Endothelial Progenitor Cells as Pathogenetic and Diagnostic Factors, and Potential Targets for GLP-1 in Combination with Metabolic Syndrome and Chronic Obstructive Pulmonary Disease

 , , , ,
, , , ,  and
and 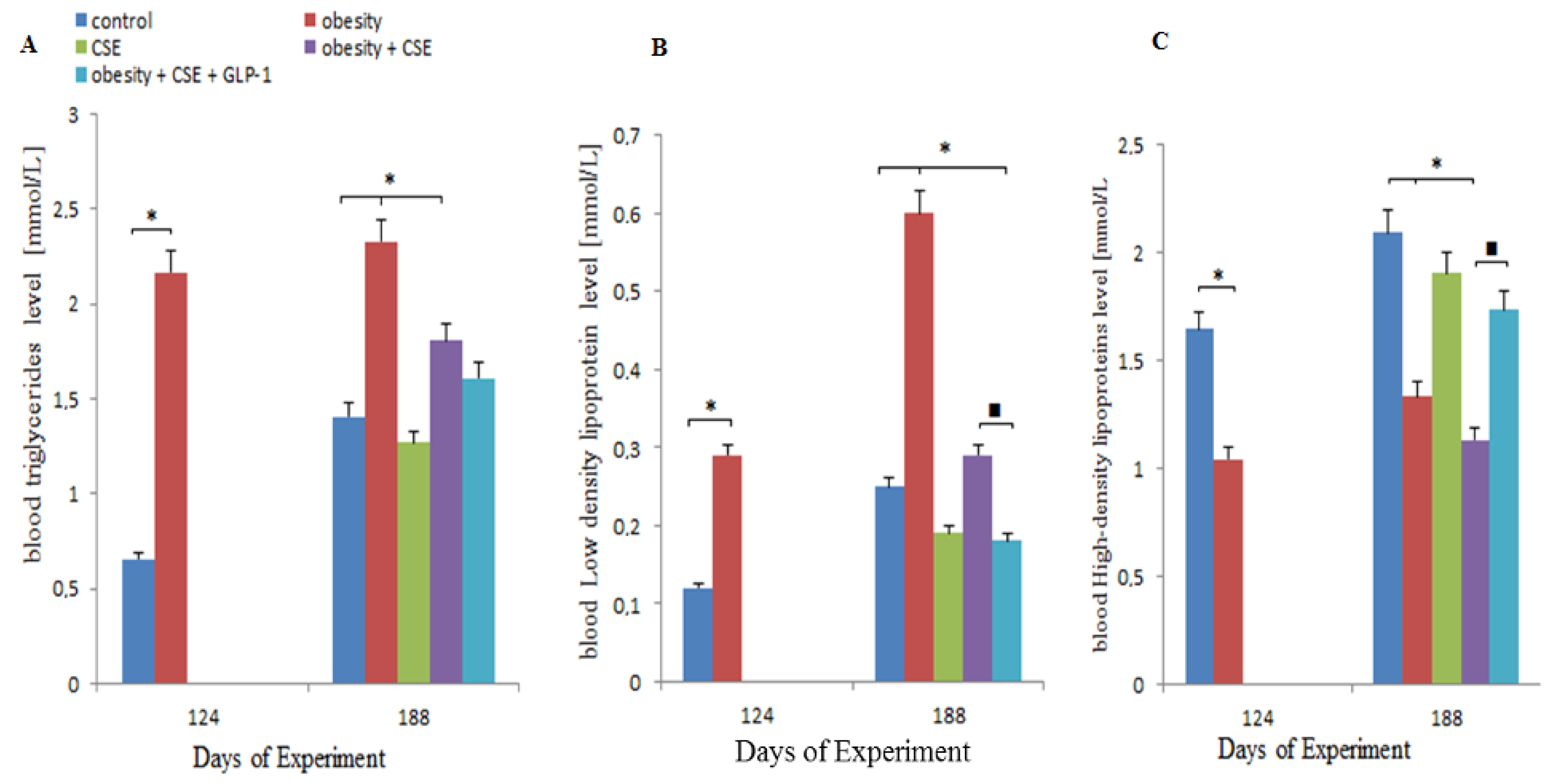{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Changes in Serum Biochemical Parameters in MSG-Treated Mice Prior to Pulmonary Emphysema Modeling
2.2. Effect of GLP-1 on Serum Lipid Profile
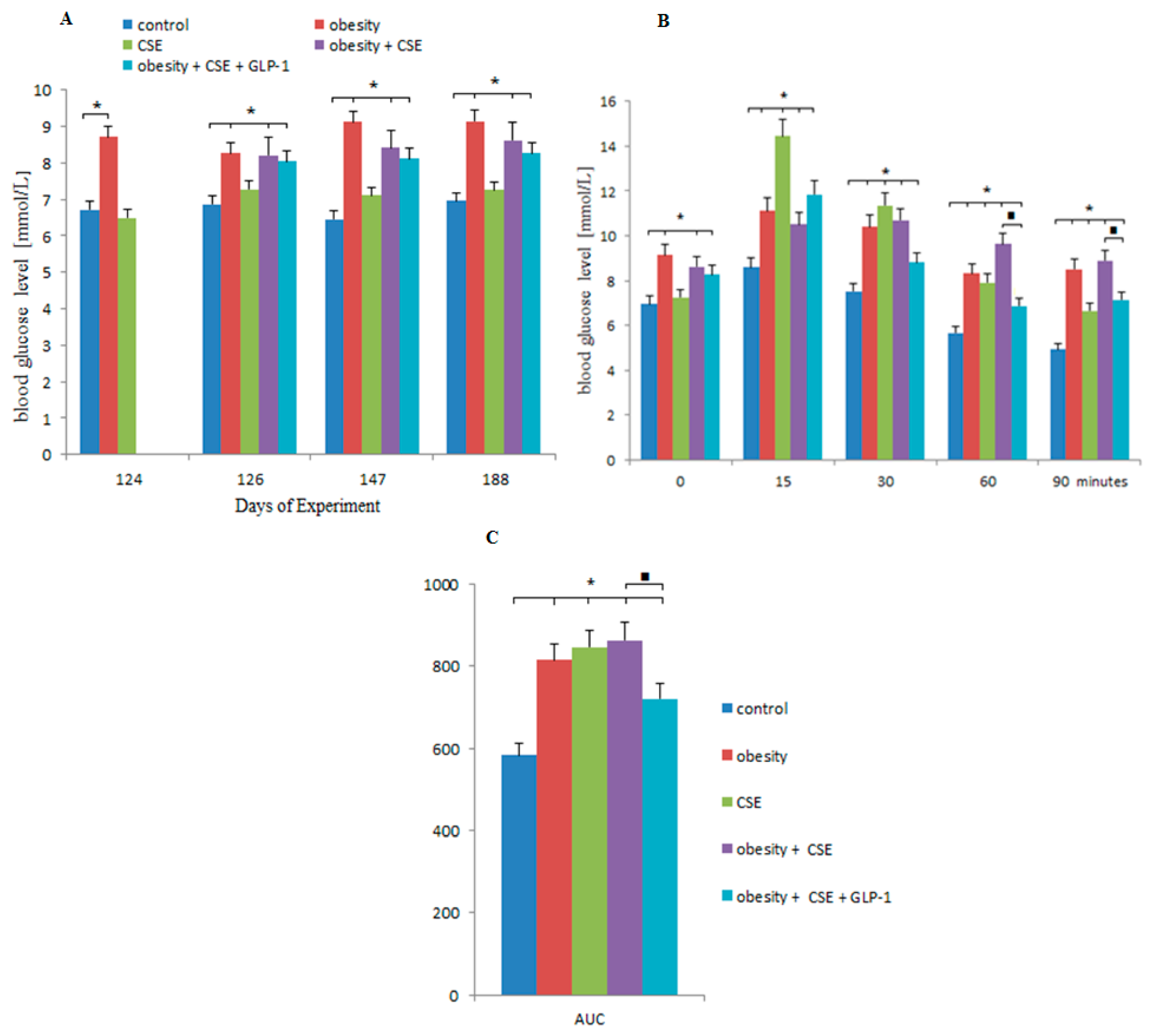
