Effects of Arachidonic and Docosohexahenoic Acid Supplementation during Gestation in Rats. Implication of Placental Oxidative Stress


 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Diet Intake
2.2. Maternal Plasma Composition
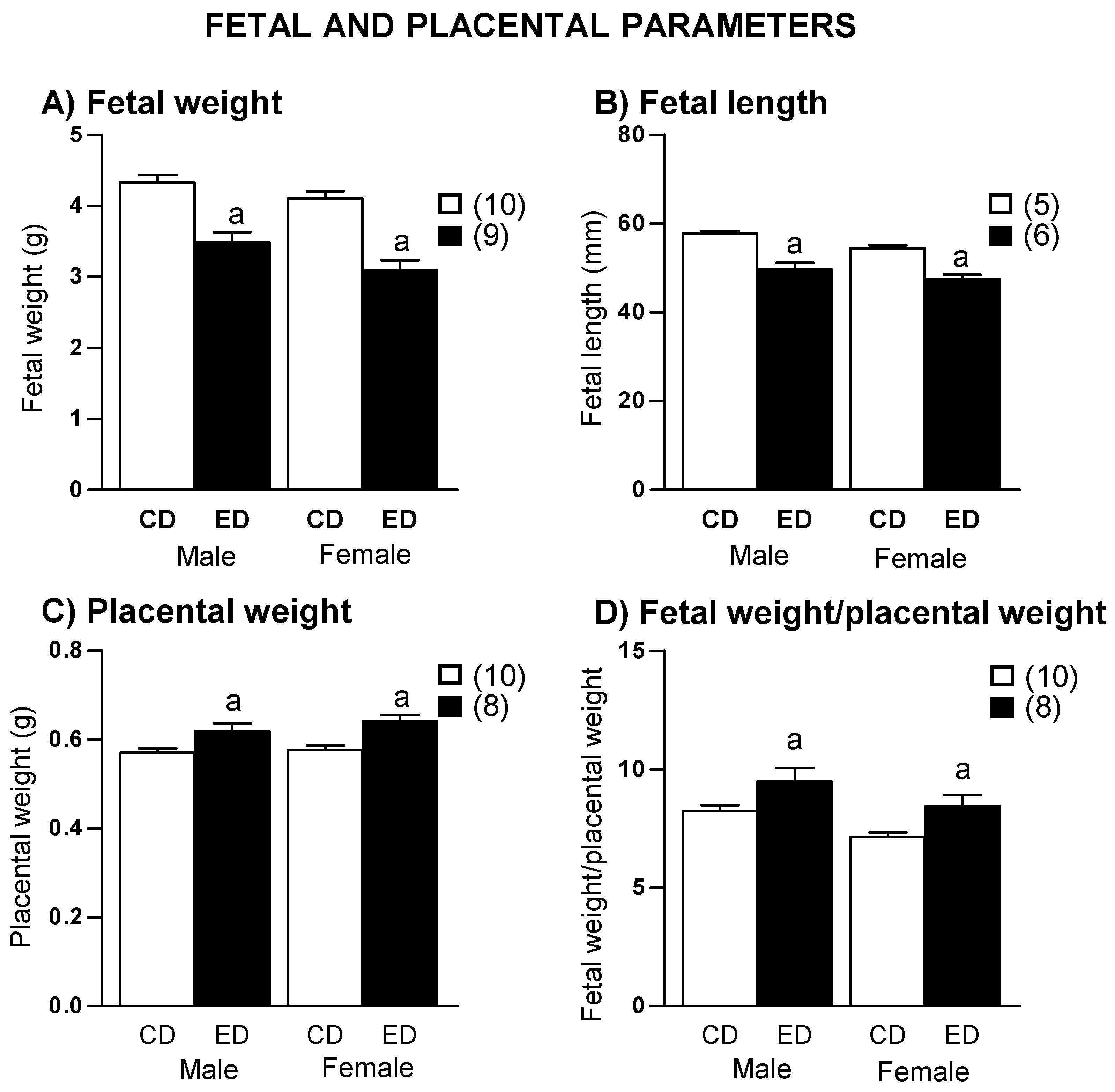
2.3. Gestation Outcome
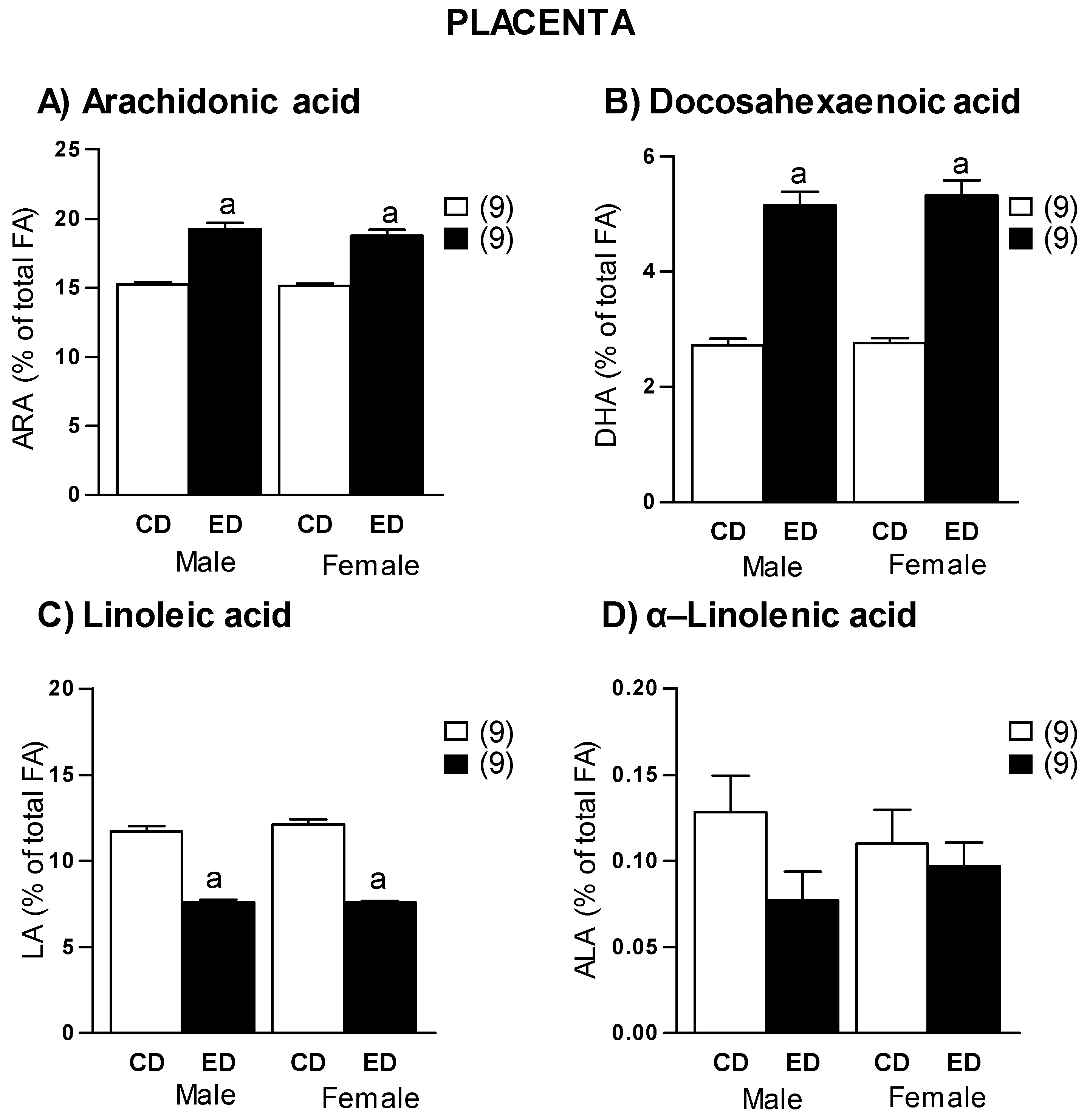
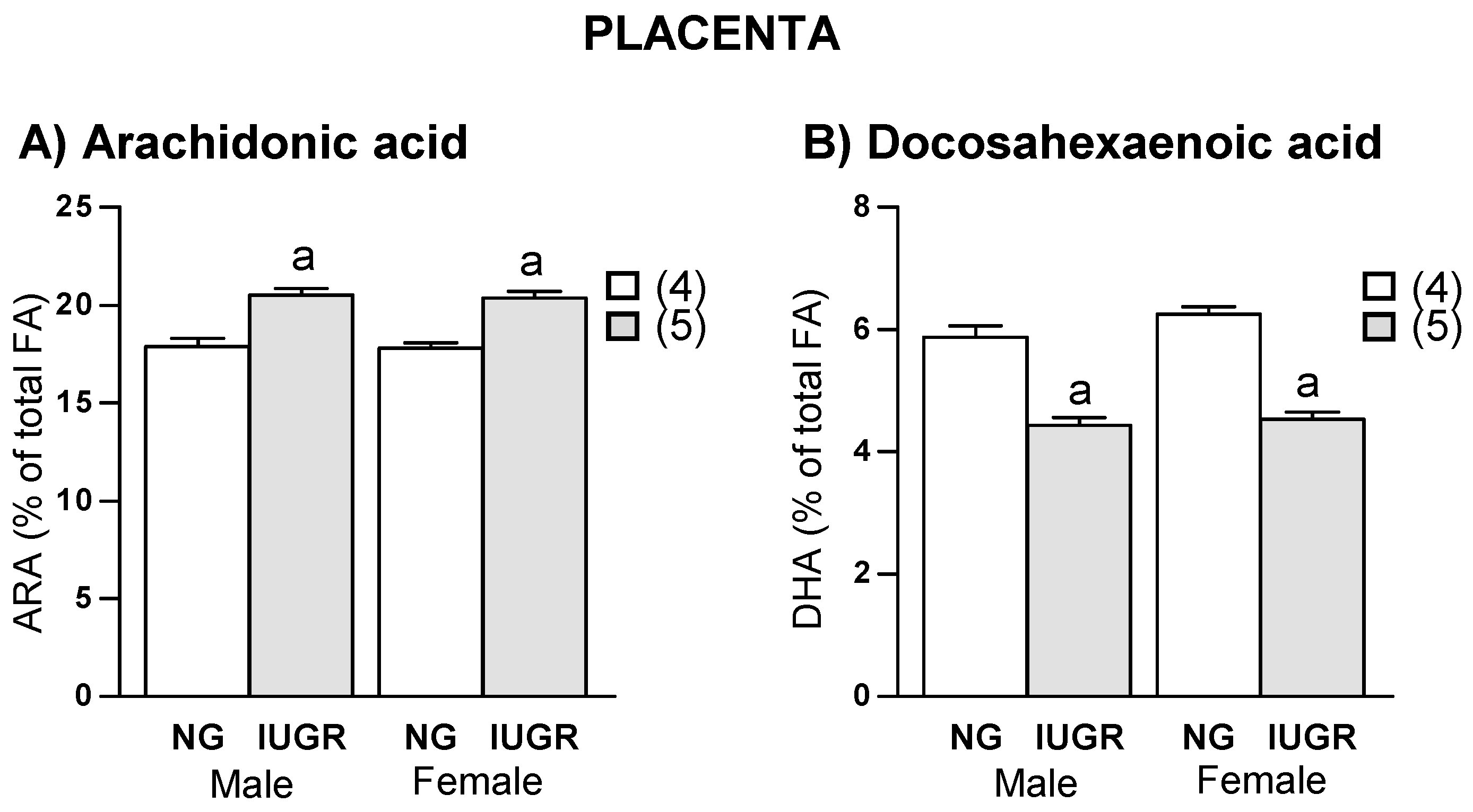
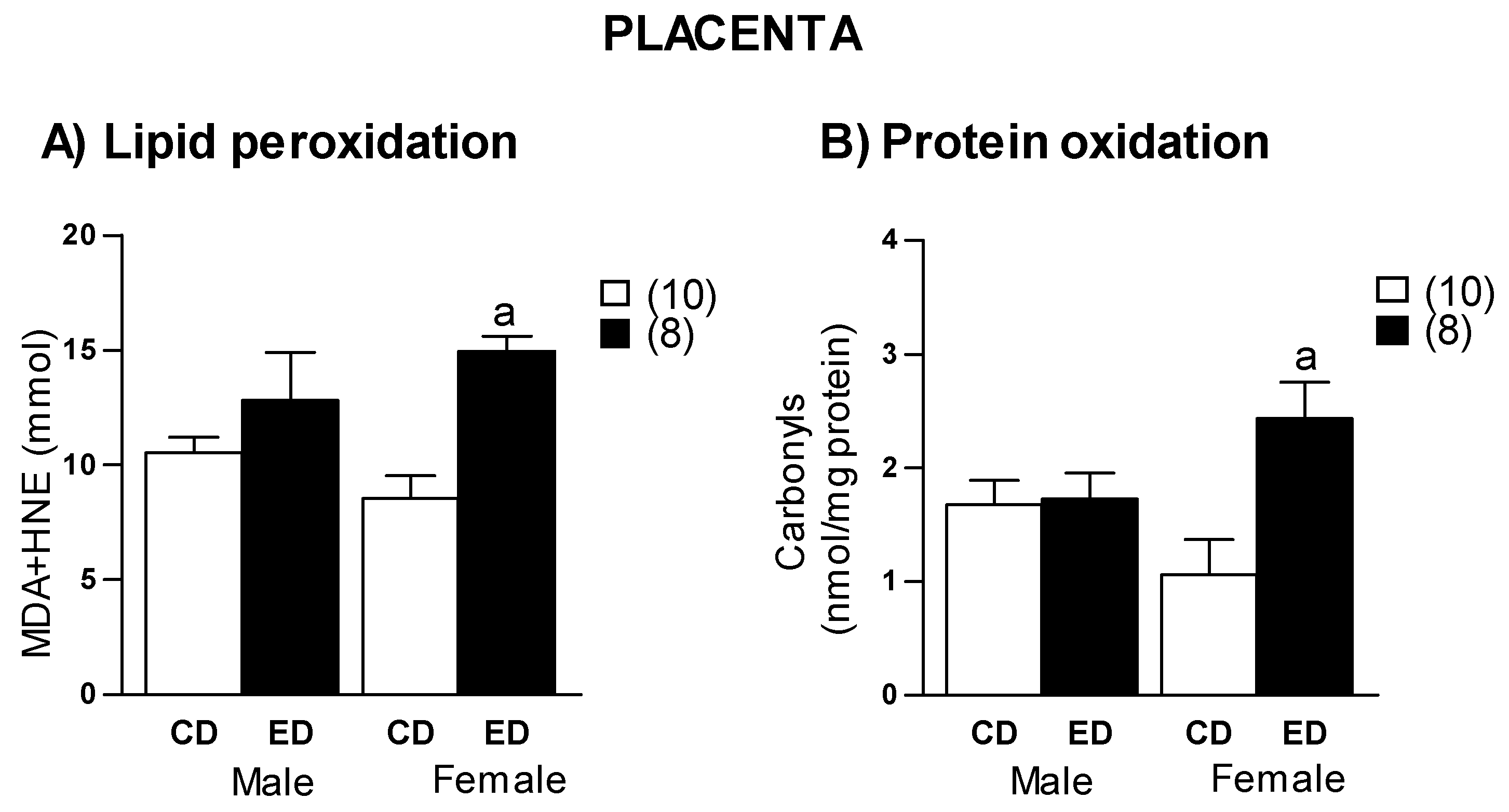
2.4. Placenta
3. Discussion
3.1. Maternal Bioavailability of LCPUFA from the Diet
3.2. Effect of Dietary Intervention on Gestation
3.3. Role of Placenta
3.4. Effect of Sex
4. Methods
4.1. Diet Preparation
4.2. Experimental Animals
4.3. Experimental Procedures
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, M.S.; Chen, Q.Z.; He, J.R.; Wei, X.L.; Lu, J.H.; Li, S.H.; Wen, X.X.; Chan, F.F.; Chen, N.N.; Qiu, L.; et al. Maternal Dietary Patterns and Fetal Growth: A Large Prospective Cohort Study in China. Nutrients 2016, 8, 257. [Google Scholar] [CrossRef]
- Lowensohn, R.I.; Stadler, D.D.; Naze, C. Current Concepts of Maternal Nutrition. Obstet. Gynecol. Surv. 2016, 71, 413–426. [Google Scholar] [CrossRef]
- Koletzko, B.; Lien, E.; Agostoni, C.; Böhles, H.; Campoy, C.; Cetin, I.; Decsi, T.; Dudenhausen, J.W.; Dupont, C.; Forsyth, S.; et al. The roles of long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy: Review of current knowledge and consensus recommendations. J. Perinat. Med. 2008, 36, 5–14. [Google Scholar] [CrossRef]
- Jones, M.L.; Mark, P.J.; Waddell, B.J. Maternal dietary omega-3 fatty acids and placental function. Reproduction 2014, 147, R143–R152. [Google Scholar] [CrossRef]
- Haggarty, P. Fatty acid supply to the human fetus. Annu. Rev. Nutr. 2010, 30, 237–255. [Google Scholar] [CrossRef]
