Comparative Proteomic Analysis Reveals Elevated Capacity for Photosynthesis in Polyphenol Oxidase Expression-Silenced Clematis terniflora DC. Leaves

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Cloning, Sequence Analysis, and Phylogenetic Tree Analysis of CtPPO
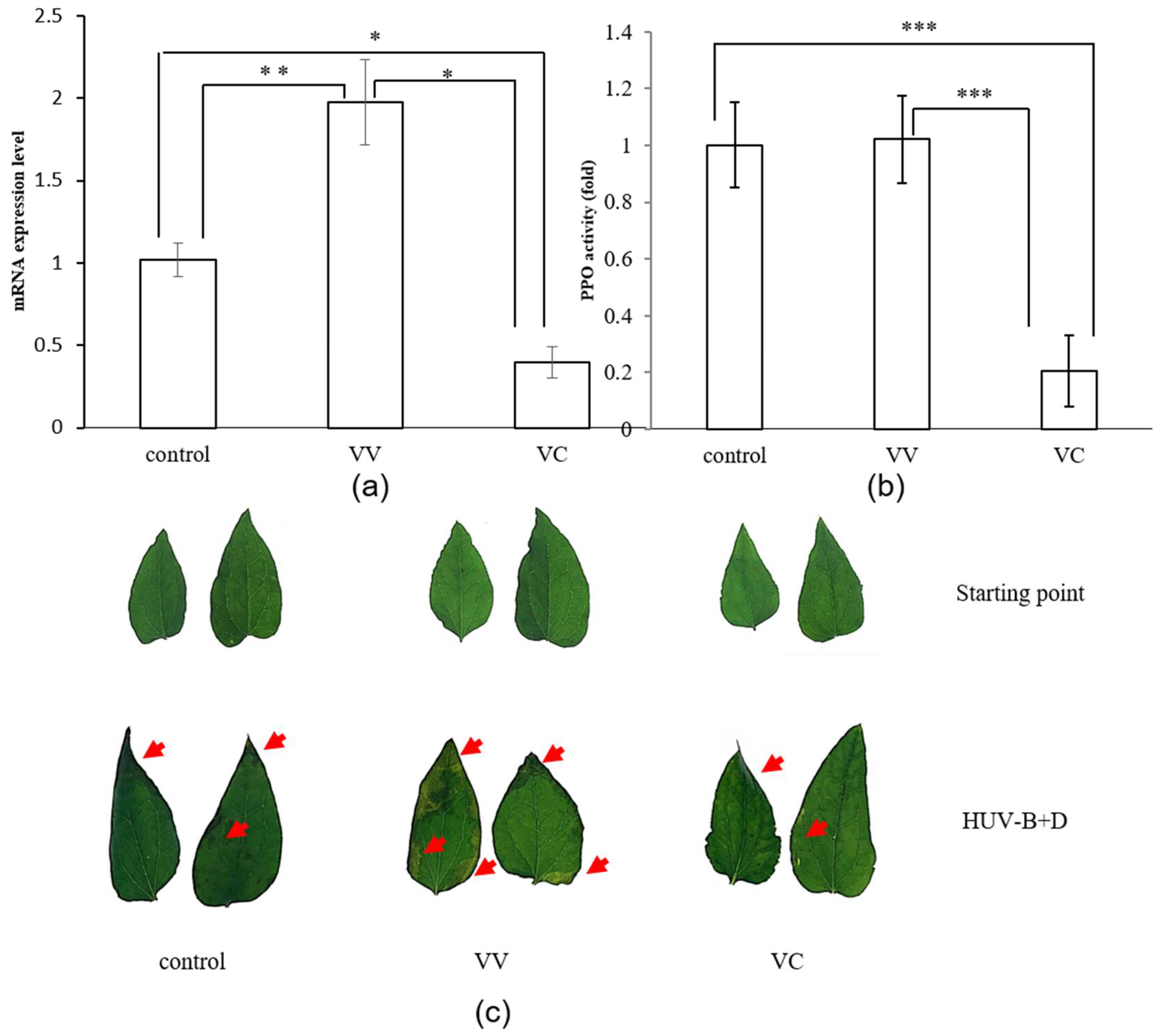
2.2. Virus-Induced Gene Silencing of CtPPO in C. terniflora DC
2.3. Effects of VIGS-CtPPO and VIGS-Vector on Leaf Proteins in C. terniflora DC.
2.4. Effects of High-Level UV-B and Dark Treatment on Proteins in VIGS-CtPPO and VIGS-Vector C. terniflora DC. Leaves
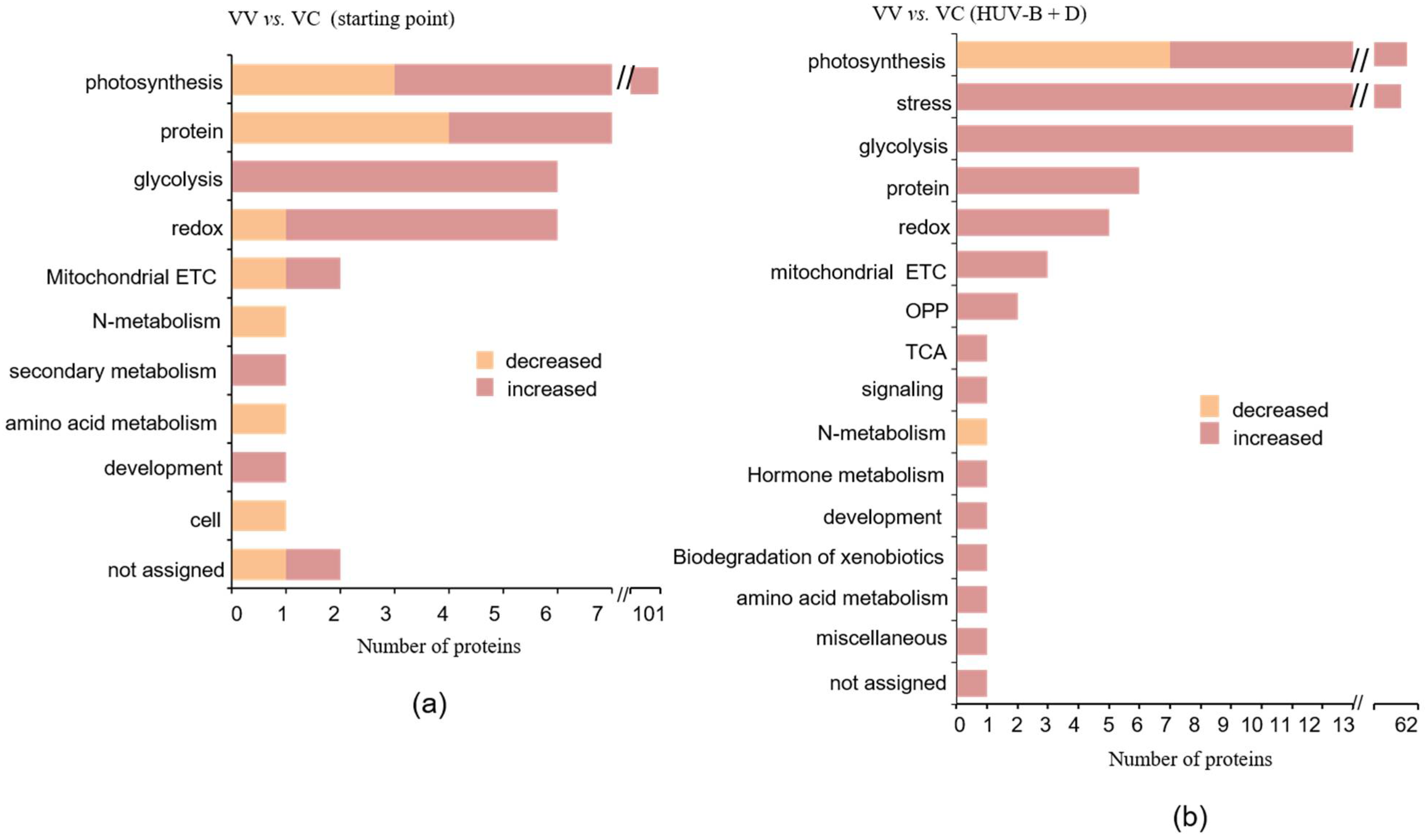
2.5. Integrated Analysis of Proteins in VIGS-CtPPO and VIGS-Vector C. terniflora Differentially Expressed after HUV-B+D
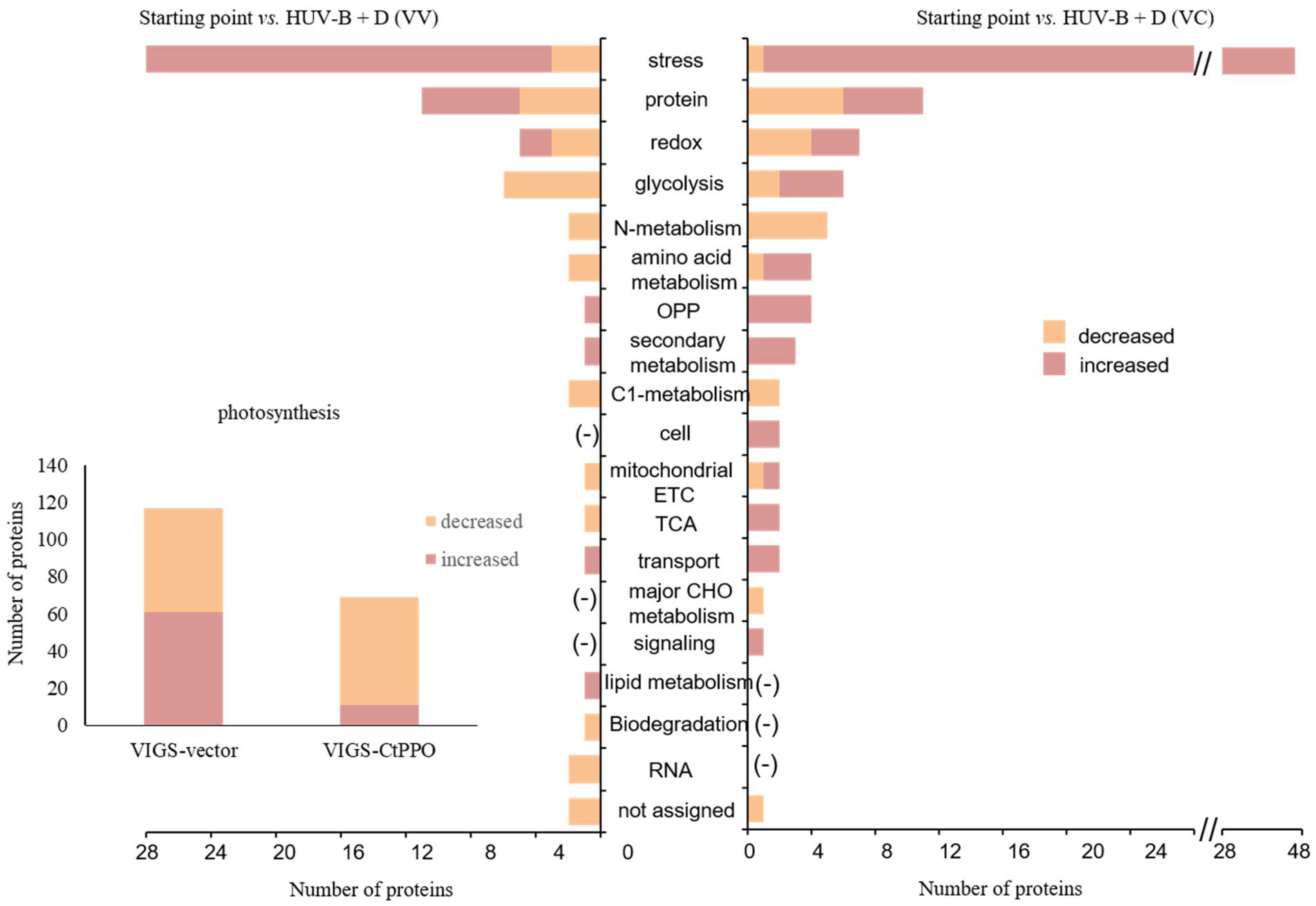
2.6. MapMan Analysis of the VIGS-CtPPO and VIGS-Vector C. terniflora Leaf Proteomic Data at the Starting Point and after HUV-B+D
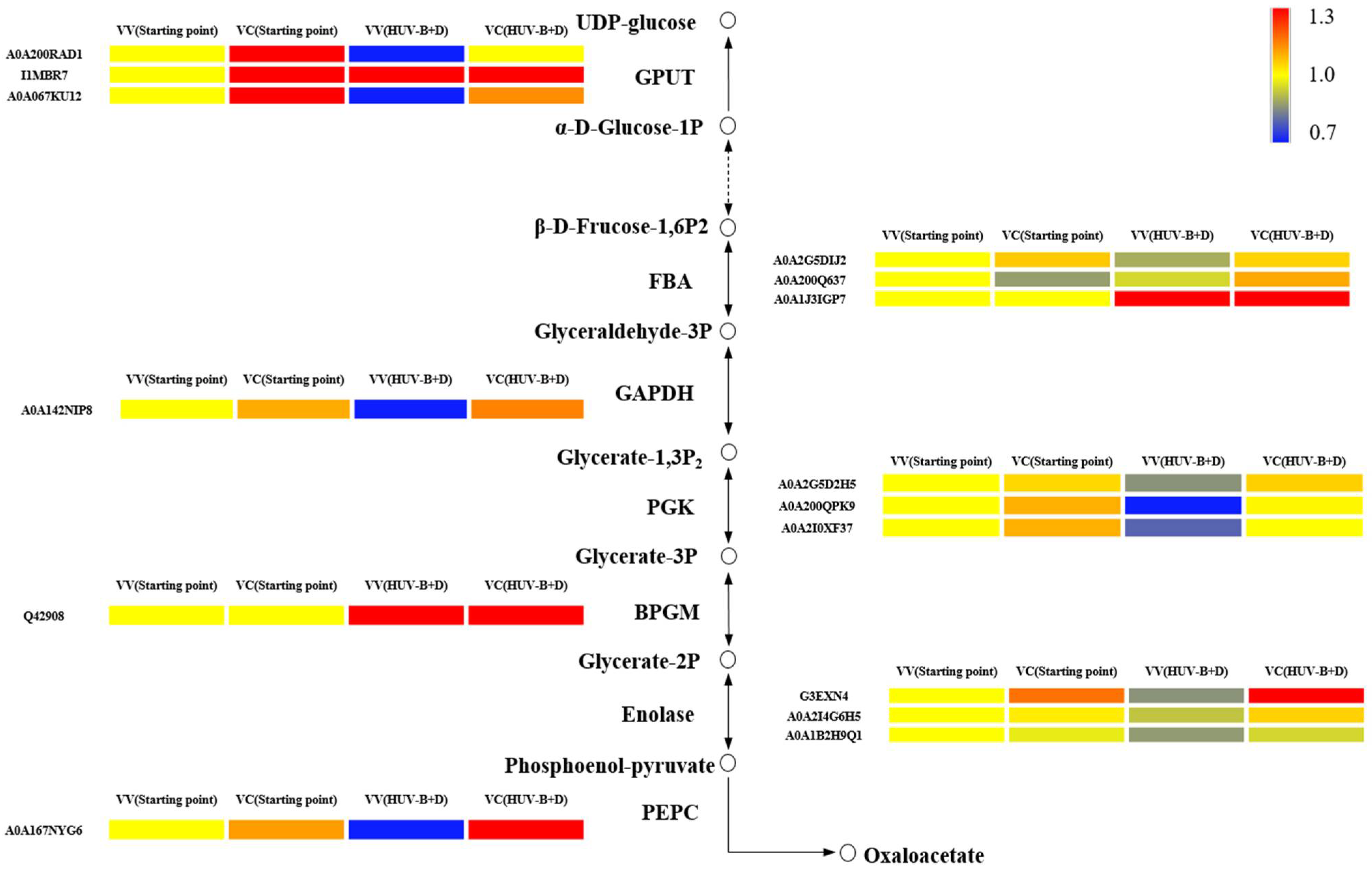
2.7. Expression Profile Analysis of Glycolysis-Related Proteins in C. terniflora DC. Leaves
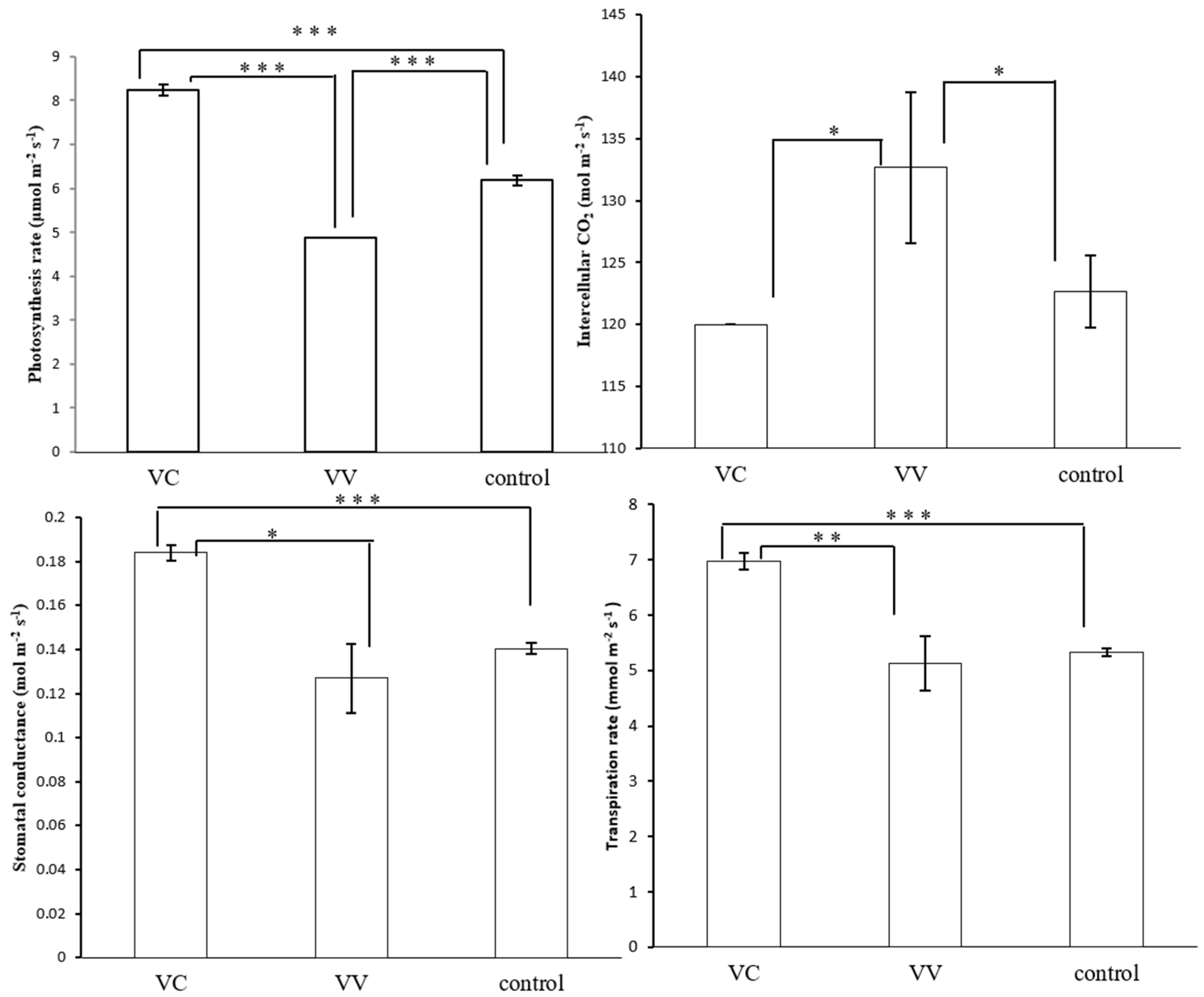
2.8. Effects of VIGS-CtPPO on Photosynthesis Characteristics in C. terniflora DC. Leaves
2.9. Effects of VIGS-CtPPO on ATP Synthase in C. terniflora DC. Leaves
3. Discussion
3.1. Silencing of PPO Promoted the Light Reaction in C. terniflora
3.2. Silencing of PPO Activated the Calvin Cycle in C. terniflora
3.3. Silencing of PPO Enhanced the glycolysis in C. terniflora
3.4. Artificial Interference with PPO Activity Has Potential Applications in Agricultural Production
4. Material and Methods
4.1. Plant Materials and Growth Conditions
4.2. High Level UVB and Dark Treatment
4.3. RNA Extraction and Cloning of CtPPO Gene
4.4. Construction of CtPPO Virus-Derived Vectors
4.5. Virus-Induced Gene Silencing
4.6. Protein Extraction, Enrichment, and Digestion for Proteomics Analysis
4.7. Nano-HPLC-MS/MS Analysis
4.8. Protein Identification Based on Mass Spectrometry Data
4.9. Functional Annotation
4.10. Cluster Analysis of Protein Abundance
4.11. Phylogenetic Analysis
4.12. qRT-PCR
4.13. PPO Enzymatic Activity
4.14. Analysis of Photosynthesis Characteristics
4.15. Analysis of Chlorophyll Content
4.16. Statistical Analysis
4.17. Accession Codes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3-PGA | 3-Phosphoglycerate |
| ABA | Abscisic acid |
| ABC | Ammonium bicarbonate |
| ACN | Acetonitrile |
| ATP | Adenosine triphosphate |
| BPGM | Phosphoglycerate mutase |
| C. terniflora | Clematis terniflora |
| CtPPO | PPO gene in C. terniflora |
| DEPs | Differentially expressed proteins |
| ETC | Electron transport chain |
| FBA | Fructose-1,6-bisphosphate aldolase |
| FBPase | Fructose-1,6-bisphosphatase |
| Fd | Ferredoxin |
| FTR | Fd-TRX reductase |
| GAP | Glyceraldehyde 3-phosphate |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GPI | Glucose-6-phosphate isomerase |
| GPUT | UTP-glucose-1-phosphate uridylytransferase |
| HCD | High energy collisional dissociation |
| HUV-B+D | High level of UV-B and dark |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MES | 2-(N-morpholino) ethanesulfonic acid |
| NTRC | NADPH-dependent TRX reductase |
| ORF | Open reading frame |
