Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy


Abstract
1. Introduction
2. From Tumor-Angiogenesis Factor (TAF) through Vascular Endothelial Growth Factor (VEGF) to Endoglin
3. Angiogenesis in HCC
4. Endoglin—Characteristics of the Molecule
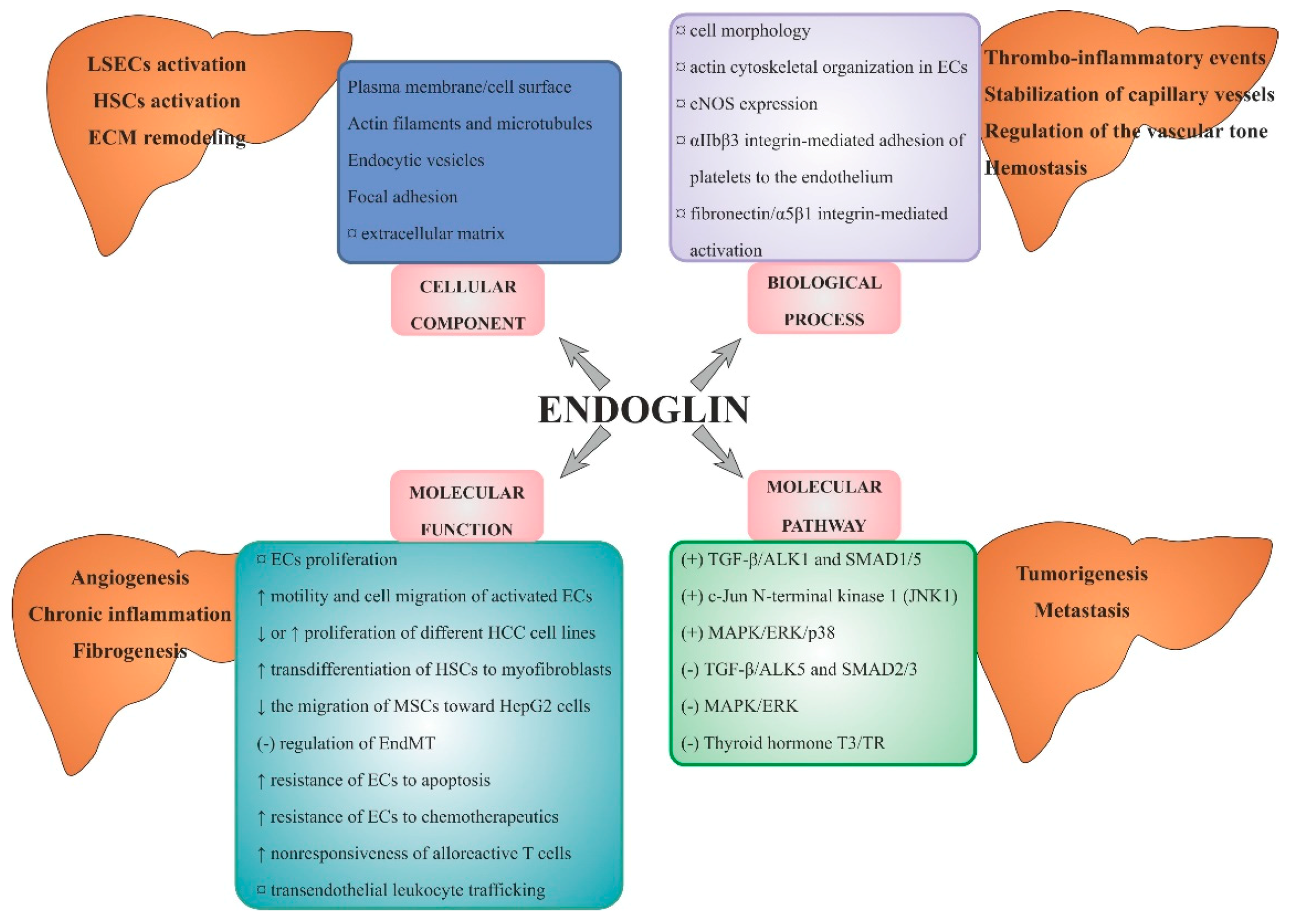
4.1. Endoglin Structure and Best-Known Functions
4.2. Endoglin and Signalling Pathways
4.3. Endoglin and Cellular Effects
4.4. Endoglin and the Most Common Diseases
4.5. Endoglin and Factors Inducing and Inhibiting Its Production
5. Endoglin in the Pathogenesis of HCC—Experimental and Clinical Studies
5.1. Endoglin and Liver Fibrosis
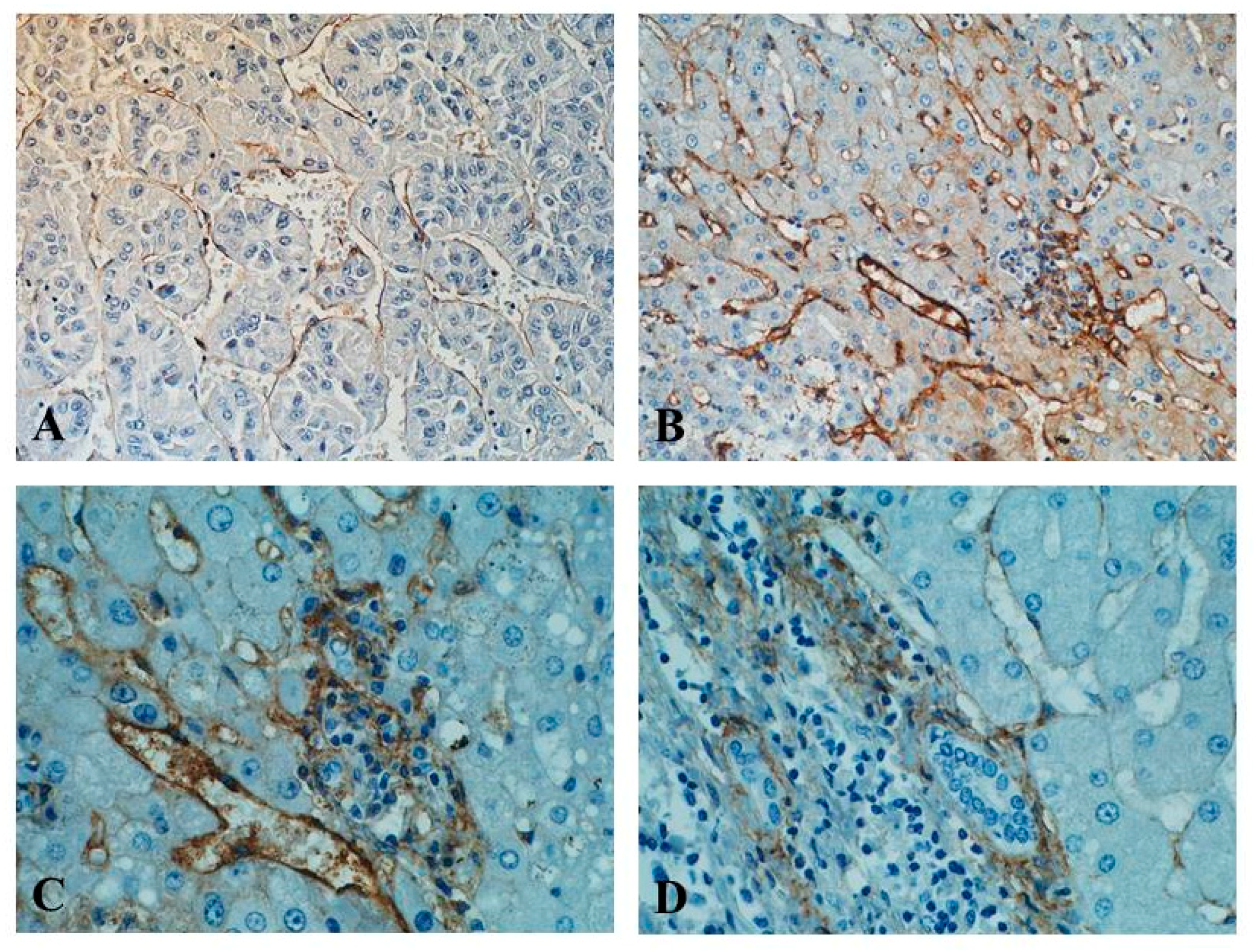
5.2. Endoglin as a Tissue Marker of Tumour Angiogenesis in HCC
5.3. MVD-CD105 as a Prognostic Factor in HCC
5.4. Soluble Endoglin and/or Serum Endoglin mRNA Level in HCC as a Complementary Biomarkers
6. Endoglin in Pathomechanisms of HCC
6.1. Endothelial Progenitor Cells (EPCs) in the Hypoxic Area of HCC
6.2. CD105-Positive Endothelial and Non-Endothelial Cells in HCC
6.3. Endoglin as a Modulator of Tumour Angiogenesis, Tumour Proliferation, Migration, Invasion and Metastasis
7. Endoglin and a New form of Anti-Angiogenic Therapy in HCC
7.1. Animal Studies
7.2. Human Studies
7.3. Nanoparticles as a Potential Way of HCC Therapy
7.4. Summary and Perspectives
8. The Main Headlines of the Review and Conclusions
- Endoglin (CD105) is involved in the process of HCC neoangiogenesis (arterialization and sinusoidal capillarization) under intratumoral hypoxia and acidosis, the main important characteristics of the tumour microenvironment.