- McCowen, K.C.; Bistrian, B.R. Essential fatty acids and their derivatives. Curr. Opin. Gastroenterol. 2005, 21, 207–215. [Google Scholar] [CrossRef]
- Haggarty, P. Placental regulation of fatty acid delivery and its effect on fetal growth—A review. Placenta 2002, 23 (Suppl. A), S28–S38. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Imhoff-Kunsch, B.; Briggs, V.; Goldenberg, T.; Ramakrishnan, U. Effect of n-3 long-chain polyunsaturated fatty acid intake during pregnancy on maternal, infant, and child health outcomes: A systematic review. Paediatr. Perinat. Epidemiol. 2012, 26 (Suppl. 1), 91–107. [Google Scholar] [CrossRef]
- Saccone, G.; Berghella, V.; Maruotti, G.M.; Sarno, L.; Martinelli, P. Omega-3 supplementation during pregnancy to prevent recurrent intrauterine growth restriction: Systematic review and meta-analysis of randomized controlled trials. Ultrasound Obstet. Gynecol. 2015, 46, 659–664. [Google Scholar] [CrossRef]
- Koletzko, B.; Boey, C.C.; Campoy, C.; Carlson, S.E.; Chang, N.; Guillermo-Tuazon, M.A.; Joshi, S.; Prell, C.; Quak, S.H.; Sjarif, D.R.; et al. Current information and Asian perspectives on long-chain polyunsaturated fatty acids in pregnancy, lactation, and infancy: Systematic review and practice recommendations from an early nutrition academy workshop. Ann. Nutr. Metab. 2014, 65, 49–80. [Google Scholar] [CrossRef] [PubMed]
- Fountain, E.D.; Mao, J.; Whyte, J.J.; Mueller, K.E.; Ellersieck, M.R.; Will, M.J.; Roberts, R.M.; MacDonald, R.; Rosenfeld, C.S. Effects of diets enriched in omega-3 and omega-6 polyunsaturated fatty acids on offspring sex-ratio and maternal behavior in mice. Biol. Reprod. 