Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection

 ,
, 
 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
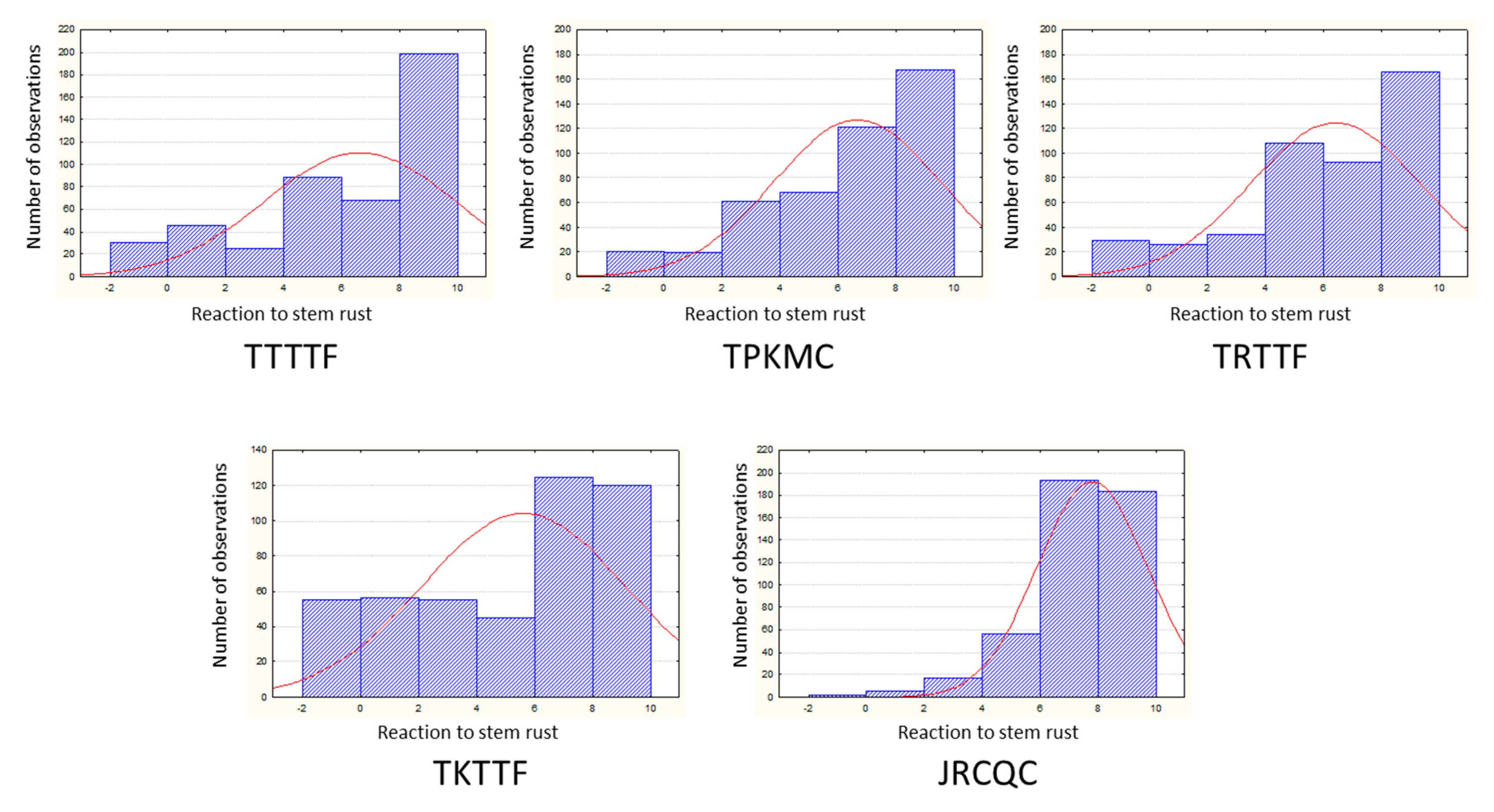
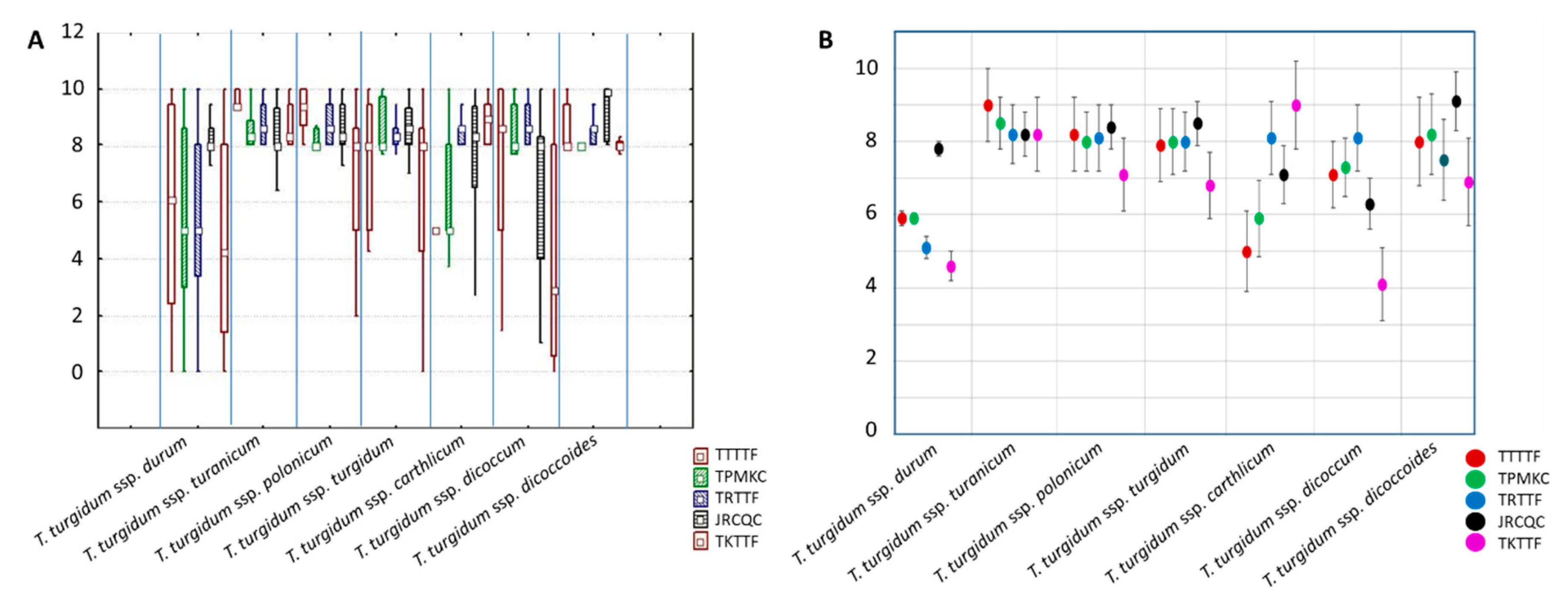
2.1. Evaluation of the Tetraploid Wheat Collection for Resistance to Stem Rust
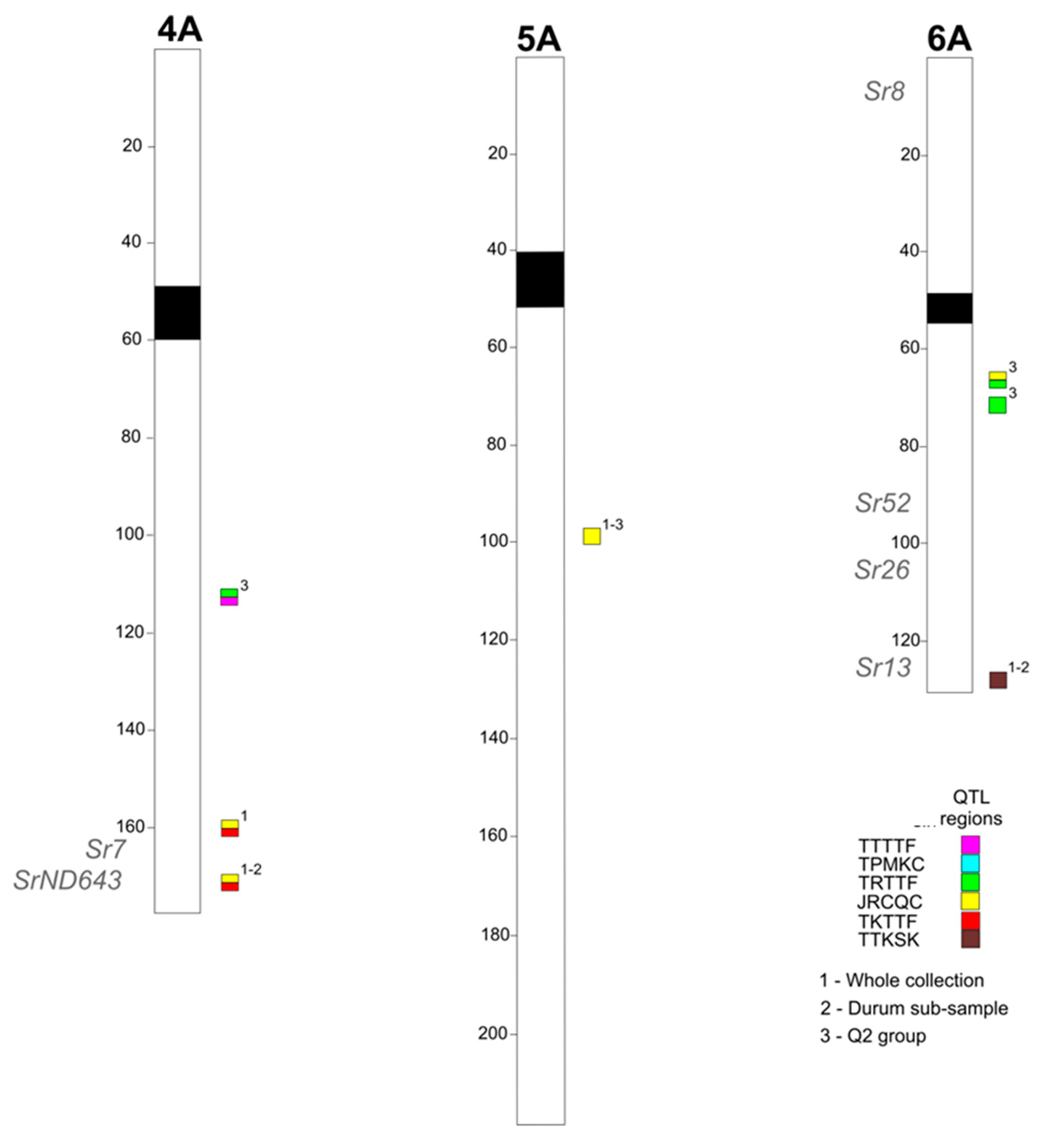
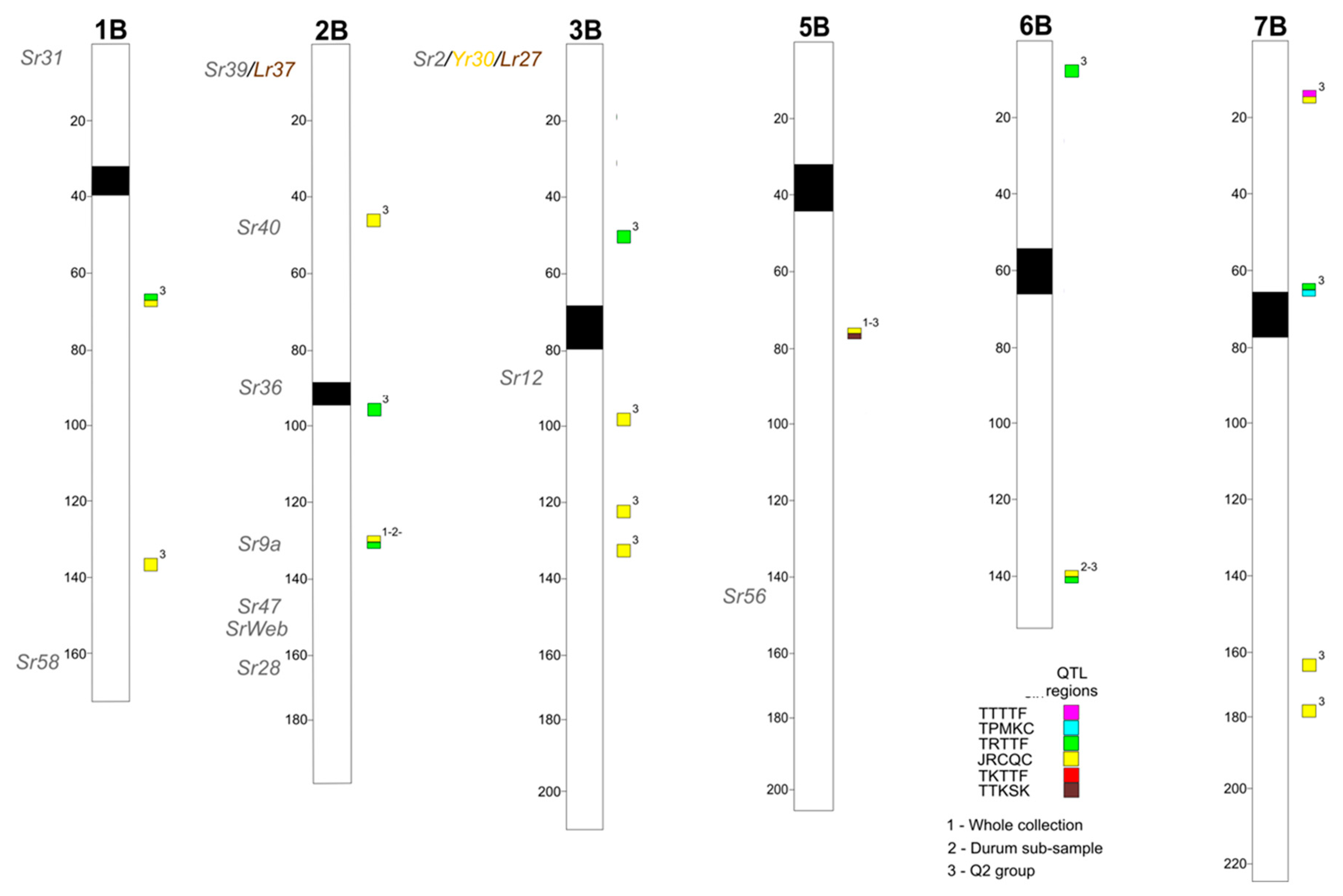
2.2. Association Mapping
2.3. Search for Disease-Related Genes in the Chromosome Regions Corresponding to QTLs for Stem Rust Resistance
3. Discussion
4. Materials and Methods
4.1. Plant Material and Pgt Races
4.2. Phenotypic Evaluation and Statistical Analysis
4.3. Genotyping
4.4. Association Mapping Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Letta, T.; Olivera, P.; Maccaferri, M.; Jin, Y.; Ammar, K.; Badebo, A.; Salvi, S.; Noli, E.; Crossa, J.; Tuberosa, R. Association mapping reveals novel stem rust resistance loci in durum wheat at the seedling stage. Plant Genome 2014, 7, 1–13. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Crossa, J.; Yuen, J.; Djurle, A. Effect of leaf rust on grain yield and yield traits of durum wheats with race-specific and slow-rusting resistance to leaf rust. Plant Dis. 2006, 90, 1065–1072. [Google Scholar] [CrossRef]
- Stakman, E.C.; Harrar, J.G. Principles of Plant Pathology; Ronald Press Co.: New York, NY, USA, 1957. [Google Scholar]
- Eversmeyer, M.G.; Kramer, C.L. Epidemiology of wheat leaf and stem rust in the central Great Plains of the USA. Annu. Rev. Phytopathol. 2000, 38, 491–513. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Ann. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.; Morris, C.; Appels, R.; Xia, X. Catalogue of gene symbols for wheat: 2015–2016 supplement. Ann. Wheat Newsl. 2016, 58, 1–18. [Google Scholar]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Crossa, J.; Huerta-Espino, J.; Lan, C.; Bhavani, S.; Rutkoski, J.E.; Poland, J.A.; Bergstrom, G.C.; et al. Genomic and pedigree-based prediction for leaf, stem, and stripe rust resistance in wheat. Theor. Appl. Genet. 2017, 130, 1415–1430. [Google Scholar] [CrossRef] [PubMed]