2.3. Effect of GLP-1 on Blood Glucose, GTT and AUC
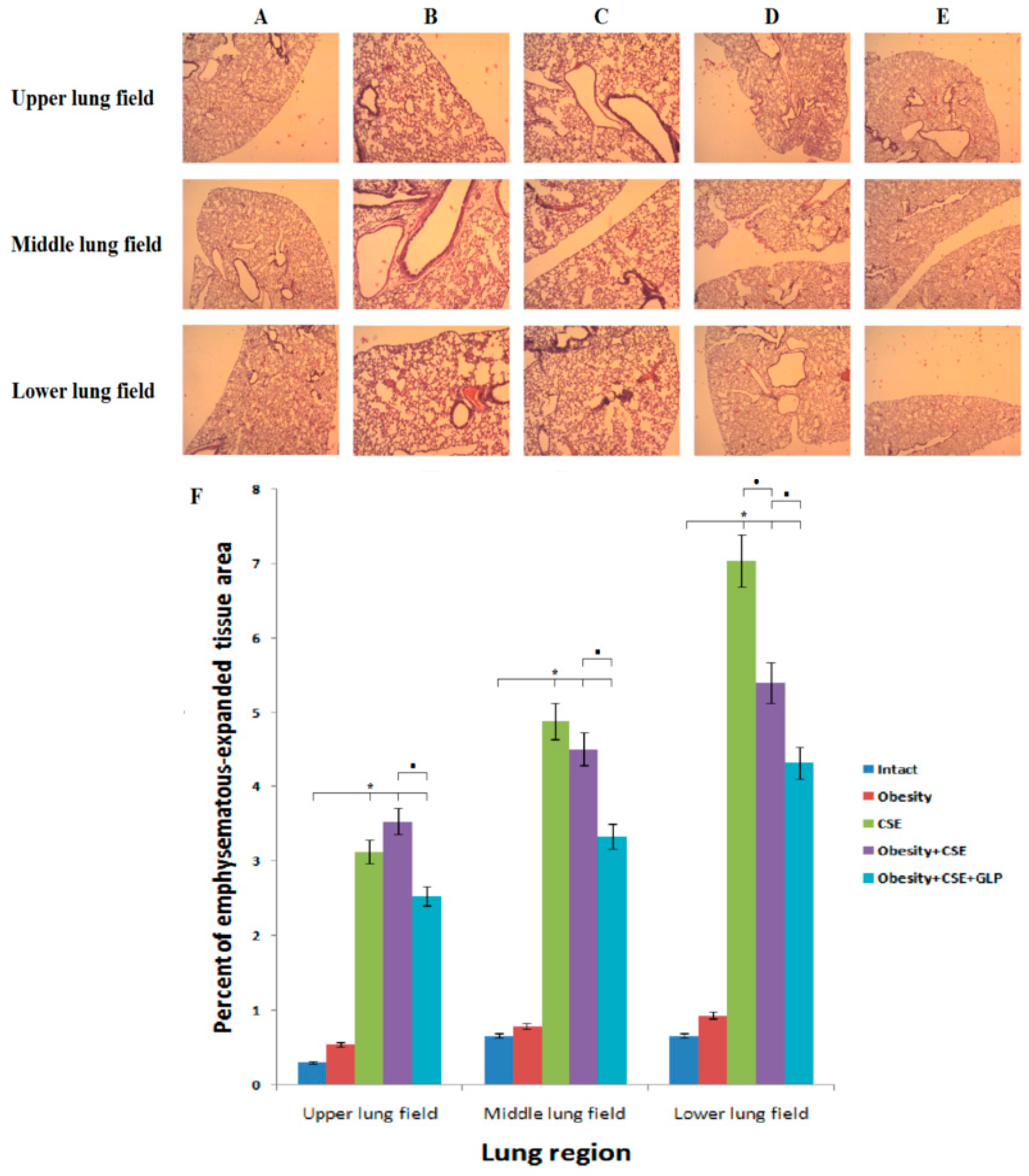
2.4. Effect of GLP-1 on Pathological Changes of the Lungs
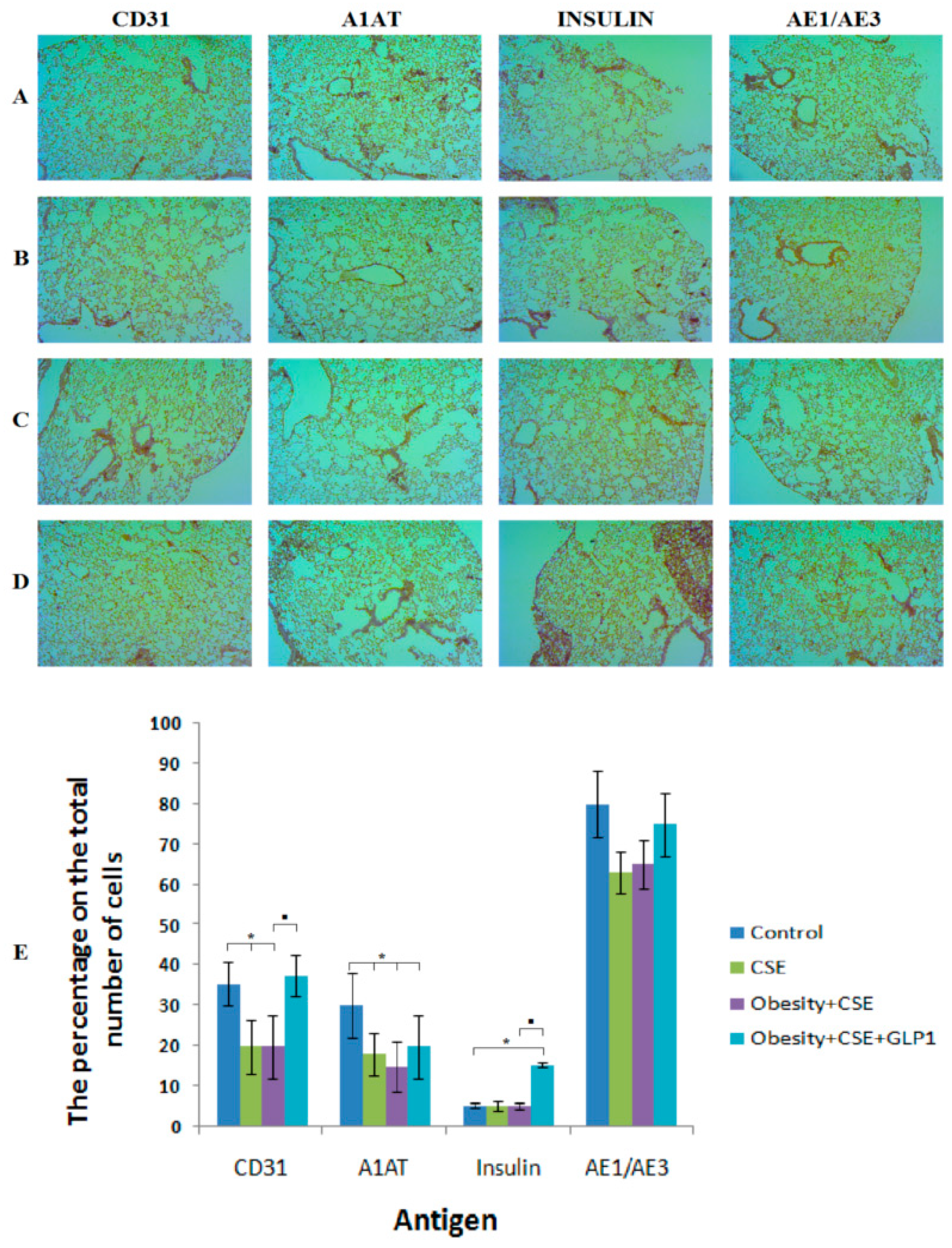
2.5. Immunohistochemical Examination of the Lungs
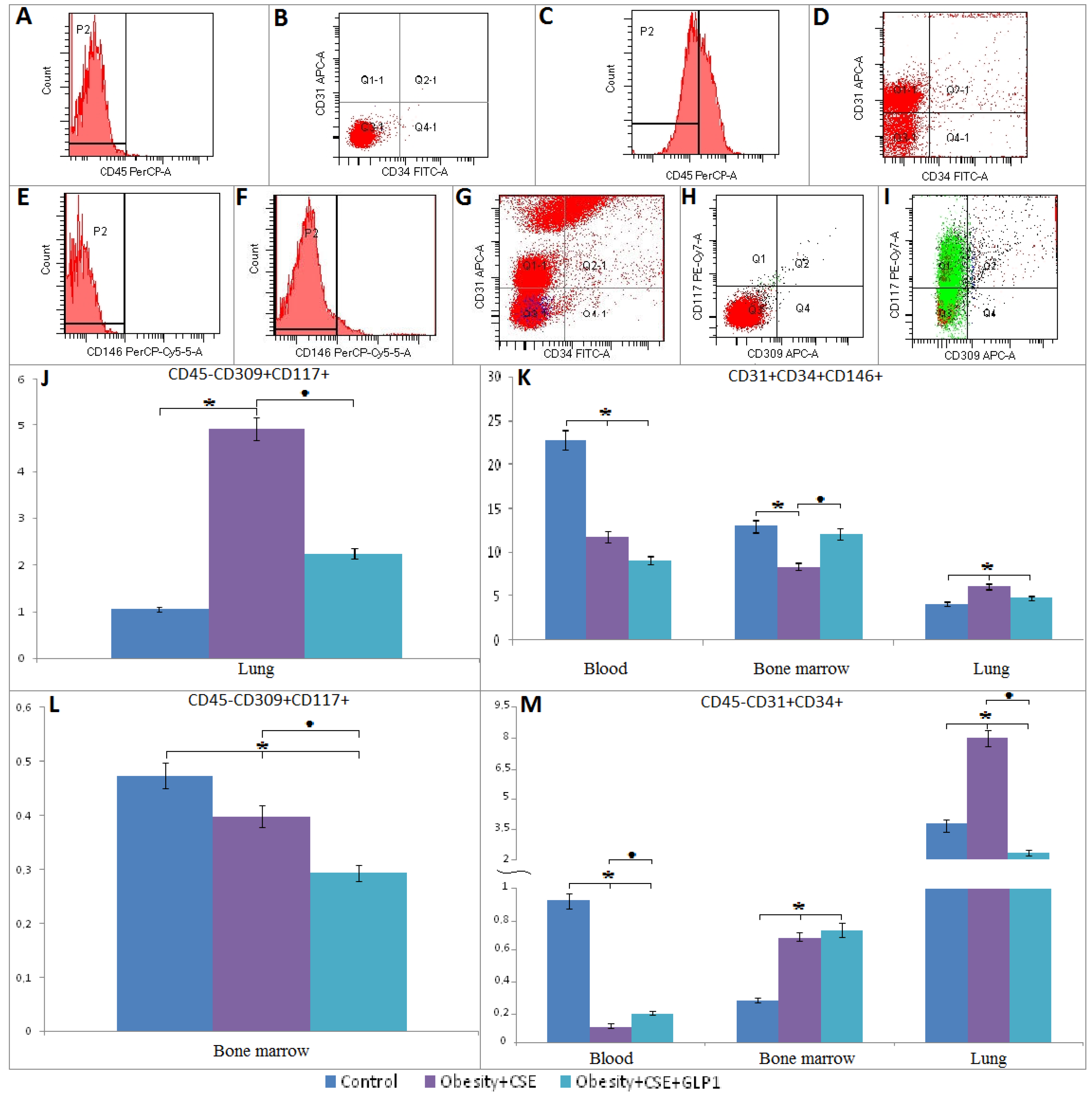
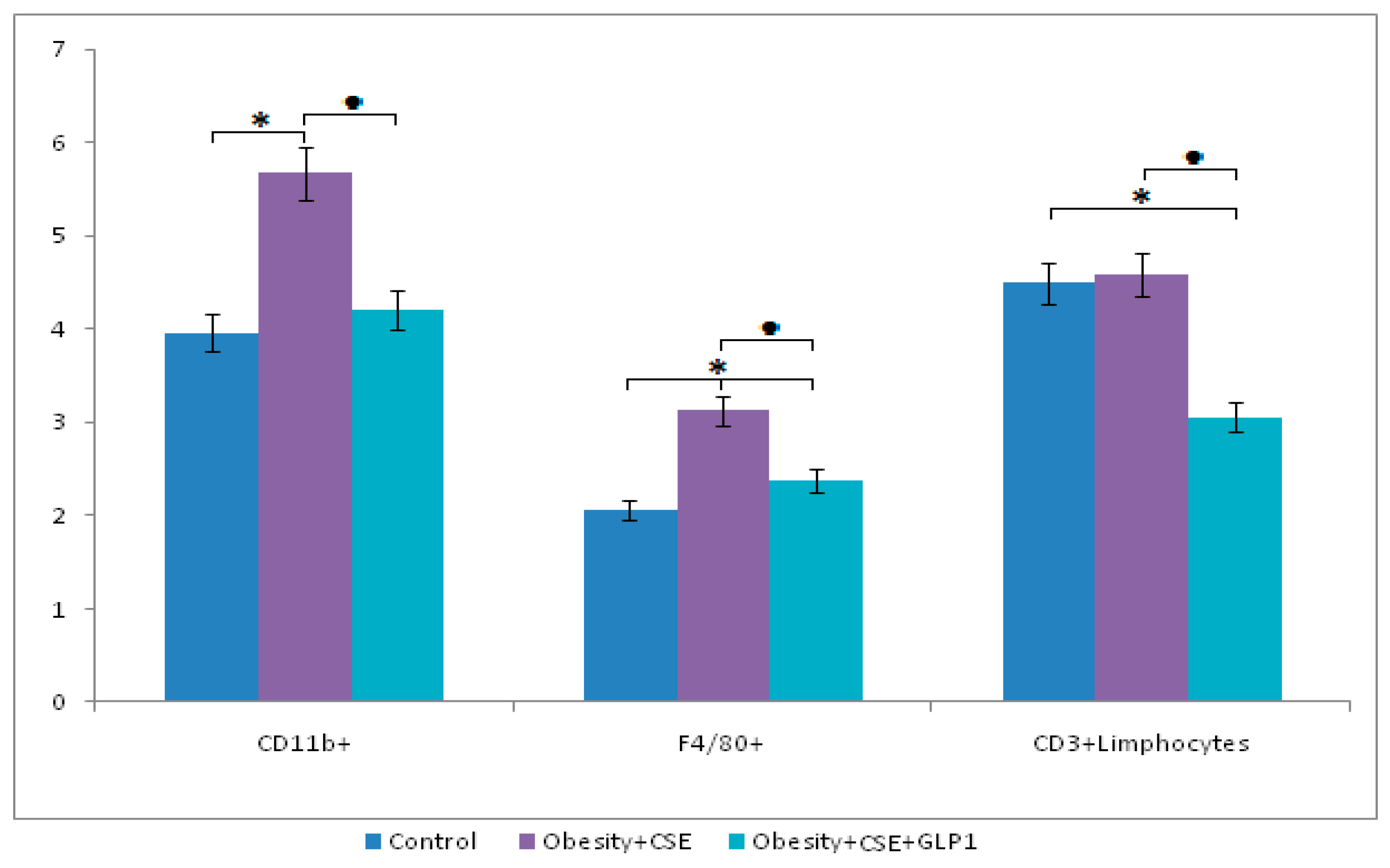
2.6. Flow Cytometric Analysis
2.6.1. Endothelial Progenitor Cells
2.6.2. Inflammatory Cells
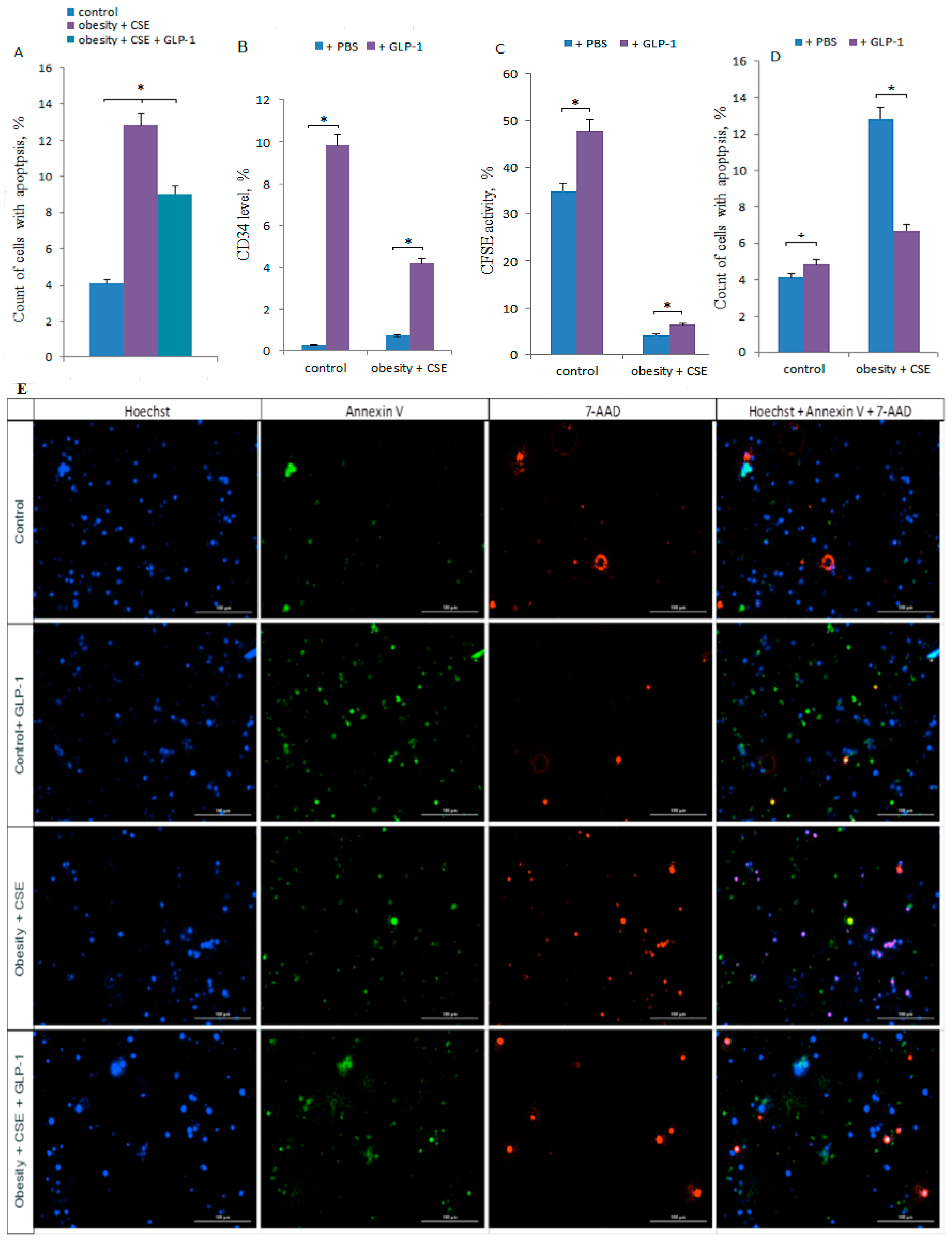
2.7. Effect of GLP-1 on Apoptosis of CD31+ Lung Cells in vivo
