Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats



Abstract
1. Introduction
2. Results
2.1. Characterization of Potent Major Compounds
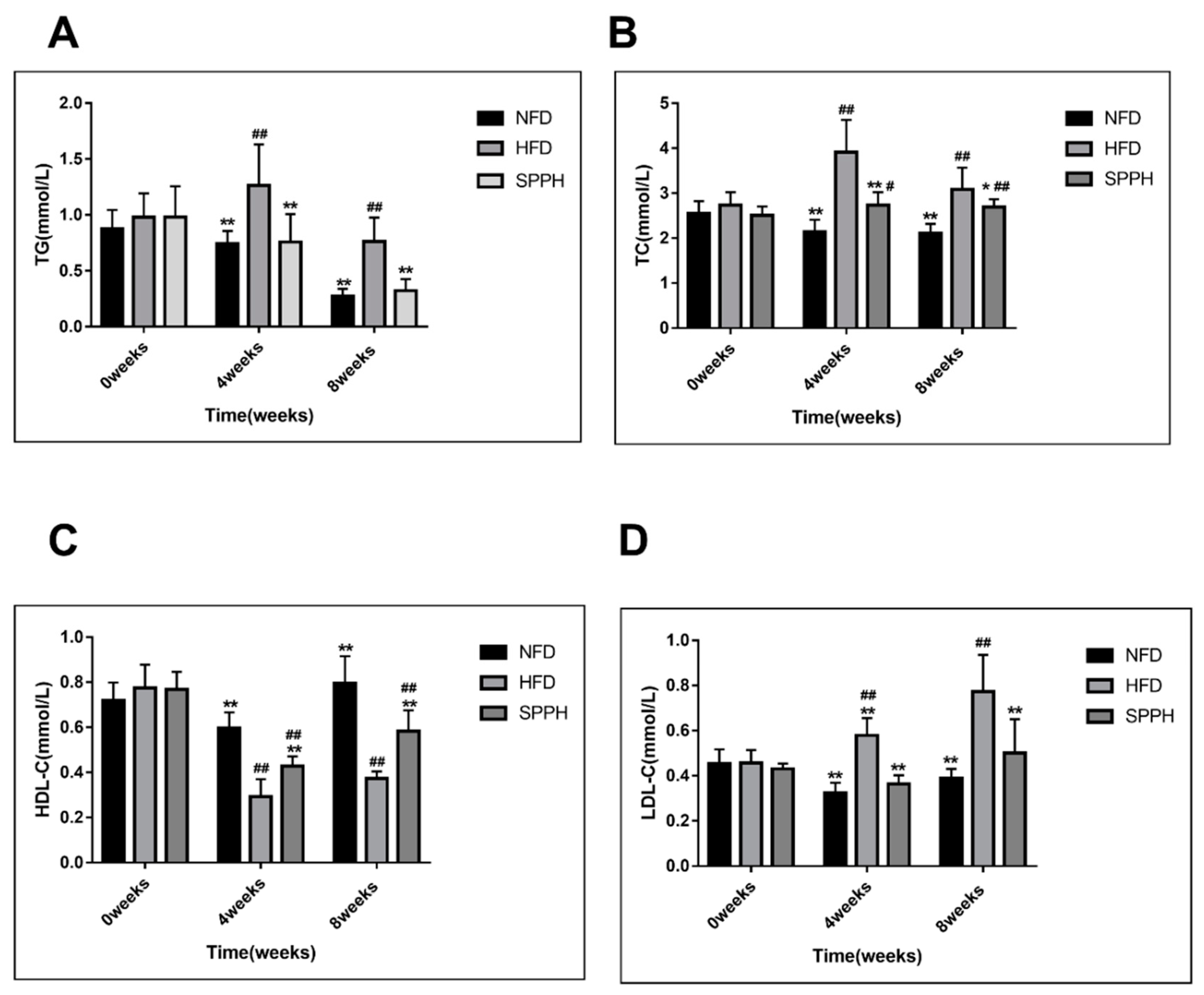
2.2. Effect of SPPH on Body Weight and Serum Lipids of HFD-fed Rats
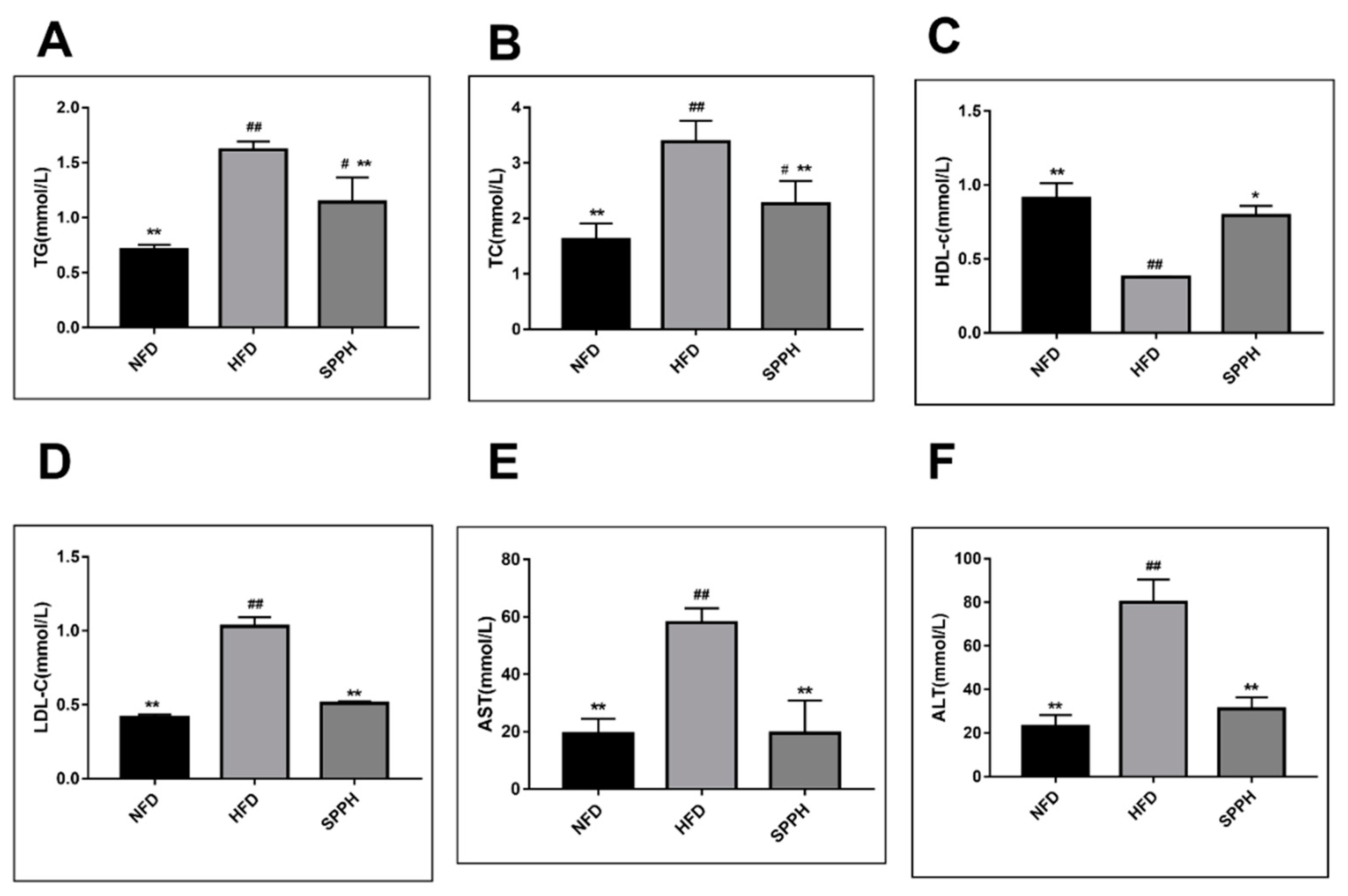
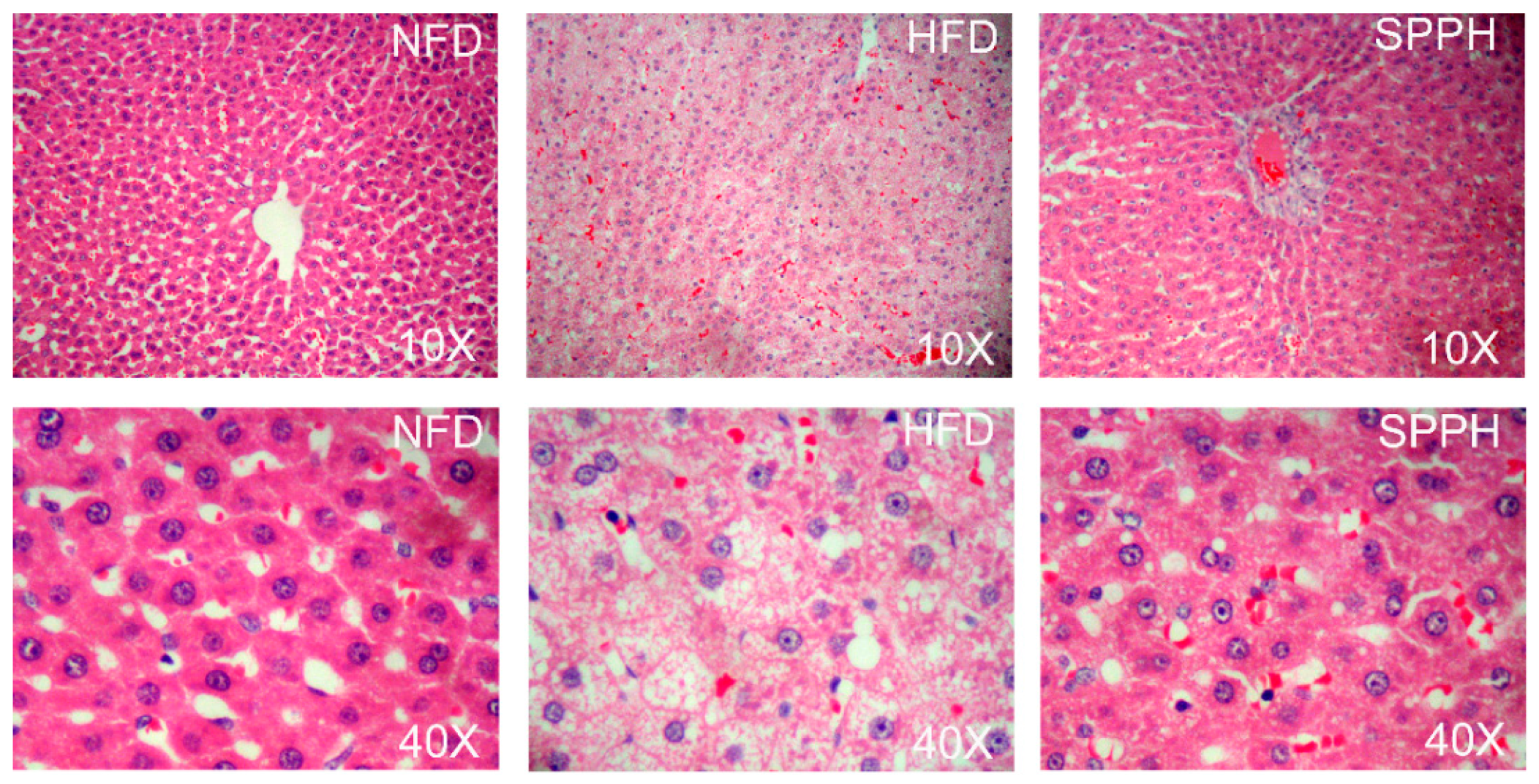
2.3. Effect of SPPH on Liver Function and Hepatic Steatosis
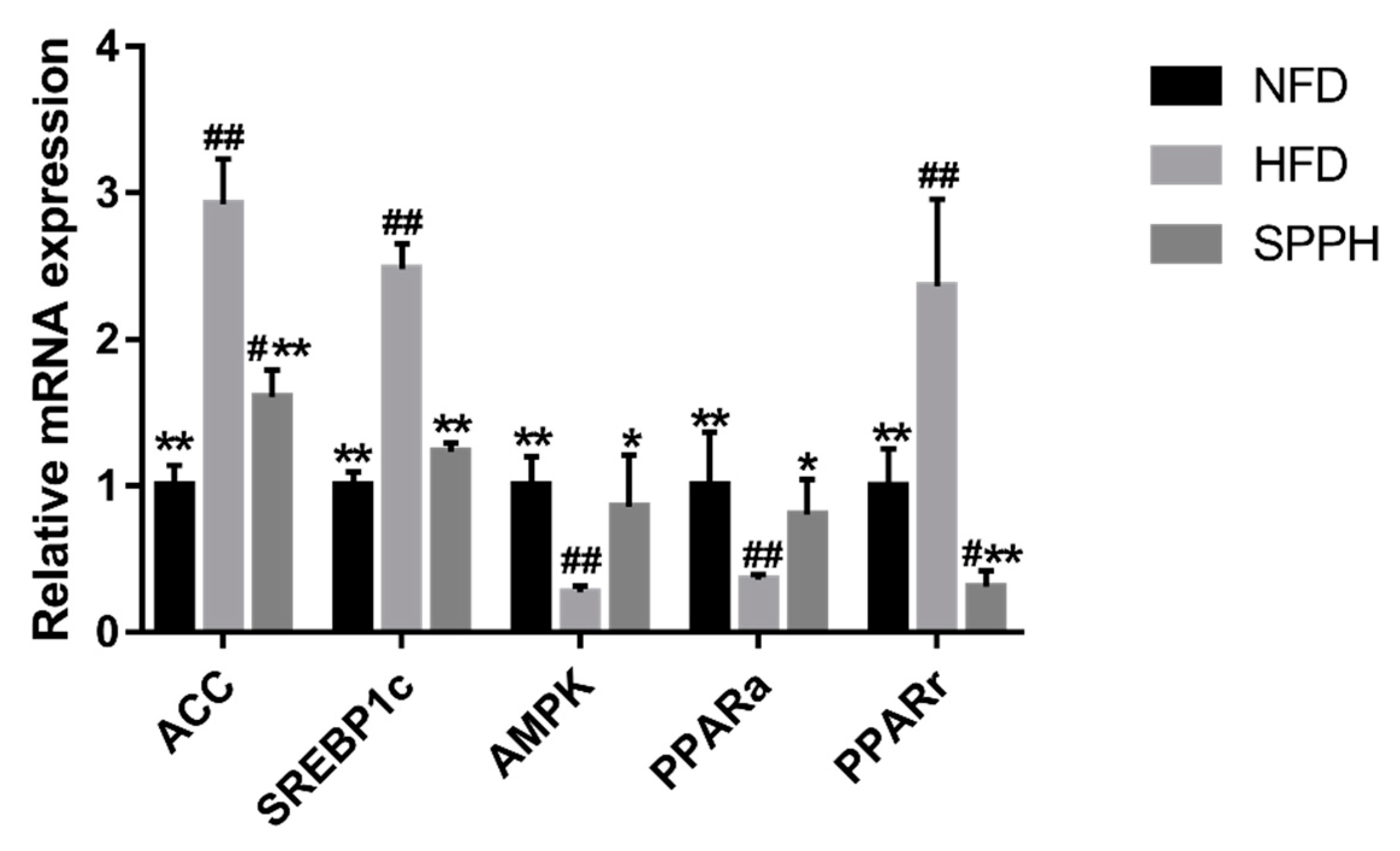
2.4. Effect of SPPH on the Expressions of Genes Involved in Lipid and Glucose Metabolism at the mRNA Level
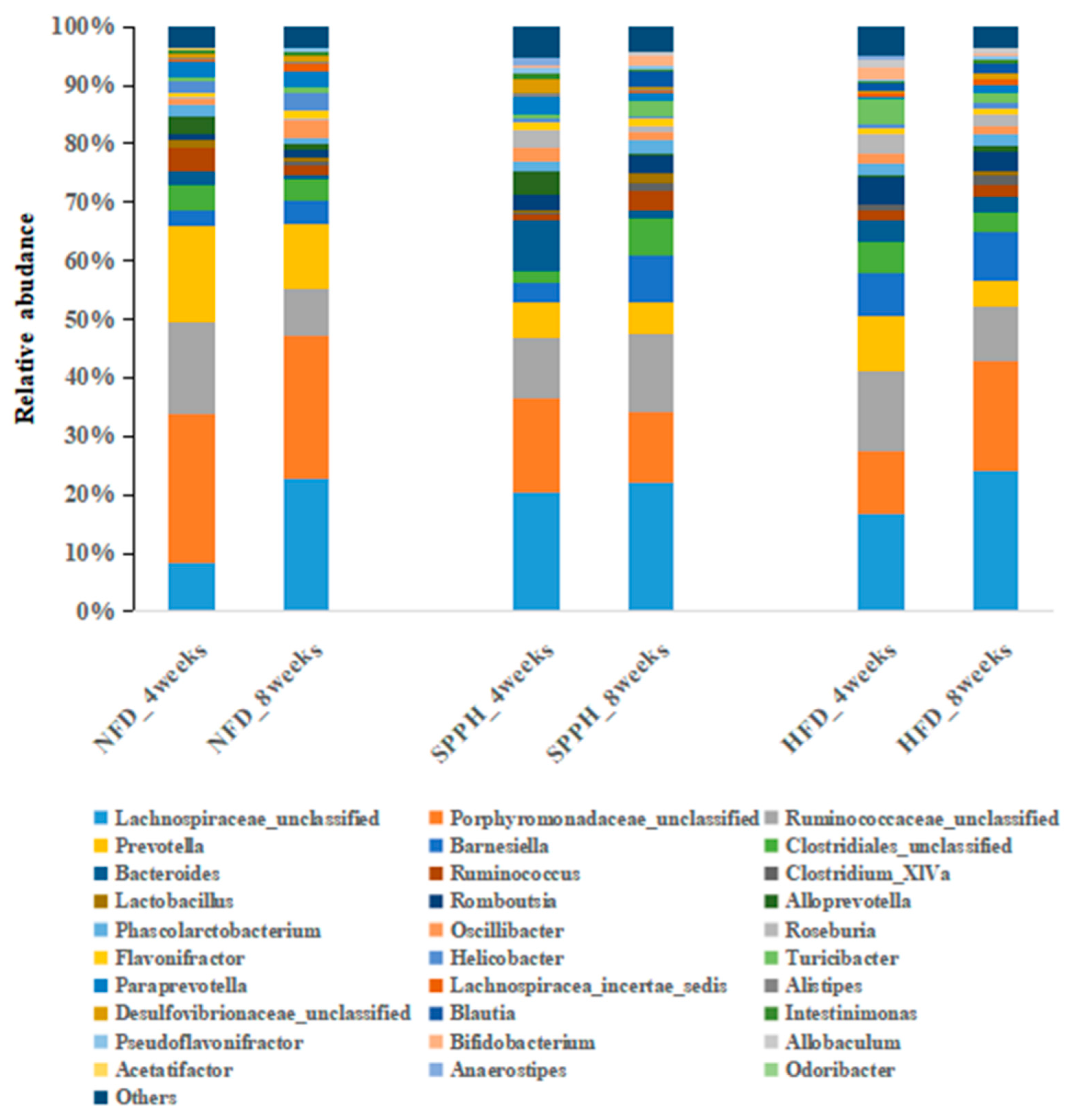
2.5. SPPH Modulates Gut Microbiota of HFD-Fed Rats
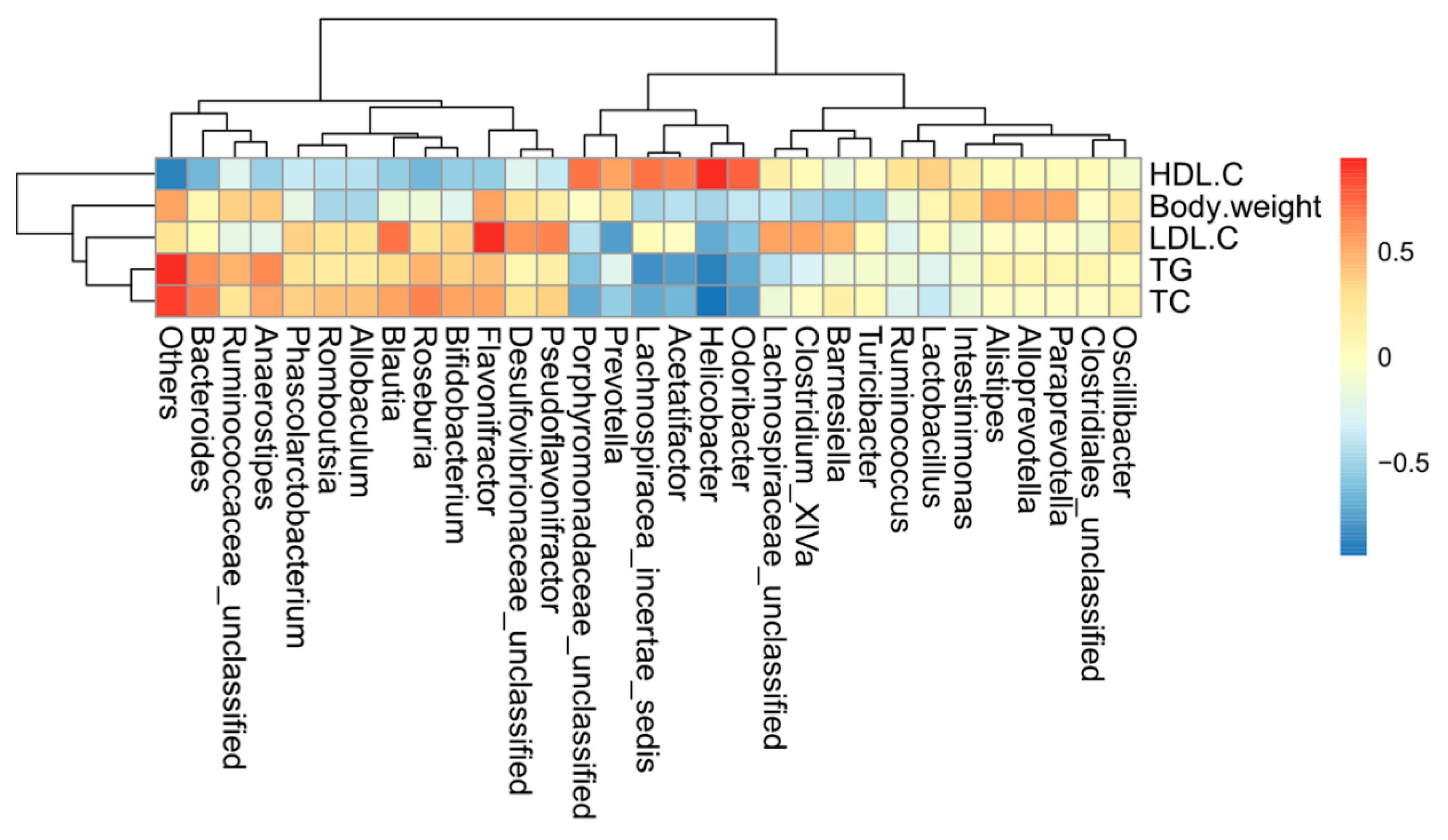
2.6. Correlations of Biochemical Data and Key Phylotypes of Caecal Microbiota
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Preparation of SPPH
4.3. HPLC-MS/MS Analysis of SPPH
4.4. Animals and Experimental Design
4.5. Serum Samples Preparation
4.6. Liver Homogenate Preparation
4.7. Biochemical Assays of Serum and Liver Tissue
4.8. Liver Histopathological Analysis
4.9. mRNA Preparation and Gene Expression
4.10. Dynamic Profile of Intestinal Microflora in Response to SPPH
5. Bioinformatics Analysis
6. Statistical Analysis
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical Approval
References
- Abliz, A.; Aji, Q.; Abdusalam, E.; Sun, X.; Abdurahman, A.; Zhou, W.; Moore, N.; Umar, A. Effect of Cydonia oblonga Mill. leaf extract on serum lipids and liver function in a rat model of hyperlipidaemia. J. Ethnopharmacol. 2014, 151, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Fidèle, N.; Joseph, B.; Emmanuel, T.; Théophile, D. Hypolipidemic, antioxidant and anti-atherosclerogenic effect of aqueous extract leaves of Cassia. occidentalisLinn (Caesalpiniaceae) in diet-induced hypercholesterolemic rats. BMC Complement. Altern. Med. 2017, 17, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Dong, H.; Fang, K.; Gong, J.; Lu, F. Effects of green tea on lipid metabolism in overweight or obese people: A meta-analysis of randomized controlled trials. Mol. Nutr. Food Res. 2017, 62, 1601122–1601134. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.Y.; Jung, U.J.; Park, T.; Yun, J.W.; Choi, M.S. Luteolin attenuates hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue in mice with diet-induced obesity. Diabetes 2015, 64, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Hurt, R.T.; Edakkanambeth, V.J.; Ebbert, J.O. New pharmacological treatments for the management of obesity. Curr. Gastroenterol. Rep. 2014, 16, 394. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.J.; Tschöp, M.H.; Wilding, J.H. Anti-obesity drugs: Past, present and future. Dis. Model. Mech. 2012, 5, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.T.; Chen, H.L.; Wu, H.S.; Ho, M.H.; Chong, K.Y.; Chen, C.M. Kefir Peptides Prevent Hyperlipidemia and Obesity in High Fat Diet-Induced Obese Rats via Lipid Metabolism Modulation. Mol. Nutr. Food Res. 2017, 62. [Google Scholar] [CrossRef]
- Baldassano, S.; Amato, A.; Caldara, G.F.; Mulè, F. Glucagon-like peptide-2 treatment improves glucose dysmetabolism in mice fed a high-fat diet. Endocrine 2016, 54, 1–9. [Google Scholar] [CrossRef]
- Kusumoto, Y.; Irie, J.; Iwabu, K.; Tagawa, H.; Itoh, A.; Kato, M.; Kobayashi, N.; Tanaka, K.; Kikuchi, R.; Fujita, M.; et al. Bile acid binding resin prevents fat accumulation through intestinal microbiota in high-fat diet-induced obesity in mice. Metab. Clin. Exp. 2017, 71, 1–6. [Google Scholar] [CrossRef]
- Zhang, W.L.; Zhu, L.; Jiang, J.G. Active ingredients from natural botanicals in the treatment of obesity. Obes. Rev. 2015, 15, 957–967. [Google Scholar] [CrossRef]
- Le, T.M.; Knulst, A.C.; Röckmann, H. Anaphylaxis to Spirulina confirmed by skin prick test with ingredients of Spirulina tablets. Food Chem. Toxicol. 2014, 74, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Piñero Estrada, J.E.; Bermejo, B.P.; Am, V.D.F. Antioxidant activity of different fractions of Spirulina platensis protean extract. Farmaco 2001, 56, 497–500. [Google Scholar] [CrossRef]
- Abdelkhalek, N.K.; Ghazy, E.W.; Abdel-Daim, M.M. Pharmacodynamic interaction of Spirulina platensis and deltamethrin in freshwater fish Nile tilapia, Oreochromis niloticus: Impact on lipid peroxidation and oxidative stress. Environ. Sci. Pollut. Res. 2015, 22, 3023–3031. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.K.; Van, D.W.J.; Med, M.E.J. Effect of spirulina on the secretion of cytokines from peripheral blood mononuclear cells. Food 2000, 3, 135–175. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Bhadouria, P.; Bisen, P.S. Nutritional and therapeutic potential of Spirulina. Curr. Pharm. Biotechnol. 2005, 6, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Abdeldaim, M.M.; Farouk, S.M.; Madkour, F.F.; Azab, S.S. Anti-inflammatory and immunomodulatory effects of Spirulina platensis in comparison to Dunaliella salina in acetic acid-induced rat experimental colitis. Immunopharmacol. Immunotoxicol. 2015, 37, 126–139. [Google Scholar] [CrossRef]
