Luben, R.; Ferrari, P.; Slimani, N.; Norat, T.; Clavel-Chapelon, F.; Kesse, E. Dietary fibre in food and protection against colorectal cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC): An observational study. Lancet 2003, 361, 1496–1501. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134 (Suppl. 12), 3479–3485. [Google Scholar] [CrossRef]
- Li, W.; Nie, S.; Xie, M.; Chen, Y.; Li, C.; Zhang, H. A major green tea component, (-)-epigallocatechin-3-gallate, ameliorates doxorubicin-mediated cardiotoxicity in cardiomyocytes of neonatal rats. J. Agric. Food Chem. 2010, 58, 8977–8982. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, M.; Wu, T.; Dai, S.; Xu, J.; Zhou, Z. The anti-obesity effect of green tea polysaccharides, polyphenols and caffeine in rats fed with a high-fat diet. Food Funct. 2015, 6, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhao, Y.; Sun, Y.; Yang, X. Protective effects of Ziyang tea polysaccharides on CCl4-induced oxidative liver damage in mice. Food Chem. 2014, 143, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, B.; Gu, Y. Pharmacological evaluation of tea polysaccharides with antioxidant activity in gastric cancer mice. Carbohydr. Polym. 2012, 90, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Liu, X.; Shen, M.Y.; Nie, S.P.; Zhang, H.; Li, C.; Gong, D.M.; Xie, M.Y. Purification, physicochemical characterisation and anticancer activity of a polysaccharide from Cyclocarya paliurus leaves. Food Chem. 2013, 136, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Hwang, D.; Suh, H.J.; Kim, T.Y.; Yu, K.W.; Shin, K.S. Antitumor and antimetastatic activities of rhamnogalacturonan-II-type polysaccharide isolated from mature leaves of green tea via activation of macrophages and natural killer cells. Int. J. Biol. Macromol. 2017, 99, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Dongfeng, W.; Dongfeng, W.; Chenghong, W.; Chenghong, W.; Jun, L.; Guiwen, Z. Components and activity of polysaccharides from coarse tea. J. Agric. Food Chem. 2001, 49, 507–510. [Google Scholar] [CrossRef]
- Nie, S.P.; Xie, M.Y. A review on the isolation and structure of tea polysaccharides and their bioactivities. Food Hydrocoll. 2011, 25, 144–149. [Google Scholar] [CrossRef]
- Fan, D.; He, T.; Wang, Y.; Kong, G.; Jiang, T.; Zhou, D. Production, preliminary characterization and antitumor activity (SKOV-3 cell lines) in vitro of glycans from green tea. Carbohydr. Polym. 2011, 86, 1651–1656. [Google Scholar] [CrossRef]
- Wei, X.; Mao, F.; Cai, X.; Wang, Y. Composition and bioactivity of polysaccharides from tea seeds obtained by water extraction. Int. J. Biol. Macromol. 2011, 49, 587–590. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Shi, X.; Zhao, Y.; Tian, L.; Wang, D.; Yang, X. Inhibitory Effects and Molecular Mechanisms of Selenium-Containing Tea Polysaccharides on Human Breast Cancer MCF-7 Cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar] [CrossRef] [PubMed]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhao, Y.; Sun, Y.; Yang, S.; Yang, X. Characterisation of polysaccharides from green tea of Huangshan Maofeng with antioxidant and hepatoprotective effects. Food Chem. 2013, 141, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Cao, H. Polysaccharides from Chinese tea: Recent advance on bioactivity and function. Int. J. Biol. Macromol. 2013, 62, 76–79. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Gonçalves, G.; de Sá-Nakanishi, A.B.; Wendt, M.M.N.; Comar, J.F.; Amado, C.A.B.; Bracht, A.; Peralta, R.M. Green tea extract improves the oxidative state of the liver and brain in rats with adjuvant-induced arthritis. Food. Funct. 2015, 6, 2701–2711. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, X.; Lu, G.; Picinich, S.C. Cancer prevention by tea: Animal studies, molecular mechanisms and human relevance. Nat. Rev. Cancer 2009, 9, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ding, C.; Yuan, S.; Zhang, Z.; Du, L.; Yuan, M. Extraction, purification and characterization of polysaccharides from Hawk tea. Carbohydr. Polym. 2014, 99, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Jiang, H. A review on the structure-function relationship aspect of polysaccharides from tea materials. Crit. Rev. Food Sci. Nutr. 2015, 55, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Jin, M.L.; Morris, G.A.; Zha, X.Q.; Chen, H.Q.; Yi, Y.; Li, J.E.; Wang, Z.J.; Gao, J.; Nie, S.P.; et al. Advances on bioactive polysaccharides from medicinal plants. Crit. Rev. Food Sci. Nutr. 2016, 56, S60–S84. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Tang, W.; Jin, M.L.; Li, J.E.; Xie, M.Y. Recent advances in bioactive polysaccharides from Lycium barbarum L.; Zizyphus jujuba Mill, Plantago spp., and Morus spp.: Structures and functionalities. Food Hydrocoll. 2016, 60, 148–160. [Google Scholar] [CrossRef]
- Yin, X.; Ying, J.; Li, L.; Zhang, H.; Wang, H. A meta-analysis of lentinan injection combined with chemotherapy in the treatment of nonsmall cell lung cancer. Indian J. Cancer 2015, 52, e29–e31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Nie, S.; Huang, D.; Huang, J.; Feng, Y.; Xie, M. A polysaccharide from Ganoderma atrum inhibits tumor growth by induction of apoptosis and activation of immune response in CT26-bearing mice. J. Agric. Food Chem. 2014, 62, 9296–9304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Nie, S.; Huang, D.; Feng, Y.; Xie, M. A novel polysaccharide from Ganoderma atrum exerts antitumor activity by activating mitochondria-mediated apoptotic pathway and boosting the immune system. J. Agric. Food Chem. 2014, 62, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Neufert, C.; Becker, C.; Neurath, M.F. An inducible mouse model of colon carcinogenesis for the analysis of sporadic and inflammation-driven tumor progression. Nat. Protoc. 2007, 2, 1998–2004. [Google Scholar] [CrossRef] [PubMed]
- Paschos, K.A.; Majeed, A.W.; Bird, N.C. Natural history of hepatic metastases from colorectal cancer—Pathobiological pathways with clinical significance. World. J. Gastroenterol. 2014, 20, 3719–3737. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.T.; Li, H.Y.; Chang, N.S.; Lin, C.H.; Chen, Y.C.; Lu, P.J. WWOX suppresses prostate cancer cell progression through cyclin D1-mediated cell cycle arrest in the G1 phase. Cell Cycle 2015, 14, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhang, L.; Lv, W.; Dong, C.; Wang, Y.; Zhang, J. Overexpression of cyclin D1 in meningioma is associated with malignancy grade and causes abnormalities in apoptosis, invasion and cell cycle progression. Med. Oncol. 2015, 32, 439. [Google Scholar] [CrossRef] [PubMed]
- Simile, M.M.; De Miglio, M.R.; Muroni, M.R.; Frau, M.; Asara, G.; Serra, S.; Mutoni, M.D.; Seddaiu, M.A.; Daino, L.; Feo, F.; et al. Down-regulation of c-myc and Cyclin D1 genes by antisense oligodeoxy nucleotides inhibits the expression of E2F1 and in vitro growth of HepG2 and Morris 5123 liver cancer cells. Carcinogenesis 2004, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Yang, C.J.; Lin, C.Y.; Lee, Y.S.; Wu, J.M. Control of stability of cyclin D1 by quinone reductase 2 in CWR22Rv1 prostate cancer cells. Carcinogenesis 2012, 33, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Peng, X.; Zhou, C.; Zhong, Y.; Chen, X.; Qiu, Y.; Ji, M.; Gong, M.; Kong, D. Stellettin B Induces G1 Arrest, Apoptosis and Autophagy in Human Non-small Cell Lung Cancer A549 Cells via Blocking PI3K/Akt/mTOR Pathway. Sci. Rep. 2016, 6, 27071. [Google Scholar] [CrossRef] [PubMed]
