7-Methylguanosine Modifications in Transfer RNA (tRNA)


Abstract
1. Introduction
2. Structural Analyses and Catalytic Mechanisms of m7G Methyltransferases
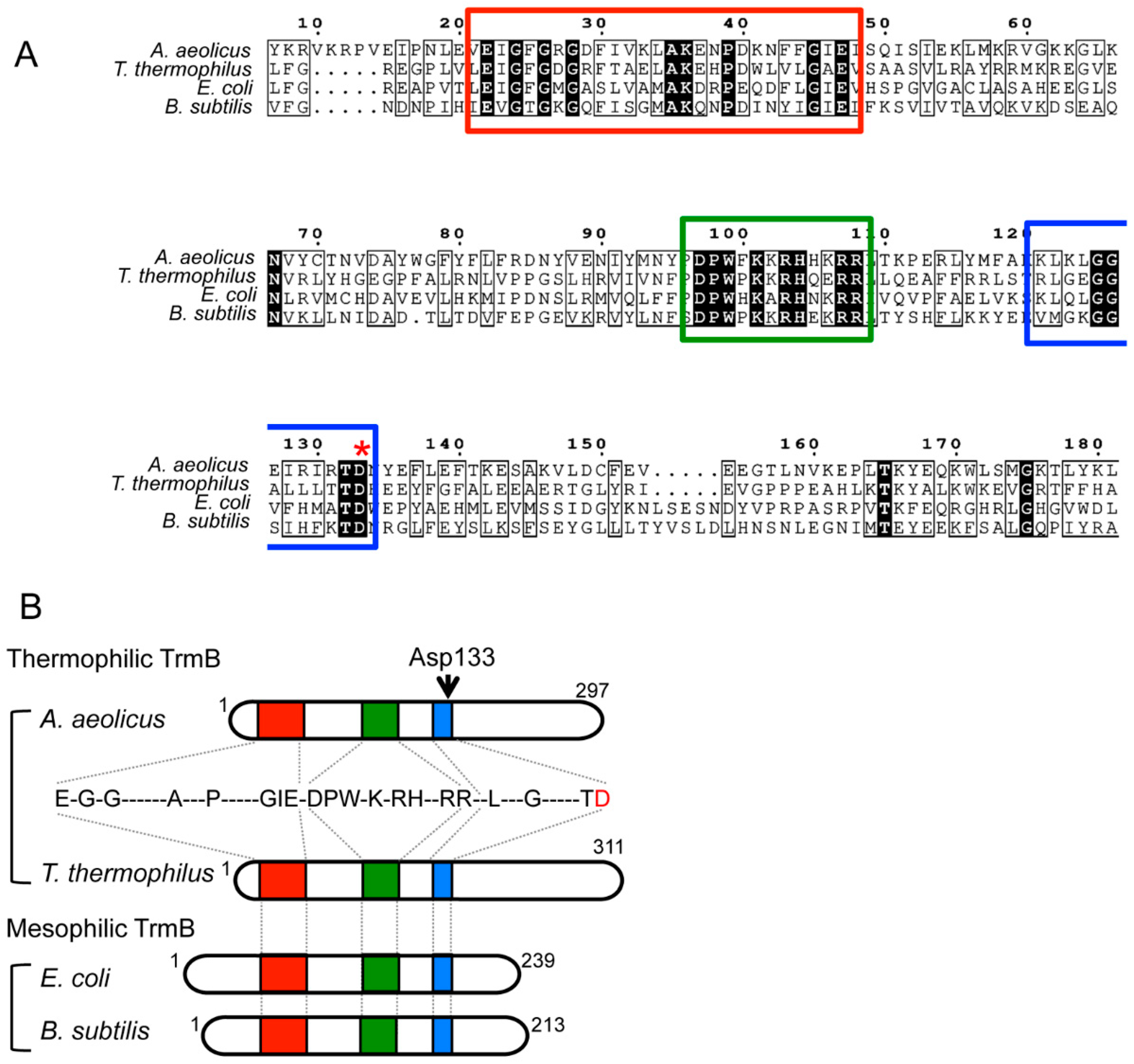
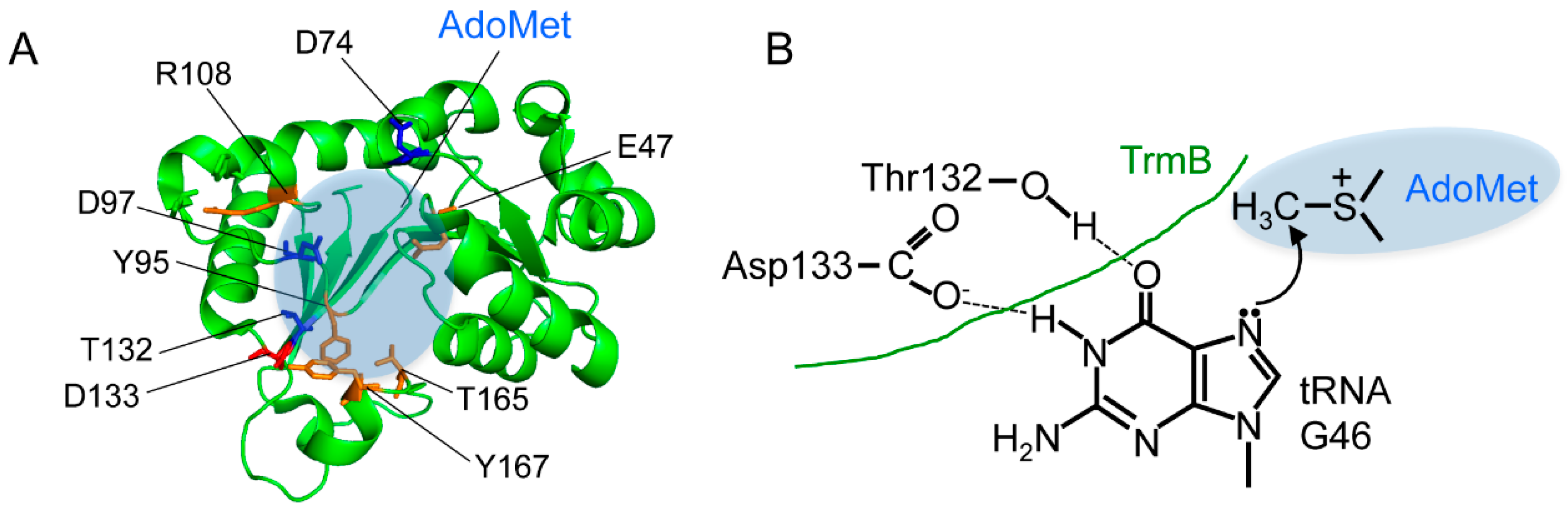
2.1. Eubacterial tRNA m7G46 Methyltransferases (TrmB)
2.2. Heterodimeric tRNA m7G Methyltransferase of Yeast (Trm8/Trm82)
2.3. mRNA Cap m7G Methyltransferase
3. Physiological Functions
3.1. tRNA m7G46 Modification in Yeast
3.2. tRNA m7G46 Modification in Thermophilic Eubacteria
3.3. Involvement of tRNA m7G46 Modification in Fungal Pathogenicity
3.4. Involvement of tRNA m7G46 Methyltransferase in Diseases
4. Perspective
Funding
Acknowledgements
Conflicts of Interest
References
- Kim, S.H.; Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sussman, J.L.; Wang, A.H.J.; Seeman, N.C.; Rich, A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science 1974, 185, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Limbach, P.A.; Crain, P.F.; Mccloskey, J.A. Summary: The modified nucleosides of RNA. Nucleic Acids Res. 1994, 22, 2183–2196. [Google Scholar] [CrossRef] [PubMed]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Pi Atkowski, P.-L.; Zej Bagí Nski, B.-L.; Wirecki, T.K.; De Crécy, V.; Crécy-Lagard, C.; Ross, R.; Limbach, P.A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2017, 46, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Juhling, F.; Morl, M.; Hartmann, R.K.; Sprinzl, M.; Stadler, P.F.; Putz, J. tRNAdb 2009: Compilation of tRNA sequences and tRNA genes. Nucleic Acids Res. 2009, 37, D159–D162. [Google Scholar] [CrossRef] [PubMed]
- Schubert, H.L.; Blumenthal, R.M.; Cheng, X. Many paths to methyltransfer: A chronicle of convergence. Trends Biochem. Sci. 2003, 28, 329–335. [Google Scholar] [CrossRef]
- Hori, H. Methylated nucleosides in tRNA and tRNA methyltransferases. Front. Genet. 2014, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, C.G.; Crain, P.F.; Gupta, R.; Hashizume, T.; Hocart, C.H.; Kowalak, J.A.; Pomerantz, S.C.; Stetter, K.O.; McCloskey, J.A. Posttranscriptional modification of tRNA in Thermophilic archaea (Archaebacteria). J. Bacteriol. 1991, 173, 3138–3148. [Google Scholar] [CrossRef] [PubMed]
- Dalluge, J.J.; Hamamoto, T.; Horikoshi, K.; Morita, R.Y.; Stetter, K.O.; McCloskey, J.A. Posttranscriptional modification of tRNA in psychrophilic bacteria. J. Bacteriol. 1997, 179, 1918–1923. [Google Scholar] [CrossRef] [PubMed]
- Stange, N.; Beier, H. A cell-free plant extract for accurate pre-tRNA processing, splicing and modification. EMBO J. 1987, 6, 2811–2818. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Sussman, J.L.; Suddath, F.L.; Quigley, G.J.; Mcpherson, A.; Wang, A.H.J.; Seeman, N.C.; Rich, A. The General Structure of Transfer RNA Molecules (base stacking/hydrogen bonding/tRNA sequences/tRNA conformation). Proc. Natl. Acad. Sci. USA 1974, 71, 4970–4974. [Google Scholar] [CrossRef]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 A resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Rich, A.; RajBhandary, U.L. Transfer RNA: Molecular Structure, Sequence, and Properties. Annu. Rev. Biochem. 1976, 45, 805–860. [Google Scholar] [CrossRef]