- Upasan, C.D.; Balaraman, R. Protective effect of Spirulina on lead induced deleterious changes in the lipid peroxidation and endogenous antioxidants in rats. Phytother. Res. 2010, 17, 330–334. [Google Scholar] [CrossRef]
- Upasani, C.D.; Khera, A.; Balarama, R. Effect of lead with vitamin E, C, or Spirulina on malondialdehyde, conjugated dienes and hydroperoxides in rats. Indian J. Exp. Biol. 2001, 39, 70–74. [Google Scholar]
- Zhang, H.Q.; Lin, A.P.; Sun, Y.; Deng, Y.M. Chemo- and radio-protective effects of polysaccharide of Spirulina platensis on hemopoietic system of mice and dogs. Acta Pharmacol. Sin. 2001, 22, 1121–1124. [Google Scholar]
- Iwata, K.; Inayama, T.; Kato, T. Effects of Spirulina platensis on plasma lipoprotein lipase activity in fructose-induced hyperlipidemic rats. J. Nutr. Sci. Vitaminol. 1990, 36, 165. [Google Scholar] [CrossRef]
- Pérez-Vega, J.; Olivera-Castillo, A.L.; Gómez-Rui, J.Á.; Hernández-Ledesma, B. Release of multifunctional peptides by gastrointestinal digestion of sea cucumber (Isostichopus badionotus). J. Funct. Foods 2013, 5, 869–877. [Google Scholar] [CrossRef]
- Balakumar, P.; Balakumar, M.; Arora, K.; Singh, M. Emerging role of PPAR ligands in the management of diabetic nephropathy. Pharmacol. Res. 2009, 60, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, P.; Rose, M.; Singh, M. PPAR ligands: Are they potential agents for cardiovascular disorders? Pharmacology 2007, 80, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Contreras, A.V.; Torres, N.; Tovar, A.R. SPAR-α as a key nutritional and environmental sensor for metabolic adaptation. Advances in Nutrition: An International. Rev. J. 2013, 4, 439–452. [Google Scholar] [CrossRef]
- Yuan, H.D.; Kim, S.J.; Quan, H.Y.; Im, B.O.; Chung, S.H.; Lee, K.H.; Cho, D.H.; Cho, J.Y. Ginseng Leaf Extract Prevents High Fat Diet-Induced Hyperglycemia and Hyperlipidemia through AMPK Activation. J. Ginseng. Res. 2010, 34, 369–375. [Google Scholar] [CrossRef]
- Velagapudi, V.R.; Hezaveh, R.; Reigstad, C.S.; Gopalacharyulu, P.; Yetukuri, L.; Islam, S.; Felin, J.; Perkins, R.; Borén, J.; Oresic, M.; et al. The gut microbiota modulates host energy and lipid metabolism in mice. J. Lipid Res. 2010, 51, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, S.E.; Kirchgessner, A. Gut microbiota and sirtuins in obesity-related inflammation and bowel dysfunction. J. Transl. Med. 2011, 9, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Wu, J. LC-MS/MS coupled with QSAR modeling in characterising of angiotensin I-converting enzyme inhibitory peptides from soybean proteins. Food Chem. 2013, 141, 2682–2690. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.Z.; Li, N.; Tuo, Z.D.; Li, J.L.; Xing, S.S.; Li, B.B.; Zhang, L.; Lee, H.S.; Chen, J.G.; Cui, L. Effects of Morus root bark extract and active constituents on blood lipids in hyperlipidemia rats. J. Ethnopharmacol. 2016, 180, 54–59. [Google Scholar] [CrossRef]