- McVey, D.V.; Long, D.L.; Roberts, J.J. Races of Puccinia graminis in the United States during 1997 and 1998. Plant Dis. 2002, 86, 568–572. [Google Scholar] [CrossRef]
- Jin, Y.; Singh, R.P.; Ward, R.W.; Wanyera, R.; Kinyua, M.; Njau, P.; Pretorius, Z.A. Characterization of seedling infection types and adult plant infection responses of monogenic Sr gene lines to race TTKS of Puccinia graminis f. sp. tritici. Plant Dis. 2007, 91, 1096–1099. [Google Scholar] [CrossRef]
- Jin, Y.; Szabo, L.J.; Pretorius, Z.A.; Singh, R.P.; Ward, R.; Fetch, T. Detection of virulence to resistance gene Sr24 within race TTKS of Puccinia graminis f. sp. tritici. Plant Dis. 2008, 92, 923–926. [Google Scholar] [CrossRef]
- Rouse, M.N.; Nirmala, J.; Pretorius, Z.A.; Hiebert, C.W. Characterization of Sr9h, a wheat stem rust resistance allele effective to Ug99. Theor. Appl. Genet. 2014, 127. [Google Scholar] [CrossRef]
- Patpour, M.; Hovmøller, M.S.; Justesen, A.F.; Newcomb, M.; Olivera, P.; Jin, Y.; Szabo, L.J.; Hodson, D.; Shahin, A.A.; Wanyera, R.; Habarurema, I.; et al. Emergence of virulence to SrTmp in the Ug99 race Group of Wheat Stem Rust, Puccinia graminis f. sp. tritici, in Africa. Plant Dis. 2016, 100, 522. [Google Scholar] [CrossRef]
- Olivera, P.D.; Jin, Y.; Rouse, M.; Badebo, A.; Fetch, T.; Singh, R.P.; Yahyaoui, A.M. Races of Puccinia graminis f. sp. tritici with combined virulence to Sr13 and Sr9e in a field stem rust screening nursery in Ethiopia. Plant Dis. 2012, 96, 623–628. [Google Scholar] [CrossRef]
- Olivera, P.D.; Newcomb, M.; Szabo, L.J.; Rouse, M.; Johnson, J.; Gale, S.; Luster, D.G.; Hodson, D.; Cox, J.A.; Burgin, L.; et al. Phenotypic and Genotypic characterization of Race TKTTF of Puccinia graminis f. sp. tritici that caused a wheat stem rust epidemic in southern Ethiopia in 2013–2014. Phytopathology 2015, 105, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Hodson, D.P.; Nazari, K.; Park, R.F.; Hansen, J.; Lassen, P.; Arista, J.; Fetch, T.; Hovmøller, M.S.; Jin, Y.; Pretorius, Z.A.; et al. Putting Ug99 on the map: An update on current and future monitoring 2011. In Proceedings of the BGRI Technical Workshop, Saint Paul, MN, USA, 13 June 2011; pp. 3–13. [Google Scholar]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Huerta-Espino, J.; Kinyua, M.G.; Wanyera, R.; Njau, P.; Ward, R.W. Current status, likely migration and strategies to mitigate the threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. CAB Rev. 2006, 1, 1–13. [Google Scholar] [CrossRef]
- Simons, K.; Abate, Z.; Chao, S.; Zhang, W.; Rouse, M.; Jin, Y.; Elias, E.; Dubcovsky, J. Genetic mapping of stem rust resistance gene Sr13 in tetraploid wheat (Triticum turgidum ssp. durum L.). Theor. Appl. Genet. 2011, 122, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Zhang, J.; Bulli, P.; Abate, Z.; Chao, S.; Cantu, D.; Bossolini, E.; Chen, X.; Pumphrey, M.; Dubcovsky, J. A genome-wide association study of resistance to stripe rust (Puccinia striiformis f. sp. tritici) in a worldwide collection of hexaploid spring wheat (Triticum aestivum L.). G3 Genes Genom. Genet. 2015, 5, 449–465. [Google Scholar] [CrossRef]
- Gupta, P.K.; Pawan, S.; Kulwal, P.L. Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 2005, 57, 461–485. [Google Scholar] [CrossRef]
- Laidò, G.; Marone, D.; Russo, M.A.; Colecchia, S.A.; Mastrangelo, A.M.; De Vita, P.; Papa, R. Linkage disequilibrium and association mapping in tetraploid wheat. PLoS ONE 2014, 9, e95211. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Sanguineti, M.C.; Demontis, A.; El-Ahmed, A.; del Moral, L.G.; Maalouf, F.; Nachit, M.; Nserallah, N.; Ouabbou, H.; Rhouma, S.; et al. Association mapping in durum wheat grown across a broad range of water regimes. J. Exp. Bot. 2011, 62, 409–438. [Google Scholar] [CrossRef]