- Quigley, G.J.; Rich, A. Structural domains of transfer RNA molecules. Science 1976, 194, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Henkin, T.M.; Agris, P.F. tRNA regulation of gene expression: Interactions of an mRNA 5’-UTR with a regulatory tRNA. RNA 2006, 12, 1254–1261. [Google Scholar] [CrossRef]
- Agris, P.F.; Sierzputowska-Gracz, H.; Smith, C. Transfer RNA contains sites of localized positive charge: Carbon NMR studies of [13C]methyl-enriched Escherichia coli and yeast tRNAPhe. Biochemistry 1986, 25, 5126–5131. [Google Scholar] [CrossRef] [PubMed]
- Jakab, G.; Kis, M.; Palfi, Z.; Solymosy, F. Nucleotide sequence of chloroplast tRNALeu/UA m7G/from Chlamydomonas reinhardtii. Nucleic Acids Res. 1990, 18, 7444. [Google Scholar] [CrossRef]
- Matsuyama, S.; Ueda, T.; Crain, P.F.; Mccloskey, J.A.; Watanabe, K. A Novel Wobble Rule Found in Starfish Mitochondria. J. Biol. Chem. 1998, 273, 3363–3368. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Ueda, T.; Watanabe, K. 7-Methylguanosine at the anticodon wobble position of squid mitochondrial tRNA Ser GCU: Molecular basis for assignment of AGA/AGG codons as serine in invertebrate mitochondria. Biochim. Biophys. Acta 1998, 1399, 78–82. [Google Scholar] [CrossRef]
- Watanabe, K.; Yokobori, S.-I. tRNA Modification and Genetic Code Variations in Animal Mitochondria. Res. J. Nucleic Acids 2011, 2011. [Google Scholar] [CrossRef]
- Tomikawa, C.; Ohira, T.; Inoue, Y.; Kawamura, T.; Yamagishi, A.; Suzuki, T.; Hori, H. Distinct tRNA modifications in the thermo-acidophilic archaeon, Thermoplasma acidophilum. FEBS Lett. 2013, 587, 3575–3580. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Q.; Yang, W.; Gao, Y.; Teng, M.; Niu, L. Monomeric tRNA (m7G46) methyltransferase from Escherichia coli presents a novel structure at the function-essential insertion. Proteins Struct. Funct. Bioinf. 2009, 76, 512–515. [Google Scholar] [CrossRef]
- Purta, E.; van Vliet, F.; Tricot, C.; de Bie, L.G.; Feder, M.; Skowronek, K.; Droogmans, L.; Bujnicki, J.M. Sequence-structure-function relationships of a tRNA (m7G46) methyltransferase studied by homology modeling and site-directed mutagenesis. Proteins Struct. Funct. Genet. 2005, 59, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Watanabe, K.; Ikeuchi, Y.; Suzuki, T.; Endo, Y.; Hori, H. Substrate tRNA recognition mechanism of tRNA (m7G46) methyltransferase from Aquifex aeolicus. J. Biol. Chem. 2004, 279, 49151–49159. [Google Scholar] [CrossRef]
- Tomikawa, C.; Ochi, A.; Hori, H. The C-terminal region of thermophilic tRNA (m7G46) methyltransferase (TrmB) stabilizes the dimer structure and enhances fidelity of methylation. Proteins 2008, 71, 1400–1408. [Google Scholar] [CrossRef]