- Endoglin is expressed preferentially on activated liver sinusoidal ECs (LSECs) which are characterized by enhanced angiogenic activity, spontaneous motility, greater capacity for migration, and increased resistance to apoptosis in response to TGF-β1.
- In HCC, three patterns of the CD105 expression in ECs are most commonly described (sinusoid-like, branching, and small endothelial sprouts).
- Both CD105-positive LSAECs and CD105-positive non-ECs (normal and cancerous hepatocytes, hepatic stellate cells, human liver-derived stem cells/progenitor cells, human adipose-derived stem cells) play a role in the pathogenesis of HCC.
- The large spectrum of endoglin effects in the liver is cell-type and HCC stage-specific.
- High expression of endoglin in non-tumour tissue suggests that the microenvironment might play an especially important role in the progression of HCC.
- For a reliable assessment of the diagnostic and prognostic role of endoglin in HCC, simultaneous examination of different tumour areas, with a comparison to healthy control, unification of research protocols, and verification of MVD-CD105 counting systems, are required.
- Endoglin (mRNA, protein) tissue expression level examination should be combined with measurement of serum levels of Sol-ENG and other HCC biomarkers and/or determination of the histological stage of the tumour, to successfully predict HCC and effectively apply anti-CD105 therapy.
- Due to the high heterogeneity of pathological changes in HCC and tumour evolution, the effective anti-CD105 therapy in this cancer is not yet available, remaining a goal of the precision medicine development.
9. A Few Key Points
- The main mechanism that stimulates endoglin production in the HCC is the hypoxia of the tumour environment. However, influence of other factors was also proven (e.g., HCV infection, alcohol abuse).
- In HCC-associated angiogenesis in vivo, endoglin expression mainly concerns tumour ECs (TECs), activated ECs in adjacent non-tumour area, and different non-endothelial cells (e.g., hepatic stellate cells).
- In HCC, there is a tight relation between overproduction of endoglin by proliferation tumour ECs (TECs) and creation of sinusoid-like, branching, and small endothelial sprouts. The same relation is not observed between endoglin expression and vasculogenic mimicry.
- Higher endoglin expression in non-tumour area, as compared with tumour tissue, suggests that this location may play a very important role in the progression of HCC.
- The large spectrum of endoglin effects in the liver is cell-type specific, depends on aetiology (higher expression in HCV infection), and is HCC stage-specific (higher expression in HCC with cirrhosis).
- Due to the heterogeneity of HCC and multifactorial aetiology of the cancer, the satisfactory effects of anti-CD105 monotherapy (TRC105) are not yet available, remaining a goal of the precision medicine development.
Author Contributions
Conflicts of Interest
Abbreviations
| aa | Amino Acids |
| ADCC | Antibody-Dependent Cellular Cytotoxicity |
| AFP | Alpha-Fetoprotein |
| ALK1/2/5 | Activin Receptor-Like Kinase 1/2/5 |
| ASF/SF2 | Alternative Splicing Factor/Splicing Factor-2 |
| BMPs | Bone Morphogenetic Proteins |
| CD | Cluster of Differentiation 31, 34, 105, etc. |
| CSS | Cancer-Specific Survival |
| CTGF | Connective Tissue Growth Factor |
| CXCL12 | C-X-C Motif Chemokine 12 or Stromal Cell-Derived Factor 1 (SDF1) |
| DFS | Disease-Free Survival |
| ECs | Endothelial Cells |
| ECM | Extracellular Matrix |
| EGF/R | Epidermal Growth Factor/Receptor |
| EndMT | Endothelial-to-Mesenchymal Transition |
| ENG | Endoglin (CD105) |
| eNOS | Endothelial Nitric Oxide Synthase |
| EPCs/BM-EPCs | Endothelial Progenitor Cells; Bone-Marrow EPCs |
| ERK1/2 | Extracellular Signal-Regulated Kinase 1/2 |
| FGF/bFGF | Fibroblast Growth Factor/Basic FGF |
| HBV/HCV | Hepatitis B/C Virus |
| HCC | Hepatocellular Carcinoma |
| HHT1 | Hemorrhagic Teleangiectasia Type 1 |
| HIF | Hypoxia-Inducible Transcription Factor |
| HRE | Hypoxia-Responsive Element |
| HSCs | Hepatic Stellate Cells |
| HUVECs | Human Umbilical Vein ECs |
| ID1 | DNA Binding Protein 1 |
| IHC | Immunohistochemistry |
| IL | Interleukin |
| INR | International Normalized Ratio of Prothrombin Time of Blood Coagulation |
| KLF6 | Kruppel-Like Factor 6 |
| L-ENG | Long-ENG |
| LOXL2 | Lysyl Oxidase Like-2 |
| LSECs | Liver Sinusoidal Endothelial Cells |
| MAPK | Mitogen-Activated Protein Kinase (Originally Called ERK) |
| MMP2, -14 | Metalloproteinase 2, -14 |
| MVD/IMVD | Microvessels Density/Intratumoral MVD |
| NF-κB | Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells |
| OR | Odds Ratio |
| OS | Overall Survival |
| PD-ECGF | Platelet-Derived Endothelial Cell Growth Factor |
| PDGF/R | Platelet-Derived Growth Factor/Receptor |
| PFS | Progression-Free Survival |
| PIGF | Placental Growth Factor |
| PVTT | Portal Vein Tumour Thrombus |
| RGD domain | Arg-Gly-Asp Domain |
| S-ENG | Short ENG |
| αSMA | Smooth Muscle Actin α |
| SMAD | The Main Signal Transducers for Receptors of TGF-β Family |
| Sol-ENG | Soluble Endoglin |
| TAF | Tumour-Angiogenesis Factor |
| TARE | Transarterial Radioembolization |
| TECs | Tumour Endothelial Cells |
| TGF-α, β | Transforming Growth Factor α, β |
| TIMP1/2 | Tissue Inhibitors of Metalloproteinase 1 and 2 |
| TNF-α | Tumour Necrosis Factor α |
| TNM | Classification of Malignant Tumours (T—tumour; N—lymph nodes; M—metastasis) |
| TRC | Anti-CD105 Monoclonal Antibody |
| TRE | Thyroid Hormone Response Element |
| TSP-1 | Thrombospondin-1 |
| VEGF/R | Vascular Endothelial Growth Factor/Receptor |
| ZRP-1 | Zyxin-Related Protein 1 |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Singh, A.K.; Kumar, R.; Pandey, A.K. Hepatocellular Carcinoma: Causes, Mechanism of Progression and Biomarkers. Curr. Chem. Genom. Transl. Med. 2018, 12, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Poon, R.T.; Ng, I.O.; Lau, C.; Yu, W.C.; Yang, Z.F.; Fan, S.T.; Wong, J. Tumor microvessel density as a predictor of recurrence after resection of hepatocellular carcinoma: A prospective study. J. Clin. Oncol. 2002, 20, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.F.; Poon, R.T. Vascular changes in hepatocellular carcinoma. Anat. Rec. (Hoboken) 2008, 291, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Da Motta Girardi, D.; Correa, T.S.; Crosara Teixeira, M.; Dos Santos Fernandes, G. Hepatocellular Carcinoma: Review of Targeted and Immune Therapies. J. Gastrointest. Cancer 2018, 49, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Medavaram, S.; Zhang, Y. Emerging therapies in advanced hepatocellular carcinoma. Exp. Hematol. Oncol. 2018, 7, 17. [Google Scholar] [CrossRef]
- Peng, S.; Wang, Y.; Peng, H.; Chen, D.; Shen, S.; Peng, B.; Chen, M.; Lencioni, R.; Kuang, M. Autocrine vascular endothelial growth factor signaling promotes cell proliferation and modulates sorafenib treatment efficacy in hepatocellular carcinoma. Hepatology 2014, 60, 1264–1277. [Google Scholar] [CrossRef]