- Maccaferri, M.; Cane, M.A.; Sanguineti, M.C.; Salvi, S.; Colalongo, M.C.; Massi, A.; Clarke, F.; Knox, R.; Pozniak, C.J.; Clarke, J.M.; et al. A consensus framework map of durum wheat (Triticum durum Desf.) suitable for linkage disequilibrium analysis and genome-wide association mapping. BMC Genom. 2014, 15, 873. [Google Scholar] [CrossRef]
- Somers, D.J.; Banks, T.; DePauw, R.; Fox, S.; Clarke, J.; Pozniak, C.; McCartney, C. Genome-wide linkage disequilibrium analysis in bread wheat and durum wheat. Genome 2007, 50, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Wang, Y.; Chao, S.; Li, T.; Liu, H.; Wang, L.; Zhang, X. The iSelect 9K SNP analysis revealed polyploidization induced revolutionary changes and intense human selection causing strong haplotype blocks in wheat. Sci. Rep. 2017, 7, 41247. [Google Scholar] [CrossRef] [PubMed]
- Ovenden, B.; Milgate, A.; Wade, L.J.; Rebetzke, G.J.; Holland, J.B. Genome-Wide Associations for Water-Soluble Carbohydrate Concentration and Relative Maturity in Wheat Using SNP and DArT Marker Arrays. G3 Genes Genom. Genet. 2017, 7, 2821–2830. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Laidò, G.; Panio, G.; Marone, D.; Russo, M.A.; Ficco, D.B.; Giovanniello, V.; Cattivelli, L.; Steffenson, B.; De Vita, P.; Mastrangelo, A.M. Identification of new resistance loci to African stem rust race TTKSK in tetraploid wheats based on linkage and genome-wide association mapping. Front. Plant Sci. 2015, 6, 1033. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90000 single nucleotide polymorphism array. Plant Biotech. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef]
- Letta, T.; Maccaferri, M.; Badebo, A.; Ammar, K.; Ricci, A.; Crossa, J.; Tuberosa, R. Searching for novel sources of field resistance to Ug99 and Ethiopian stem rust races in durum wheat via association mapping. Theor. Appl. Genet. 2013, 126, 1237–1256. [Google Scholar] [CrossRef]
- Haile, J.K.; Nachit, M.M.; Hammer, K.; Badebo, A.; Roder, M.S. QTL mapping of resistance to race Ug99 of Puccinia graminis f. sp. Tritici in durum wheat (Triticum durum Desf.). Mol. Breed. 2012, 30, 1479–1493. [Google Scholar] [CrossRef]