- Tomikawa, C.; Takai, K.; Hori, H. Kinetic characterization of substrate-binding sites of thermostable tRNA methyltransferase (TrmB). J. Biochem. 2018, 163, 133–142. [Google Scholar] [CrossRef]
- Tomikawa, C.; Yokogawa, T.; Kanai, T.; Hori, H. N7-Methylguanine at position 46 (m7G46) in tRNA from Thermus thermophilus is required for cell viability at high temperatures through a tRNA modification network. Nucleic Acids Res. 2010, 38, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Zegers, I.; Gigot, D.; van Vliet, F.; Tricot, C.; Phane Aymerich, S.; Bujnicki, J.M.; Kosinski, J.; Droogmans, L. Crystal structure of Bacillus subtilis TrmB, the tRNA (m7G46) methyltransferase. Nucleic Acids Res. 2006, 34, 1925–1934. [Google Scholar] [CrossRef]
- Alexandrov, A.; Martzen, M.R.; Phizicky, E.M. Two proteins that form a complex are required for 7-methylguanosine modification of yeast tRNA. RNA 2002, 8, 1253–1266. [Google Scholar] [CrossRef]
- Leulliot, N.; Chaillet, M.; Durand, D.; Ulryck, N.; Blondeau, K.; van Tilbeurgh, H. Article Structure of the Yeast tRNA m7G Methylation Complex. Structure 2007, 16, 52–61. [Google Scholar] [CrossRef]
- Matsumoto, K.; Tomikawa, C.; Toyooka, T.; Ochi, A.; Takano, Y.; Takayanagi, N.; Abe, M.; Endo, Y.; Hori, H. Production of yeast tRNA (m7G46) methyltransferase (Trm8–Trm82 complex) in a wheat germ cell-free translation system. J. Biotechnol. 2008, 133, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Suzuki, T. Precuesors of tRNAs are stabilized by methylguanosine cap structures. Nat. Chem. Biol. 2016, 12, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Takano, Y.; Takayanagi, N.; Hori, H.; Ikeuchi, Y.; Suzuki, T.; Kimura, A.; Okuno, T. A gene involved in modifying transfer RNA is required for fungal pathogenicity and stress tolerance of Colletotrichum lagenarium. Mol. Microbiol. 2006, 60, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Shatkin, A.J. Capping of eucaryotic mRNAs. Cell 1976, 9, 645–653. [Google Scholar] [CrossRef]
- Furuichi, Y.; Miura, K. A blocked structure at the 5’ terminus of mRNA from cytoplasmic polyhedrosis virus. Nature 1975, 253, 374–375. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Schwer, B.; Shuman, S. Yeast mRNA cap methyltransferase is a 50-kilodalton protein encoded by an essential gene. Mol. Cell. Biol. 1995, 15, 4167–4174. [Google Scholar] [CrossRef] [PubMed]
- Beauclerk, A.A.D.; Cundliffe, E. Sites of action of two ribosomal RNA methylases responsible for resistance to aminoglycosides. J. Mol. Biol. 1987, 193, 661–671. [Google Scholar] [CrossRef]
- Holmes, D.J.; Cundliffe, E. Analysis of a ribosomal RNA methylase gene from Streptomyces tenebrarius which confers resistance to gentamicin. Mol. Gen. Genet. 1991, 229, 229–237. [Google Scholar] [CrossRef]
- Doi, Y.; Wachino, J.-I.; Arakawa, Y. Aminoglycoside Resistance: The Emergence of Acquired 16S Ribosomal RNA Methyltransferases. Infect. Dis. Clin. N. Am. 2016, 30, 523–537. [Google Scholar] [CrossRef]
