Different Achilles Tendon Pathologies Show Distinct Histological and Molecular Characteristics

 ,
,
Abstract

1. Introduction
2. Results
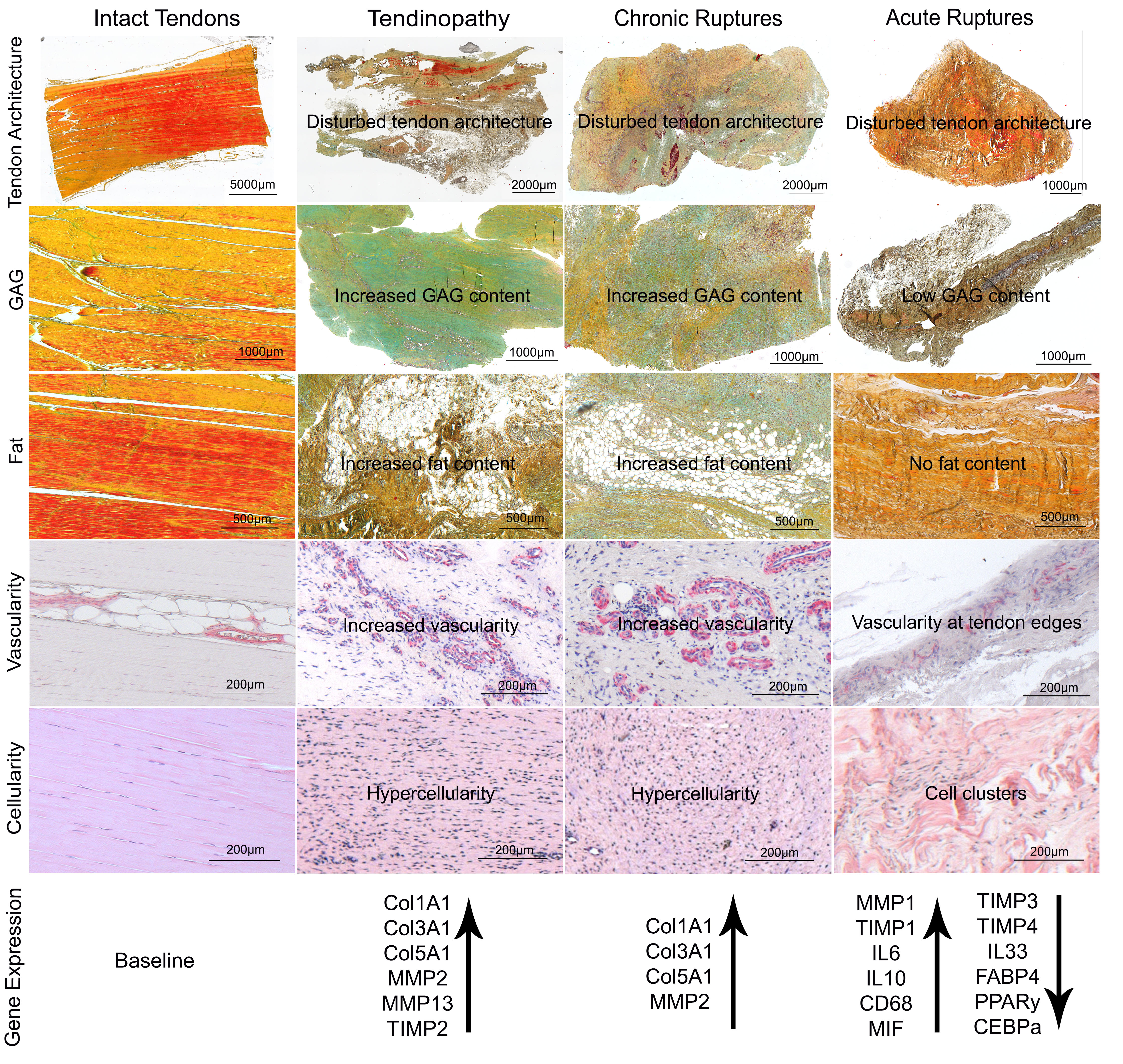
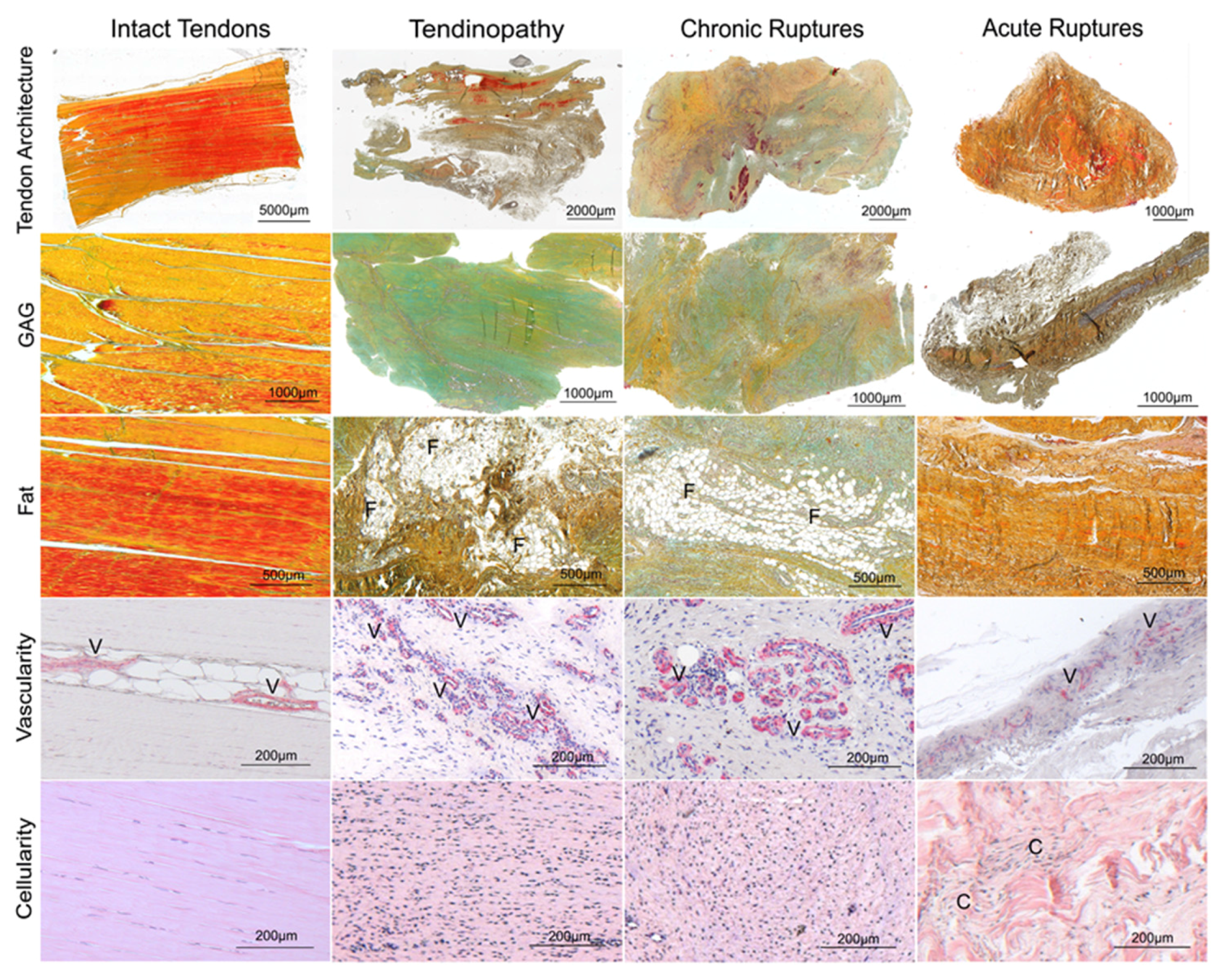
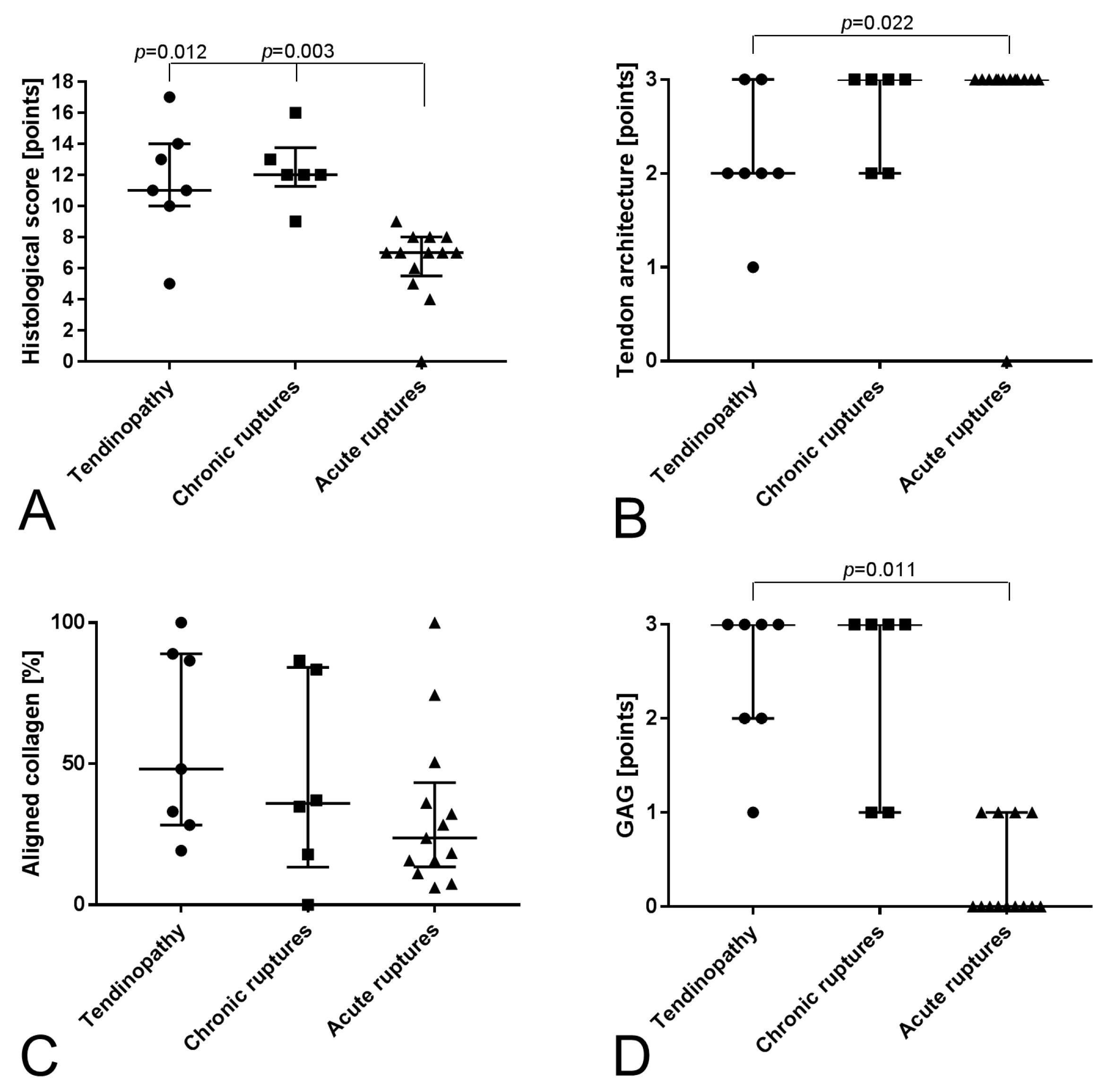
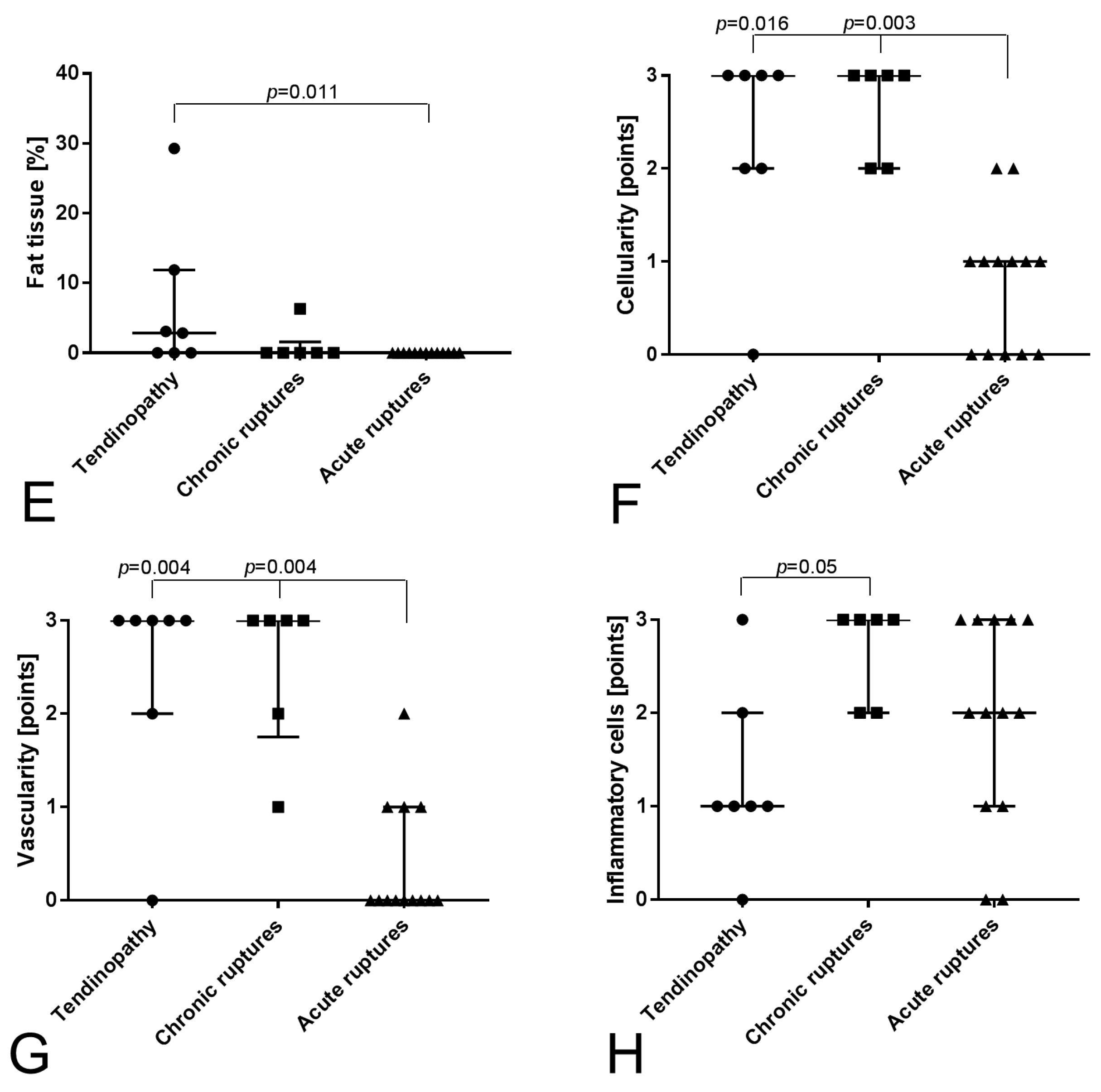
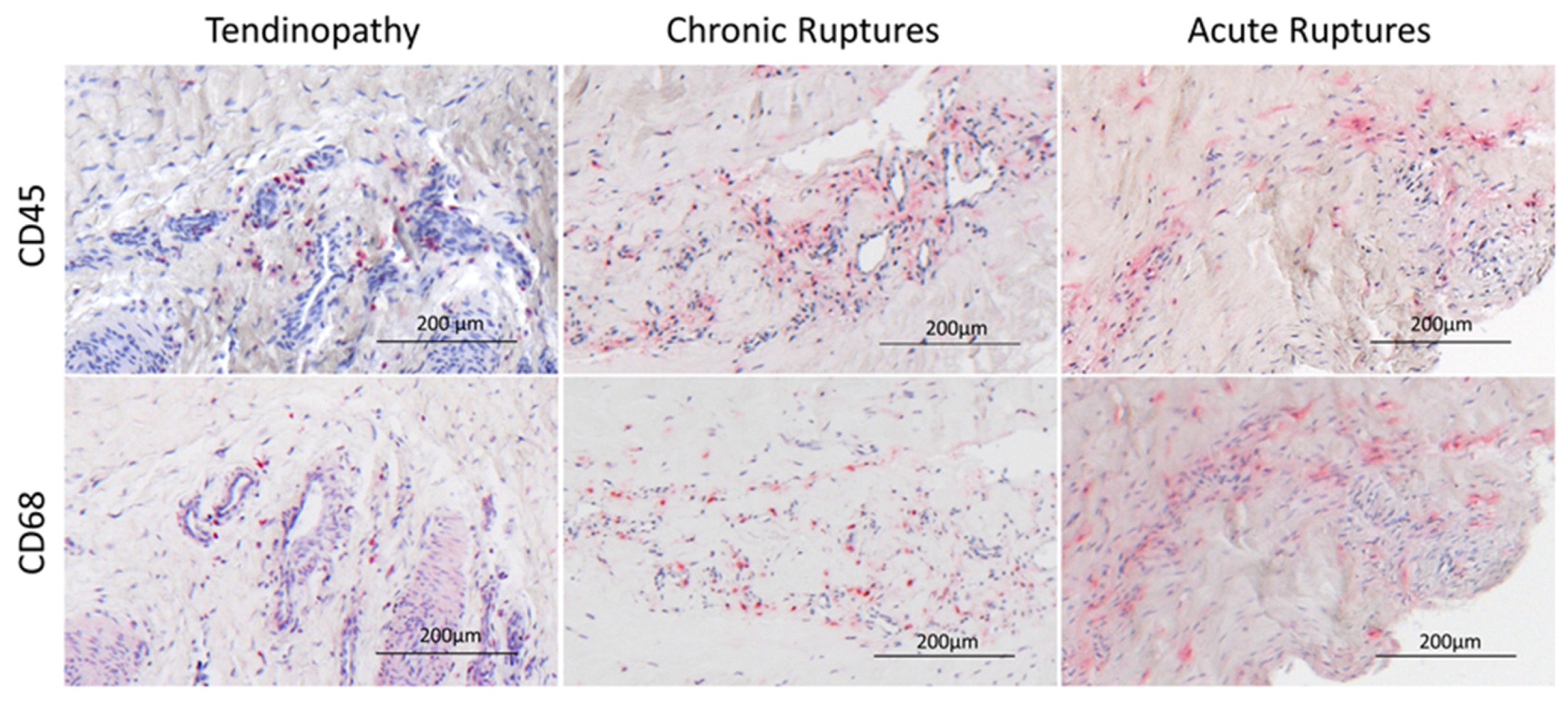
2.1. Histological Analysis
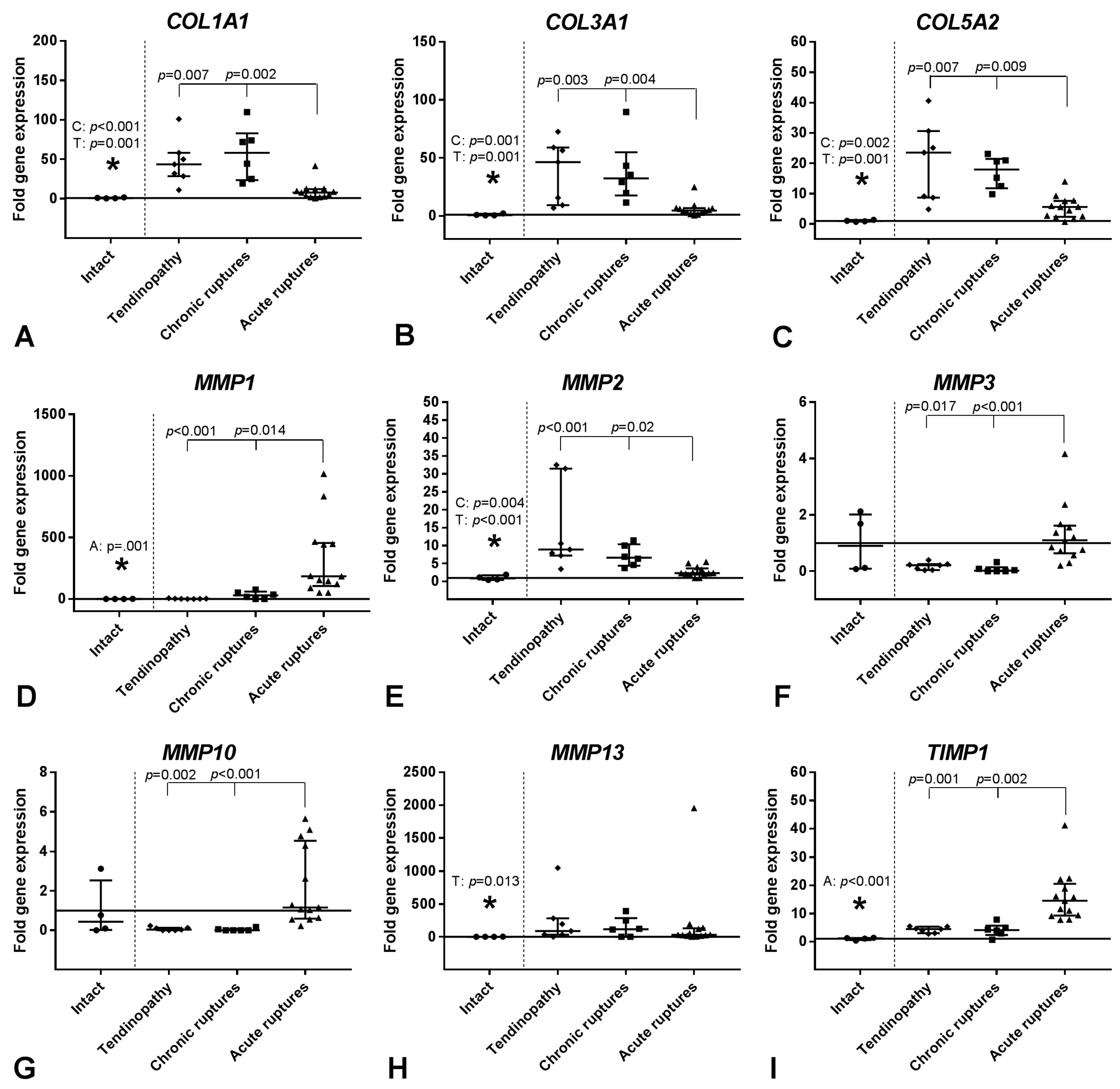
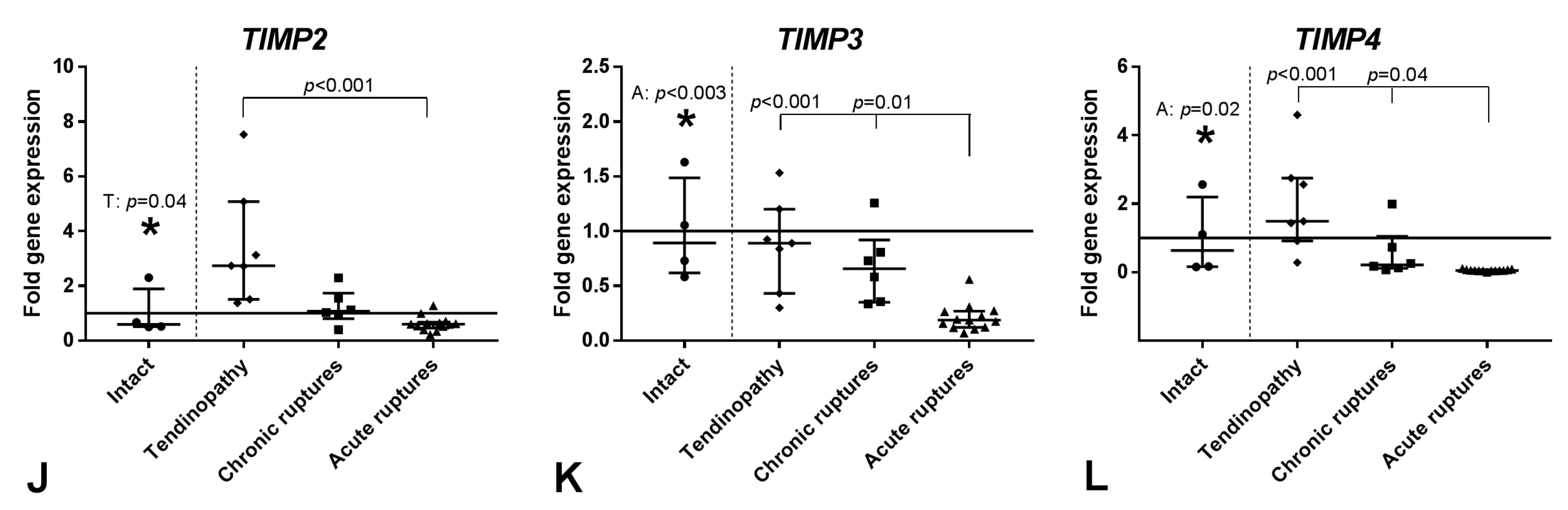
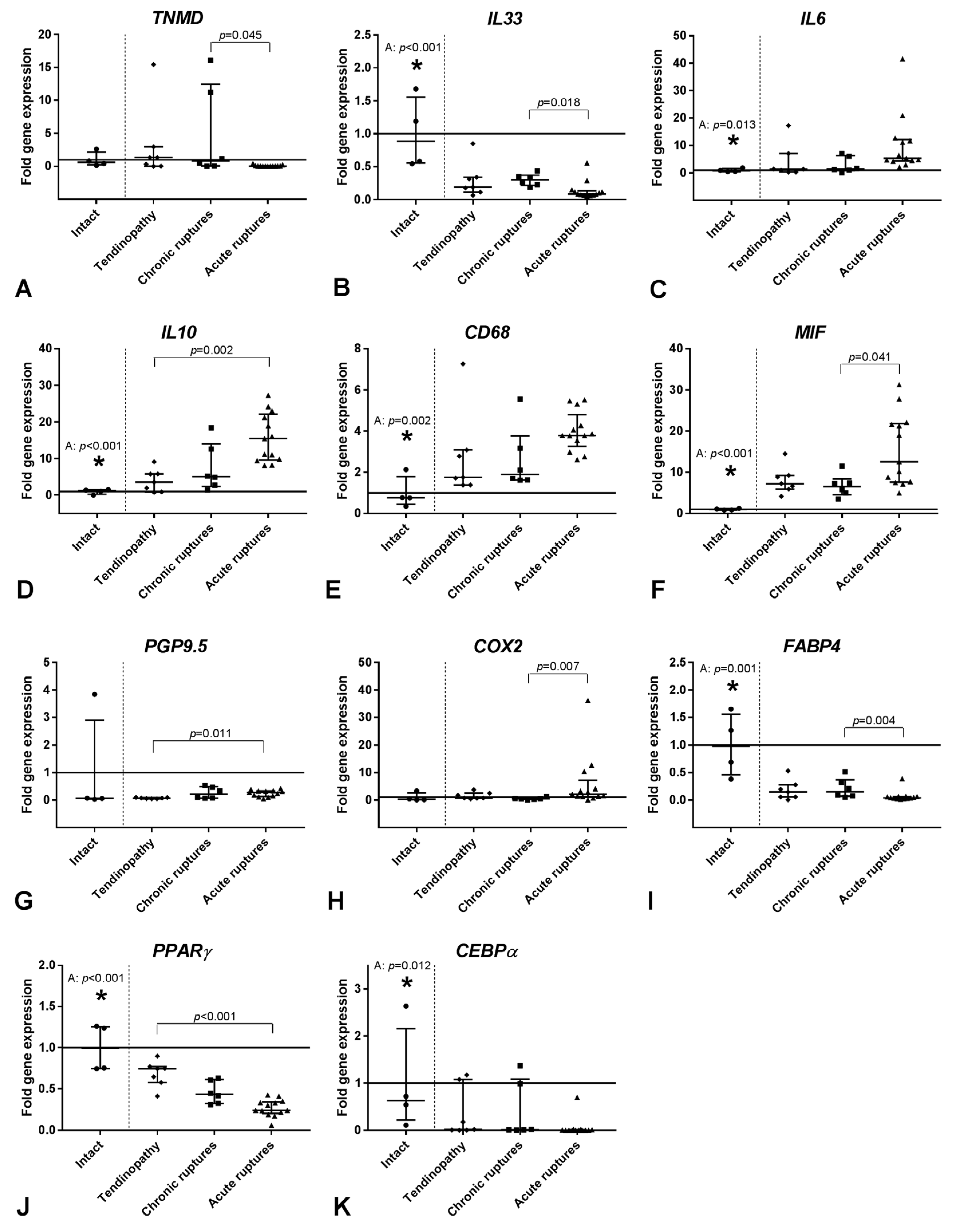
2.2. Gene Expression
3. Discussion
Limitations
4. Materials and Methods
4.1. Tendon Sampling
4.2. Histological Evaluation
4.3. Immunohistochemistry
4.4. qRT-PCR
4.5. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sobhani, S.; Dekker, R.; Postema, K.; Dijkstra, P.U. Epidemiology of ankle and foot overuse injuries in sports: A systematic review. Scand. J. Med. Sci. Sports 2013, 23, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.M.; Cook, J.L.; Bonar, F.; Harcourt, P.; Astrom, M. Histopathology of common tendinopathies. Update and implications for clinical management. Sports Med. 1999, 27, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Maffulli, N.; Longo, U.G.; Denaro, V. Novel approaches for the management of tendinopathy. J. Bone Jt. Surg. Am. 2010, 92, 2604–2613. [Google Scholar] [CrossRef]
- Leung, J.L.; Griffith, J.F. Sonography of chronic Achilles tendinopathy: A case-control study. J. Clin. Ultrasound 2008, 36, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Longo, U.G.; Ronga, M.; Maffulli, N. Achilles tendinopathy. Sports Med. Arthrosc. 2009, 17, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Maffulli, N.; Longo, U.G.; Franceschi, F.; Rabitti, C.; Denaro, V. Movin and Bonar scores assess the same characteristics of tendon histology. Clin. Orthop. Relat. Res. 2008, 466, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Maffulli, N.; Longo, U.G.; Maffulli, G.D.; Rabitti, C.; Khanna, A.; Denaro, V. Marked pathological changes proximal and distal to the site of rupture in acute Achilles tendon ruptures. Knee Surg. Sports Traumatol. Arthrosc. 2011, 19, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Pingel, J.; Lu, Y.; Starborg, T.; Fredberg, U.; Langberg, H.; Nedergaard, A.; Weis, M.; Eyre, D.; Kjaer, M.; Kadler, K.E. 3-D ultrastructure and collagen composition of healthy and overloaded human tendon: Evidence of tenocyte and matrix buckling. J. Anat. 2014, 224, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Movin, T.; Gad, A.; Reinholt, F.P.; Rolf, C. Tendon pathology in long-standing achillodynia. Biopsy findings in 40 patients. Acta Orthop. Scand. 