2008, 78, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Amusquivar, E.; Ruperez, F.J.; Barbas, C.; Herrera, E. Low arachidonic acid rather than alpha-tocopherol is responsible for the delayed postnatal development in offspring of rats fed fish oil instead of olive oil during pregnancy and lactation. J. Nutr. 2000, 130, 2855–2865. [Google Scholar] [CrossRef]
- Uauy-Dagach, R.; Mena, P. Nutritional role of omega-3 fatty acids during the perinatal period. Clin. Perinatol. 1995, 22, 157–175. [Google Scholar] [CrossRef]
- Jimenez, M.J.; Bocos, C.; Panadero, M.; Herrera, E. Fish oil diet in pregnancy and lactation reduces pup weight and modifies newborn hepatic metabolic adaptations in rats. Eur. J. Nutr. 2017, 56, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Song, J.H.; Fujimoto, K.; Miyazawa, T. Polyunsaturated (n-3) fatty acids susceptible to peroxidation are increased in plasma and tissue lipids of rats fed docosahexaenoic acid-containing oils. J. Nutr. 2000, 130, 3028–3033. [Google Scholar] [CrossRef]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef]
- Mori, T.A. Effect of fish and fish oil-derived omega-3 fatty acids on lipid oxidation. Redox Rep. 2004, 9, 193–197. [Google Scholar] [CrossRef]
- Leghi, G.E.; Muhlhausler, B.S. The effect of n-3 LCPUFA supplementation on oxidative stress and inflammation in the placenta and maternal plasma during pregnancy. Prostaglandins Leukot. Essent. Fatty Acids 2016, 113, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Arab, L. Biomarkers of fat and fatty acid intake. J. Nutr. 2003, 133 (Suppl. 3), 925S–932S. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Cooke, C.L.; Davidge, S.T. In Utero Origins of Hypertension: Mechanisms and Targets for Therapy. Physiol. Rev. 2016, 96, 549–603. [Google Scholar] [CrossRef] [PubMed]