- Tong, K.L.; Chan, K.L.; AbuBakar, S.; Low, B.S.; Ma, H.Q.; Wong, P.F. The In Vitro and In Vivo Anti-Cancer Activities of a Standardized Quassinoids Composition from Eurycoma longifolia on LNCaP Human Prostate Cancer Cells. PLoS ONE 2015, 10, 121752. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Iizuka, S.; Yoshida, M.; Tsunematsu, T.; Kondo, T.; Subarnbhesaj, A.; Deraz, E.M.; Siriwardena, S.B.; Tahara, H.; Ishimaru, N.; et al. Matrix Metalloproteinase-13 (MMP-13) Directly and Indirectly Promotes Tumor Angiogenesis. J. Biol. Chem. 2012, 287, 38716–38728. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, M.; Nasser, M.W.; Ravi, J.; Wani, N.A.; Ahirwar, D.K.; Zhao, H.; Oghumu, S.; Satoskar, A.R.; Shilo, K.; Carson, W.E.; et al. Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: Novel anti-tumor mechanisms of Cannabidiol in breast cancer. Mol. Oncol. 2015, 9, 906–919. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Nie, S.P.; Wang, J.Q.; Huang, D.F.; Li, W.J.; Xie, M.Y. Signaling Pathway Involved in the Immunomodulatory Effect of Ganoderma atrum Polysaccharide in Spleen Lymphocytes. J. Agric. Food Chem. 2015, 63, 2734–2740. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, H.; Wang, Y.; Nie, S.; Li, C.; Xie, M. Acetylation and carboxymethylation of the polysaccharide from Ganoderma atrum and their antioxidant and immunomodulating activities. Food Chem. 2014, 156, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, W.D.; Li, Y. Antitumor and immunomodulatory activity of polysaccharide isolated from Trametes orientalis. Carbohydr. Polym. 2015, 131, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zou, G.; Li, C. Antitumor and immunomodulatory activities of a water-soluble polysaccharide from Chaenomeles speciosa. Carbohydr. Polym. 2015, 132, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Nie, S.; Xie, M. Tumor microenvironment as a new target for tumor immunotherapy of polysaccharides. Crit. Rev. Food Sci. Nutr. 2016, 56, S85–S94. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.S.; Nie, S.P.; Feng, Y.L.; Wang, J.Q.; Que, Z.Q. Isolation, purification and physico-chemical properties of acidic polysaccharides from green tea. Food Sci. 2014, 35, 57–60. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mg/kg) | Colon Length (cm) | Tumor Number |
|---|---|---|---|
| AOM/DSS | 0 | 6.582 ± 0.250 | 16.667 ± 2.582 |
| 50 | 7.117 ± 0.380 | 11.667 ± 2.160 Δ | |
| AOM/DSS + TPS | 100 | 7.747 ± 0.318 ΔΔ | 8.833 ± 1.472 ΔΔ |
| 200 | 8.272 ± 0.450 ΔΔ | 4.833 ± 1.722 ΔΔ |
| Group | Dose (mg/kg) | Spleen Index (mg/g) | Thymus Index (mg/g) |
|---|---|---|---|
| Normal | 200 | 5.732 ± 0.359 ΔΔ | 1.773 ± 0.051 ΔΔ |
| Normal + TPS | 0 | 6.455 ± 0.341 **Δ | 2.232 ± 0.090 **ΔΔ |
| AOM/DSS | 0 | 7.035 ± 0.206 ** | 1.957 ± 0.181 |
| 50 | 6.843 ± 0.262 ** | 2.402 ± 0.250 **ΔΔ | |
| AOM/DSS + TPS | 100 | 6.672 ± 0.229 ** | 2.408 ± 0.116 **ΔΔ |
| 200 | 6.510 ± 0.256 *Δ | 2.672 ± 0.093 **ΔΔ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.-Q.; Li, H.-S.; Nie, S.-P.; Shen, M.-Y.; Hu, J.-L.; Xie, M.-Y. Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells. Int. J. Mol. Sci. 2018, 19, 506. https://doi.org/10.3390/ijms19020506
Liu L-Q, Li H-S, Nie S-P, Shen M-Y, Hu J-L, Xie M-Y. Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells. International Journal of Molecular Sciences. 2018; 19(2):506. https://doi.org/10.3390/ijms19020506
Chicago/Turabian StyleLiu, Li-Qiao, Hai-Shan Li, Shao-Ping Nie, Ming-Yue Shen, Jie-Lun Hu, and Ming-Yong Xie. 2018. "Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells" International Journal of Molecular Sciences 19, no. 2: 506. https://doi.org/10.3390/ijms19020506
APA StyleLiu, L.-Q., Li, H.-S., Nie, S.-P., Shen, M.-Y., Hu, J.-L., & Xie, M.-Y. (2018). Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells. International Journal of Molecular Sciences, 19(2), 506. https://doi.org/10.3390/ijms19020506