- Song, Y.; Lee, S.J.; Jang, S.H.; Ji, H.H.; Song, Y.M.; Ko, Y.G.; Kim, H.D.; Min, W.; Kang, S.N.; Cho, J.H. Sasa borealis stem extract attenuates hepatic steatosis in high-fat diet-induced obese rats. Nutrients 2014, 6, 2179–2195. [Google Scholar] [CrossRef] [PubMed]
- Lópezmartí, J.; Díazsilva, M.; Salas, A.; Del, M.G.M.; Fernández-López, J.; Remesar, X.; Alemany, M.; Fernándezlópez, J. Oleoyl-estrone induces the massive loss of body weight in Zucker fa/fa rats fed a high-energy hyperlipidic diet. J. Nutr. Biochem. 2000, 11, 530–535. [Google Scholar] [CrossRef]
- Kusunoki, M.; Hara, T.; Tsutsumi, K.; Nakamura, T.; Miyata, T.; Sakakibara, F.; Sakamoto, S.; Ogawa, H.; Nakaya, Y.; Storlien, L.H. The lipoprotein lipase activator, NO-1886, suppresses fat accumulation and insulin resistance in rats fed a high-fat diet. Diabetologia 2000, 43, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.N.; Bok, S.H.; Lee, M.K.; Kim, H.J.; Jeon, S.M.; Do, J.M.; Shin, S.K.; Ha, T.Y.; Choi, M.S. Anti-obesity and hypolipidemic effects of a proprietary herb and fiber combination (S&S PWH) in rats fed high-fat diet. J. Med. Food 2008, 11, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Maebara, Y.; Hayashi, M.; Nagae, R.; Tokuyama, S.; Michinaga, S. Endothelins reciprocally regulate VEGF-A and angiopoietin-1 production in cultured rat astrocytes: Implications on astrocytic proliferation. Glia 2012, 60, 1954–1963. [Google Scholar] [CrossRef]
- Shearer, K.D.; Fragoso, Y.D.; Clagett-Dame, M.; Mccaffery, P.; Tokuyama, S.; Michinaga, S. Astrocytes as a regulated source of retinoic acid for the brain. Glia 2012, 60, 1964–1976. [Google Scholar] [CrossRef]
- Deedwania, P.C.; Pedersen, T.R.; Demicco, D.A.; Breazna, A.; Betteridge, D.J.; Hitman, G.A.; Durrington, P.; Neil, A. Differing predictive relationships between baseline LDL-C, systolic blood pressure, and cardiovascular outcomes. Int. J. Cardiol. 2016, 222, 548–556. [Google Scholar] [CrossRef]