- Ebos, J.M.; Kerbel, R.S. Antiangiogenic therapy: Impact on invasion, disease progression, and metastasis. Nat. Rev. Clin. Oncol. 2011, 8, 210–221. [Google Scholar] [CrossRef]
- Seon, B.K.; Haba, A.; Matsuno, F.; Takahashi, N.; Tsujie, M.; She, X.; Harada, N.; Uneda, S.; Tsujie, T.; Toi, H.; et al. Endoglin-targeted cancer therapy. Curr. Drug Deliv. 2011, 8, 135–143. [Google Scholar] [CrossRef]
- Finnson, K.W.; Philip, A. Endoglin in liver fibrosis. J. Cell Commun. Signal. 2012, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Paschoal, J.P.; Bernardo, V.; Canedo, N.H.; Ribeiro, O.D.; Caroli-Bottino, A.; Pannain, V.L. Microvascular density of regenerative nodule to small hepatocellular carcinoma by automated analysis using CD105 and CD34 immunoexpression. BMC Cancer 2014, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Alsamman, M.; Sterzer, V.; Meurer, S.K.; Sahin, H.; Schaeper, U.; Kuscuoglu, D.; Strnad, P.; Weiskirchen, R.; Trautwein, C.; Scholten, D. Endoglin in human liver disease and murine models of liver fibrosis-A protective factor against liver fibrosis. Liver Int. 2018, 38, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Y.; Lu, W.Q.; Huang, G.W.; Wang, W. Correlation between CD105 expression and postoperative recurrence and metastasis of hepatocellular carcinoma. BMC Cancer 2006, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Pan, Y.; Chen, J.; Sun, X.; Qiu, Y.; Ding, Y. Endoglin (CD105) expression in angiogenesis of primary hepatocellular carcinomas: Analysis using tissue microarrays and comparisons with CD34 and VEGF. Ann. Clin. Lab. Sci. 2007, 37, 39–48. [Google Scholar] [PubMed]
- Li, Y.; Zhai, Z.; Liu, D.; Zhong, X.; Meng, X.; Yang, Q.; Liu, J.; Li, H. CD105 promotes hepatocarcinoma cell invasion and metastasis through VEGF. Tumour Biol. 2015, 36, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, E.; Rizk, M.; Stanzel, S.; Hellerbrand, C.; Lammert, F.; Trautwein, C.; Wasmuth, H.E.; Gressner, A.M. Elevation of endoglin (CD105) concentrations in serum of patients with liver cirrhosis and carcinoma. Eur. J. Gastroenterol. Hepatol. 2007, 19, 755–761. [Google Scholar] [CrossRef]
- Elnemr, D.M.; Abdel-Azeez, H.A.; Labib, H.A.; Abo-Taleb, F.M. Clinical relevance of serum endoglin level in Egyptian hepatocellular carcinoma patients. Clin. Lab. 2012, 58, 1023–1028. [Google Scholar]
- Teama, S.; Fawzy, A.; Teama, S.; Helal, A.; Drwish, A.D.; Elbaz, T.; Desouky, E. Increased Serum Endoglin and Transforming Growth Factor β1 mRNA Expression and Risk of Hepatocellular Carcinoma in Cirrhotic Egyptian Patients. Asian Pac. J. Cancer Prev. 2016, 17, 2429–2434. [Google Scholar]
- Yu, D.; Zhuang, L.; Sun, X.; Chen, J.; Yao, Y.; Meng, K.; Ding, Y. Particular distribution and expression pattern of endoglin (CD105) in the liver of patients with hepatocellular carcinoma. BMC Cancer 2007, 7, 122. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.H.; Guo, P.; Yan, L.N.; He, D. Tumor microvascular density detected by anti-CD105 and anti-CD34 in hepatocellular carcinoma patients and its predictive value of tumor recurrence after liver transplantation. Sichuan Da Xue Xue Bao Yi Xue Ban 2010, 41, 818–821. [Google Scholar] [PubMed]
- Ribeiro, O.D.; Canedo, N.H.; Pannain, V.L. Immunohistochemical angiogenic biomarkers in hepatocellular carcinoma and cirrhosis: Correlation with pathological features. Clinics (Sao Paulo) 2016, 71, 639–643. [Google Scholar] [CrossRef]
- Bernabeu, C.; Lopez-Novoa, J.M.; Quintanilla, M. The emerging role of TGF-β superfamily coreceptors in cancer. Biochim. Biophys. Acta 2009, 1792, 954–973. [Google Scholar] [CrossRef] [PubMed]
- López-Novoa, J.M.; Bernabeu, C. The physiological role of endoglin in the cardiovascular system. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H959–H974. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y. Tumor angiogenesis and anti-angiogenic therapy. Keio J. Med. 2012, 61, 47–56. [Google Scholar] [CrossRef]
- Döme, B.; Hendrix, M.J.; Paku, S.; Tóvári, J.; Tímár, J. Alternative vascularization mechanisms in cancer: Pathology and therapeutic implications. Am. J. Pathol. 2007, 170, 1–15. [Google Scholar] [CrossRef]
- Folkman, J.; Merler, E.; Abernathy, C.; Williams, G. Isolation of a tumor factor responsible for angiogenesis. J. Exp. Med. 1971, 133, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Brem, H.; Folkman, J. Inhibition of tumor angiogenesis mediated by cartilage. J. Exp. Med. 1975, 141, 427–439. [Google Scholar] [CrossRef]
- Senger, D.R.; Galli, S.J.; Dvorak, A.M.; Perruzzi, C.A.; Harvey, V.S.; Dvorak, H.F. Tumor cells secrete a vascular permeability factor that promotes accumulation of ascites fluid. Science 1983, 219, 983–985. [Google Scholar] [CrossRef]
- Yu, D.; Sun, X.; Qiu, Y.; Zhou, J.; Wu, Y.; Zhuang, L.; Chen, J.; Ding, Y. Identification and clinical significance of mobilized endothelial progenitor cells in tumor vasculogenesis of hepatocellular carcinoma. Clin. Cancer Res. 2007, 13, 3814–3824. [Google Scholar] [CrossRef]
- Yu, D.C.; Chen, J.; Ding, Y.T. Hypoxic and highly angiogenic non-tumor tissues surrounding hepatocellular carcinoma: The ‘niche’ of endothelial progenitor cells. Int. J. Mol. Sci. 2010, 11, 2901–2909. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, X.; Zhu, D.; Liu, T.; Liang, X.; Liu, F.; Zhang, Y.; Dong, X.; Sun, B. HIF-1α promoted vasculogenic mimicry formation in hepatocellular carcinoma through LOXL2 up-regulation in hypoxic tumor microenvironment. J. Exp. Clin. Cancer Res. 2017, 36, 60. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Dor, Y.; Herbert, J.M.; Fukumura, D.; Brusselmans, K.; Dewerchin, M.; Neeman, M.; Bono, F.; Abramovitch, R.; Maxwell, P.; et al. Role of HIF-1alpha in hypoxia-mediated apoptosis, cell proliferation and tumour angiogenesis. Nature 1998, 394, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Shahneh, F.Z.; Baradaran, B.; Zamani, F.; Aghebati-Maleki, L. Tumor angiogenesis and anti-angiogenic therapies. Hum. Antibodies 2013, 22, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Hillen, F.; Griffioen, A.W. Tumour vascularization: Sprouting angiogenesis and beyond. Cancer Metastasis Rev. 2007, 26, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.K.; Srinivasan, R.; Chawla, Y.K.; Chakraborti, A. Vascular endothelial growth factor: Evidence for autocrine signaling in hepatocellular carcinoma cell lines affecting invasion. Indian J. Cancer 2016, 53, 542–547. [Google Scholar] [CrossRef]
- Fukumura, D.; Xavier, R.; Sugiura, T.; Chen, Y.; Park, E.C.; Lu, N.; Selig, M.; Nielsen, G.; Taksir, T.; Jain, R.K.; Seed, B. Tumor induction of VEGF promoter activity in stromal cells. Cell 1998, 94, 715–725. [Google Scholar] [CrossRef]
- Gougos, A.; Letarte, M. Identification of a human endothelial cell antigen with monoclonal antibody 44G4 produced against a pre-B leukemic cell line. J. Immunol. 1988, 141, 1925–1933. [Google Scholar]