2.8. Effect of GLP-1 on CD31+ Lung Cells in vitro
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Obesity Experimental Design
4.3. Exposure to Cigarette Smoke Extract
4.4. Test Preparation
4.5. Distribution of Animals Into Groups
4.6. Blood Glucose Determination and Glucose Tolerance Test (GTT)
4.7. Lipid Profile Determination
4.8. Morphological Examination of Lungs
4.9. Immunohistochemical Examination of the Lungs
4.10. Flow Cytometric Analysis
4.11. Lung Tissue Dissociation, Isolation and Magnetic Separation of CD31+ Lung Endothelial Cells
4.12. Cultivation of CD31+ Lung Endothelial Cells with GLP-1
4.13. Cellular Imaging
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AUC | area under the curve |
| COPD | chronic obstructive pulmonary disease |
| CSE | cigarette smoke extract |
| EPC | endothelial progenitor cells |
| GLP-1 | glucagon-like peptide-1 |
| GTT | glucose tolerance test |
| HDL | high-density lipoproteins |
| LDL | low-density lipoproteins |
| LPS | lipopolysaccharide |
| MS | metabolic syndrome |
| MSG | monosodium glutamate |
| TG | triglycerides |
References
- Matsushita, K.; Dzau, V.J. Mesenchymal stem cells in obesity: Insights for translational applications. Lab. Investig. 2017, 97, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation, 2017. Available online: https://www.idf.org/e-library/epidemiology-research/diabetes-atlas/134-idf-diabetes-atlas-8th-edition.html (accessed on 22 January 2019).