- Wada, H.; Dohi, T.; Miyauchi, K.; Shitara, J.; Endo, H.; Doi, S.; Konishi, H.; Naito, R.; Tsuboi, S.; Ogita, M.; et al. Pre-procedural neutrophil-to-lymphocyte ratio and long-term cardiac outcomes after percutaneous coronary intervention for stable coronary artery disease. Atherosclerosis 2017, 265, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Fujisawa, K.; Ito, E.; Idei, S.; Kawaguchi, N.; Kimoto, M.; Hiemori, M.; Tsuji, H. Improvement of obesity and glucose tolerance by acetate in Type 2 diabetic Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Biosci. Biotechnol. Biochem. 2007, 71, 1236–1243. [Google Scholar] [CrossRef]
- Reddy, J.K.; Rao, M.S. Lipid Metabolism and Liver Inflammation. II. Fatty liver disease and fatty acid oxidation. Am. J. Physiol. 2006, 290, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K.I.; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Bhutani, K.K.; Birari, R.B.; Kapat, K. Potential anti-obesity and lipid lowering natural products: A review. Nat. Prod. Commun. 2007, 2, 331–348. [Google Scholar]
- Sunhee, C.; Miyeon, K.; Daieun, S. Spirulina prevents atherosclerosis by reducing hypercholesterolemia in rabbits fed a high-cholesterol diet. Sci. Vitaminol. 2010, 56, 34–40. [Google Scholar] [CrossRef]
- Jin, D.; Xu, Y.; Mei, X.; Meng, Q.; Gao, Y.; Li, B. Antiobesity and lipid lowering effects of theaflavins on high-fat diet induced obese rats. J. Funct. Foods 2013, 5, 1142–1150. [Google Scholar] [CrossRef]
- Torres-Duran, P.V.; Aldo, F.H.; Juarez-Oropeza, M.A. Antihyperlipemic and antihypertensive effects of Spirulina maximain an open sample of mexican population: A preliminary report. Lipids Health Dis. 2007, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Niaz, M.A.; Ghosh, S. Hypolipidemic and antioxidant effects of Commiphora mukul as an adjunct to dietary therapy in patients with hypercholesterolemia. Cardiovasc. Drugs Ther. 1994, 8, 659–664. [Google Scholar] [CrossRef]
- Birari, R.; Javia, V.; Bhutani, K.K. Antiobesity and lipid lowering effects of Murraya koenigii (L.) Spreng leaves extracts and mahanimbine on high fat diet induced obese rats. Fitoterapia 2010, 81, 1129–1133. [Google Scholar] [CrossRef]
- Chandrasekaran, C.V.; Vijayalakshmim, M.A.; Prakash, K. Review Article: Herbal Approach for Obesity Management. Am. J. Plant Sci. 2012, 3, 1003–1014. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, M.S.; Jo, K.; Hwang, J.K. Piperidine alkaloids from Piper retrofractum Vahl. protect against high-fat diet-induced obesity by regulating lipid metabolism and activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2011, 411, 219–225. [Google Scholar] [CrossRef]
