Hydrophilic Linear Peptide with Histidine and Lysine Residues as a Key Factor Affecting Antifungal Activity

 , and
, and
Abstract
1. Introduction
2. Results and Discussion
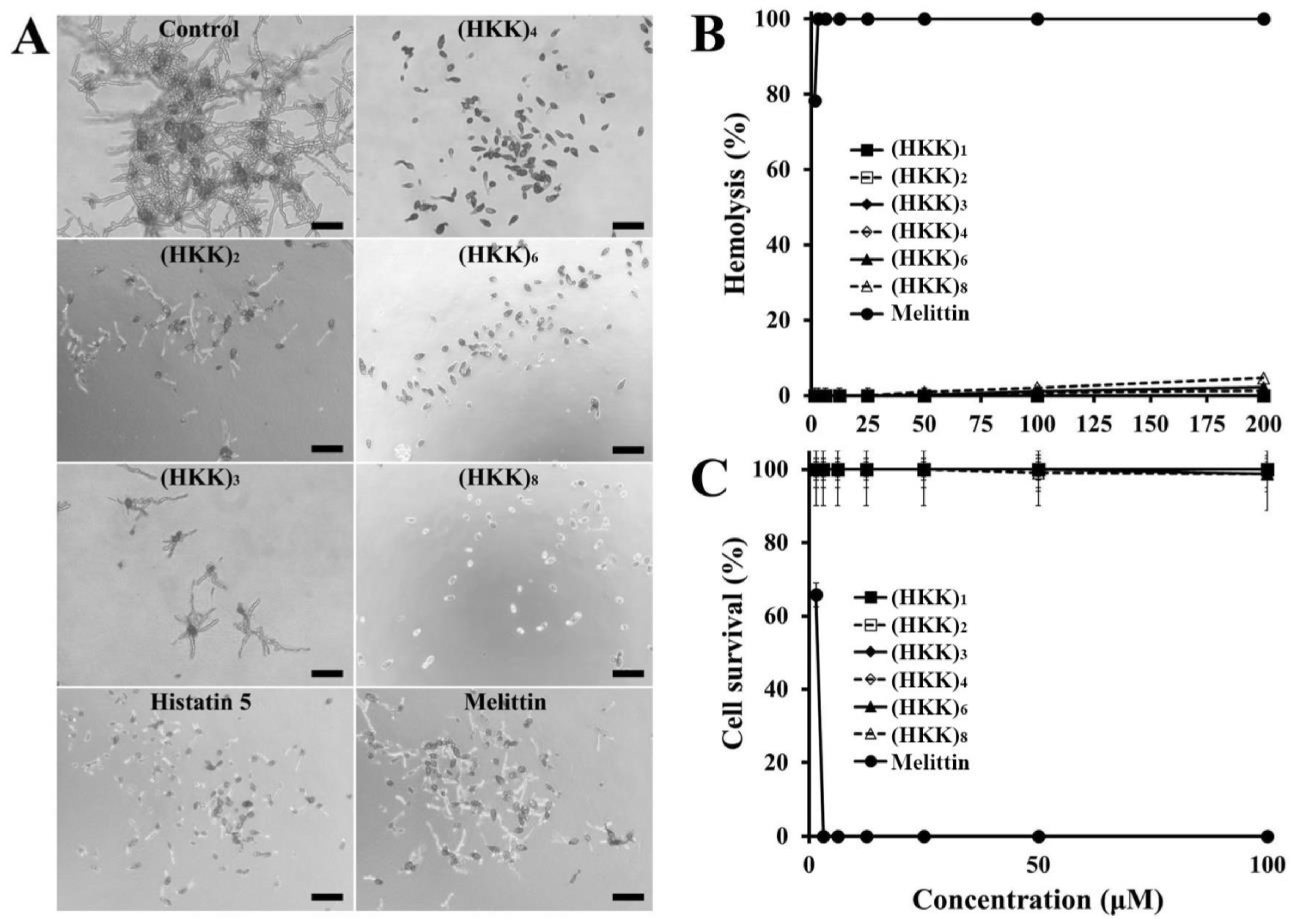
2.1. Design, Antifungal Activity, and Cytotoxic Effects of Peptides
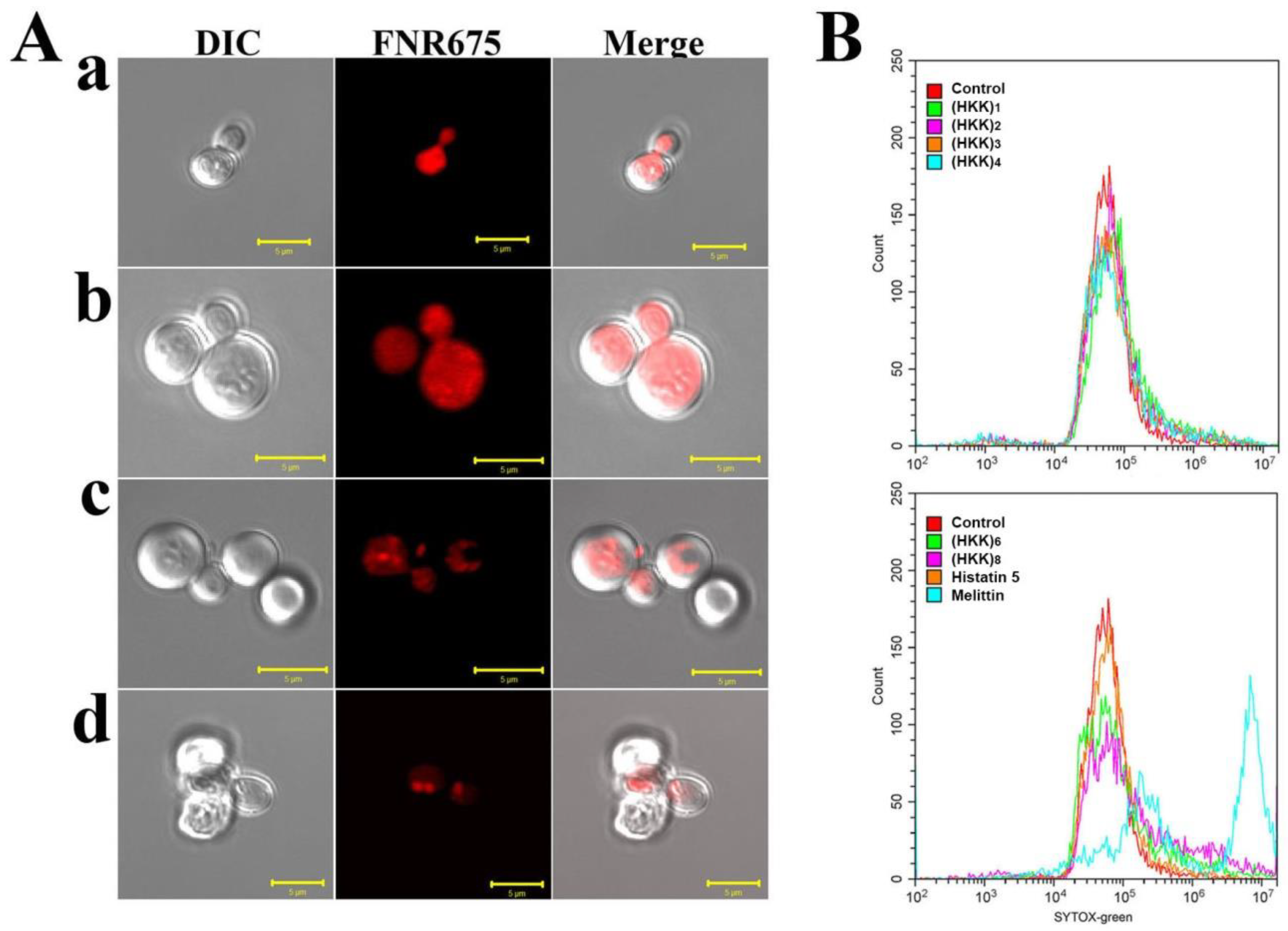
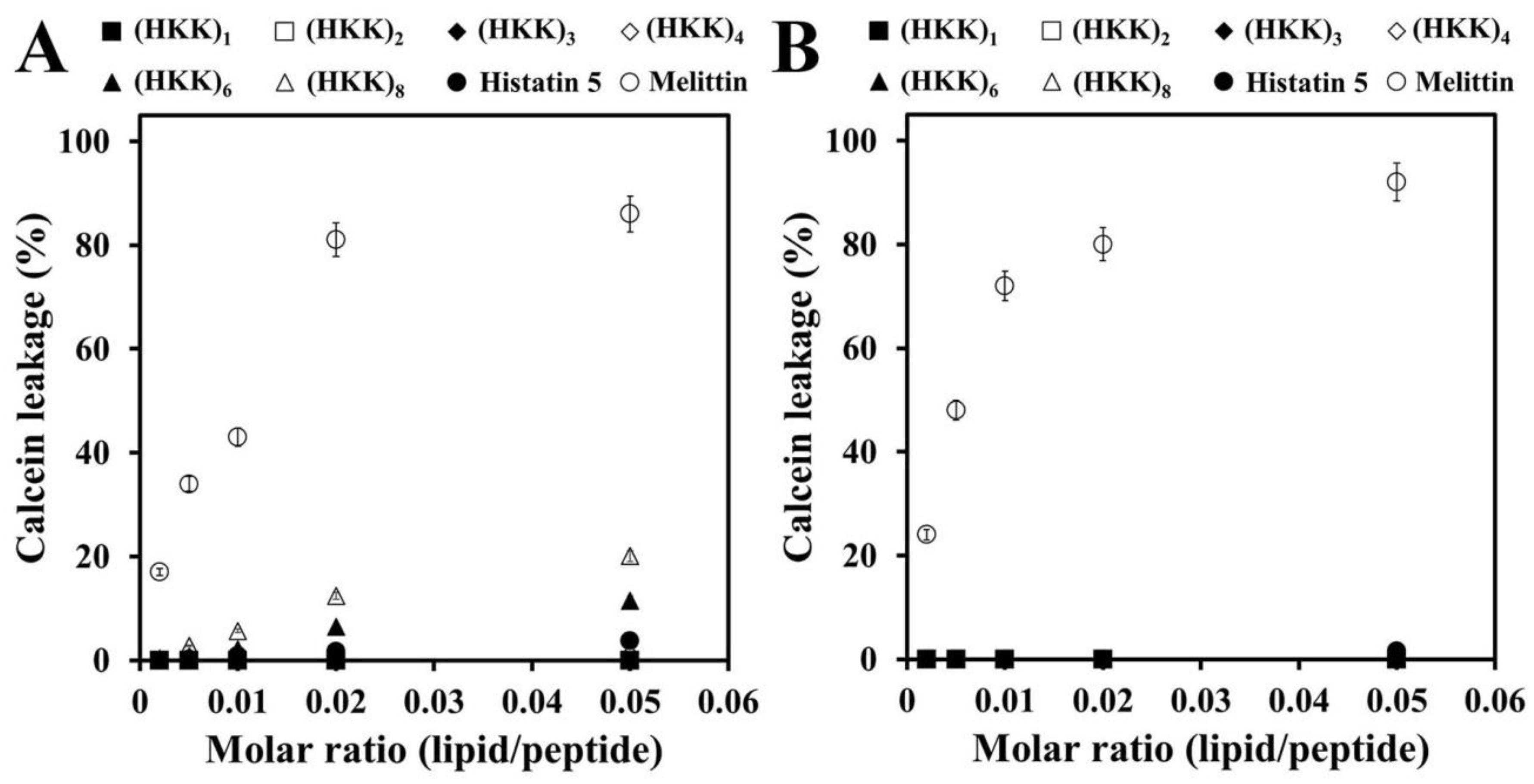
2.2. Localization and Membrane-Permeable Effects of HKK Peptides
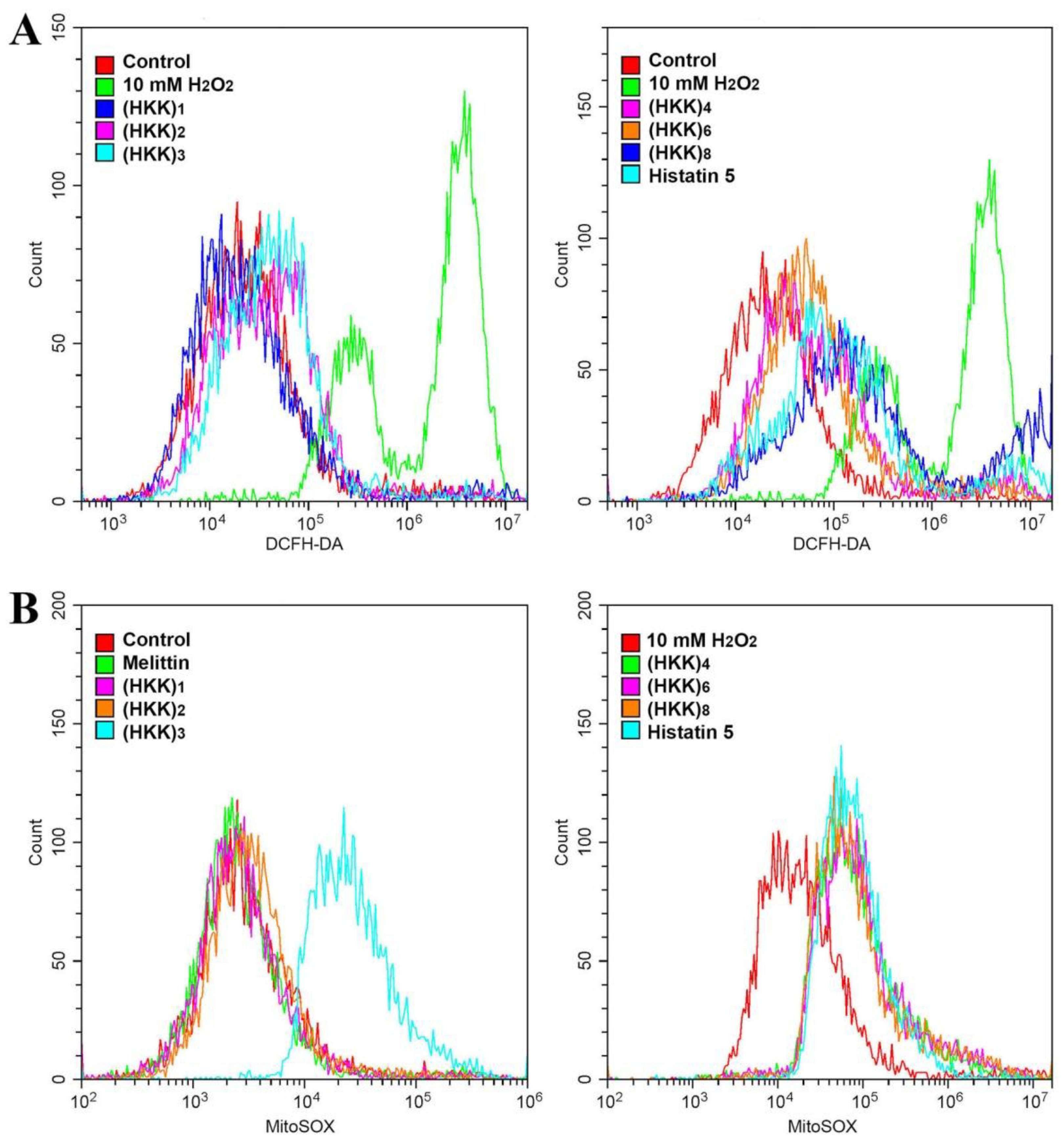
2.3. Reactive Oxygen Species (ROS) Production via Mitochondrial Damage
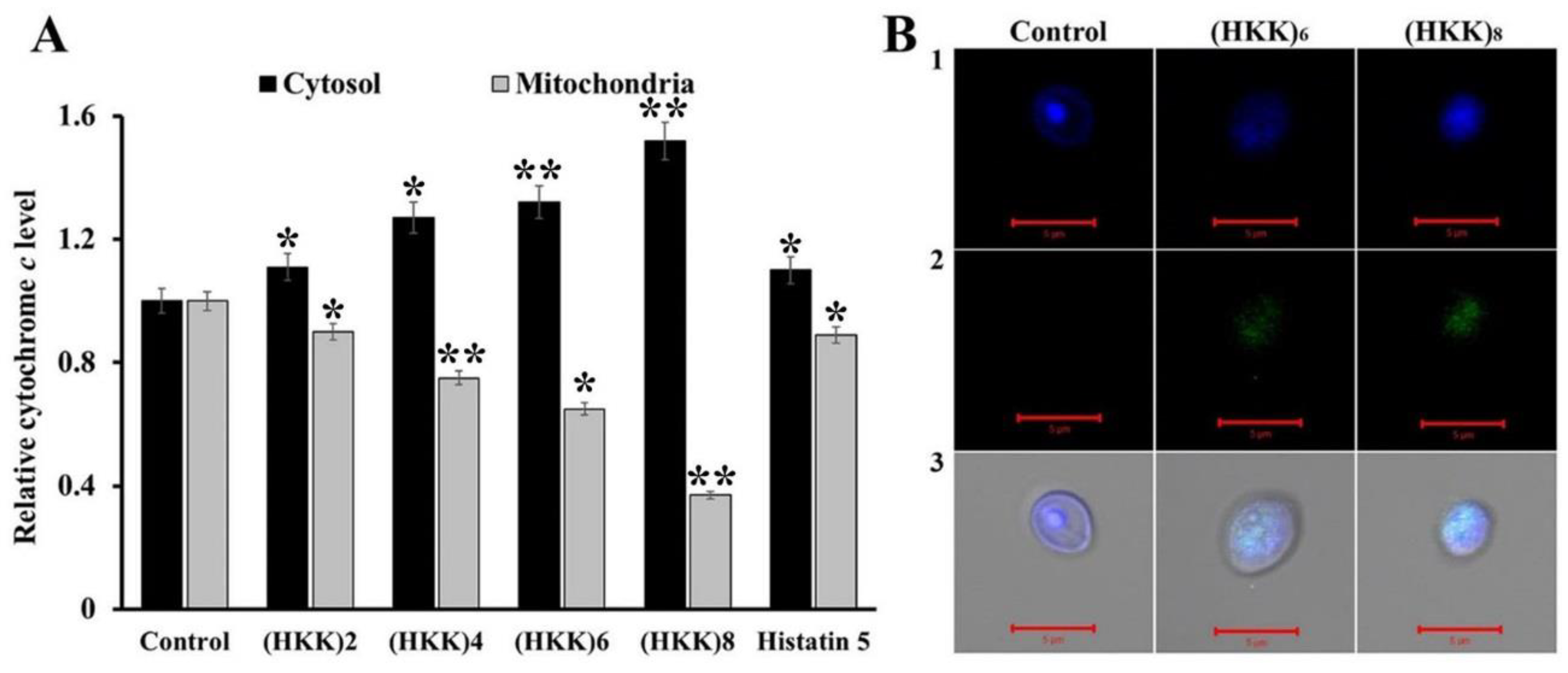
2.4. Apoptosis Induction
2.5. Morphological Alterations in C. albicans Cells with HKK Peptides
3. Materials and Methods
3.1. Materials
3.2. Peptide Synthesis
3.3. Antifungal Assay
3.4. Hemolysis and Cytotoxicity
3.5. CLSM
3.6. SYTOX Green Uptake
3.7. Calcein Leakage
3.8. Measurement of Intracellular ROS and Mitochondrial SOX
3.9. Release of Cytochrome c
3.10. TUNEL Assay
3.11. SEM
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 3004404. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, P.; Dromer, F.; Lortholary, O.; Dannaoui, E. Efficacy of amphotericin B in combination with flucytosine against flucytosine-susceptible or flucytosine-resistant isolates of Cryptococcus neoformans during disseminated murine cryptococcosis. Antimicrob. Agents Chemother. 