1997, 68, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.C.; Chan, B.P.; Wang, W.; Pau, H.M.; Chan, K.M.; Rolf, C.G. Increased expression of matrix metalloproteinase 1 (MMP1) in 11 patients with patellar tendinosis. Acta Orthop. Scand. 2002, 73, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, S.M.; van der Merwe, L.; Ribbans, W.J.; Smith, R.K.; Schwellnus, M.P.; Collins, M. Variants within the MMP3 gene are associated with Achilles tendinopathy: Possible interaction with the COL5A1 gene. Br. J. Sports Med. 2009, 43, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Ireland, D.; Harrall, R.; Curry, V.; Holloway, G.; Hackney, R.; Hazleman, B.; Riley, G. Multiple changes in gene expression in chronic human Achilles tendinopathy. Matrix Biol. 2001, 20, 159–169. [Google Scholar] [CrossRef]
- Alfredson, H.; Lorentzon, M.; Backman, S.; Backman, A.; Lerner, U.H. cDNA-arrays and real-time quantitative PCR techniques in the investigation of chronic Achilles tendinosis. J. Orthop. Res. 2003, 21, 970–975. [Google Scholar] [CrossRef]
- Battery, L.; Maffulli, N. Inflammation in overuse tendon injuries. Sports Med. Arthrosc. 2011, 19, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Silbernagel, K.G.; Siljeholm, C.; di Iorio, A.; de Amicis, D.; Salini, V.; Werner, S.; Paganelli, R. Pathogenesis of tendinopathies: Inflammation or degeneration? Arthritis Res. Ther. 2009, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Del Buono, A.; Battery, L.; Denaro, V.; Maccauro, G.; Maffulli, N. Tendinopathy and inflammation: Some truths. Int. J. Immunopathol. Pharmacol. 2011, 24, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Fredberg, U.; Stengaard-Pedersen, K. Chronic tendinopathy tissue pathology, pain mechanisms, and etiology with a special focus on inflammation. Scand. J. Med. Sci. Sports 2008, 18, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Kannus, P.; Jozsa, L. Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients. J. Bone Jt. Surg. Am. 1991, 73, 1507–1525. [Google Scholar] [CrossRef]