- Grundy, S.M. Metabolic syndrome pandemic. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Díez-Manglano, J.; Barquero-Romero, J.; Almagro, P.; Cabrera, F.J.; López García, F.; Montero, L.; Soriano, J.B.; Working Group on COPD; Spanish Society of Internal Medicine. COPD patients with and without metabolic syndrome: Clinical and functional differences. Intern. Emerg. Med. 2014, 9, 419–425. [Google Scholar] [CrossRef]
- Eckel, R.H.; Kahn, S.E.; Ferrannini, E.; Goldfine, A.B.; Nathan, D.M.; Schwartz, M.W.; Smith, R.J.; Smith, S.R. Obesity and type 2 diabetes: What can be unified and what needs to be individualized? Diabetes Care 2011, 34, 1424–1430. [Google Scholar] [CrossRef] [PubMed]
- El-Badri, N.; Ghoneim, M.A. Mesenchymal stem cell therapy in diabetes mellitus: Progress and challenges. J. Nucleic Acids 2013, 2013, 194858. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, D.E.; Ciavaglia, C.E.; Neder, J.A. When obesity and chronic obstructive pulmonary disease collide. Physiological and clinical consequences. Ann. Am. Thorac. Soc. 2014, 11, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.; LeVan, T. Obesity and chronic obstructive pulmonary disease: Recent knowledge and future directions. Curr. Opin. Pulm. Med. 2017, 23, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Lamonaca, P.; Prinzi, G.; Kisialiou, A.; Cardaci, V.; Fini, M.; Russo, P. Metabolic Disorder in Chronic Obstructive Pulmonary Disease (COPD) Patients: Towards a Personalized Approach Using Marine Drug Derivatives. Mar. Drugs 2017, 15, 81. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.H.; Janaway, C.H.; Philips, B.J.; Brennan, A.L.; Baines, D.L.; Wood, D.M.; Jones, P.W. Hyperglycaemia is associated with poor outcomes in patients admitted to hospital with acute exacerbations of chronic obstructive pulmonary disease. Thorax 2006, 61, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Edwards, L.D.; Agusti, A.; Bakke, P.; Calverley, P.M.; Celli, B.; Coxson, H.O.; Crim, C.; Lomas, D.A.; Miller, B.E.; et al. Comorbidity, systemic inflammation and outcomes in the ECLIPSE cohort. Respir. Med. 2013, 107, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Parappil, A.; Depczynski, B.; Collett, P.; Marks, G.B. Effect of comorbid diabetes on length of stay and risk of death in patients admitted with acute exacerbations of COPD. Respirology 2010, 15, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Wang, R.; Wang, J.; Bunjhoo, H.; Xu, Y.; Xiong, W. Body mass index and mortality in chronic obstructive pulmonary disease: A meta-analysis. PLoS ONE 2012, 7, e43892. [Google Scholar] [CrossRef] [PubMed]
- Divo, M.; Cote, C.; de Torres, J.P.; Casanova, C.; Marin, J.M.; Pinto-Plata, V.; Zulueta, J.; Cabrera, C.; Zagaceta, J.; Hunninghake, G.; et al. Comorbidities and risk of mortality in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2012, 186, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rio, F.; Soriano, J.B.; Miravitlles, M.; Muñoz, L.; Duran-Tauleria, E.; Sánchez, G.; Sobradillo, V.; Ancochea, J. Impact of obesity on the clinical profile of a population-based sample with chronic obstructive pulmonary disease. PLoS ONE 2014, 9, e105220. [Google Scholar] [CrossRef] [PubMed]
- Piazzolla, G.; Castrovilli, A.; Liotino, V.; Vulpi, M.R.; Fanelli, M.; Mazzocca, A.; Candigliota, M.; Berardi, E.; Resta, O.; Sabbà, C.; et al. Metabolic syndrome and Chronic Obstructive Pulmonary Disease (COPD): The interplay among smoking, insulin resistance and vitamin D. PLoS ONE 2017, 12, e0186708. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, O.; Karampitsakos, T.; Barbayianni, I.; Chrysikos, S.; Xylourgidis, N.; Tzilas, V.; Bouros, D.; Aidinis, V.; Tzouvelekis, A. Metabolic Disorders in Chronic Lung Diseases. Front. Med. 2017, 4, 246. [Google Scholar] [CrossRef] [PubMed]
- Mirrakhimov, A.E. Chronic obstructive pulmonary disease and glucose metabolism: A bitter sweet symphony. Cardiovasc. Diabetol. 2012, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Ambasta, R.K.; Kohli, H.; Kumar, P. Multiple therapeutic effect of endothelial progenitor cell regulated by drugs in diabetes and diabetes related disorder. J. Transl. Med. 2017, 15, 185. [Google Scholar] [CrossRef] [PubMed]
- Peinado, V.I.; Pizarro, S.; Barberà, J.A. Pulmonary vascular involvement in COPD. Chest 2008, 134, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Agustí, A.; Barberà, J.A.; Wouters, E.F.M.; Peinado, V.; Jeffery, P.K. Lungs, Bone Marrow, and Adipose Tissue. A Network Approach to the Pathobiology of Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2013, 188, 1396–1406. [Google Scholar] [CrossRef] [PubMed]
- Janssen, W.J.; Yunt, Z.X.; Muldrow, A.; Kearns, M.T.; Kloepfer, A.; Barthel, L.; Bratton, D.L.; Bowler, R.P.; Henson, P.M. Circulating Hematopoietic Progenitor Cells are Decreased in COPD. COPD 2014, 11, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; She, M.; Xu, M.; Chen, H.; Li, J.; Chen, X.; Zheng, D.; Liu, J.; Chen, S.; Zhu, J.; et al. GLP-1 treatment protects endothelial cells from oxidative stress-induced autophagy and endothelial dysfunction. Int. J. Biol. Sci. 2018, 14, 1696–1708. [Google Scholar] [CrossRef] [PubMed]