- Hall, D.; Tegstrom, C.; Ingvarsson, P.K. Using association mapping to dissect the genetic basis of complex traits in plants. Brief. Funct. Genom. 2010, 9, 157–165. [Google Scholar] [CrossRef]
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotech. J. 2015, 13, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, J.; Saini, J.; Newcomb, M.; Olivera, P.; Gale, S.; Klindworth, D.; Elias, E.; Talbert, L.; Chao, S.; Faris, J.; et al. Discovery of a novel stem rust resistance allele in durum wheat that exhibits differential reactions to Ug99 isolates. G3 Genes Genomes Genet. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bhavani, S.; Singh, R.P.; Argillier, O.; Huerta-Espino, J.; Singh, S.; Njau, P.; Brun, S.; Lacam, S.; Desmouceaux, N. Mapping durable adult plant stem rust resistance to the race Ug99 group in six CIMMYT wheats. In Proceedings of the BGRI Technical Workshop, Saint Paul, MN, USA, 13 June 2011; pp. 43–53. [Google Scholar]
- Basnet, B.R.; Singh, S.; Lopez-Vera, E.E.; Huerta-Espino, J.; Bhavani, S.; Jin, Y.; Rouse, M.N.; Singh, R.P. Molecular mapping and validation of SrND643: A new wheat gene for resistance to the stem rust pathogen Ug99 race group. Phytopathology 2015, 105, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Bajgain, P.; Rouse, M.N.; Bhavani, S.; Anderson, J.A. QTL mapping of adult plant resistance to Ug99 stem rust in the spring wheat population RB07/MN06113-8. Mol. Breed. 2015, 35, 170. [Google Scholar] [CrossRef]
- Muleta, K.T.; Rouse, M.N.; Rynearson, S.; Chen, X.; Buta, B.G.; Pumphrey, M.O. Characterization of molecular diversity and genome-wide mapping of loci associated with resistance to stripe rust and stem rust in Ethiopian bread wheat accessions. BMC Plant Biol. 2017, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Tsilo, T.J.; Jin, Y.; Anderson, J.A. Microsatellite markers linked to stem rust resistance allele Sr9a in wheat. Crop Sci. 2007, 47, 2013–2020. [Google Scholar] [CrossRef]