- Schubert, T.E.; Weidler, C.; Lerch, K.; Hofstadter, F.; Straub, R.H. Achilles tendinosis is associated with sprouting of substance P positive nerve fibres. Ann. Rheum. Dis. 2005, 64, 1083–1086. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Lian, O.; Bahr, R.; Hart, D.A.; Duronio, V.; Khan, K.M. Increased mast cell numbers in human patellar tendinosis: Correlation with symptom duration and vascular hyperplasia. Br. J. Sports Med. 2008, 42, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Behzad, H.; Sharma, A.; Mousavizadeh, R.; Lu, A.; Scott, A. Mast cells exert pro-inflammatory effects of relevance to the pathophyisology of tendinopathy. Arthritis Res. Ther. 2013, 15, R184. [Google Scholar] [CrossRef] [PubMed]
- Kragsnaes, M.S.; Fredberg, U.; Stribolt, K.; Kjaer, S.G.; Bendix, K.; Ellingsen, T. Stereological quantification of immune-competent cells in baseline biopsy specimens from achilles tendons: Results from patients with chronic tendinopathy followed for more than 4 years. Am. J. Sports Med. 2014, 42, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Hueber, A.J.; Reilly, J.H.; Xu, Y.; Fazzi, U.G.; Murrell, G.A.; McInnes, I.B. Inflammation is present in early human tendinopathy. Am. J. Sports Med. 2010, 38, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Dakin, S.G.; Newton, J.; Martinez, F.O.; Hedley, R.; Gwilym, S.; Jones, N.; Reid, H.A.B.; Wood, S.; Wells, G.; Appleton, L.; et al. Chronic inflammation is a feature of Achilles tendinopathy and rupture. Br. J. Sports Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rees, J.D.; Maffulli, N.; Cook, J. Management of tendinopathy. Am. J. Sports Med. 2009, 37, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Alfredson, H.; Lorentzon, R. Chronic tendon pain: No signs of chemical inflammation but high concentrations of the neurotransmitter glutamate. Implications for treatment? Curr. Drug Targets 2002, 3, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Dean, B.J.; Snelling, S.J.; Dakin, S.G.; Murphy, R.J.; Javaid, M.K.; Carr, A.J. Differences in glutamate receptors and inflammatory cell numbers are associated with the resolution of pain in human rotator cuff tendinopathy. Arthritis Res. Ther. 2015, 17, 176. [Google Scholar] [CrossRef] [PubMed]
- Spang, C.; Harandi, V.M.; Alfredson, H.; Forsgren, S. Marked innervation but also signs of nerve degeneration in between the Achilles and plantaris tendons and presence of innervation within the plantaris tendon in midportion Achilles tendinopathy. J. Musculoskelet. Neuronal Interact. 2015, 15, 197–206. [Google Scholar] [PubMed]