| PDS | Phytoene desaturase |
| PEP | Phosphoenolpyruvate |
| PEPC | Phosphoenolpyruvate carboxylase |
| PGK | Phosphoglycerate kinase |
| PPO | Polyphenol oxidase |
| PRK | Phosphoribulokinase |
| PS I | Photosystem I |
| PS II | Photosystem II |
| PS | Photosystem |
| RACE | 5′ Rapid amplification of cDNA ends |
| Rubisco | Ribulose-1,5-bisphosphate carboxylase/oxygenase |
| RuBP | Ribulose 1,5-bisphosphate |
| SBPase | Sedoheptulose-1,7-bisphosphatase |
| TCA | Tricarboxylic acid |
| TFA | Trifluoroacetic acid |
| TK | Transketolase |
| TRV | Tobacco rattle virus |
| TRX | Thioredoxin |
| UTR | Untranslated Region |
| VIGS | Virus-induced gene silencing |
| VC | VIGS-CtPPO |
| VV | VIGS-vector |
References
- Hart, E.H.; Onime, L.A.; Davies, T.E.; Morphew, R.M.; Kingston-Smith, A.H. The effects of PPO activity on the proteome of ingested red clover and implications for improving the nutrition of grazing cattle. J. Proteom. 2016, 141, 67–76. [Google Scholar] [CrossRef]
- Kroll, J.; Rawel, H.M. Reactions of Plant Phenols with Myoglobin: Influence of Chemical Structure of the Phenolic Compounds. J. Food Sci. 2010, 66, 48–58. [Google Scholar] [CrossRef]
- Nakayama, T.; Yonekura-Sakakibara, K.; Sato, T.; Kikuchi, S.; Fukui, Y.; Fukuchi-Mizutani, M.; Ueda, T.; Nakao, M.; Tanaka, Y.; Kusumi, T.; et al. Aureusidin synthase: A polyphenol oxidase homolog responsible for flower coloration. Science 2000, 290, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Gandia-Herrero, F.; Escribano, J.; Garcia-Carmona, F. Characterization of the monophenolase activity of tyrosinase on betaxanthins: The tyramine-betaxanthin/dopamine-betaxanthin pair. Planta 2005, 222, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef]
- Thipyapong, P.; Mahanil, S.; Bhonwong, A.; Attajarusit, J.; Stout, M.J.; Steffens, J.C. Increasing Resistance of Tomato to Lepidopteran Insects by Overexpression of Polyphenol Oxidase. Acta Hortic. 2006, 22, 29–38. [Google Scholar] [CrossRef]
- Jia, H.; Zhao, P.; Wang, B.; Tariq, P.; Zhao, F.; Zhao, M.; Wang, Q.; Yang, T.; Fang, J. Overexpression of Polyphenol Oxidase Gene in Strawberry Fruit Delays the Fungus Infection Process. Plant Mol. Biol. Report. 2016, 34, 592–606. [Google Scholar] [CrossRef]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Antioxidant defences in olive trees during drought stress: Changes in activity of some antioxidant enzymes. Funct. Plant Biol. 2005, 32, 351–364. [Google Scholar] [CrossRef]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef]
- Murniati, A.; Buchari, B.; Gandasasmita, S.; Nurachman, Z.; Nurhanifah, N. Characterization of Polyphenol Oxidase Application as Phenol Removal in Extracts of Rejected White Oyster Mushrooms (Pleurotus ostreatus). Orient. J. Chem. 2018, 34, 1457–1468. [Google Scholar] [CrossRef]
- Sommer, A.; Ne’Eman, E.; Koussevitzky, S.; Hunt, M.D.; Steffens, J.C.; Mayer, A.M.; Harel, E. The Inhibition by Cu2+ of the Import of Polyphenol Oxidase into Chloroplasts. Photosynth. Light Biosph. 1995, 3, 827–830. [Google Scholar]
- Koussevitzky, S.; Ne’Eman, E.; Harel, E. Import of polyphenol oxidase by chloroplasts is enhanced by methyl jasmonate. Planta 2004, 219, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef] [PubMed]