- Jonker, L.; Arthur, H.M. Endoglin expression in early development is associated with vasculogenesis and angiogenesis. Mech. Dev. 2002, 110, 193–196. [Google Scholar] [CrossRef]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A marker of tumor vasculature and potential target for therapy. Clin. Cancer. Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Nassiri, F.; Cusimano, M.D.; Scheithauer, B.W.; Rotondo, F.; Fazio, A.; Yousef, G.M.; Syro, L.V.; Kovacs, K.; Lloyd, R.V. Endoglin (CD105): A review of its role in angiogenesis and tumor diagnosis, progression and therapy. Anticancer Res. 2011, 31, 2283–2290. [Google Scholar] [PubMed]
- Abel, T.; El Filali, E.; Waern, J.; Schneider, I.C.; Yuan, Q.; Münch, R.C.; Hick, M.; Warnecke, G.; Madrahimov, N.; Kontermann, R.E.; et al. Specific gene delivery to liver sinusoidal and artery endothelial cells. Blood 2013, 122, 2030–2038. [Google Scholar] [CrossRef] [PubMed]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.E. Liver sinusoidal endothelial cells: Physiology and role in liver diseases. J. Hepatol. 2017, 66, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Minhajat, R.; Mori, D.; Yamasaki, F.; Sugita, Y.; Satoh, T.; Tokunaga, O. Organ-specific endoglin (CD105) expression in the angiogenesis of human cancers. Pathol. Int. 2006, 56, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Quackenbush, E.J.; Letarte, M. Identification of several cell surface proteins of non-T, non-B acute lymphoblastic leukemia by using monoclonal antibodies. J. Immunol. 1985, 134, 1276–1285. [Google Scholar] [PubMed]
- Rokhlin, O.W.; Cohen, M.B.; Kubagawa, H.; Letarte, M.; Cooper, M.D. Differential expression of endoglin on fetal and adult hematopoietic cells in human bone marrow. J. Immunol. 1995, 154, 4456–4465. [Google Scholar] [PubMed]
- Liu, Q.; Zhang, R.Z.; Li, D.; Cheng, S.; Yang, Y.H.; Tian, T.; Pan, X.R. Muse Cells, a New Type of Pluripotent Stem Cell Derived from Human Fibroblasts. Cell Reprogram. 2016, 18, 67–77. [Google Scholar] [CrossRef]
- El-Kehdy, H.; Sargiacomo, C.; Fayyad-Kazan, M.; Fayyad-Kazan, H.; Lombard, C.; Lagneaux, L.; Sokal, E.; Najar, M.; Najimi, M. Immunoprofiling of Adult-Derived Human Liver Stem/Progenitor Cells: Impact of Hepatogenic Differentiation and Inflammation. Stem Cells Int. 2017, 2017, 2679518. [Google Scholar] [CrossRef]
- Lin, Y.H.; Huang, Y.H.; Wu, M.H.; Wu, S.M.; Chi, H.C.; Liao, C.J.; Chen, C.Y.; Tseng, Y.H.; Tsai, C.Y.; Tsai, M.M.; et al. Thyroid hormone suppresses cell proliferation through endoglin-mediated promotion of p21 stability. Oncogene 2013, 32, 3904–3914. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.J.; Jang, I.K.; Kim, H.E.; Lee, D.H.; Park, J.K.; Lee, S.K.; Yoon, H.H. In vitro differentiation of human liver-derived stem cells with mesenchymal characteristics into immature hepatocyte-like cells. Transplant. Proc. 2014, 46, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Deng, J.; Jiang, Q.; Wang, Y.; Zhang, Y.; Yao, Y.; Cheng, F.; Chen, X.; Xu, F.; Huang, M.; et al. Rapid generation of functional hepatocyte-like cells from human adipose-derived stem cells. Stem Cell. Res. Ther. 2016, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, P.S.; Bose, B. Hepatic perivascular mesenchymal stem cells with myogenic properties. J. Tissue Eng. Regen. Med. 2018, 12, e1297–e1310. [Google Scholar] [CrossRef]
- Tanigawa, N.; Lu, C.; Mitsui, T.; Miura, S. Quantitation of sinusoid-like vessels in hepatocellular carcinoma: Its clinical and prognostic significance. Hepatology 1997, 26, 1216–1223. [Google Scholar] [PubMed]
- Muto, J.; Shirabe, K.; Sugimachi, K.; Maehara, Y. Review of angiogenesis in hepatocellular carcinoma. Hepatol. Res. 2015, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.N.; Kim, Y.B.; Yang, K.M.; Park, C. Increased expression of vascular endothelial growth factor and angiogenesis in the early stage of multistep hepatocarcinogenesis. Arch. Pathol. Lab. Med. 2000, 124, 1061–1065. [Google Scholar] [PubMed]
- Sanz-Cameno, P.; Trapero-Marugán, M.; Chaparro, M.; Jones, E.A.; Moreno-Otero, R. Angiogenesis: From chronic liver inflammation to hepatocellular carcinoma. J. Oncol. 2010, 2010, 272170. [Google Scholar] [CrossRef] [PubMed]
- Segatelli, V.; de Oliveira, E.C.; Boin, I.F.; Ataide, E.C.; Escanhoela, C.A. Evaluation and comparison of microvessel density using the markers CD34 and CD105 in regenerative nodules, dysplastic nodules and hepatocellular carcinoma. Hepatol. Int. 2014, 8, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.C.; Brasil, I.R.C.; Torres, A.F.C.; Tavora, F. The Evaluation of Angiogenesis Markers in Hepatocellular Carcinoma and Precursor Lesions in Liver Explants from a Single Institution. Appl. Immunohistochem. Mol. Morphol. 2018, 26, 330–336. [Google Scholar] [CrossRef]
- Pang, R.; Poon, R.T. Angiogenesis and antiangiogenic therapy in hepatocellular carcinoma. Cancer Lett. 2006, 242, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ahmad, M.K.; Waseem, M.; Pandey, A.K. Drug targets for cancer treatment: An overview. Med. Chem. 2015, 5, 115–123. [Google Scholar] [CrossRef]
- Zhu, A.X.; Duda, D.G.; Sahani, D.V.; Jain, R.K. HCC and angiogenesis: Possible targets and future directions. Nat. Rev. Clin. Oncol. 2011, 8, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, S.; Zhang, D.; Du, J.; Guo, H.; Zhao, X.; Zhang, W.; Hao, X. Vasculogenic mimicry is associated with high tumor grade, invasion and metastasis, and short survival in patients with hepatocellular carcinoma. Oncol. Rep. 2006, 16, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Diaz, L.A.; Polyak, K.; Meszler, L.; Romans, K.; Guinan, E.C.; Antin, J.H.; Myerson, D.; Hamilton, S.R.; Vogelstein, B.; et al. Contribution of bone marrow-derived endothelial cells to human tumor vasculature. Nat. Med. 2005, 11, 261–262. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Pang, R.W.; Lau, C.; Sun, C.K.; Yu, W.C.; Fan, S.T.; Poon, R.T. Significance of circulating endothelial progenitor cells in hepatocellular carcinoma. Hepatology 2006, 44, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Shao, Q.; Sun, X.; Deng, Z.; Yuan, X.; Yu, D.; Zhou, X.; Ding, Y. The mobilization, recruitment and contribution of bone marrow-derived endothelial progenitor cells to the tumor neovascularization occur at an early stage and throughout the entire process of hepatocellular carcinoma growth. Oncol. Rep. 2012, 28, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhou, H.; Luo, J.; Cui, Y.; Li, H.; Zhang, W.; Fang, F.; Li, Q.; Zhang, T. Different but synergistic effects of bone marrow-derived VEGFR2+ and VEGFR2-CD45+ cells during hepatocellular carcinoma progression. Oncol. Lett. 2017, 13, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Gougos, A.; Letarte, M. Primary structure of endoglin, an RGD-containing glycoprotein of human endothelial cells. J. Biol. Chem. 1990, 265, 8361–8364. [Google Scholar] [PubMed]