2006, 50, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Park, Y.; Hahm, K.S. The role of antimicrobial peptides in preventing multidrug-resistant bacterial infections and biofilm formation. Int. J. Mol. Sci. 2011, 12, 5971–5992. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.H. Tryptophan-rich and proline-Rich antimicrobial peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Augusto, M.T.; Felício, M.R.; Hollmann, A.; Franco, O.L.; Gonçalves, S.; Santos, N.C. Designing improved active peptides for therapeutic approaches against infectious diseases. Biotechnol. Adv. 2018, 36, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Hung, Y.J.; Kim, I.R.; Lee, Y.; Kim, Y.M.; Jang, M.K.; Lee, J.R. Functional characterization of thioredoxin h type 5 with antimicrobial activity from Arabidopsis thaliana. Biotechnol. Bioprocess Eng. 2017, 22, 129–135. [Google Scholar] [CrossRef]
- Lee, J.K.; Park, S.C.; Hahm, K.S.; Park, Y. A helix-PXXP-helix peptide with antibacterial activity without cytotoxicity against MDRPA-infected mice. Biomaterials 2014, 35, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Lau, Q.Y.; Ng, F.M.; Cheong, J.W.; Yap, Y.Y.; Tan, Y.Y.; Jureen, R.; Hill, J.; Chia, C.S. Discovery of an ultra-short linear antibacterial tetrapeptide with anti-MRSA activity from a structure-activity relationship study. Eur. J. Med. Chem. 2015, 105, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Lequin, O.; Piesse, C.; Goasdoué, N.; Foulon, T.; Nicolas, P.; Ladram, A. Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide. J. Biol. Chem. 2010, 285, 16880–16892. [Google Scholar] [CrossRef] [PubMed]