- Velásquez, A.C.; Chakravarthy, S.; Martin, G.B. Virus-induced gene silencing (VIGS) in Nicotiana benthamiana and tomato. J. Vis. Exp. 2009, 28, 1292. [Google Scholar]
- Lee, W.S.; Rudd, J.J.; Kanyuka, K. Virus induced gene silencing (VIGS) for functional analysis of wheat genes involved in Zymoseptoria tritici susceptibility and resistance. Fungal Genet. Biol. 2015, 79, 84–88. [Google Scholar] [CrossRef]
- Zhao, D.; Zhao, J.R.; Huang, X.; Li, N.; Liu, Y.; Huang, Z.J.; Zhang, Z.Y. Functional Analysis of TNBL1 Gene in Wheat Defense Response to Barley yellow dwarf virus Using BSMV-VIGS Technique. Acta Agron. Sin. 2011, 37, 2106–2110. [Google Scholar] [CrossRef]
- Groszyk, J.; Kowalczyk, M.; Yanushevska, Y.; Stochmal, A.; Rakoczy-Trojanowska, M.; Orczyk, W. Identification and VIGS-based characterization of Bx1 ortholog in rye (Secale cereale L.). PLoS ONE 2017, 12, e0171506. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Long, L.; Zhu, L.F.; Rong, Y.M.; Lu, X.H.; Sun, M.Y.; Zhang, L.; Tian, J.K. Proteomic and Virus-induced Gene Silencing (VIGS) Analyses Reveal That Gossypol, Brassinosteroids, and Jasmonic acid Contribute to the Resistance of Cotton to Verticillium dahliae. Mol. Cell. Proteom. 2013, 12, 3690–3703. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.D.; Yuan, X.Y.; Chen, S.; Meng, L.; Fu, D. Role of the tomato TAGL1 gene in regulating fruit metabolites elucidated using RNA sequence and metabolomics analyses. PLoS ONE 2018, 13, e0199083. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.J.; Zeng, L. Elucidating the role of highly homologous Nicotiana benthamiana ubiquitin E2 gene family members in plant immunity through an improved virus-induced gene silencing approach. Plant Methods 2017, 13, 59. [Google Scholar] [CrossRef]
- Chen, R.Z.; Cui, L.; Guo, Y.J.; Rong, Y.M.; Lu, X.H.; Sun, M.Y.; Zhang, L.; Tian, J.K. In vivo study of four preparative extracts of Clematis terniflora, DC. for antinociceptive activity and anti-inflammatory activity in rat model of carrageenan-induced chronic non-bacterial prostatitis. J. Ethnopharmacol. 2011, 134, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Huang, H.T.; Huang, S.Y.; Lin, Z.H.; Shen, C.C.; Tsai, W.J.; Kuo, Y.H. Antioxidant and Anti-Inflammatory Phenolic Glycosides from Clematis tashiroi. J. Nat. Prod. 2015, 78, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Huang, Y.H.; Yan, C.Y.; Wei, X.D.; Hou, J.Q.; Pu, J.X.; Lv, J.X. N-acetylcysteine Ameliorates Prostatitis via miR-141 Regulating Keap1/Nrf2 Signaling. Inflammation 2016, 39, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.X.; Wang, X.; Gao, C.X.; Chen, M.; Guan, Q.J.; Tian, J.K.; Komatsu, S. Proteomic and Metabolomic Analyses of Leaf from Clematis terniflora DC. Exposed to High-Level Ultraviolet-B Irradiation with Dark Treatment. J. Proteome Res. 2015, 15, 2643–2657. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.B.; Yang, B.X.; Zhang, L.; Lu, Y.Z.; Gong, M.H.; Tian, J.K. An in vivo, and in vitro, assessment of the anti-inflammatory, antinociceptive, and immunomodulatory activities of Clematis terniflora, DC. extract, participation of aurantiamide acetate. J. Ethnopharmacol. 2015, 169, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.X.; Yang, B.X.; Zhang, D.D.; Chen, M.; Tian, J.K. Enhanced metabolic process to indole alkaloids in Clematis terniflora, DC. after exposure to high level of UV-B irradiation followed by the dark. BMC Plant Biol. 2016, 16, 231. [Google Scholar] [CrossRef]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef]