- Maring, J.A.; Trojanowska, M.; Ten Dijke, P. Role of endoglin in fibrosis and scleroderma. Int. Rev. Cell Mol. Biol. 2012, 297, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.C.; Sasaki, R.; Meyer, K.; Ray, R. Hepatitis C Virus Core Protein Modulates Endoglin (CD105) Signaling Pathway for Liver Pathogenesis. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Cheifetz, S.; Bellón, T.; Calés, C.; Vera, S.; Bernabeu, C.; Massagué, J.; Letarte, M. Endoglin is a component of the transforming growth factor-β receptor system in human endothelial cells. J. Biol. Chem. 1992, 267, 19027–19030. [Google Scholar] [PubMed]
- Fonsatti, E.; Del Vecchio, L.; Altomonte, M.; Sigalotti, L.; Nicotra, M.R.; Coral, S.; Natali, P.G.; Maio, M. Endoglin: An accessory component of the TGF-β-binding receptor-complex with diagnostic, prognostic, and bioimmunotherapeutic potential in human malignancies. J. Cell Physiol. 2001, 188, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lebrin, F.; Deckers, M.; Bertolino, P.; Ten Dijke, P. TGF-β receptor function in the endothelium. Cardiovasc. Res. 2005, 65, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Ríus, C.; Smith, J.D.; Almendro, N.; Langa, C.; Botella, L.M.; Marchuk, D.A.; Vary, C.P.; Bernabéu, C. Cloning of the promoter region of human endoglin, the target gene for hereditary hemorrhagic telangiectasia type 1. Blood 1998, 92, 4677–4690. [Google Scholar] [PubMed]
- Bellón, T.; Corbí, A.; Lastres, P.; Calés, C.; Cebrián, M.; Vera, S.; Cheifetz, S.; Massague, J.; Letarte, M.; Bernabéu, C. Identification and expression of two forms of the human transforming growth factor-β-binding protein endoglin with distinct cytoplasmic regions. Eur. J. Immunol. 1993, 23, 2340–2345. [Google Scholar] [CrossRef] [PubMed]
- Blanco, F.J.; Bernabeu, C. Alternative splicing factor or splicing factor-2 plays a key role in intron retention of the endoglin gene during endothelial senescence. Aging Cell 2011, 10, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gómez, E.; Eleno, N.; López-Novoa, J.M.; Ramirez, J.R.; Velasco, B.; Letarte, M.; Bernabéu, C.; Quintanilla, M. Characterization of murine S-endoglin isoform and its effects on tumor development. Oncogene 2005, 24, 4450–4461. [Google Scholar] [CrossRef]
- Meurer, S.K.; Tihaa, L.; Borkham-Kamphorst, E.; Weiskirchen, R. Expression and functional analysis of endoglin in isolated liver cells and its involvement in fibrogenic Smad signalling. Cell Signal. 2011, 23, 683–699. [Google Scholar] [CrossRef]
- Velasco, S.; Alvarez-Muñoz, P.; Pericacho, M.; Dijke, P.T.; Bernabéu, C.; López-Novoa, J.M.; Rodríguez-Barbero, A. L- and S-endoglin differentially modulate TGFβ1 signaling mediated by ALK1 and ALK5 in L6E9 myoblasts. J. Cell Sci. 2008, 121, 913–919. [Google Scholar] [CrossRef]
- Gougos, A.; St Jacques, S.; Greaves, A.; O’Connell, P.J.; d’Apice, A.J.; Bühring, H.J.; Bernabeu, C.; van Mourik, J.A.; Letarte, M. Identification of distinct epitopes of endoglin, an RGD-containing glycoprotein of endothelial cells, leukemic cells, and syncytiotrophoblasts. Int. Immunol. 1992, 4, 83–92. [Google Scholar] [CrossRef]
- Rossi, E.; Sanz-Rodriguez, F.; Eleno, N.; Düwell, A.; Blanco, F.J.; Langa, C.; Botella, L.M.; Cabañas, C.; Lopez-Novoa, J.M.; Bernabeu, C. Endothelial endoglin is involved in inflammation: Role in leukocyte adhesion and transmigration. Blood 2013, 121, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gómez, E.; Del Castillo, G.; Juan Francisco, S.; López-Novoa, J.M.; Bernabéu, C.; Quintanilla, M. The role of the TGF-β coreceptor endoglin in cancer. Sci. World J. 2010, 10, 2367–2384. [Google Scholar] [CrossRef] [PubMed]
- Hawinkels, L.J.; Kuiper, P.; Wiercinska, E.; Verspaget, H.W.; Liu, Z.; Pardali, E.; Sier, C.F.; ten Dijke, P. Matrix metalloproteinase-14 (MT1-MMP)-mediated endoglin shedding inhibits tumor angiogenesis. Cancer Res. 2010, 70, 4141–4150. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Vara, E.; Blanco, F.J.; Roqué, M.; Friedman, S.L.; Suzuki, T.; Botella, L.M.; Bernabeu, C. Transcription factor KLF6 upregulates expression of metalloprotease MMP14 and subsequent release of soluble endoglin during vascular injury. Angiogenesis 2016, 19, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Esteo, M.; Sanchez-Elsner, T.; Letamendia, A.; Bernabeu, C. Extracellular and cytoplasmic domains of endoglin interact with the transforming growth factor-β receptors I and II. J. Biol. Chem. 2002, 277, 29197–29209. [Google Scholar] [CrossRef] [PubMed]
- Blanco, F.J.; Santibanez, J.F.; Guerrero-Esteo, M.; Langa, C.; Vary, C.P.; Bernabeu, C. Interaction and functional interplay between endoglin and ALK-1, two components of the endothelial transforming growth factor-β receptor complex. J. Cell Physiol. 2005, 204, 574–584. [Google Scholar] [CrossRef]
- Pomeraniec, L.; Hector-Greene, M.; Ehrlich, M.; Blobe, G.C.; Henis, Y.I. Regulation of TGF-β receptor hetero-oligomerization and signaling by endoglin. Mol. Biol. Cell. 2015, 26, 3117–3127. [Google Scholar] [CrossRef]
- Lebrin, F.; Goumans, M.J.; Jonker, L.; Carvalho, R.L.; Valdimarsdottir, G.; Thorikay, M.; Mummery, C.; Arthur, H.M.; ten Dijke, P. Endoglin promotes endothelial cell proliferation and TGF-β/ALK1 signal transduction. EMBO J. 2004, 23, 4018–4028. [Google Scholar] [CrossRef]
- Kapur, N.K.; Morine, K.J.; Letarte, M. Endoglin: A critical mediator of cardiovascular health. Vasc. Health Risk Manag. 2013, 9, 195–206. [Google Scholar] [CrossRef]
- Rodríguez-Barbero, A.; Obreo, J.; Alvarez-Munoz, P.; Pandiella, A.; Bernabéu, C.; López-Novoa, J.M. Endoglin modulation of TGF-β1-induced collagen synthesis is dependent on ERK1/2 MAPK activation. Cell Physiol. Biochem. 2006, 18, 135–142. [Google Scholar]
- Lee, N.Y.; Blobe, G.C. The interaction of endoglin with β-arrestin2 regulates transforming growth factor-β-mediated ERK activation and migration in endothelial cells. J. Biol. Chem. 2007, 282, 21507–21517. [Google Scholar] [CrossRef]
- Scherner, O.; Meurer, S.K.; Tihaa, L.; Gressner, A.M.; Weiskirchen, R. Endoglin differentially modulates antagonistic transforming growth factor-β1 and BMP-7 signaling. J. Biol. Chem. 2007, 282, 13934–13943. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Mythreye, K.; Golzio, C.; Katsanis, N.; Blobe, G.C. Endoglin mediates fibronectin/α5β1 integrin and TGF-β pathway crosstalk in endothelial cells. EMBO J. 2012, 31, 3885–3900. [Google Scholar] [CrossRef]
- Rossi, E.; Pericacho, M.; Bachelot-Loza, C.; Pidard, D.; Gaussem, P.; Poirault-Chassac, S.; Blanco, F.J.; Langa, C.; González-Manchón, C.; Novoa, J.M.L.; et al. Human endoglin as a potential new partner involved in platelet-endothelium interactions. Cell Mol. Life Sci. 2018, 75, 1269–1284. [Google Scholar] [CrossRef] [PubMed]
- Onoe, T.; Ohdan, H.; Tokita, D.; Hara, H.; Tanaka, Y.; Ishiyama, K.; Asahara, T. Liver sinusoidal endothelial cells have a capacity for inducing nonresponsiveness of T cells across major histocompatibility complex barriers. Transpl. Int. 2005, 18, 206–214. [Google Scholar] [CrossRef]