- Vylkova, S. Environmental pH modulation by pathogenic fungi as a strategy to conquer the host. PLoS Pathog. 2017, 13, e1006149. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Volk, M.; Shah, K.; Clerkin, K.; Liang, J.F. Constructing bioactive peptides with pH-dependent activities. Peptides 2009, 30, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.E.; Daum, J. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef] [PubMed]
- Koppenol, W.H. The Haber-Weiss cycle—70 years later. Redox Rep. 2001, 6, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C.; Almeida, J.I.; Lima, I.S.; Kapitão, A.S.; Gozzelino, R. Iron metabolism and the inflammatory response. IUBMB Life 2017, 69, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Falcone, C.; Mazzoni, C. External and internal triggers of cell death in yeast. Cell. Mol. Life Sci. 2016, 73, 2237–2250. [Google Scholar] [CrossRef] [PubMed]
- Ramsdale, M. Programmed cell death in pathogenic fungi. Biochim. Biophys. Acta 2008, 1783, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Austriaco, N. Aging and cell death in the other yeasts, Schizosaccharomyces pombe and Candida albicans. FEMS Yeast Res. 2014, 14, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed]
- Vandenbosch, D.; Braeckmans, K.; Nelis, H.J.; Coenye, T. Fungicidal activity of miconazole against Candida spp. biofilms. J. Antimicrob. Chemother. 2010, 65, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Lee, D.G. Reactive oxygen species modulate itraconazole-induced apoptosis via mitochondrial disruption in Candida albicans. Free Radic. Res. 2018, 52, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Manoharlal, R.; Puri, N.; Prasad, R. Antifungal curcumin induces reactive oxygen species and triggers an early apoptosis but prevents hyphae development by targeting the global repressor TUP1 in Candida albicans. Biosci. Rep. 2010, 30, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, D.G. Novel antifungal mechanism of resveratrol: Apoptosis inducer in Candida albicans. Curr. Microbiol. 2015, 70, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.B.; Ban, F.F.; Li, H.B.; Qian, P.P.; Shen, Q.S.; Zhao, Y.Y.; Mo, H.Z.; Zhou, X. Thymol induces conidial apoptosis in Aspergillus flavus via stimulating K+ eruption. J. Agric. Food Chem. 2018, 66, 8530–8536. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liao, K.; Hang, C.; Wang, D. Honokiol induces reactive oxygen species-mediated apoptosis in Candida albicans through mitochondrial dysfunction. PLoS ONE. 2017, 12, e0172228. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hwang, J.S.; Hwang, I.S.; Cho, J.; Lee, E.; Kim, Y.; Lee, D.G. Coprisin-induced antifungal effects in Candida albicans correlate with apoptotic mechanisms. Free Radic. Biol. Med. 2012, 52, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Ogawa, K.; Fukuda, M.; Ohori, M.; Kanaoka, T.; Tanaka, T.; Taniguchi, M.; Sagehashi, Y. Rice defensin OsAFP1 is a new drug candidate against human pathogenic fungi. Sci. Rep. 2018, 8, 11434. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kim, I.R.; Kim, J.Y.; Lee, Y.; Kim, E.J.; Jung, J.H.; Jung, Y.J.; Jang, M.K.; Lee, J.R. Molecular mechanism of Arabidopsis thaliana profilins as antifungal proteins. Biochim. Biophys. Acta 2018, 1862, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Cheong, M.S.; Kim, E.J.; Kim, J.H.; Chi, Y.H.; Jang, M.K. Antifungal effect of Arabidopsis SGT1 proteins via mitochondrial reactive oxygen species. J. Agric. Food Chem. 2017, 65, 8340–8347. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Jung, Y.J.; Lee, Y.; Kim, I.R.; Seol, M.A.; Kim, E.J.; Jang, M.K.; Lee, J.R. Functional characterization of the Arabidopsis universal stress protein AtUSP with an antifungal activity. Biochem. Biophys. Res. Commun. 2017, 486, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Hwang, J.S.; Lee, D.G. Identification of a novel antimicrobial peptide, scolopendin 1, derived from centipede Scolopendra subspinipes mutilans and its antifungal mechanism. Insect Mol. Biol. 2014, 23, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kim, J.Y.; Shin, S.O.; Jeong, C.Y.; Kim, M.H.; Shin, S.Y.; Cheong, G.W.; Park, Y.; Hahm, K.S. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 2006, 343, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Forsman, J.; Woodward, C.E. Molecular simulations of melittin-induced membrane pores. J. Phys. Chem. B 2017, 121, 10209–10214. [Google Scholar] [CrossRef] [PubMed]
- Pino-Angeles, A.; Lazaridis, T. Effects of peptide charge, orientation, and concentration on melittin transmembrane pores. Biophys. J. 2018, 114, 2865–2874. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC (µM) a | |||||||
|---|---|---|---|---|---|---|---|
| (HKK)1 | (HKK)2 | (HKK)3 | (HKK)4 | (HKK)6 | (HKK)8 | Melittin | |
| A. flavus | 256 | 64 | 16 | 8 | 4 | 2 | 8 |
| C. gloeosporioides | 1024 | 16 | 8 | 2 | 1 | 1 | 16 |
| C. albicans | 1024 | 64 | 64 | 32 | 16 | 8 | 8 |
| C. albicans 14001 b | 1024 | 64 | 64 | 32 | 16 | 8 | 8 |
| C. albicans 14004 b | 1024 | 64 | 64 | 32 | 16 | 8 | 8 |
| C. albicans 14007 b | 1024 | 64 | 64 | 32 | 16 | 8 | 8 |
| C. albicans 14020 b | 1024 | 64 | 64 | 32 | 16 | 8 | 8 |
| C. krusei | 512 | 64 | 16 | 4 | 4 | 4 | 8 |
| C. parasilopsis | 1024 | 64 | 32 | 32 | 16 | 8 | 8 |
| F. graminearum | 512 | 64 | 32 | 16 | 8 | 4 | 8 |
| F. moniliforme | 256 | 32 | 16 | 4 | 2 | 2 | 8 |
| F. oxysporum | 256 | 64 | 32 | 4 | 2 | 2 | 8 |
| F. solani | 256 | 32 | 16 | 4 | 2 | 1 | 8 |