- Marone, D.; Russo, M.A.; Laidò, G.; De Vita, P.; Papa, R.; Blanco, A.; Gadaleta, A.; Rubiales, D.; Mastrangelo, A.M. Genetic basis of qualitative and quantitative resistance to powdery mildew in wheat: From consensus regions to candidate genes. BMC Genom. 2013, 14, 562. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Mazzucotelli, E.; Marone, D.; Crosatti, C.; Michelotti, V.; Valè, G.; Mastrangelo, A.M. Regulation and evolution of NLR genes: A close interconnection for plant immunity. Int. J. Mol. Sci. 2018, 19, E1662. [Google Scholar] [CrossRef]
- Roelfs, A.P.; Martens, J.W. An international system of nomenclature for Puccinia graminis f. sp. tritici. Phytopathology 1988, 78, 526–533. [Google Scholar] [CrossRef]
- Mamo, B.E.; Kevin, P.S.; Brueggeman, R.S.; Steffenson, B.J. Genetic characterization of resistance to wheat stem rust race TTKSK in landrace and wild barley accessions identifies the Rpg4/Rpg5 locus. Phytopathology 2015, 105, 99–109. [Google Scholar] [CrossRef]
- Steffenson, B.J.; Jin, Y.; Brueggeman, R.S.; Kleinhofs, A.; Sun, Y. Resistance to stem rust race TTKSK maps to the rpg4/Rpg5 complex of chromosome 5H of barley. Phytopathology 2009, 99, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Stakman, E.C.; Stewart, P.M.; Loegering, W. Identification of Physiologic Races of Puccinia graminis var. tritici; US Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1962.
- Zhang, D.; Bowden, R.L.; Yu, J.; Carver, B.F.; Bai, G. Association Analysis of Stem Rust Resistance in U.S. Winter Wheat. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Texcoco, Mexico, 1992. [Google Scholar]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P.J. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Royo, C.; Maccaferri, M.; Álvaro, F.; Moragues, M.; Sanguineti, M.C.; Tuberosa, R.; Maalouf, F.; García Del Moral, L.F.; Demontis, A.; Rhouma, S.; et al. Understanding the relationships between genetic and phenotypic structures of a collection of elite durum wheat accessions. Field Crops Res. 2010, 119, 91–105. [Google Scholar] [CrossRef]