- Khan, K.M.; Cook, J.L.; Maffulli, N.; Kannus, P. Where is the pain coming from in tendinopathy? It may be biochemical, not only structural, in origin. Br. J. Sports Med. 2000, 34, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Legerlotz, K.; Jones, E.R.; Screen, H.R.; Riley, G.P. Increased expression of IL-6 family members in tendon pathology. Rheumatology 2012, 51, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Burssens, P.; Forsyth, R.; Steyaert, A.; Van Ovost, E.; Praet, M.; Verdonk, R. Influence of burst TENS stimulation on collagen formation after Achilles tendon suture in man. A histological evaluation with Movat’s pentachrome stain. Acta Orthop. Belg. 2005, 71, 342–346. [Google Scholar] [PubMed]
- Pingel, J.; Fredberg, U.; Qvortrup, K.; Larsen, J.O.; Schjerling, P.; Heinemeier, K.; Kjaer, M.; Langberg, H. Local biochemical and morphological differences in human Achilles tendinopathy: A case control study. BMC Musculoskelet. Disord. 2012, 13, 53. [Google Scholar] [CrossRef]
- Goncalves-Neto, J.; Witzel, S.S.; Teodoro, W.R.; Carvalho-Junior, A.E.; Fernandes, T.D.; Yoshinari, H.H. Changes in collagen matrix composition in human posterior tibial tendon dysfunction. Jt. Bone Spine 2002, 69, 189–194. [Google Scholar] [CrossRef]
- September, A.V.; Schwellnus, M.P.; Collins, M. Tendon and ligament injuries: The genetic component. Br. J. Sports Med. 2007, 41, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Magra, M.; Maffulli, N. Genetic aspects of tendinopathy. J. Sci. Med. Sport 2008, 11, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Mokone, G.G.; Schwellnus, M.P.; Noakes, T.D.; Collins, M. The COL5A1 gene and Achilles tendon pathology. Scand. J. Med. Sci. Sports 2006, 16, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Hunziker, E.B.; Fassler, R.; Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Gilchrist, D.S.; Akbar, M.; Reilly, J.H.; Kerr, S.C.; Campbell, A.L.; Murrell, G.A.; Liew, F.Y.; Kurowska-Stolarska, M.; McInnes, I.B. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nat. Commun. 2015, 6, 6774. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar] [PubMed]
- Del Buono, A.; Oliva, F.; Longo, U.G.; Rodeo, S.A.; Orchard, J.; Denaro, V.; Maffulli, N. Metalloproteases and rotator cuff disease. J. Shoulder Elbow Surg. 2012, 21, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Riley, G.P.; Curry, V.; DeGroot, J.; van El, B.; Verzijl, N.; Hazleman, B.L.; Bank, R.A. Matrix metalloproteinase activities and their relationship with collagen remodelling in tendon pathology. Matrix Biol. 2002, 21, 185–195. [Google Scholar] [CrossRef]