- Sanz-Rodriguez, F.; Guerrero-Esteo, M.; Botella, L.M.; Banville, D.; Vary, C.P.; Bernabéu, C. Endoglin regulates cytoskeletal organization through binding to ZRP-1, a member of the Lim family of proteins. J. Biol. Chem. 2004, 279, 32858–32868. [Google Scholar] [CrossRef]
- Larson, A.M. Liver disease in hereditary hemorrhagic telangiectasia. J. Clin. Gastroenterol. 2003, 36, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Llorca, O.; Trujillo, A.; Blanco, F.J.; Bernabeu, C. Structural model of human endoglin, a transmembrane receptor responsible for hereditary hemorrhagic telangiectasia. J. Mol. Biol. 2007, 365, 694–705. [Google Scholar] [CrossRef]
- Helmo, F.R.; Lopes, A.M.M.; Carneiro, A.C.D.M.; Campos, C.G.; Silva, P.B.; Dos Reis Monteiro, M.L.G.; Rocha, L.P.; Dos Reis, M.A.; Etchebehere, R.M.; Machado, J.R.; et al. Angiogenic and antiangiogenic factors in preeclampsia. Pathol. Res. Pract. 2018, 214, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.X.; Zhang, X.T.; Liao, Y.Q.; Zhang, Q.Y.; Chen, H.; Lin, M.; Kumar, S. Relationship between expression of CD105 and growth factors in malignant tumors of gastrointestinal tract and its significance. World J. Gastroenterol. 2003, 9, 2866–2869. [Google Scholar] [CrossRef]
- Dassoulas, K.; Gazouli, M.; Theodoropoulos, G.; Christoni, Z.; Rizos, S.; Zisi-Serbetzoglou, A.; Glava, C.; Karantanos, T.; Klonaris, C.; Karakitsos, P. Vascular endothelial growth factor and endoglin expression in colorectal cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Kobayashi, S.; Ohtsuka, M.; Kimura, F.; Shimizu, H.; Yoshidome, H.; Miyazaki, M. Specific expression of endoglin (CD105) in endothelial cells of intratumoral blood and lymphatic vessels in pancreatic cancer. Pancreas 2008, 37, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Poon, R.T.; Sun, C.K.; Xue, W.C.; Fan, S.T. Clinicopathological and prognostic implications of endoglin (CD105) expression in hepatocellular carcinoma and its adjacent non-tumorous liver. World J. Gastroenterol. 2005, 11, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Yang, L.; Zhao, W.; Chen, H.; He, S. A comparison of CD105 and CD31 expression in tumor vessels of hepatocellular carcinoma by tissue microarray and flow cytometry. Exp. Ther. Med. 2018, 16, 2881–2888. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, C.; Cunha, S.I.; Zhai, Z.; Cortez, E.; Pardali, E.; Johnson, J.R.; Franco, M.; Páez-Ribes, M.; Cordiner, R.; Fuxe, J.; et al. Deficiency for endoglin in tumor vasculature weakens the endothelial barrier to metastatic dissemination. J. Exp. Med. 2013, 210, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Van Le, B.; Franke, D.; Svergun, D.I.; Han, T.; Hwang, H.Y.; Kim, K.K. Structural and functional characterization of soluble endoglin receptor. Biochem. Biophys. Res. Commun. 2009, 383, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Varejckova, M.; Gallardo-Vara, E.; Vicen, M.; Vitverova, B.; Fikrova, P.; Dolezelova, E.; Rathouska, J.; Prasnicka, A.; Blazickova, K.; Micuda, S.; et al. Soluble endoglin modulates the pro-inflammatory mediators NF-κB and IL-6 in cultured human endothelial cells. Life Sci. 2017, 175, 52–60. [Google Scholar] [CrossRef]
- Fernández-Ruiz, E.; St-Jacques, S.; Bellón, T.; Letarte, M.; Bernabéu, C. Assignment of the human endoglin gene (END) to 9q34-->qter. Cytogenet. Cell Genet. 1993, 64, 204–207. [Google Scholar] [CrossRef]
- ENG Gene (Protein Coding). Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=ENG&keywords=CD105 (accessed on 15 October 2018).
- Meurer, S.K.; Tihaa, L.; Lahme, B.; Gressner, A.M.; Weiskirchen, R. Identification of endoglin in rat hepatic stellate cells: New insights into transforming growth factor β receptor signaling. J. Biol. Chem. 2005, 280, 3078–3087. [Google Scholar] [CrossRef]
- Meurer, S.K.; Alsamman, M.; Sahin, H.; Wasmuth, H.E.; Kisseleva, T.; Brenner, D.A.; Trautwein, C.; Weiskirchen, R.; Scholten, D. Overexpression of endoglin modulates TGF-β1-signalling pathways in a novel immortalized mouse hepatic stellate cell line. PLoS ONE 2013, 8, e56116. [Google Scholar] [CrossRef]
- Guerrero-Esteo, M.; Lastres, P.; Letamendía, A.; Pérez-Alvarez, M.J.; Langa, C.; López, L.A.; Fabra, A.; García-Pardo, A.; Vera, S.; Letarte, M.; Bernabéu, C. Endoglin overexpression modulates cellular morphology, migration, and adhesion of mouse fibroblasts. Eur. J. Cell Biol. 1999, 78, 614–623. [Google Scholar] [CrossRef]
- Shchyogolev, A.I.; Tumanova, U.N.; Dubova, E.A. Morphometric evaluation of vascularization of hepatic focal nodular hyperplasia. Bull. Exp. Biol. Med. 2013, 156, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wu, J. Hypoxia inducible factor in hepatocellular carcinoma: A therapeutic target. World J. Gastroenterol. 2015, 21, 12171–12178. [Google Scholar] [CrossRef]
- Sánchez-Elsner, T.; Botella, L.M.; Velasco, B.; Langa, C.; Bernabéu, C. Endoglin expression is regulated by transcriptional cooperation between the hypoxia and transforming growth factor-β pathways. J. Biol. Chem. 2002, 277, 43799–43808. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Guo, B.; Ding, S.; Rius, C.; Langa, C.; Kumar, P.; Bernabeu, C.; Kumar, S. TNF alpha down-regulates CD105 expression in vascular endothelial cells: A comparative study with TGF β 1. Anticancer Res. 2003, 23, 1189–1196. [Google Scholar] [PubMed]
- Tian, F.; Zhou, A.X.; Smits, A.M.; Larsson, E.; Goumans, M.J.; Heldin, C.H.; Borén, J.; Akyürek, L.M. Endothelial cells are activated during hypoxia via endoglin/ALK-1/SMAD1/5 signaling in vivo and in vitro. Biochem. Biophys. Res. Commun. 2010, 392, 283–288. [Google Scholar] [CrossRef]
- Wang, J.M.; Kumar, S.; van Agthoven, A.; Kumar, P.; Pye, D.; Hunter, R.D. Irradiation induces up-regulation of E9 protein (CD105) in human vascular endothelial cells. Int. J. Cancer 1995, 62, 791–796. [Google Scholar] [CrossRef]
- Gressner, A.M.; Weiskirchen, R. Modern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-β as major players and therapeutic targets. J. Cell Mol. Med. 2006, 10, 76–99. [Google Scholar] [CrossRef]
- García-Pozo, L.; Miquilena-Colina, M.E.; Lozano-Rodríguez, T.; García-Monzón, C. Endoglin: Structure, biological functions, and role in fibrogenesis. Rev. Esp. Enferm. Dig. 2008, 100, 355–360. [Google Scholar] [CrossRef]
- Wiercinska, E.; Wickert, L.; Denecke, B.; Said, H.M.; Hamzavi, J.; Gressner, A.M.; Thorikay, M.; ten Dijke, P.; Mertens, P.R.; Breitkopf, K.; et al. Id1 is a critical mediator in TGF-β-induced transdifferentiation of rat hepatic stellate cells. Hepatology 2006, 43, 1032–1041. [Google Scholar] [CrossRef]