- Breseghello, F.; Sorrells, M.E. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics 2006, 172, 1165–1177. [Google Scholar] [CrossRef]
- Cleveland, W.S. Robust locally weighted regression and smoothing scatterplots. J. Am. Stat. Assoc. 1979, 74, 829–836. [Google Scholar] [CrossRef]
- Ardlie, K.G.; Kruglyak, L.; Seielstad, M. Patterns of linkage disequilibrium in the human genome. Nat. Rev. Genet. 2002, 3, 299–309. [Google Scholar] [CrossRef]
- Papa, R.; Bellucci, E.; Rossi, M.; Leonardi, S.; Rau, D.; Gepts, P.; Nanni, L.; Attene, G. Tagging the signatures of domestication in common bean (Phaseolus vulgaris) by means of pooled DNA samples. Ann. Bot. 2007, 100, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Mosig, M.O.; Lipkin, E.; Khutoreskaya, G.; Tchourzyna, E.; Soller, M. Whole genome scan for quantitative trait loci affecting milk protein percentage in Israeli-Holstein cattle, by means of selective milk DNA pooling in a daughter design, using an adjusted false discovery rate criterion. Genetics 2001, 157, 1683–1698. [Google Scholar] [PubMed]
- Benjamini, Y.; Hockberg, Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef]
- Rutkoski, J.; Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Poland, J. Genetic gain from phenotypic and genomic selection for quantitative resistance to stem rust of wheat. Plant Genome 2015, 8, 2. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Race | CV | Mean | Range | Min–Max | Genetic Variance | MSD | H2 (%) |
|---|---|---|---|---|---|---|---|
| TTTTF | 0.48 | 6.65 | 10 | 0–10 | 10.34 | 2.05 | 90 |
| TPMKC | 0.41 | 6.69 | 10 | 0–10 | 7.86 | 1.92 | 89 |
| TRTTF | 0.44 | 6.43 | 10 | 0–10 | 8.16 | 1.73 | 91 |
| JRCQC | 0.23 | 7.82 | 10 | 0–10 | 3.28 | 1.62 | 81 |
| TKTTF | 0.60 | 5.64 | 10 | 0–10 | 11.70 | 1.95 | 92 |