- Körner, M.; Stöckli, M.; Waser, B.; Reubi, J.C. GLP-1 receptor expression in human tumors and human normal tissues: Potential for in vivo targeting. J. Nucl. Med. 2007, 48, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.V.; Linderholm, A.; Haczku, A.; Kenyon, N. Glucagon-like peptide 1: A potential anti-inflammatory pathway in obesity-related asthma. Pharmacol. Ther. 2017, 180, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yi, H.; Zhao, C.; Zhang, Y.; Zhu, L.; Liu, B.; He, P.; Zhou, M. Glucagon-like peptide-1 receptor (GLP-1R) signaling ameliorates dysfunctional immunity in COPD patients. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 3191–3202. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Jun, H.S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediators Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef] [PubMed]
- Sjøberg, K.A.; Rattigan, S.; Jeppesen, J.F.; Lundsgaard, A.M.; Holst, J.J.; Kiens, B. Differential effects of glucagon-like peptide-1 on microvascular recruitment and glucose metabolism in short- and long-term insulin resistance. J. Physiol. 2015, 593, 2185–2198. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pakhomova, A.V.; Krupin, V.A.; Pershina, O.V.; Pan, E.S.; Ermolaeva, L.A.; Vaizova, O.E.; Rybalkina, O.Y.; Dygai, A.M. Response of Inflammatory Mediators, Extracellular Matrix Proteins and Stem and Progenitor Cells to Emphysema. Bull. Exp. Biol. Med. 2016, 161, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pakhomova, A.V.; Ermakova, N.N.; Pershina, O.V.; Krupin, V.A.; Pan, E.S.; Kudryashova, A.I.; Ermolaeva, L.A.; Dygai, A.M. Role of Tissue-Specific Stem and Progenitor Cells in the Regeneration of the Pancreas and Testicular Tissue in Diabetic Disorders. Bull. Exp. Biol. Med. 2016, 3, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Matysková, R.; Maletínská, L.; Maixnerová, J.; Pirník, Z.; Kiss, A.; Zelezná, B. Comparison of the obesity phenotypes related to monosodium glutamate effect on arcuate nucleus and/or the high fat diet feeding in C57BL/6 and NMRI mice. Physiol. Res. 2008, 57, 727–734. [Google Scholar] [PubMed]
- Sasaki, Y.; Suzuki, W.; Shimada, T.; Iizuka, S.; Nakamura, S.; Nagata, M.; Fujimoto, M.; Tsuneyama, K.; Hokao, R.; Miyamoto, K.; et al. Dose dependent development of diabetes mellitus and non-alcoholic steatohepatitis in monosodium glutamate-induced obese mice. Life Sci. 2009, 85, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Salisu, N.; Aliyu, F.S.; Garba, H.; Garba, M.H. Anaemogenic, Obesogenic and Thermogenic Potentials of Graded Doses of Monosodium Glutamate Sub-Acutely Fed to Experimental Wistar Rats. Curr. Clin. Pharmacol. 2018, 13, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pakhomova, A.V.; Epanchintsev, A.A.; Stronin, O.V.; Ermakova, N.N.; Pershina, O.V.; Ermolaeva, L.A.; Krupin, V.A.; Kudryashova, A.I.; Zhdanov, V.V.; et al. Role of β Cell Precursors in the Regeneration of Insulin-Producing Pancreatic β Cells under the Influence of Glucagon-Like Peptide 1. Bull. Exp. Biol. Med. 2018, 165, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.F.; Tracy, R.P.; Parikh, M.A.; Hoffman, E.A.; Shimbo, D.; Austin, J.H.; Smith, B.M.; Hueper, K.; Vogel-Claussen, J.; Lima, J.; et al. Endothelial progenitor cells in chronic obstructive pulmonary disease and emphysema. PLoS ONE 2017, 12, e0173446. [Google Scholar] [CrossRef] [PubMed]
- Salter, B.; Sehmi, R. The role of bone marrow-derived endothelial progenitor cells and angiogenic responses in chronic obstructive pulmonary disease. J. Thorac. Dis. 2017, 9, 2168–2177. [Google Scholar] [CrossRef] [PubMed]
- Burger, P.E.; Coetzee, S.; McKeehan, W.L.; Kan, M.; Cook, P.; Fan, Y.; Suda, T.; Hebbel, R.P.; Novitzky, N.; Muller, W.A.; et al. Fibroblast growth factor receptor-1 is expressed by endothelial progenitor cells. Blood 2002, 100, 3527–3535. [Google Scholar] [CrossRef] [PubMed]
- Rogliani, P.; Calzetta, L.; Capuani, B.; Facciolo, F.; Cazzola, M.; Lauro, D.; Matera, M.G. Glucagon-Like Peptide 1 Receptor: A Novel Pharmacological Target for Treating Human Bronchial Hyperresponsiveness. Am. J. Respir. Cell Mol. Biol. 2016, 55, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Viby, N.E.; Isidor, M.S.; Buggeskov, K.B.; Poulsen, S.S.; Hansen, J.B.; Kissow, H. Glucagon-like peptide-1 (GLP-1) reduces mortality and improves lung function in a model of experimental obstructive lung disease in female mice. Endocrinology 2013, 154, 4503–4511. [Google Scholar] [CrossRef] [PubMed]
- Mendivil, C.O.; Koziel, H.; Brain, J.D. Metabolic hormones, apolipoproteins, adipokines, and cytokines in the alveolar lining fluid of healthy adults: Compartmentalization and physiological correlates. PLoS ONE 2015, 10, e0123344. [Google Scholar] [CrossRef] [PubMed]
- Pierrou, S.; Broberg, P.; O’Donnell, R.A.; Pawłowski, K.; Virtala, R.; Lindqvist, E.; Richter, A.; Wilson, S.J.; Angco, G.; Möller, S.; et al. Expression of genes involved in oxidative stress responses in airway epithelial cells of smokers with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2007, 175, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Chilosi, M.; Poletti, V.; Rossi, A. The pathogenesis of COPD and IPF: Distinct horns of the same devil? Respir. Res. 2012, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Hull, D.; Guo, K.; Barton, D.; Hazlehurst, J.M.; Gathercole, L.L.; Nasiri, M.; Yu, J.; Gough, S.C.; Newsome, P.N.; et al. Glucagon-like peptide 1 decreases lipotoxicity in non-alcoholic steatohepatitis. J. Hepatol. 