- Holley, R.W.; Apgar, J.; Everett, G.A.; Madison, J.T.; Marquisee, M.; Merrill, S.H.; Penswick, J.R.; Zamir, A. Structure of a Ribonucleic Acid. Science 1965, 147, 1462–1465. [Google Scholar] [CrossRef]
- Wintermeyer, W.; Zachau, H.G. A specific chemical chain scission of tRNA at 7-methylguanosine. FEBS Lett. 1970, 11, 160–164. [Google Scholar] [CrossRef]
- Wintermeyer, W.; Zachau, H.G. Tertiary structure interactions of 7-methylguanosine in yeast tRNA Phe as studied by borohydride reduction. FEBS Lett. 1975, 58, 306–309. [Google Scholar] [CrossRef]
- Donis-Keller, H.; Maxam, A.M.; Gilbert, W. Mapping adenines, guanines, and pyrimidines in RNA. Nucleic Acids Res. 1977, 4, 2527–2538. [Google Scholar] [CrossRef] [PubMed]
- Behm-Ansmant, I.; Helm, M.; Motorin, Y. Use of specific chemical reagents for detection of modified nucleotides in RNA. J. Nucleic Acids 2011, 2011, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Zueva, V.S.; Mankin, A.S.; Bogdanov, A.A.; Baratova, L.A. Specific fragmentation of tRNA and rRNA at a 7-methylguanine residue in the presence of methylated carrier RNA. Eur. J. Biochem. 1985, 146, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Kahle, D.; Wehmeyer, U.; Char, S.; Krupp, G. The methylation of one specific guanosine in a pre-tRNA prevents cleavage by RNase P and by the catalytic M1 RNA. Nucleic Acids Res. 1990, 18, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Munns, T.W.; Liszewski, M.K.; Sims, H.F. Characterization of antibodies specific for N6-methyladenosine and for 7-methylguanosine. Biochemistry 1977, 16, 2163–2168. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ikeuchi, Y.; Noma, A.; Suzuki, T.; Sakaguchi, Y. Mass spectrometric identification and characterization of RNA-modifying enzymes. Methods Enzymol. 2007, 425, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Marchand, V.; Ayadi, L.; Ernst, F.G.M.; Hertler, J.; Bourguignon-Igel, V.; Galvanin, A.; Kotter, A.; Helm, M.; Lafontaine, D.L.J.; Motorin, Y. AlkAniline-Seq: Profiling of m7G and m3C RNA Modifications at Single Nucleotide Resolution. Angew. Chem. Int. Ed. 2018. [Google Scholar] [CrossRef]
- Hurwitz, J.; Gold, M.; Ander, M. The Enzymatic Methylation of Ribonucleic Acid and Deoxyribonucleic Acid. J. Biol. Chem. 1964, 239, 3462–3473. [Google Scholar]
- Aschhoff, H.J.; Elten, H.; Arnold, H.H.; Mahal, G.; Kersten, W.; Kersten, H. 7-Methylguanine specific tRNA-methyltransferase from Escherichia coli. Nucleic Acids Res. 1976, 3, 3109–3122. [Google Scholar] [CrossRef]
- Cimino, F.; Traboni, C.; Colonna, A.; Izzo, P.; Salvatore, F. Purification and properties of several transfer RNA methyltransferases from S. typhimurium. Mol. Cell. Biochem. 1981, 36, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Colonna, A.; Ciliberto, G.; Santamaria, R.; Cimino, F.; Salvatore, F. Isolation and characterization of a tRNA(guanine-7-)-methyltransferase from Salmonella typhimurium. Mol. Cell. Biochem. 1983, 52, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Morozov, I.A.; Gambaryan, A.S.; Lvova, T.N.; Nedospasov, A.A.; Venkstern, T.V. Purification and Characterization of tRNA (Adenine-1-)-Methyltransferase from Thermus flavus Strain 71. Eur. J. Biochem. 1982, 129, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Koski, R.A.; Clarkson, S.G. Synthesis and maturation of Xenopus laevis methionine tRNA gene transcripts in homologous cell-free extracts. J. Biol. Chem. 1982, 257, 4514–4521. [Google Scholar] [PubMed]