- Klatte-Schulz, F.; Aleyt, T.; Pauly, S.; Geissler, S.; Gerhardt, C.; Scheibel, M.; Wildemann, B. Do Matrix Metalloproteases and Tissue Inhibitors of Metalloproteases in Tenocytes of the Rotator Cuff Differ with Varying Donor Characteristics? Int. J. Mol. Sci. 2015, 16, 13141–13157. [Google Scholar] [CrossRef] [PubMed]
- Minkwitz, S.; Schmock, A.; Kurtoglu, A.; Tsitsilonis, S.; Manegold, S.; Wildemann, B.; Klatte-Schulz, F. Time-Dependent Alterations of MMPs, TIMPs and Tendon Structure in Human Achilles Tendons after Acute Rupture. Int. J. Mol. Sci. 2017, 18, 2199. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.A. Directing transition from innate to acquired immunity: Defining a role for IL-6. J. Immunol. 2005, 175, 3463–3468. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.W.; Cardenas, L.; Glaser, D.L.; Soslowsky, L.J. Tendon healing in interleukin-4 and interleukin-6 knockout mice. J. Biomech. 2006, 39, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Muller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Durgam, S.; Stewart, M. Cellular and Molecular Factors Influencing Tendon Repair. Tissue Eng. Part B Rev. 2017, 23, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Dakin, S.G.; Werling, D.; Hibbert, A.; Abayasekara, D.R.; Young, N.J.; Smith, R.K.; Dudhia, J. Macrophage sub-populations and the lipoxin A4 receptor implicate active inflammation during equine tendon repair. PLoS ONE 2012, 7, e32333. [Google Scholar] [CrossRef] [PubMed]
- Stolk, M.; Klatte-Schulz, F.; Schmock, A.; Minkwitz, S.; Wildemann, B.; Seifert, M. New insights into tenocyte-immune cell interplay in an in vitro model of inflammation. Sci. Rep. 2017, 7, 9801. [Google Scholar] [CrossRef] [PubMed]
- Klatte-Schulz, F.; Gerhardt, C.; Scheibel, M.; Wildemann, B.; Pauly, S. Relationship between muscle fatty infiltration and the biological characteristics and stimulation potential of tenocytes from rotator cuff tears. J. Orthop. Res. 2014, 32, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Klatte-Schulz, F.; Pauly, S.; Scheibel, M.; Greiner, S.; Gerhardt, C.; Hartwig, J.; Schmidmaier, G.; Wildemann, B. Characteristics and stimulation potential with BMP-2 and BMP-7 of tenocyte-like cells isolated from the rotator cuff of female donors. PLoS ONE 2013, 8, e67209. [Google Scholar] [CrossRef]