- Nardozza, L.M.; Caetano, A.C.; Zamarian, A.C.; Mazzola, J.B.; Silva, C.P.; Marçal, V.M.; Lobo, T.F.; Peixoto, A.B.; Júnior, E.A. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Faraci, M.; Renda, E.; Monte, S.; Di Prima, F.A.; Valenti, O.; De Domenico, R.; Giorgio, E.; Hyseni, E. Fetal growth restriction: Current perspectives. J. Prenat. Med. 2011, 5, 31–33. [Google Scholar] [PubMed]
- Dickinson, H.; Moss, T.J.; Gatford, K.L.; Moritz, K.M.; Akison, L.; Fullston, T.; Hryciw, D.H.; Maloney, C.A.; Morris, M.J.; Wooldridge, A.L.; et al. A review of fundamental principles for animal models of DOHaD research: An Australian perspective. J. Dev. Orig. Health Dis. 2016, 7, 449–472. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E. Implications of dietary fatty acids during pregnancy on placental, fetal and postnatal development—A review. Placenta 2002, 23 (Suppl. A), S9–S19. [Google Scholar] [CrossRef] [PubMed]
- Hayward, C.E.; Lean, S.; Sibley, C.P.; Jones, R.L.; Wareing, M.; Greenwood, S.L.; Dilworth, M.R. Placental Adaptation: What Can We Learn from Birthweight: Placental Weight Ratio? Front. Physiol. 2016, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Chassen, S.S.; Ferchaud-Roucher, V.; Gupta, M.B.; Jansson, T.; Powell, T.L. Alterations in placental long chain polyunsaturated fatty acid metabolism in human intrauterine growth restriction. Clin. Sci. 2018, 132, 595–607. [Google Scholar] [CrossRef]
- Jauniaux, E.; Poston, L.; Burton, G.J. Placental-related diseases of pregnancy: Involvement of oxidative stress and implications in human evolution. Hum. Reprod. Update 2006, 12, 747–755. [Google Scholar] [CrossRef]