- Drabkin, H.J.; RajBhandary, U.L. Site-specific mutagenesis on a human initiator methionine tRNA gene within a sequence conserved in all eukaryotic initiator tRNAs and studies of its effects on in vitro transcription. J. Biol. Chem. 1985, 260, 5580–5587. [Google Scholar] [PubMed]
- Garcia, G.R.; Goodenough-Lashhua, D.M. Modification and Editing of RNA; Grosjean, B., Ed.; American Society of Microbiology: Washington, DC, USA, 1998; ISBN 9781555818296. [Google Scholar]
- Alexandrov, A.; Grayhack, E.J.; Phizicky, E.M. tRNA m7G methyltransferase Trm8p/Trm82p: Evidence linking activity to a growth phenotype and implicating Trm82p in maintaining levels of active Trm8p. RNA 2005, 11, 821. [Google Scholar] [CrossRef]
- De Bie, L.G.S.; Roovers, M.; Oudjama, Y.; Wattiez, R.; Tricot, C.; Stalon, V.; Droogmans, L.; Bujnicki, J.M. The yggH Gene of Escherichia coli Encodes a tRNA (m7G46) Methyltransferase. J. Bacteriol. 2003, 185, 3238–3243. [Google Scholar] [CrossRef]
- Ahn, H.J.; Kim, H.-W.; Yoon, H.-J.; Lee, B.; Suh, S.W.; Yang, J.K. Crystal structure of tRNA(m1G37)methyltransferase: Insights into tRNA recognition. EMBO J. 2003, 22, 2593–2603. [Google Scholar] [CrossRef]
- Nureki, O.; Watanabe, K.; Fukai, S.; Ishii, R.; Endo, Y.; Hori, H.; Yokoyama, S. Deep Knot Structure for Construction of Active Site and Cofactor Binding Site of tRNA Modification Enzyme. Structure 2004, 12, 593–602. [Google Scholar] [CrossRef]
- Hori, H. Transfer RNA methyltransferases with a SpoU-TrmD (SPOUT) fold and their modified nucleosides in tRNA. Biomolecules 2017, 7, 23. [Google Scholar] [CrossRef]
- Dégut, C.; Ponchon, L.; Folly-Klan, M.; Barraud, P.; Tisné, C. The m1A58 modification in eubacterial tRNA: An overview of tRNA recognition and mechanism of catalysis by TrmI. Biophys. Chem. 2016, 210, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Phan, L.; Hinnebusch, A.G. The Gcd10p/Gcd14p complex is the essential two-subunit tRNA(1-methyladenosine) methyltransferase of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 5173–5178. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, S.K.; Bujnicki, J.M.; Grosjean, H.; Lapeyre, B. Trm11p and Trm112p are both required for the formation of 2-methylguanosine at position 10 in yeast tRNA. Mol. Cell. Biol. 2005, 25, 4359–4370. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, Y.; Wang, C.; Fan, X.; Jiang, X.; Ebrahimi, M.; Qiao, Z.; Niu, L.; Teng, M.; Li, X. Crystal structure of the two-subunit tRNA m1A58 methyltransferase TRM6-TRM61 from Saccharomyces cerevisiae. Sci. Rep. 2016, 6, 32562. [Google Scholar] [CrossRef] [PubMed]