- Sekiya, M.; Yahagi, N.; Matsuzaka, T.; Najima, Y.; Nakakuki, M.; Nagai, R.; Ishibashi, S.; Osuga, J.; Yamada, N.; Shimano, H. Polyunsaturated fatty acids ameliorate hepatic steatosis in obese mice by SREBP-1 suppression. Hepatology 2003, 38, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Shimano, H.; Horton, J.D.; Shimomura, I.; EHammer, R.; Brown, M.S.; Goldstein, J.L. Isoform 1c of sterol regulatory element binding protein is less active than isoform 1a in livers of transgenic mice and in cultured cells. J. Clin. Investig. 1997, 99, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.X.; Shen, W.; Sun, H. Effects of nuclear receptor FXR on the regulation of liver lipid metabolism in patients with non-alcoholic fatty liver disease. Hepatol. Int. 2010, 4, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Senda, M.; Lin, S.; Goto, T.; Yano, M.; Sasaki, T.; Murakami, S.; Kawada, T. Auraptene regulates gene expression involved in lipid metabolism through PPARα activation in diabetic obese mice. Mol. Nutr. Food Res. 2011, 55, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Inada, Y.; Nakano, S.; Tamura, T.; Takahashi, T.; Maruyama, K.; Yamazaki, Y.; Kuroda, J.; Shibata, N. Effects of bezafibrate, PPAR pan-agonist, and GW501516, PPARδ agonist, on development of steatohepatitis in mice fed a methionine- and choline-deficient diet. Eur. J. Pharmacol. 2006, 536, 182–191. [Google Scholar] [CrossRef] [PubMed]
- EAkiyama, T.; Sakai, S.; Lambert, G. Conditional disruption of the peroxisome proliferator-activated receptor gamma gene in mice results in lowered expression of ABCA1, ABCG1, and apoE in macrophages and reduced cholesterol efflux. Mol. Cell. Biol. 2002, 22, 2607–2619. [Google Scholar] [CrossRef]
- Foretz, M.; Viollet, B. Regulation of hepatic metabolism by AMPK. J. Hepatol. 2011, 54, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Wright, H.M.; Wright, M.; Spiegelman, B.M. ADD1/SREBP1 activates PPARγ through the production of endogenous ligand. Proc. Natl. Acad. Sci. USA 1998, 95, 4333–4337. [Google Scholar] [CrossRef]
- Berger, J.P. Role of PPARgamma, transcriptional cofactors, and adiponectin in the regulation of nutrient metabolism, adipogenesis and insulin action: View from the chair. Int. J. Obes. 2005, 29, S3. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D.E. The mechanism of action of PPARs. Ann. Rev. Med. 2002, 53, 409–444. [Google Scholar] [CrossRef] [PubMed]
- Lei, F.; Zhang, X.N.; Zhang, W.; Wang, D.; Xie, W.; Su, H.; Du, L.J. Evidence of anti-obesity effects of the pomegranate leaf extract in high-fat diet induced obese mice. Int. J. Obes. 2007, 31, 1023. [Google Scholar] [CrossRef] [PubMed]