- Cuffe, J.S.; Xu, Z.C.; Perkins, A.V. Biomarkers of oxidative stress in pregnancy complications. Biomark. Med. 2017, 11, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; de Pablo, A.L.; Condezo-Hoyos, L.; Martín-Cabrejas, M.A.; Aguilera, Y.; Ruiz-Hurtado, G.; Gutierrez-Arzapalo, P.Y.; Ramiro-Cortijo, D.; Fernández-Alfonso, M.S.; del Carmen Gonzalez, M.; et al. Fetal undernutrition is associated with perinatal sex-dependent alterations in oxidative status. J. Nutr. Biochem. 2015, 26, 1650–1659. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; de Pablo, A.L.; García-Prieto, C.F.; Somoza, B.; Quintana-Villamandos, B.; de Diego, J.J.; Gutierrez-Arzapalo, P.Y.; Ramiro-Cortijo, D.; González, M.C.; Arribas, S.M. Long term effects of fetal undernutrition on rat heart. Role of hypertension and oxidative stress. PLoS ONE 2017, 12, e0171544. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Reyes-Hernández, C.G.; López de Pablo, A.L.; González, M.C.; Arribas, S.M. Implication of Oxidative Stress in Fetal Programming of Cardiovascular Disease. Front. Physiol. 2018, 9, 602. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zhang, X.; Sieli, P.T.; Falduto, M.T.; Torres, K.E.; Rosenfeld, C.S. Contrasting effects of different maternal diets on sexually dimorphic gene expression in the murine placenta. Proc. Natl. Acad. Sci. USA 2010, 107, 5557–5562. [Google Scholar] [CrossRef] [PubMed]
- Carnielli, V.P.; Pederzini, F.; Vittorangeli, R.; Luijendijk, I.H.; Boomaars, W.E.; Pedrotti, D.; Sauer, P.J. Plasma and red blood cell fatty acid of very low birth weight infants fed exclusively with expressed preterm human milk. Pediatr. Res. 1996, 39, 671–679. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef]