- Mazauric, M.-H.; Dirick, L.; Purushothaman, S.K.; Björk, G.R.; Lapeyre, B. Trm112p is a 15-kDa zinc finger protein essential for the activity of two tRNA and one protein methyltransferases in yeast. J. Biol. Chem. 2010, 285, 18505–18515. [Google Scholar] [CrossRef] [PubMed]
- Guy, M.P.; Phizicky, E.M. Two-subunit enzymes involved in eukaryotic post-transcriptional tRNA modification. RNA Biol. 2014, 11, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Van Tran, N.; Muller, L.; Ross, R.L.; Lestini, R.; Létoquart, J.; Ulryck, N.; Limbach, P.A.; de Crécy-Lagard, V.; Cianférani, S.; Graille, M. Evolutionary insights into Trm112-methyltransferase holoenzymes involved in translation between archaea and eukaryotes. Nucleic Acids Res. 2018, 46, 8483–8499. [Google Scholar] [CrossRef]
- Matsumoto, K.; Toyooka, T.; Tomikawa, C.; Ochi, A.; Takano, Y.; Takayanagi, N.; Endo, Y.; Hori, H. RNA recognition mechanism of eukaryote tRNA (m7G46) methyltransferase (Trm8–Trm82 complex). FEBS Lett. 2007, 581, 1599–1604. [Google Scholar] [CrossRef]
- Fabrega, C.; Hausmann, S.; Shen, V.; Shuman, S.; Lima, C.D. Structure and Mechanism of mRNA Cap (Guanine-N7) Methyltransferase. Mol. Cell 2004, 13, 77–89. [Google Scholar] [CrossRef]
- Shuman, S.; Morham, S.G. Domain structure of vaccinia virus mRNA capping enzyme. Activity of the Mr 95,000 subunit expressed in Escherichia coli. J. Biol. Chem. 1990, 265, 11967–11972. [Google Scholar]
- Higman, M.A.; Bourgeois, N.; Niles, E.G. The vaccinia virus mRNA (guanine-N7-)-methyltransferase requires both subunits of the mRNA capping enzyme for activity. J. Biol. Chem. 1992, 267, 16430–16437. [Google Scholar]
- Cong, P.; Shuman, S. Methyltransferase and subunit association domains of vaccinia virus mRNA capping enzyme. J. Biol. Chem. 1992, 267, 16424–16429. [Google Scholar] [PubMed]
- Mao, X.; Shuman, S. Intrinsic RNA (guanine-7) methyltransferase activity of the vaccinia virus capping enzyme D1 subunit is stimulated by the D12 subunit. Identification of amino acid residues in the D1 protein required for subunit association and methyl group transfer. J. Biol. Chem. 1994, 269, 24472–24479. [Google Scholar] [PubMed]
- Niewmierzycka, A.; Clarke, S. S-Adenosylmethionine-dependent methylation in Saccharomyces cerevisiae. Identification of a novel protein arginine methyltransferase. J. Biol. Chem. 1999, 274, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H. Modification and Editing of RNA: Historical Overview and Important Facts to Remember; Springer: Berlin/Heidelberg, Germany, 2005; pp. 1–22. [Google Scholar]