- Yang, B.X.; Guan, Q.J.; Tian, J.K.; Komatsu, S. Transcriptomic and proteomic analyses of leaves from Clematis terniflora, DC. under high level of ultraviolet-B irradiation followed by dark treatment. J. Proteom. 2016, 150, 323–340. [Google Scholar] [CrossRef]
- Joliot, P.; Joliot, A. Cyclic electron flow in C3 plants. BBA Bioenerg. 2015, 1757, 362–368. [Google Scholar] [CrossRef]
- Arnon, D.I.; Whatley, F.R.; Allen, M.B. Vitamin K as a cofactor of photosynthetic phosphorylation. BBA Biochim. Biophys. Acta 1955, 16, 607–608. [Google Scholar] [CrossRef]
- Grondelle, R.V.; Zuber, H. The light reactions of photosynthesis. Proc. Natl. Acad. Sci. USA 1971, 68, 2883–2892. [Google Scholar] [CrossRef]
- Karlusich, J.J.P.; Carrillo, N. Evolution of the acceptor side of photosystem I: Ferredoxin, flavodoxin, and ferredoxin-NADP +, oxidoreductase. Photosynth. Res. 2017, 134, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Aliverti, A.; Pandini, V.; Pennati, A.; Rosa, M.; Zanetti, G. Structural and functional diversity of ferredoxin-NADP(+) reductases. Arch. Biochem. Biophys. 2008, 474, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Hisabori, T.; Sunamura, E.I.; Kim, Y.; Konno, H. The Chloroplast ATP Synthase Features the Characteristic Redox Regulation Machinery. Antioxid. Redox Signal. 2013, 19, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Kohzuma, K.; Bosco, C.D.; Meurer, J.; Kramer, D.M. Light- and Metabolism-related Regulation of the Chloroplast ATP Synthase Has Distinct Mechanisms and Functions. J. Biol. Chem. 2013, 288, 13156–13163. [Google Scholar] [CrossRef] [PubMed]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Shu, S.; Chen, L.; Lu, W.; Sun, J.; Guo, S.; Yuan, Y.; Li, J. Effects of exogenous spermidine on photosynthetic capacity and expression of Calvin cycle genes in salt-stressed cucumber seedlings. J. Plant Res. 2014, 127, 763–773. [Google Scholar] [CrossRef]
- Hanke, G.; Mulo, P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ. 2013, 36, 1071–1084. [Google Scholar] [CrossRef]
- Brandes, H.K.; Hartman, F.C.; Lu, T.Y.S.; Larimer, F.W. Efficient Expression of the Gene for Spinach Phosphoribulokinase in Pichia pastoris and Utilization of the Recombinant Enzyme to Explore the Role of Regulatory Cysteinyl Residues by Site-directed Mutagenesis. J. Biol. Chem. 1996, 271, 6490–6496. [Google Scholar] [CrossRef]
- Nikkanen, L.; Rintamäki, E. Thioredoxin-dependent regulatory networks in chloroplasts under fluctuating light conditions. Philos. Trans. R Soc. Lond. 2014, 369, 20130224. [Google Scholar] [CrossRef]