- Condezo-Hoyos, L.; Arribas, S.M.; Abderrahim, F.; Somoza, B.; Gil-Ortega, M.; Díaz-Gil, J.J.; Conde, M.V.; Susin, C.; González, M.C. Liver growth factor treatment reverses vascular and plasmatic oxidative stress in spontaneously hypertensive rats. J. Hypertens. 2012, 30, 1185–1194. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | |||
|---|---|---|---|---|
| CD | ED | CD | ED | |
| CRL (mm) | 44.7 ± 0.4 | 42.7 ± 0.6 a | 42.7 ± 0.5 | 40.4 ± 0.6 a |
| BMI (g/cm2) | 0.254 ± 0.004 | 0.218 ± 0.004 b | 0.258 ± 0.004 | 0.204 ± 0.004 b |
| Diet Composition | CD | ED |
|---|---|---|
| Total digestible nutrients (%) | 77.8 | 73.2 |
| Calories provided by protein (%) | 25.9 | 25.7 |
| Calories provided by CHO (%) | 64.8 | 62.9 |
| Calories provided by fat (%) | 9.3 | 10.7 |
| Protein content (%) | 22.0 | 22.0 |
| CHO (nitrogen free extract) content (%) | 55.0 | 53.9 |
| Total fat content (%) | 4.4 | 4.4 |
| Ash content (%) | 5.4 | 5.4 |
| Fiber maximum content (%) | 4.1 | 4.3 |
| Fat source | Soybean oil | Formulaid™ 2:1 |
| Fatty Acids (%) | ED | CD |
|---|---|---|
| Lauric Acid (12:0) | 1.3 | - |
| Myristic Acid (14:0) | 3.1 | 0.1 |
| Palmitic Acid (16:0) | 14.2 | 16.3 |
| Palmitoleic Acid (16:1w7) | 0.5 | 0.2 |
| Stearic Acid (18:0) | 4.5 | 2.7 |
| Oleic Acid (18:1w9) | 20.8 | 20.4 |
| Linoleic Acid (18:2w6) | 27.5 | 53.5 |
| Linolenic Acid (18:3w6) | 1.0 | - |
| Alpha Linoleic Acid (18:3w3) | 3.0 | 4.6 |
| Arachidic Acid (20:0) | 0.4 | 0.3 |
| Gondoic Acid (20:1w9) | 0.2 | 0.3 |
| Eicosadienoic Acid (20:2w6) | 0.2 | - |
| Eicosatrienoic Acid (20:3w6) | 0.9 | - |
| Arachidonic Acid (20:4w6) | 13.5 | - |
| Eicosapentaenoic Acid (20:5w3) | 0.1 | - |
| Behenic Acid (22:0) | 0.5 | - |
| Docosahexaenoic Acid (22:6w3) | 6.3 | - |