- Klatte-Schulz, F.; Pauly, S.; Scheibel, M.; Greiner, S.; Gerhardt, C.; Schmidmaier, G.; Wildemann, B. Influence of age on the cell biological characteristics and the stimulation potential of male human tenocyte-like cells. Eur. Cell Mater. 2012, 24, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Pingel, J.; Petersen, M.C.; Fredberg, U.; Kjaer, S.G.; Quistorff, B.; Langberg, H.; Hansen, J.B. Inflammatory and Metabolic Alterations of Kager’s Fat Pad in Chronic Achilles Tendinopathy. PLoS ONE 2015, 10, e0127811. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.N. Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells. Vitam. Horm. 2006, 74, 443–477. [Google Scholar] [CrossRef] [PubMed]
- Gomez, R.; Lago, F.; Gomez-Reino, J.; Dieguez, C.; Gualillo, O. Adipokines in the skeleton: Influence on cartilage function and joint degenerative diseases. J. Mol. Endocrinol. 2009, 43, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nishio, Y.; Nishihira, J.; Ishibashi, T.; Kato, H.; Minami, A. Role of macrophage migration inhibitory factor (MIF) in peripheral nerve regeneration: Anti-MIF antibody induces delay of nerve regeneration and the apoptosis of Schwann cells. Mol. Med. 2002, 8, 509–520. [Google Scholar] [PubMed]
- Kendal, A.; Snelling, S.; Dakin, S.; Stace, E.; Mouthuy, P.A.; Carr, A. Resorbable electrospun polydioxanone fibres modify the behaviour of cells from both healthy and diseased human tendons. Eur. Cell Mater. 2017, 33, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Nissinen, L.; Kahari, V.M. Matrix metalloproteinases in inflammation. Biochim. Biophys. Acta 2014, 1840, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Itaya, T.; Kagami, H.; Okada, K.; Yamawaki, A.; Narita, Y.; Inoue, M.; Sumita, Y.; Ueda, M. Characteristic changes of periodontal ligament-derived cells during passage. J. Periodontal Res. 2009, 44, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Brophy, R.H.; Rai, M.F.; Zhang, Z.; Torgomyan, A.; Sandell, L.J. Molecular analysis of age and sex-related gene expression in meniscal tears with and without a concomitant anterior cruciate ligament tear. J. Bone Jt. Surg. Am. 2012, 94, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Simon, P. Q-Gene: Processing quantitative real-time RT-PCR data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n-Value | Age (Mean ± SD) | BMI (Mean ± SD) | Sex (Female/Male) |
|---|---|---|---|---|
| Tendinopathy | 7 | 45.3 ± 7.9 years | 26.8 ± 5.2 | 3/4 |
| Chronic Rupture | 6 | 52.2 ± 12.6 years * | 28.6 ± 5.2 | 1/5 |
| Acute Rupture | 13 | 32.0 ± 3.1 years * | 23.5 ± 1.9 | 2/11 |
| Intact (patient) | 4 | 37.8 ± 12.8 years | 30.5 ± 3.5 | 2/2 |
| Intact (cadaver) | 5 | 83.4 ± 2.9 years | - | 3/2 |