- Herbert, T.; Moschen, A.R.; Kaneider, N.C. Pathways of liver injury in alcoholic liver disease. J. Hepatol. 2011, 55, 1159–1161. [Google Scholar] [CrossRef]
- Dolan, K.T.; Chang, E.B. Diet, gut microbes, and the pathogenesis of inflammatory bowel diseases. Mol. Nutr. Food Res. 2016, 61, 1–38. [Google Scholar] [CrossRef]
- Kimura, R.; Takahashi, N.; Murota, K.; Yamada, Y.; Niiya, S.; Kanzaki, N.; Murakami, Y.; Moriyama, T.; Goto, T.; Kawada, T. Activation of peroxisome proliferator-activated receptor-α (PPARα) suppresses postprandial lipidemia through fatty acid oxidation in enterocytes. Biochem. Biophys. Res. Commun. 2011, 410, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Taminiau, B.; Walgrave, H.; Daube, G.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota? Nutrients 2017, 9, 633–642. [Google Scholar] [CrossRef]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.M.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K.; et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef]
- Ubeda, C. Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect. Immun. 2013, 81, 965–973. [Google Scholar] [CrossRef]
- Shang, Q.; Song, G.; Zhang, M.; Zhang, M.F.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia, population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G Protein-coupled Receptors GPR41 and GPR43 Are Activated by Propionate and Other Short Chain Carboxylic Acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Henaomejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.L.; Pan, Y.Y.; Li, L.; Li, T.T.; Lv, X.C. Ethanol extract of Ganoderma lucidum ameliorates lipid metabolic disorders and modulates the gut microbiota composition in high-fat diet fed rats. Food Funct. 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.N.; Wang, S.Y.; Huang, S.L.; Huang, Y.F. Optimization of Chelation of Whey Protein Hydrolysate with Calcium Using Response Surface Method. Adv. Mater. Res. 2013, 781–784, 1856–1860. [Google Scholar] [CrossRef]
- Luo, Y.C.; Gang, C.; Bo, L.; Guo, Y.; Yetukuri, L.; VIslam, S.; Felin, J.; Perkins, R.; Borén, J.; Oresic, M.; et al. Evaluation of antioxidative and hypolipidemic properties of a novel functional diet formulation of Auricularia auricula and Hawthorn. Innov. Food Sci. Emerg. 2009, 10, 215–221. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–505. [Google Scholar] [PubMed]
- Cheng, H.; Xu, N.; Zhao, W.; Su, J.; Lian, M.; Xie, Z.; Wu, X.; Li, Q. (−)-Epicatechin regulates blood lipids and attenuates hepatic steatosis in rats fed high-fat diet†. Mol. Nutr. Food Res. 2017, 61, 1700303. [Google Scholar] [CrossRef]