- Thormählen, I.; Meitzel, T.; Groysman, J.; Öchsner, A.B.; von Roepenach-Lahaye, E.; Naranjo, B.; Cejudo, F.J.; Geigenberger, P. Thioredoxin f1 and NADPH-Dependent Thioredoxin Reductase C Have Overlapping Functions in Regulating Photosynthetic Metabolism and Plant Growth in Response to Varying Light Conditions. Plant Physiol. 2015, 169, 1766–1786. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A.; Ne’Eman, E.; Steffens, J.C.; Mayer, A.M.; Harel, E. Import, targeting, and processing of a plant polyphenol oxidase. Plant Physiol. 1994, 105, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Li, G.Q.; Dong, Z.; Zhou, W. Transformation of tobacco plants by Yali PPO-GFP fusion gene and observation of subcellular localization. Am. J. Transl. Res. 2016, 8, 698. [Google Scholar] [PubMed]
- Richter, A.S.; Peter, E.; Rothbart, M.; Schlicke, H.; Toivola, J.; Rintamäki, E.; Grimm, B. Posttranslational influence of NADPH-dependent thioredoxin reductase C on enzymes in tetrapyrrole synthesis. Plant Physiol. 2013, 162, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Lepisto, A.; Kangasjarvi, S.; Luomala, E.M.; Brader, G.; Sipari, N.; Keränen, M.; Keinänen, M.; Rintamäki, E. Chloroplast NADPH-Thioredoxin Reductase Interacts with Photoperiodic Development in Arabidopsis. Plant Physiol. 2009, 149, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Toivola, J.; Rintamäki, E. Crosstalk between chloroplast thioredoxin systems in regulation of photosynthesis. Plant Cell Environ. 2016, 39, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef]
- Rosa-Téllez, S.; Anoman, A.D.; Flores-Tornero, M.; Toujani, W.; Alseek, S.; Femie, A.R.; Nebauer, S.G.; Muñoz-Bertomeu, J.; Segura, J.; Ros, R. Phosphoglycerate Kinases Are Co-Regulated to Adjust Metabolism and to Optimize Growth. Plant Physiol. 2017, 176, 1182–1198. [Google Scholar] [CrossRef]
- Zhao, Z.; Assmann, S.M. The glycolytic enzyme, phosphoglycerate mutase, has critical roles in stomatal movement, vegetative growth, and pollen production in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 5179–5189. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef]
- Bai, J.; Liu, J.; Jiao, W.; Sa, R.; Zhang, N.; Jia, R. Proteomic analysis of salt-responsive proteins in oat roots (Avena sativa L.). J. Sci. Food Agric. 2016, 96, 3867–3875. [Google Scholar] [CrossRef] [PubMed]
- Azoulay-Shemer, T.; Palomares, A.; Bagheri, A.; Israelsson-Nordstrom, M.; Engineer, C.B.; Bargmann, B.O.; Stephan, A.B.; Schroeder, J.I. Guard cell photosynthesis is critical for stomatal turgor production, yet does not directly mediate CO2- and ABA-induced stomatal closing. Plant J. 2015, 83, 567–581. [Google Scholar] [CrossRef]
- Guo, L.; Devaiah, S.P.; Narasimhan, R.; Pan, X.; Zhang, Y.; Zhang, W.; Wang, X. Cytosolic glyceraldehyde-3-phosphate dehydrogenases interact with phospholipase Dδ to transduce hydrogen peroxide signals in the Arabidopsis response to stress. Plant Cell 2012, 24, 2200–2212. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, F.; Raja, N.I.; Razzaq, A.; Komatsu, S. Proteomic and physiological analyses of wheat seeds exposed to copper and iron nanoparticles. BBA Proteins Proteom. 2017, 1865, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Feillet, P.; Autran, J.C.; Icard-Vernière, C. Pasta brownness: An assessment. J. Cereal Sci. 2000, 32, 215–233. [Google Scholar] [CrossRef]