| Gene | Accession No. | Primer Sequence |
|---|---|---|
| 18S rRNA | NM_022551 | Forward: 5′ CGGAAAATAGCCTTTGCCATC 3′ |
| Reverse: 5′ AGTTCTCCCGCCCTCTTGGT 3′ | ||
| COL1A1 | NM_000088.3 | Forward: 5′ TGACCTCAAGATGTGCCACT 3′ |
| Reverse: 5′ ACCAGACATGCCTCTTGTCC 3′ | ||
| COL3A1 | NM_000090.3 | Forward: 5′ GCTGGCATCAAAGGACATCG 3′ |
| Reverse: 5′ TGTTACCTCGAGGCCCTGGT 3′ | ||
| COL5A2 | NM_000393 | Forward: 5′ AGAAGCCTCCCAGAACATCA 3′ |
| Reverse: 5′ ACAGTCTTGCCCACATTTCC 3′ | ||
| MMP1 | NM_002421.3 | Forward: 5′ CACGCCAGATTTGCCAAGAG 3′ |
| Reverse: 5′ GTCCCGATGATCTCCCCTGA 3′ | ||
| MMP2 | NM_ 004530 | Forward: 5′ TGGATGATGCCTTTGCTCGT 3′ |
| Reverse: 5′ CCAGGAGTCCGTCCTTACCG 3′ | ||
| MMP3 | NM_002422.3 | Forward: 5′ TGGGCCAGGGATTAATGGAG 3′ |
| Reverse: 5′ GGCCAATTTCATGAGCAGCA 3′ | ||
| MMP9 | NM_004994.2 | Forward: 5′ GGGACGCAGACATCGTCATC 3′ |
| Reverse: 5′ GGGACCACAACTCGTCATCG 3′ | ||
| MMP10 | NM_ 002425 | Forward: 5′ CCACCTGGACCTGGGCTTTA 3′ |
| Reverse: 5′ GAACTGGGCGAGCTCTGTGA 3′ | ||
| MMP13 | NM_002427.3 | Forward: 5′ CCTTCCCAGTGGTGGTGATG 3′ |
| Reverse: 5′ CGGAGCCTCTCAGTCATGGA 3′ | ||
| TIMP1 | NM_003254.2 | Forward: 5′ TTGGCTGTGAGGAATGCACA 3′ |
| Reverse: 5′ AAGGTGACGGGACTGGAAGC 3′ | ||
| TIMP2 | NM_003255.4 | Forward: 5′ CCTGAGCACCACCCAGAAGA 3′ |
| Reverse: 5′ TCCATCCAGAGGCACTCGTC 3′ | ||
| TIMP3 | NM_000362.4 | Forward: 5′ CCGAGGCTTCACCAAGATGC 3′ |
| Reverse: 5′ GCCATCATAGACGCGACCTG 3′ | ||
| TIMP4 | NM_003256.3 | Forward: 5′ GAAGCCAACAGCCAGAAGCA 3′ |
| Reverse: 5′ TTCCCTCTGCACCAAGGACA 3′ | ||
| TNMD | NM_022144.2 | Forward: 5′ TTGAAGACCCACGAAGTAGA 3′ |
| Reverse: 5′ ATGACATGGAGCACACTTTC 3′ | ||
| IL33 | NM_033439 | Forward: 5′ CCAACAGAAGGCCAAAGAAG 3′ |
| Reverse: 5′ AAGGCAAAGCACTCCACAGT 3′ | ||
| sST2 | NM_003856 | Forward: 5′ CAACAAGAGGAAGGGCAAAA 3′ |
| Reverse: 5′ CAAATTCAGGGCCAGACAGT 3′ | ||
| IL1β | NM_000576 | Forward: 5′ TCCAGGAGAATGACCTGAGC 3′ |
| Reverse: 5′ GTGATCGTACAGGTGCATCG 3′ | ||
| TNFα | NM_000594 | Forward: 5′ AGCCCATGTTGTAGCAAACC 3′ |
| Reverse: 5′ GAGGTACAGGCCCTCTGATG 3′ | ||
| IL6 | NM_000600 | Forward: 5′ TGAGGAGACTTGCCTGGTGA 3′ |
| Reverse: 5′ TTGGGTCAGGGGTGGTTATT 3′ | ||
| IL10 | NM_000572 | Forward: 5′ TGAGAACAGCTGCACCCACT 3′ |
| Reverse: 5′ GGCAACCCAGGTAACCCTTA 3′ | ||
| TGFβ1 | NM_000660.4 | Forward: 5′ AAGGACCTCGGCTGGAAGTG 3′ |
| Reverse: 5′ AGGGCCAGGACCTTGCTGTA 3′ | ||
| CD3 | NM_000073 | Forward: 5′ CTGGGAAGTAATGCCAAGGA 3′ |
| Reverse: 5′ CCAACAGCAAGGACGAAAAT 3′ | ||
| CD68 | NM_001040059 | Forward: 5′ CCACCTGCTTCTCTCATTCC 3′ |