- Clemente, M.; Núñez, O.; Lorente, R.; Rincón, D.; Matilla, A.; Salcedo, M.; Catalina, M.V.; Ripoll, C.; Iacono, O.L.; Bañares, R.; et al. Increased intrahepatic and circulating levels of endoglin, a TGF-β1 co-receptor, in patients with chronic hepatitis C virus infection: Relationship to histological and serum markers of hepatic fibrosis. J. Viral. Hepat. 2006, 13, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Prystupa, A.; Kiciński, P.; Luchowska-Kocot, D.; Błażewicz, A.; Kurys-Denis, E.; Niedziałek, J.; Sak, J.; Panasiuk, L. Relationships between serum selenium and zinc concentrations versus profibrotic and proangiogenic cytokines (FGF-19 and endoglin) in patients with alcoholic liver cirrhosis. Ann. Agric. Environ. Med. 2017, 24, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tang, Z.Y.; Fan, J.; Wu, Z.Q.; Li, X.M.; Liu, Y.K.; Liu, F.; Sun, H.C.; Ye, S.L. Expression of platelet-derived endothelial cell growth factor and vascular endothelial growth factor in hepatocellular carcinoma and portal vein tumor thrombus. J. Cancer Res. Clin. Oncol. 2000, 126, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, L.; Lin, Q.; Ren, W.; Xu, G. Prognostic value of endoglin-assessed microvessel density in cancer patients: A systematic review and meta-analysis. Oncotarget 2017, 9, 7660–7671. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, Q.; Guo, L.; Han, C. The prognostic correlation between CD105 expression level in tumor tissue and peripheral blood and sunitinib administration in advanced hepatocellular carcinoma. Cancer Biol. Ther. 2018, 25, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Takahashi, T.; Masuda, H.; Kalka, C.; Chen, D.; Iwaguro, H.; Inai, Y.; Silver, M.; Isner, J.M. VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 1999, 18, 3964–3972. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Muro, H.; Suzuki, S.; Sakaguchi, T.; Konno, H.; Baba, S.; Syed, A.S. Immunohistochemical studies on endothelial cell phenotype in hepatocellular carcinoma. Hepatology 1997, 26, 407–415. [Google Scholar] [CrossRef]
- Benetti, A.; Berenzi, A.; Gambarotti, M.; Garrafa, E.; Gelati, M.; Dessy, E.; Portolani, N.; Piardi, T.; Giulini, S.M.; Caruso, A.; et al. Transforming growth factor-β1 and CD105 promote the migration of hepatocellular carcinoma-derived endothelium. Cancer Res. 2008, 68, 8626–8634. [Google Scholar] [CrossRef]
- Xiong, Y.Q.; Sun, H.C.; Zhang, W.; Zhu, X.D.; Zhuang, P.Y.; Zhang, J.B.; Wang, L.; Wu, W.Z.; Qin, L.X.; Tang, Z.Y. Human hepatocellular carcinoma tumor-derived endothelial cells manifest increased angiogenesis capability and drug resistance compared with normal endothelial cells. Clin. Cancer Res. 2009, 15, 4838–4846. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, L.; Chen, X.; Qian, H.; Zhang, S.; Chen, Y.; Luo, R.; Shao, J.; Liu, H.; Chen, J. Phenotypic and functional characterization of tumor-derived endothelial cells isolated from primary human hepatocellular carcinoma. Hepatol. Res. 2018. [Google Scholar] [CrossRef]
- Warrington, K.; Hillarby, M.C.; Li, C.; Letarte, M.; Kumar, S. Functional role of CD105 in TGF-β1 signalling in murine and human endothelial cells. Anticancer Res. 2005, 25, 1851–1864. [Google Scholar] [PubMed]
- Kwon, Y.C.; Bose, S.K.; Steele, R.; Meyer, K.; Di Bisceglie, A.M.; Ray, R.B.; Ray, R. Promotion of Cancer Stem-Like Cell Properties in Hepatitis C Virus-Infected Hepatocytes. J. Virol. 2015, 89, 11549–11556. [Google Scholar] [CrossRef] [PubMed]
- Mardomi, A.; Sabzichi, M.; Hussein Somi, M.; Shanehbandi, D.; Rahbarghazi, R.; Taj Sanjarani, O.; Samadi, N. Trafficking mechanism of bone marrow-derived mesenchymal stem cells toward hepatocellular carcinoma HepG2 cells by modulating Endoglin, CXCR4 and TGF-β. Cell Mol. Biol. (Noisy-le-grand) 2016, 62, 81–86. [Google Scholar]
- Casadei Gardini, A.; Santini, D.; Aprile, G.; Silvestris, N.; Felli, E.; Foschi, F.G.; Ercolani, G.; Marisi, G.; Valgiusti, M.; Passardi, A.; et al. Antiangiogenic agents after first line and sorafenib plus chemoembolization: A systematic review. Oncotarget 2017, 8, 66699–66708. [Google Scholar] [CrossRef]
- Tan, G.H.; Huang, F.Y.; Wang, H.; Huang, Y.H.; Lin, Y.Y.; Li, Y.N. Immunotherapy of hepatoma with a monoclonal antibody against murine endoglin. World J. Gastroenterol. 2007, 13, 2479–2483. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Kang, G.; Wang, T.; Huang, H. Tumor angiogenesis and anti-angiogenic gene therapy for cancer. Oncol. Lett. 2018, 16, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Tansi, F.; Rüger, R.; Kollmeier, A.M.; Rabenhold, M.; Steiniger, F.; Kontermann, R.E.; Teichgraeber, U.K.; Fahr, A.; Hilger, I. Endoglin based in vivo near-infrared fluorescence imaging of tumor models in mice using activatable liposomes. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1389–1400. [Google Scholar] [CrossRef]
- Kim, H.C. Radioembolization for the treatment of hepatocellular carcinoma. Clin. Mol. Hepatol. 2017, 23, 109–114. [Google Scholar] [CrossRef]
- Nolan-Stevaux, O.; Zhong, W.; Culp, S.; Shaffer, K.; Hoover, J.; Wickramasinghe, D.; Ruefli-Brasse, A. Endoglin requirement for BMP9 signaling in endothelial cells reveals new mechanism of action for selective anti-endoglin antibodies. PLoS ONE 2012, 7, e50920. [Google Scholar] [CrossRef]
- Duan, C.L.; Hou, G.H.; Liu, Y.P.; Liang, T.; Song, J.; Han, J.K.; Zhang, C. Tumor vascular homing endgolin-targeted radioimmunotherapy in hepatocellular carcinoma. Tumour Biol. 2014, 35, 12205–12215. [Google Scholar] [CrossRef]
- Duffy, A.G.; Ma, C.; Ulahannan, S.V.; Rahma, O.E.; Makarova-Rusher, O.; Cao, L.; Yu, Y.; Kleiner, D.E.; Trepel, J.; Lee, M.J.; et al. Phase I and preliminary phase II study of TRC105 in combination with sorafenib in hepatocellular carcinoma. Clin. Cancer Res. 2017, 23, 4633–4641. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.L.; Li, Y.; Yang, W.; Zhang, C. Combination of 131 I-anti-endoglin monoclonal antibody and 5-fluorouracil may be a promising combined-modality radioimmunotherapy strategy for the treatment of hepatocellular carcinoma. Biotechnol. Biotechnol. Equip. 2018. [Google Scholar] [CrossRef]
- Rosen, L.S.; Hurwitz, H.I.; Wong, M.K.; Goldman, J.; Mendelson, D.S.; Figg, W.D.; Spencer, S.; Adams, B.J.; Alvarez, D.; Seon, B.K.; et al. A phase I first-in-human study of TRC105 (Anti-Endoglin Antibody) in patients with advanced cancer. Clin. Cancer Res. 2012, 18, 4820–4829. [Google Scholar] [CrossRef] [PubMed]
- Karzai, F.H.; Apollo, A.B.; Cao, L.; Madan, R.A.; Adelberg, D.E.; Parnes, H.; McLeod, D.G.; Harold, N.; Peer, C.; Yu, Y.; et al. A phase I study of TRC105 anti-endoglin (CD105) antibody in metastatic castration-resistant prostate cancer. BJU Int. 2015, 116, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.S.; Robert, F.; Matei, D.; Mendelson, D.S.; Goldman, J.W.; Chiorean, E.G.; Strother, R.M.; Seon, B.K.; Figg, W.D.; Peer, C.J.; et al. An open-label phase Ib dose-escalation study of TRC105 (anti-endoglin antibody) with bevacizumab in patients with advanced cancer. Clin. Cancer Res. 2014, 20, 5918–5926. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.G.; Ulahannan, S.V.; Cao, L.; Rahma, O.E.; Makarova-Rusher, O.V.; Kleiner, D.E.; Fioravanti, S.; Walker, M.; Carey, S.; Yu, Y.; et al. A phase II study of TRC105 in patients with hepatocellular carcinoma who have progressed on sorafenib. United Eur. Gastroenterol. J. 2015, 3, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Dorff, T.B.; Longmate, J.A.; Pal, S.K.; Stadler, W.M.; Fishman, M.N.; Vaishampayan, U.N.; Rao, A.; Pinksi, J.K.; Hu, J.S.; Quinn, D.I.; et al. Bevacizumab alone or in combination with TRC105 for patients with refractory metastatic renal cell cancer. Cancer 2017, 123, 4566–4573. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Michaelson, M.D.; Posadas, E.M.; Sonpavde, G.P.; McDermott, D.F.; Nixon, A.B.; Liu, Y.; Yuan, Z.; Seon, B.K.; Walsh, M.; et al. An open label phase Ib dose escalation study of TRC105 (anti-endoglin antibody) with axitinib in patients with metastatic renal cell carcinoma. Oncologist 2018. [Google Scholar] [CrossRef]