- Mesquita, V.L.V.; Queiroz, C. Enzymatic Browning Biochemistry of Foods; Elsevier Inc.: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Simeone, R.; Pasqualone, A.; Clodoveo, M.L.; Blanco, A. Genetic mapping of polyphenol oxidase in tetraploid wheat. Cell. Mol. Biol. Lett. 2002, 7, 763–769. [Google Scholar] [PubMed]
- Sun, J.Z.; Zhao, J.T.; Liu, D.C.; Yang, W.L.; Luo, G.B.; Zhang, L.Y.; Zhang, X.Q.; Zhang, A.M. Modification to the Test Method of Polyphenol Oxidase(PPO) Activity in Wheat Seeds and Its Usage in Breeding Programs. J. Triticeae Crops 2012, 3, 13. [Google Scholar]
- Long, S.P.; Zhu, X.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Lin, M.T.; Occhialini, A.; Andralojc, P.J.; Devonshire, J.; Hines, K.M.; Parry, M.A.; Hanson, M.R. β-Carboxysomal proteins assemble into highly organized structures in Nicotiana chloroplasts. Plant J. Cell Mol. Biol. 2014, 79, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Constabel, C.P.; Ryan, C.A. A survey of wound- and methyl jasmonate-induced leaf polyphenol oxidase in crop plants. Phytochemistry 1998, 47, 507–511. [Google Scholar] [CrossRef]
- Winters, A.L.; Minchin, F.R.; Michaelson-Yeates, T.P.; Lee, M.R.; Morris, P. Latent and active polyphenol oxidase (PPO) in red clover (Trifolium pratense) and use of a low PPO mutant to study the role of PPO in proteolysis reduction. J. Agric. Food Chem. 2008, 56, 2817–2824. [Google Scholar] [CrossRef] [PubMed]
- Becker, A. Virus-Induced Gene Silencing. Methods Mol. Biol. 2011, 236, 287–294. [Google Scholar]
- Salim, V.; Yu, F.; Altarejos, J.; Luca, V.D. Virus-induced gene silencing identifies Catharanthus roseus 7-deoxyloganic acid-7-hydroxylase, a step in iridoid and monoterpene indole alkaloid biosynthesis. Plant J. 2013, 76, 754–765. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Damasceno, C.M.B.; Saravanan, R.S.; He, Y.; Catalá, C.; Saladié, M.; Rose, J.K. Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat. Protoc. 2006, 1, 769. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.C.; Fischer, J.S.G.; Xu, T.; Yates, J.R.; Barbosa, V.C. PatternLab: From mass spectra to label-free differential shotgun proteomics. Curr. Protoc. Bioinform. 2012, 13, 13–19. [Google Scholar]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; Xu, Q.W.; et al. 2016 update of the PRIDE database and related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [PubMed]
- Usadel, B.; Nagel, A.; Thimm, O.; Redestig, H.; Blaesing, O.E.; Palacios-Rojas, N.; Selbig, J.; Hannemann, J.; Piques, M.C.; Steinhauser, D.; et al. Extension of the visualization tool MapMan to allow statistical analysis of arrays, display of corresponding genes, and comparison with known responses. Plant Physiol. 2005, 138, 1195–1204. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopaedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Hoon, M.J.L.D.; Imoto, S.; Nolan, J.; Miyano, S. Open source clustering software. Bioinformatics 2004, 20, 1453. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.; Berger, S.; Schaller, A.; Stintzi, A. Jasmonate-dependent induction of polyphenol oxidase activity in tomato foliage is important for defense against Spodoptera exiguabut not against Manduca sexta. BMC Plant Biol. 2014, 14, 257. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein ID a | Description | Abundance | FC | p-Value | Annotation b | |