| QTL Region | Interval Zavitan (Mbp) | Number of Annotated Genes | Ratio Annotated Genes/Interval Size | Number of Disease-Related Genes | Ratio Disease-Related/Annotated Genes | Ratio Disease-Related Genes/Mbp |
|---|---|---|---|---|---|---|
| 10 | 17 | 753 | 44.3 | 137 | 0.18 | 8.06 |
| 11 | 3.9 | 62 | 15.9 | 13 | 0.21 | 3.33 |
| 13 | 9.8 | 123 | 12.6 | 8 | 0.07 | 0.82 |
| 14 | 9.5 | 80 | 8.4 | 1 | 0.01 | 0.11 |
| 16 | 4.2 | 258 | 61.4 | 48 | 0.19 | 11.43 |
| 17 | 2.1 | 34 | 16.2 | 1 | 0.03 | 0.48 |
| 18 | 4.8 | 157 | 32.7 | 12 | 0.08 | 2.50 |
| 19 | 5.1 | 70 | 13.7 | 2 | 0.03 | 0.39 |
| 20 | 17 | 77 | 4.5 | 2 | 0.03 | 0.12 |
| 22 | 1.6 | 21 | 13.1 | 4 | 0.19 | 2.50 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saccomanno, A.; Matny, O.; Marone, D.; Laidò, G.; Petruzzino, G.; Mazzucotelli, E.; Desiderio, F.; Blanco, A.; Gadaleta, A.; Pecchioni, N.; et al. Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection. Int. J. Mol. Sci. 2018, 19, 3907. https://doi.org/10.3390/ijms19123907
Saccomanno A, Matny O, Marone D, Laidò G, Petruzzino G, Mazzucotelli E, Desiderio F, Blanco A, Gadaleta A, Pecchioni N, et al. Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection. International Journal of Molecular Sciences. 2018; 19(12):3907. https://doi.org/10.3390/ijms19123907
Chicago/Turabian StyleSaccomanno, Antonietta, Oadi Matny, Daniela Marone, Giovanni Laidò, Giuseppe Petruzzino, Elisabetta Mazzucotelli, Francesca Desiderio, Antonio Blanco, Agata Gadaleta, Nicola Pecchioni, and et al. 2018. "Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection" International Journal of Molecular Sciences 19, no. 12: 3907. https://doi.org/10.3390/ijms19123907
APA StyleSaccomanno, A., Matny, O., Marone, D., Laidò, G., Petruzzino, G., Mazzucotelli, E., Desiderio, F., Blanco, A., Gadaleta, A., Pecchioni, N., De Vita, P., Steffenson, B., & Mastrangelo, A. M. (2018). Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection. International Journal of Molecular Sciences, 19(12), 3907. https://doi.org/10.3390/ijms19123907