- Mashmoul, M.; Azlan, A.; Mohtarrudin, N. Protective effects of saffron extract and crocin supplementation on fatty liver tissue of high-fat diet-induced obese rats. BMC Complem. Altern. M. 2016, 16, 401–405. [Google Scholar] [CrossRef]
- Hernándezrodas, M.C.; Valenzuela, R.; Echeverría, F.; Rincón-Cervera, M.Á.; Espinosa, A.; Illesca, P.; Muñoz, P.; Corbari, A.; Romero, N.; Gonzalez-Mañan, D.; et al. Supplementation with Docosahexaenoic Acid and Extra Virgin Olive Oil Prevents Liver Steatosis Induced by a High-Fat Diet in Mice through PPAR-α and Nrf2 Upregulation with Concomitant SREBP-1c and NF-kB Downregulation. Mol. Nutr. Food Res. 2017, 61, 456–469. [Google Scholar] [CrossRef]
- Li, K.; Zhuo, C.; Teng, C.; Yu, S.; Wang, X.; Hu, Y.; Ren, G.; Yu, M.; Qu, J. Effects of Ganoderma lucidum, polysaccharides on chronic pancreatitis and intestinal microbiota in mice. Int. J. Biol. Macromol. 2016, 93, 904–912. [Google Scholar] [CrossRef]
- Henschel, A.; Anwar, M.Z.; Manohar, V. Comprehensive Meta-analysis of Ontology Annotated 16S rRNA Profiles Identifies Beta Diversity Clusters of Environmental Bacterial Communities. PLoS Comput. Biol. 2015, 11, e1004468. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Weight (g) | ||
|---|---|---|---|
| NFD | HFD | SPPH | |
| 0 Weeks | 223.52 ± 6.15 | 224.28 ± 6.58 | 227.08 ± 9.84 |
| 4 Weeks | 382.69 ± 31.65 * | 418.53 ± 18.48 # | 363.02 ± 42.65 ** |
| 8 Weeks | 374.58 ± 16.20 ** | 412.05 ± 19.21 ## | 354.09 ± 13.11 ** |
| Weight gain | 163.14 ± 14.82 ** | 215.36 ± 23.70 ## | 169.83 ± 42.68 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hua, P.; Yu, Z.; Xiong, Y.; Liu, B.; Zhao, L. Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. Int. J. Mol. Sci. 2018, 19, 4023. https://doi.org/10.3390/ijms19124023
Hua P, Yu Z, Xiong Y, Liu B, Zhao L. Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. International Journal of Molecular Sciences. 2018; 19(12):4023. https://doi.org/10.3390/ijms19124023
Chicago/Turabian StyleHua, Pengpeng, Zhiying Yu, Yu Xiong, Bin Liu, and Lina Zhao. 2018. "Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats" International Journal of Molecular Sciences 19, no. 12: 4023. https://doi.org/10.3390/ijms19124023
APA StyleHua, P., Yu, Z., Xiong, Y., Liu, B., & Zhao, L. (2018). Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. International Journal of Molecular Sciences, 19(12), 4023. https://doi.org/10.3390/ijms19124023