|---|---|---|---|---|---|---|---|
| VV | VC | ||||||
| Increased | |||||||
| 1 | F6HCT7 | Uncharacterized protein | 0.01 | 0.02 | 1.55 | 0.01 | stress |
| 2 | M5W912 | Ferredoxin-NADP reductase | 0.04 | 0.06 | 1.51 | 0.00 | PS |
| 3 | A0A2G5CER3 | Uncharacterized protein (Fragment) | 0.01 | 0.01 | 1.50 | 0.03 | signaling |
| 4 | A0A200RAD1 | UTP-glucose-1-phosphate uridylyltransferase | 0.02 | 0.03 | 1.48 | 0.03 | glycolysis |
| 5 | I6P9I5 | Cytosolic ascorbate peroxidase (Fragment) | 0.03 | 0.04 | 1.47 | 0.02 | redox |
| 6 | J3MAL0 | Ferredoxin-NADP reductase | 0.04 | 0.06 | 1.47 | 0.00 | PS |
| 7 | M4CGU3 | Uncharacterized protein | 0.02 | 0.02 | 1.45 | 0.02 | PS |
| 8 | Q42908 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase | 0.01 | 0.02 | 1.42 | 0.04 | glycolysis |
| 9 | A0A1J3H2G2 | Ubiquitin-NEDD8-like protein RUB1 (Fragment) | 0.27 | 0.38 | 1.40 | 0.01 | protein |
| 10 | A0A2G5F096 | Uncharacterized protein | 0.02 | 0.03 | 1.40 | 0.05 | development |
| 11 | C5YTC0 | Uncharacterized protein | 0.02 | 0.03 | 1.39 | 0.00 | not assigned |
| 12 | A0A067L6G5 | Uncharacterized protein | 0.02 | 0.03 | 1.39 | 0.00 | amino acid metabolism |
| 13 | W9RXI1 | Glycerate dehydrogenase | 0.08 | 0.11 | 1.38 | 0.00 | PS |
| 14 | Q1EP00 | Chlorophyll a-b binding protein, chloroplastic | 0.05 | 0.07 | 1.37 | 0.03 | PS |
| 15 | W9SCQ6 | Ferredoxin-NADP reductase | 0.05 | 0.07 | 1.36 | 0.00 | PS |
| 16 | A0A2G5DAJ3 | Carbonic anhydrase | 0.03 | 0.04 | 1.36 | 0.04 | TCA |
| 17 | W8TP69 | Glycerate dehydrogenase-like protein | 0.04 | 0.06 | 1.36 | 0.02 | PS |
| 18 | A0A0K9P513 | Phosphoglycerate kinase | 0.08 | 0.10 | 1.35 | 0.01 | PS |
| 19 | A0A0K9Q3W1 | 70 kDa heat shock protein | 0.06 | 0.07 | 1.35 | 0.01 | stress |
| 20 | A0A1J6I7J0 | 2-cys peroxiredoxin bas1, chloroplastic | 0.05 | 0.07 | 1.35 | 0.01 | redox |
| 21 | C0PRV0 | Lactoylglutathione lyase | 0.03 | 0.04 | 1.33 | 0.02 | Biodegradation of Xenobiotics |
| 22 | Q19U04 | NADH-dependent hydroxypyruvate reductase (Fragment) | 0.11 | 0.15 | 1.33 | 0.00 | PS |
| 23 | D2XUU3 | Chloroplast managanese stabilizing protein (Fragment) | 0.17 | 0.22 | 1.32 | 0.04 | PS |
| 24 | A0A0A0KBL8 | Uncharacterized protein | 0.03 | 0.04 | 1.32 | 0.00 | PS |
| 25 | A5BVF4 | Uncharacterized protein | 0.16 | 0.21 | 1.32 | 0.02 | PS |
| 26 | A0A200QG47 | Aminotransferase | 0.05 | 0.06 | 1.32 | 0.02 | PS |
| 27 | W1P8B5 | Uncharacterized protein | 0.01 | 0.01 | 1.31 | 0.05 | OPP |
| 28 | A0A0D3B1C7 | Uncharacterized protein | 0.02 | 0.02 | 1.30 | 0.02 | OPP |
| 29 | K7KB09 | Uncharacterized protein | 0.03 | 0.04 | 1.30 | 0.01 | hormone metabolism |
| 30 | K4BW79 | 2-methylene-furan-3-one reductase | 0.06 | 0.07 | 1.30 | 0.00 | misc |
| 31 | A0A2H5NQP8 | Uncharacterized protein | 0.08 | 0.10 | 1.30 | 0.01 | PS |
| 32 | A0A0D2Q3K9 | Uncharacterized protein | 0.04 | 0.05 | 1.30 | 0.00 | stress |