| Reverse: 5′ ATTGTACTCCACCGCCATGT 3′ | ||
| CD80 | NM_005191 | Forward: 5′ GCAGGGAACATCACCATCCA 3′ |
| Reverse: 5′ CAGGACAGCGTTGCCACTTC 3′ | ||
| CD206 | NM_002438 | Forward: 5′ ACTGGGGCCAAGCTTCTCTG 3′ |
| Reverse: 5′ CACAGCCACGTCCCTTCAAC 3′ | ||
| MIF | NM_002415 | Forward: 5′ GGTTCCTCTCCGAGCTCACC 3′ |
| Reverse: 5′ TAGACCCTGTCCGGGCTGAT 3′ | ||
| PGP9.5 | NM_004181 | Forward: 5′ CCATACAGGCAGCCCATGAT 3′ |
| Reverse: 5′ AGACCTTGGCAGCGTCCTTC 3′ | ||
| GAP43 | NM_002045 | Forward: 5′ CCGGCAAAGCAGGAGAAACT 3′ |
| Reverse: 5′ TGGAGGACGGCGAGTTATCA 3′ | ||
| COX2 | NM_000963 | Forward: 5′ TAGAGCCCTTCCTCCTGTGC 3′ |
| Reverse: 5′ TGGGGATCAGGGATGAACTT 3′ | ||
| FABP4 | NM_001442 | Forward: 5′ ACTGGGCCAGGAATTTGACG 3′ |
| Reverse: 5′ ATGACGCATTCCACCACCAG 3′ | ||
| PPARγ | NM_015869 | Forward: 5′ AAAGTCCTTCCCGCTGACCA 3′ |
| Reverse: 5′ GGCCACCTCTTTGCTCTGCT 3′ | ||
| ADIPOQ | NM_004797 | Forward: 5′ TGACCAGGAAACCACGACTCA 3′ |
| Reverse: 5′ CCGATGTCTCCCTTAGGACCA 3′ | ||
| CEBPα | NM_004364 | Forward: 5′ AAGGCCAAGAAGTCGGTGGA 3′ |
| Reverse: 5′ GGCGGTCATTGTCACTGGTC 3′ | ||
| SCX | Commercial Quantitect primer Assay Hs_SCXB_2_SG | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klatte-Schulz, F.; Minkwitz, S.; Schmock, A.; Bormann, N.; Kurtoglu, A.; Tsitsilonis, S.; Manegold, S.; Wildemann, B. Different Achilles Tendon Pathologies Show Distinct Histological and Molecular Characteristics. Int. J. Mol. Sci. 2018, 19, 404. https://doi.org/10.3390/ijms19020404
Klatte-Schulz F, Minkwitz S, Schmock A, Bormann N, Kurtoglu A, Tsitsilonis S, Manegold S, Wildemann B. Different Achilles Tendon Pathologies Show Distinct Histological and Molecular Characteristics. International Journal of Molecular Sciences. 2018; 19(2):404. https://doi.org/10.3390/ijms19020404
Chicago/Turabian StyleKlatte-Schulz, Franka, Susann Minkwitz, Aysha Schmock, Nicole Bormann, Alper Kurtoglu, Serafeim Tsitsilonis, Sebastian Manegold, and Britt Wildemann. 2018. "Different Achilles Tendon Pathologies Show Distinct Histological and Molecular Characteristics" International Journal of Molecular Sciences 19, no. 2: 404. https://doi.org/10.3390/ijms19020404
APA StyleKlatte-Schulz, F., Minkwitz, S., Schmock, A., Bormann, N., Kurtoglu, A., Tsitsilonis, S., Manegold, S., & Wildemann, B. (2018). Different Achilles Tendon Pathologies Show Distinct Histological and Molecular Characteristics. International Journal of Molecular Sciences, 19(2), 404. https://doi.org/10.3390/ijms19020404