2016, 64, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Finelli, C.; Sommella, L.; Gioia, S.; La Sala, N.; Tarantino, G. Should visceral fat be reduced to increase longevity? Ageing Res. Rev. 2013, 12, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; He, L.; Yan, M.Y.; Zhao, R.Q.; Li, B.; Wang, F.; Yang, Y.; Yu, H.P. Overexpression of GLP-1 receptors suppresses proliferation and cytokine release by airway smooth muscle cells of patients with chronic obstructive pulmonary disease via activation of ABCA1. Mol. Med. Rep. 2017, 16, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.P.; Poon, T.K.-Y.; Smith, G.C. Effects of Monosodium Glutamate Administration in the Neonatal Period on the Diabetic Syndrome in KK Mice. Diabetologia 1976, 12, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Nawa, A.; Fujita-Hamabe, W.; Tokuyama, S. Altered intestinal P-glycoprotein expression levels in a monosodium glutamate-induced obese mouse model. Life Sci. 2011, 89, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, D.C.; Sinzato, Y.K.; Bueno, A.; Dallaqua, B.; Lima, P.H.; Calderon, I.M.P.; Rudge, M.V.C.; Campos, K.E. Metabolic Profile and Genotoxicity in Obese Rats Exposed to Cigarette Smoke. Obesity 2013, 21, 1569–1601. [Google Scholar] [CrossRef] [PubMed]
- Campos, K.E.; Sinzato, Y.K.; de Paula Pimenta, W.; Rudge, M.V.; Damasceno, D.C. Effect of maternal obesity on the diabetes developmental in adult rat offspring. Life Sci. 2007, 81, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hanaoka, M.; Chen, P.; Droma, Y.; Voelkel, N.F.; Kubo, K. Protective effect of beraprost sodium, a stable prostacyclin analog, in the development of cigarette smoke extract-induced. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L648–L656. [Google Scholar] [CrossRef] [PubMed]
- Hanaoka, M.; Hanaoka, P.; Chen, Y.; Agatsuma, T.; Kitaguchi, Y.; Voelkel, N.F.; Kubo, K. Carbocisteine protects against emphysema induced by cigarette smoke extract in rats. Chest 2011, 139, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Chen, P.; Chen, Y.; He, S.D.; Ye, J.R.; Zhang, H.L.; Cao, J. Comparison between cigarette smoke-induced emphysema and cigarette smoke extract-induced emphysema. Tob. Induc. Dis. 2015, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Pacini, G.; Ahrén, M.; Ahrén, B. Reappraisal of the intravenous glucose tolerance index for a simple assessment of insulin sensitivity in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1316–R1324. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, H.; Majumdar, A.; Ito, S.; Alencar, A.M.; Suki, B. Quantitative characterization of airspace enlargement in emphysema. J. Appl. Physiol. 2006, 100, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Barrutia, A.; Ceresa, M.; Artaechevarria, X.; Montuenga, L.M.; Ortiz-de-Solorzano, C. Quantification of lung damage in an elastase-induced mouse model of emphysema. Int. J. Biomed. Imaging 2012, 2012, 734734. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Bartolák-Suki, E.; Parameswaran, H.; Hamakawa, H.; Suki, B. Scale dependence of structure-function relationship in the emphysematous mouse lung. Front. Physiol. 2015, 6, 146. [Google Scholar] [CrossRef] [PubMed]
- Skurikhin, E.G.; Pershina, O.V.; Reztsova, A.M.; Ermakova, N.N.; Khmelevskaya, E.S.; Krupin, V.A.; Stepanova, I.E.; Artamonov, A.V.; Bekarev, A.A.; Madonov, P.G.; et al. Modulation of Bleomycin-Induced Lung Fibrosis by Pegylated Hyaluronidase and Dopamine Receptor Antagonist in Mice. PLoS ONE 2015, 10, e0125065. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skurikhin, E.G.; Pershina, O.V.; Pakhomova, A.V.; Pan, E.S.; Krupin, V.A.; Ermakova, N.N.; Vaizova, O.E.; Pozdeeva, A.S.; Zhukova, M.A.; Skurikhina, V.E.; et al. Endothelial Progenitor Cells as Pathogenetic and Diagnostic Factors, and Potential Targets for GLP-1 in Combination with Metabolic Syndrome and Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2019, 20, 1105. https://doi.org/10.3390/ijms20051105
Skurikhin EG, Pershina OV, Pakhomova AV, Pan ES, Krupin VA, Ermakova NN, Vaizova OE, Pozdeeva AS, Zhukova MA, Skurikhina VE, et al. Endothelial Progenitor Cells as Pathogenetic and Diagnostic Factors, and Potential Targets for GLP-1 in Combination with Metabolic Syndrome and Chronic Obstructive Pulmonary Disease. International Journal of Molecular Sciences. 2019; 20(5):1105. https://doi.org/10.3390/ijms20051105
Chicago/Turabian StyleSkurikhin, Evgenii Germanovich, Olga Victorovna Pershina, Angelina Vladimirovna Pakhomova, Edgar Sergeevich Pan, Vyacheslav Andreevich Krupin, Natalia Nicolaevna Ermakova, Olga Evgenevna Vaizova, Anna Sergeevna Pozdeeva, Mariia Andreevna Zhukova, Viktoriia Evgenevna Skurikhina, and et al. 2019. "Endothelial Progenitor Cells as Pathogenetic and Diagnostic Factors, and Potential Targets for GLP-1 in Combination with Metabolic Syndrome and Chronic Obstructive Pulmonary Disease" International Journal of Molecular Sciences 20, no. 5: 1105. https://doi.org/10.3390/ijms20051105
APA StyleSkurikhin, E. G., Pershina, O. V., Pakhomova, A. V., Pan, E. S., Krupin, V. A., Ermakova, N. N., Vaizova, O. E., Pozdeeva, A. S., Zhukova, M. A., Skurikhina, V. E., Grimm, W.-D., & Dygai, A. M. (2019). Endothelial Progenitor Cells as Pathogenetic and Diagnostic Factors, and Potential Targets for GLP-1 in Combination with Metabolic Syndrome and Chronic Obstructive Pulmonary Disease. International Journal of Molecular Sciences, 20(5), 1105. https://doi.org/10.3390/ijms20051105