| 33 | A0A200PYZ1 | ATPase | 0.08 | 0.11 | 1.29 | 0.01 | PS |
| 34 | A1BQW9 | Transketolase (Fragment) | 0.05 | 0.06 | 1.29 | 0.00 | PS |
| 35 | A0A251VGE5 | Putative photosystem I PsaA/PsaB | 0.05 | 0.06 | 1.29 | 0.03 | PS |
| 36 | A0A1D8H339 | 2-Cys peroxiredoxin | 0.07 | 0.09 | 1.28 | 0.01 | redox |
| 37 | S8EAM3 | Heat shock protein hsp70 (Fragment) | 0.03 | 0.03 | 1.28 | 0.00 | stress |
| No. | Protein ID a | Description | Abundance | FC | p-Value | Annotation b | |
|---|---|---|---|---|---|---|---|
| VV | VC | ||||||
| Increased | |||||||
| 1 | A0A151U9E4 | Uncharacterized protein | 0.02 | 0.03 | 1.92 | 0.01 | redox |
| 2 | Q8M9K2 | Ribulose-bisphosphate carboxylase (Fragment) | 0.79 | 1.46 | 1.84 | 0.00 | PS |
| 3 | K8ECB3 | Thioredoxin | 0.04 | 0.08 | 1.82 | 0.00 | redox |
| 4 | A0A067KU12 | UTP-glucose-1-phosphate uridylyltransferase | 0.03 | 0.04 | 1.57 | 0.02 | glycolysis |
| 5 | A0A068TPY5 | Uncharacterized protein | 0.03 | 0.05 | 1.53 | 0.01 | PS |
| 6 | A0A2G3DEA2 | 3-oxo-Delta(4,5)-steroid 5-beta-reductase | 0.03 | 0.04 | 1.45 | 0.02 | development |
| 7 | A1X444 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large | 0.27 | 0.39 | 1.44 | 0.00 | PS |
| 8 | A0A1U8LIR6 | 2-methyl-6-phytyl-1,4-hydroquinone methyltransferase | 0.02 | 0.02 | 1.43 | 0.01 | secondary metabolism |
| 9 | A0A200RAD1 | UTP-glucose-1-phosphate uridylyltransferase | 0.03 | 0.04 | 1.42 | 0.04 | glycolysis |
| 10 | A0A2G5DW13 | Uncharacterized protein | 0.04 | 0.06 | 1.42 | 0.01 | redox |
| 11 | A0A067KC46 | Ribulose bisphosphate carboxylase small chain | 0.06 | 0.09 | 1.41 | 0.02 | PS |
| 12 | W9QII5 | Peroxiredoxin Q | 0.03 | 0.04 | 1.41 | 0.02 | redox |
| 13 | A0A061EH79 | Ribulose bisphosphate carboxylase small chain | 0.05 | 0.07 | 1.41 | 0.00 | PS |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Yang, B.; Huang, W.; Wang, T.; Li, Y.; Zhong, Z.; Yang, L.; Li, S.; Tian, J. Comparative Proteomic Analysis Reveals Elevated Capacity for Photosynthesis in Polyphenol Oxidase Expression-Silenced Clematis terniflora DC. Leaves. Int. J. Mol. Sci. 2018, 19, 3897. https://doi.org/10.3390/ijms19123897
Chen X, Yang B, Huang W, Wang T, Li Y, Zhong Z, Yang L, Li S, Tian J. Comparative Proteomic Analysis Reveals Elevated Capacity for Photosynthesis in Polyphenol Oxidase Expression-Silenced Clematis terniflora DC. Leaves. International Journal of Molecular Sciences. 2018; 19(12):3897. https://doi.org/10.3390/ijms19123897
Chicago/Turabian StyleChen, Xi, Bingxian Yang, Wei Huang, Tantan Wang, Yaohan Li, Zhuoheng Zhong, Lin Yang, Shouxin Li, and Jingkui Tian. 2018. "Comparative Proteomic Analysis Reveals Elevated Capacity for Photosynthesis in Polyphenol Oxidase Expression-Silenced Clematis terniflora DC. Leaves" International Journal of Molecular Sciences 19, no. 12: 3897. https://doi.org/10.3390/ijms19123897
APA StyleChen, X., Yang, B., Huang, W., Wang, T., Li, Y., Zhong, Z., Yang, L., Li, S., & Tian, J. (2018). Comparative Proteomic Analysis Reveals Elevated Capacity for Photosynthesis in Polyphenol Oxidase Expression-Silenced Clematis terniflora DC. Leaves. International Journal of Molecular Sciences, 19(12), 3897. https://doi.org/10.3390/ijms19123897