- Hopper, A.K.; Phizicky, E.M. tRNA transfers to the limelight. Genes Dev. 2003, 17, 162–180. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, A.; Chernyakov, I.; Gu, W.; Hiley, S.L.; Hughes, T.R.; Grayhack, E.J.; Phizicky, E.M. Rapid tRNA Decay Can Result from Lack of Nonessential Modifications. Mol. Cell 2006, 21, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Chernyakov, I.; Whipple, J.M.; Kotelawala, L.; Grayhack, E.J.; Phizicky, E.M. Degradation of several hypomodified mature tRNA species in Saccharomyces cerevisiae is mediated by Met22 and the 5’-3’ exonucleases Rat1 and Xrn1. Genes Dev. 2008, 22, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Turowski, T.W.; Karkusiewicz, I.; Kowal, J.; Boguta, M. Maf1-mediated repression of RNA polymerase III transcription inhibits tRNA degradation via RTD pathway. RNA 2012, 18, 1823–1832. [Google Scholar] [CrossRef]
- Karkusiewicz, I.; Turowski, T.W.; Graczyk, D.; Towpik, J.; Dhungel, N.; Hopper, A.K.; Boguta, M. Maf1 protein, repressor of RNA polymerase III, indirectly affects tRNA processing. J. Biol. Chem. 2011, 286, 39478–39488. [Google Scholar] [CrossRef]
- Boguta, M.; Czerska, K.; Żołądek, T. Mutation in a new gene MAF1 affects tRNA suppressor efficiency in Saccharomyces cerevisiae. Gene 1997, 185, 291–296. [Google Scholar] [CrossRef]
- Upadhya, R.; Lee, J.; Willis, I.M. Maf1 Is an Essential Mediator of Diverse Signals that Repress RNA Polymerase III Transcription. Mol. Cell 2002, 10, 1489–1494. [Google Scholar] [CrossRef]
- Watanabe, K.; Shinma, M.; Oshima, T.; Nishimura, S. Heat-induced stability of tRNA from an extreme thermophile, Thermus thermophilus. Biochem. Biophys. Res. Commun. 1976, 72, 1137–1144. [Google Scholar] [CrossRef]
- Horie, N.; Hara-Yokoyama, M.; Yokoyama, S.; Watanabe, K.; Kuchino, Y.; Nishimura, S.; Miyazawa, T. Two tRNAIle1 species from an extreme thermophile, Thermus thermophilus HB8: Effect of 2-thiolation of ribothymidine on the thermostability of tRNA. Biochemistry 1985, 24, 5711–5715. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Suzuki, T.; Terada, T.; Shirouzu, M.; Yokoyama, S.; Watanabe, K. Temperature-dependent biosynthesis of 2-thioribothymidine of Thermus thermophilus tRNA. J. Biol. Chem. 2006, 281, 2104–2113. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Watanabe, K.; Miyazawa, T. Dynamic structures and functions of transfer ribonucleic acids from extreme thermophiles. Adv. Biophys. 1987, 23, 115–147. [Google Scholar] [CrossRef]
- Ishida, K.; Kunibayashi, T.; Tomikawa, C.; Ochi, A.; Kanai, T.; Hirata, A.; Iwashita, C.; Hori, H. Pseudouridine at position 55 in tRNA controls the contents of other modified nucleotides for low-temperature adaptation in the extreme-thermophilic eubacterium Thermus thermophilus. Nucleic Acids Res. 2011, 39, 2304–2318. [Google Scholar] [CrossRef]
- Cartlidge, R.A.; Knebel, A.; Peggie, M.; Alexandrov, A.; Phizicky, E.M.; Cohen, P. The tRNA methylase METTL1 is phosphorylated and inactivated by PKB and RSK in vitro and in cells. EMBO J. 2005, 24, 1696–1705. [Google Scholar] [CrossRef]
- Sakita-Suto, S.; Kanda, A.; Suzuki, F.; Sato, S.; Takata, T.; Tatsuka, M. Aurora-B Regulates RNA Methyltransferase NSUN2. Mol. Biol. Cell 2007, 18, 1107–1117. [Google Scholar] [CrossRef]
- Gustavsson, M.; Ronne, H. Evidence that tRNA modifying enzymes are important in vivo targets for 5-fluorouracil in yeast. RNA 2008, 14, 666–674. [Google Scholar] [CrossRef]