| Lignoceric Acid (24:0) | 0.4 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Hernández, C.G.; Ramiro-Cortijo, D.; Rodríguez-Rodríguez, P.; Giambelluca, S.; Simonato, M.; González, M.d.C.; López de Pablo, A.L.; López-Giménez, M.d.R.; Cogo, P.; Sáenz de Pipaón, M.; et al. Effects of Arachidonic and Docosohexahenoic Acid Supplementation during Gestation in Rats. Implication of Placental Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 3863. https://doi.org/10.3390/ijms19123863
Reyes-Hernández CG, Ramiro-Cortijo D, Rodríguez-Rodríguez P, Giambelluca S, Simonato M, González MdC, López de Pablo AL, López-Giménez MdR, Cogo P, Sáenz de Pipaón M, et al. Effects of Arachidonic and Docosohexahenoic Acid Supplementation during Gestation in Rats. Implication of Placental Oxidative Stress. International Journal of Molecular Sciences. 2018; 19(12):3863. https://doi.org/10.3390/ijms19123863
Chicago/Turabian StyleReyes-Hernández, Cynthia Guadalupe, David Ramiro-Cortijo, Pilar Rodríguez-Rodríguez, Sonia Giambelluca, Manuela Simonato, Mª del Carmen González, Angel Luis López de Pablo, Mª del Rosario López-Giménez, Paola Cogo, Miguel Sáenz de Pipaón, and et al. 2018. "Effects of Arachidonic and Docosohexahenoic Acid Supplementation during Gestation in Rats. Implication of Placental Oxidative Stress" International Journal of Molecular Sciences 19, no. 12: 3863. https://doi.org/10.3390/ijms19123863
APA StyleReyes-Hernández, C. G., Ramiro-Cortijo, D., Rodríguez-Rodríguez, P., Giambelluca, S., Simonato, M., González, M. d. C., López de Pablo, A. L., López-Giménez, M. d. R., Cogo, P., Sáenz de Pipaón, M., Carnielli, V. P., & Arribas, S. M. (2018). Effects of Arachidonic and Docosohexahenoic Acid Supplementation during Gestation in Rats. Implication of Placental Oxidative Stress. International Journal of Molecular Sciences, 19(12), 3863. https://doi.org/10.3390/ijms19123863