| T. rubrum | 512 | 64 | 32 | 16 | 8 | 2 | 8 |
| T. virens | 512 | 32 | 16 | 16 | 8 | 4 | 32 |
| T. viride | 256 | 64 | 16 | 4 | 2 | 1 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-C.; Kim, J.-Y.; Kim, E.-J.; Cheong, G.-W.; Lee, Y.; Choi, W.; Lee, J.R.; Jang, M.-K. Hydrophilic Linear Peptide with Histidine and Lysine Residues as a Key Factor Affecting Antifungal Activity. Int. J. Mol. Sci. 2018, 19, 3781. https://doi.org/10.3390/ijms19123781
Park S-C, Kim J-Y, Kim E-J, Cheong G-W, Lee Y, Choi W, Lee JR, Jang M-K. Hydrophilic Linear Peptide with Histidine and Lysine Residues as a Key Factor Affecting Antifungal Activity. International Journal of Molecular Sciences. 2018; 19(12):3781. https://doi.org/10.3390/ijms19123781
Chicago/Turabian StylePark, Seong-Cheol, Jin-Young Kim, Eun-Ji Kim, Gang-Won Cheong, Yongjae Lee, Wonkyun Choi, Jung Ro Lee, and Mi-Kyeong Jang. 2018. "Hydrophilic Linear Peptide with Histidine and Lysine Residues as a Key Factor Affecting Antifungal Activity" International Journal of Molecular Sciences 19, no. 12: 3781. https://doi.org/10.3390/ijms19123781
APA StylePark, S.-C., Kim, J.-Y., Kim, E.-J., Cheong, G.-W., Lee, Y., Choi, W., Lee, J. R., & Jang, M.-K. (2018). Hydrophilic Linear Peptide with Histidine and Lysine Residues as a Key Factor Affecting Antifungal Activity. International Journal of Molecular Sciences, 19(12), 3781. https://doi.org/10.3390/ijms19123781