- Madhav, A.; Andres, A.; Duong, F.; Mishra, R.; Haldar, S.; Liu, Z.; Angara, B.; Gottlieb, R.; Zumsteg, Z.S.; Bhowmick, N.A. Antagonizing CD105 enhances radiation sensitivity in prostate cancer. Oncogene 2018, 37, 4385–4397. [Google Scholar] [CrossRef]
- Kumari, P.; Ghosh, B.; Biswas, S. Nanocarriers for cancer-targeted drug delivery. J. Drug Target 2016, 24, 179–191. [Google Scholar] [CrossRef]
- Abdalla, A.M.E.; Xiao, L.; Ullah, M.W.; Yu, U.; Ouyang, C.; Yang, G. Current challenges of cancer antiangiogenic therapy and the promise of nanotherapeutics. Theranostics 2018, 8, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, S. Anticancer activity of eco-friendly gold nanoparticles against lung and liver cancer cells. J. Genet. Eng. Biotechnol. 2016, 14, 195–202. [Google Scholar] [CrossRef]
- Saratale, R.G.; Benelli, G.; Kumar, G.; Kim, D.S.; Saratale, G.D. Bio-fabrication of silver nanoparticles using the leaf extract of an ancient herbal medicine, dandelion (Taraxacum officinale), evaluation of their antioxidant, anticancer potential, and antimicrobial activity against phytopathogens. Environ. Sci. Pollut. Res. Int. 2018, 25, 10392–10406. [Google Scholar] [CrossRef] [PubMed]
- Chajon, E.; Pracht, M.; De Baere, T.; Nguyen, F.; Bronowicki, J.P.; Vendrely, V.; Baumann, A.S.; Croisé-Laurent, V.; Rio, E.; Rolland, Y.; et al. NBTXR3, hafnium oxide nanoparticles in the treatment of liver cancer. A phase I/II trial. J. Clin. Oncol. 2018, 36 (Suppl. 15), e16194. [Google Scholar]
- Zhao, X.; Zhou, S.; Wang, D.; He, W.; Li, J.; Zhang, S. Dual-intelligent functionalized silica nanoparticles for liver cancer imaging and therapy. Int. J. Clin. Exp. Med. 2016, 9, 13584–13594. [Google Scholar]
- Ao, M.; Xiao, X.; Ao, Y. Low density lipoprotein modified silica nanoparticles loaded with docetaxel and thalidomide for effective chemotherapy of liver cancer. Braz. J. Med. Biol. Res. 2018, 51, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.; Lin, C.T.; Wu, H.S. Hepatocellular carcinoma cell-specific peptide ligand for targeted drug delivery. Mol. Cancer Ther. 2008, 7, 579–589. [Google Scholar] [CrossRef]
- Wu, C.H.; Lan, C.H.; Wu, K.L.; Wu, Y.M.; Jane, W.N.; Hsiao, M.; Wu, H.C. Hepatocellular carcinoma-targeted nanoparticles for cancer therapy. Int. J. Oncol. 2018, 52, 389–401. [Google Scholar] [CrossRef]
- Karmani, L.; Bouchat, V.; Bouzin, C.; Levêque, P.; Labar, D.; Bol, A.; Deumer, G.; Marega, R.; Bonifazi, D.; Haufroid, V.; et al. (89)Zr-labeled anti-endoglin antibody-targeted gold nanoparticles for imaging cancer: Implications for future cancer therapy. Nanomedicine 2014, 9, 1923–1937. [Google Scholar] [CrossRef]
- Zhang, Y.; Chang, R.; Zhao, K. CD105-Loaded docetaxel lipid microbubble combined with ultrasound–triggered microbubble destruction for inhibition of proliferation of HCC in vitro and in vivo. Ultrasound Med. Biol. 2017, 43, S1–S44. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Hong, H.; Cai, W. Multimodality molecular imaging of CD105 (Endoglin) expression. Int. J. Clin. Exp. Med. 2011, 4, 32–42. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Criteria | Characteristics | No. of Ref. |
|---|---|---|
| Gene Location and size (kb) | Chromosome 9q34.11; ~40 kb; 14–16 exons, human gene encodes 15 exons | [40,41,42,108,109] |
| Class of Genes | Protein coding; Zona pellucida family; CD molecules | [98,109] |
| No. of Transcripts | 3 | [109] |
| Protein m.w. (kd), no. of Amino Acids | Homodimer, ~180 kD; 2 subunits 95 kDa disulfide-linked; 633-658 aa | [38,42,68,75,81,98] |
| Cellular Sources in Liver | Quiescent ECs in sinusoids, arterioles and venules | [23,39,40,41,42,43,100] |
| Activated LSECs or TECs | ||
| Normal and cancerous hepatocytes | [44,45,49] | |
| HCV core-expressing human hepatocytes | [70] | |
| Myofibroblasts | [78] | |
| Hepatic stellate cells | [11,13,78,110,111] | |
| Hepatic perivascular mesenchymal stem cells | [48,53] | |
| Pericytes | [53] | |
| Adipose-derived stem cells | [52] | |
| Adult-derived human liver stem/progenitor cells | [49,51] | |
| Main Role in Liver Physiology | Angiogenesis and vascular development | [24,39,40,85,88] |
| Vascular homeostasis | ||
| Maintenance of vessel wall integrity | ||
| Main Role in Liver Pathology | Stimulation of tumour neovasculature | [88,89] |
| Vascular remodelling/modulation of vascular tone | [24,88,89] | |
| Fibrosis (stimulation/inhibition) | [11,69,89,112,113] | |
| Diagnostic biomarker in HCC | [17,18,22] | |
| Risk factor of HCC in patients with liver cirrhosis | [17,19] | |
| Predictor of HCC early recurrence/distant metastasis | [14,103] | |
| Risk factor for post-transplant recurrence | [21] | |
| Predictor of decreased survival in HCC | [14,15,104] |
| Tissue Expression | |||
|---|---|---|---|
| C | NT | HCC | No. of Ref. |
| nt | (+); Variable pattern with diffuse expression in some cases # | (+) (68% of HCC); 3 patterns of expression; ↓score ♣,♦ | [103] |
| (−) | (−) | (+) (100% of HCC) ↑score ♦,# | [14] |
| nt | (+) (100% cases); (+) in “normal” hepatocytes | (+) (80% of HCC); (+) in some cancer cells | [45] |
| (+) | ↑score; typical pattern | (+); typical pattern | [20] |
| nt | nt | (+) (90% of HCC); ↓score ♣,♦ ↑score# | [15] |
| nt | ↑↑↑score ♦,# | (−) | [21] |
| nt | ↑score in RN>DN>HCC | (+) | [12] |
| (+) | nt | ↑score ♣,♦ | [16] |
| nt | ↑score | (+); ↓score ♣ | [22] |
| nt | ↑score (100% cases) | (+) (57% of HCC) ↓score ♣,# | [104] |
| Serum Levels | |||
| (+) | ↑↑ | Sol-ENG ↑↑↑ (HCC + cirrhosis) * | [17] |
| (+) | ↑ | Sol-ENG ↑↑,♦ | [18] |
| (+) | ↑↑ | mRNA-ENG ↑↑,♦ (HCC + cirrhosis) | [19] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprzak, A.; Adamek, A. Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy. Int. J. Mol. Sci. 2018, 19, 3887. https://doi.org/10.3390/ijms19123887
Kasprzak A, Adamek A. Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy. International Journal of Molecular Sciences. 2018; 19(12):3887. https://doi.org/10.3390/ijms19123887
Chicago/Turabian StyleKasprzak, Aldona, and Agnieszka Adamek. 2018. "Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy" International Journal of Molecular Sciences 19, no. 12: 3887. https://doi.org/10.3390/ijms19123887
APA StyleKasprzak, A., & Adamek, A. (2018). Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy. International Journal of Molecular Sciences, 19(12), 3887. https://doi.org/10.3390/ijms19123887