- Okamoto, M.; Fujiwara, M.; Hori, M.; Okada, K.; Yazama, F. tRNA Modifying Enzymes, NSUN2 and METTL1, Determine Sensitivity to 5-Fluorouracil in HeLa Cells. PLoS Genet. 2014, 10, 1004639. [Google Scholar] [CrossRef]
- Wu, J.; Hou, J.H.; Hsieh, T. A new Drosophila gene wh (wuho) with WD40 repeats is essential for spermatogenesis and has maximal expression in hub cells. Dev. Biol. 2006, 296, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, R.; Abdel-Salam, G.M.; Guy, M.P.; Alomar, R.; Abdel-Hamid, M.S.; Afifi, H.H.; Ismail, S.I.; Emam, B.A.; Phizicky, E.M.; Alkuraya, F.S. Mutation in WDR4 impairs tRNA m7G46 methylation and causes a distinct form of microcephalic primordial dwarfism. Genome Biol. 2015, 16. [Google Scholar] [CrossRef]
- Trimouille, A.; Lasseaux, E.; Barat, P.; Deiller, C.; Drunat, S.; Rooryck, C.; Arveiler, B.; Lacombe, D. Further delineation of the phenotype caused by biallelic variants in the WDR4 gene. Clin. Genet. 2018, 93, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, Q.; Lelyveld, V.S.; Choe, J.; Szostak, J.W.; Gregory, R.I. Mettl1/Wdr4-Mediated m7G tRNA Methylome Is Required for Normal mRNA Translation and Embryonic Stem Cell Self-Renewal and Differentiation. Mol. Cell 2018, 71, 244–255. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
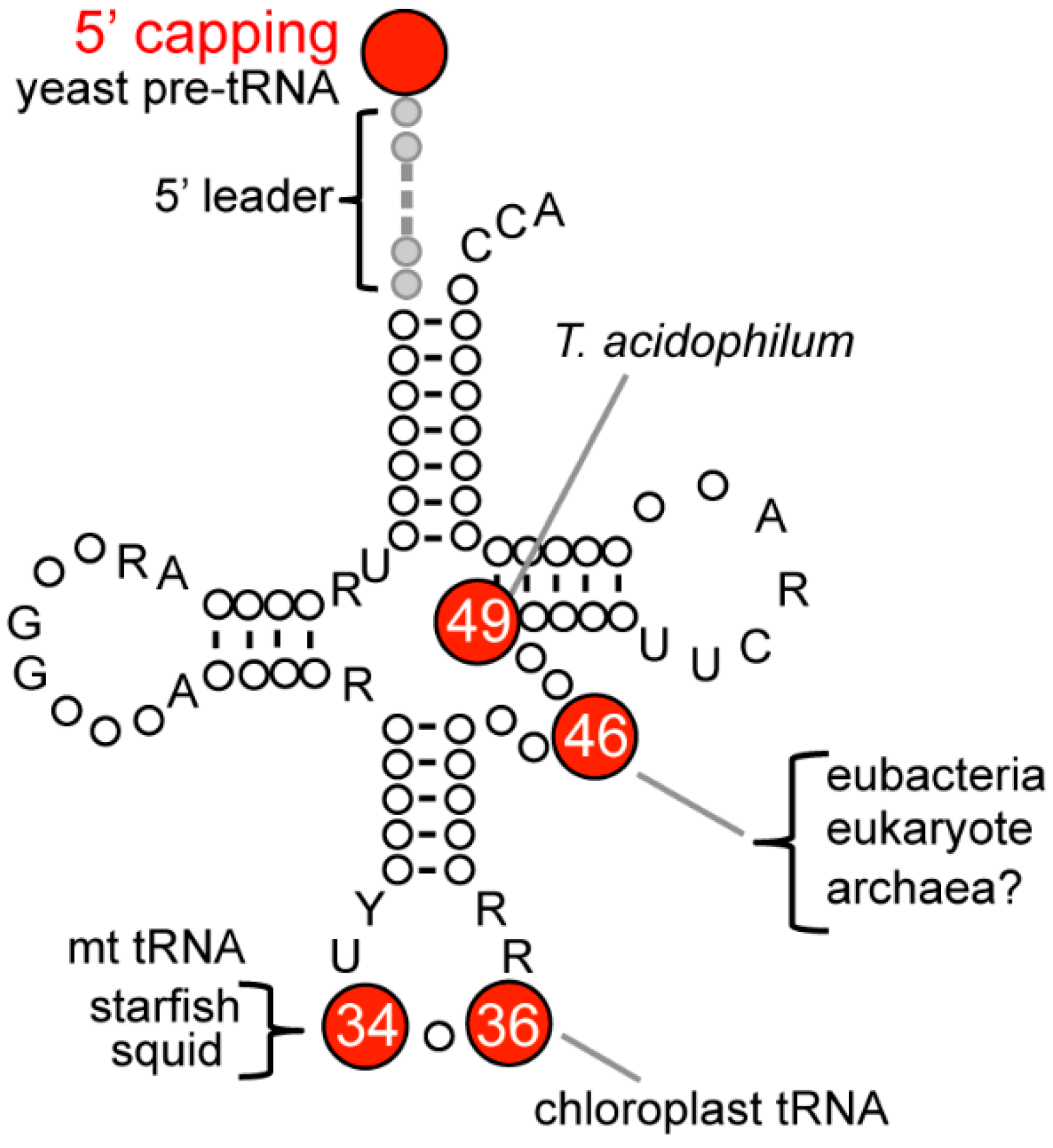
| Domains of Life | Organisms | Positions of m7G | Enzyme Names | Higher Order Structure | References |
|---|---|---|---|---|---|
| eubacteria | E. coli | 46 | TrmB | monomer | [21,22] |
| A. aeolicus | 46 | TrmB | monomer | [23,24,25] | |
| T. thermophilus | 46 | TrmB | ? | [26] | |
| B. subtilis | 46 | TrmB | homodimer | [27] | |
| S. pneumoniae | 46 | TrmB | homodimer | PDB: 1YZH | |
| archaea | T. acidophilum | 49 (tRNALeu(UAG)) | ? | ? | [7,20] |
| T. neutrophilus | ? | ? | ? | [7] | |
| eukaryote | S. cerevisiae | 46 | Trm8/Trm82 | heterodimer | [28,29,30] |
| S. cerevisiae | 5′ termini of pre-tRNA | Ceg1p | ? | [31] | |
| C. lagenarium | 46 | Aph1 | ? | [32] | |
| human | 46 | METTL1/WDR4 | ? | [21,28] | |
| C. reinhardtii | 37 (chloroplast tRNALeu(UAG)) | ? | ? | [16] | |
| A. amurensis | 34 (mt tRNASer(GCU)) | ? | ? | [17,19] | |
| L. bleekeri | 34 (mt tRNASer(GCU)) | ? | ? | [18,19] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomikawa, C. 7-Methylguanosine Modifications in Transfer RNA (tRNA). Int. J. Mol. Sci. 2018, 19, 4080. https://doi.org/10.3390/ijms19124080
Tomikawa C. 7-Methylguanosine Modifications in Transfer RNA (tRNA). International Journal of Molecular Sciences. 2018; 19(12):4080. https://doi.org/10.3390/ijms19124080
Chicago/Turabian StyleTomikawa, Chie. 2018. "7-Methylguanosine Modifications in Transfer RNA (tRNA)" International Journal of Molecular Sciences 19, no. 12: 4080. https://doi.org/10.3390/ijms19124080
APA StyleTomikawa, C. (2018). 7-Methylguanosine Modifications in Transfer RNA (tRNA). International Journal of Molecular Sciences, 19(12), 4080. https://doi.org/10.3390/ijms19124080