New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets


Abstract
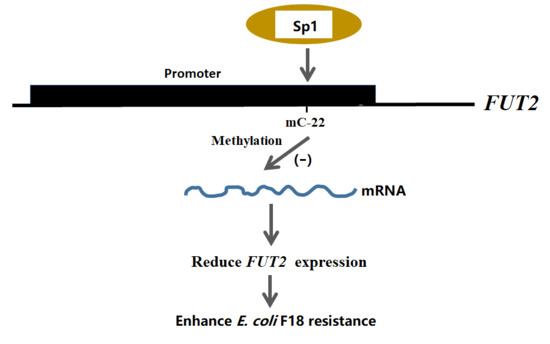
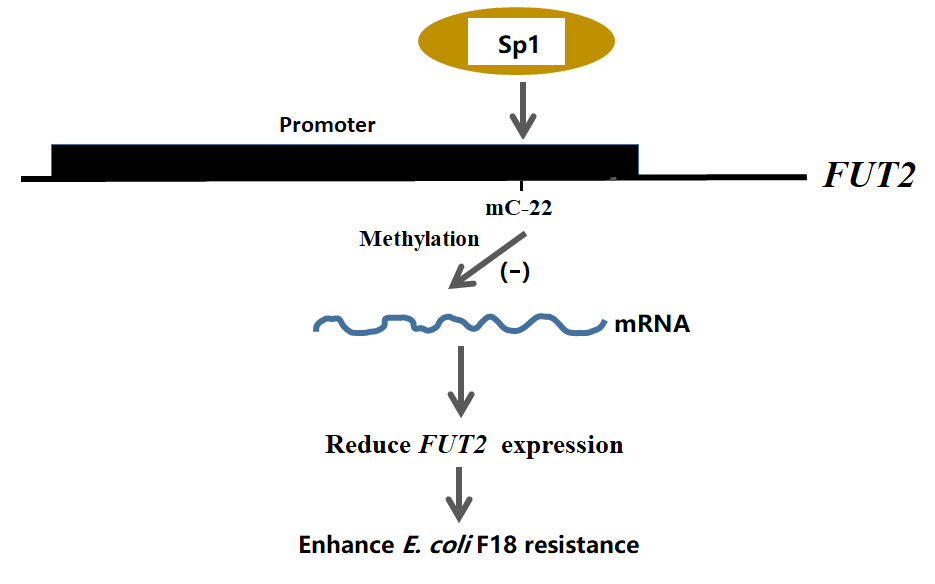{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
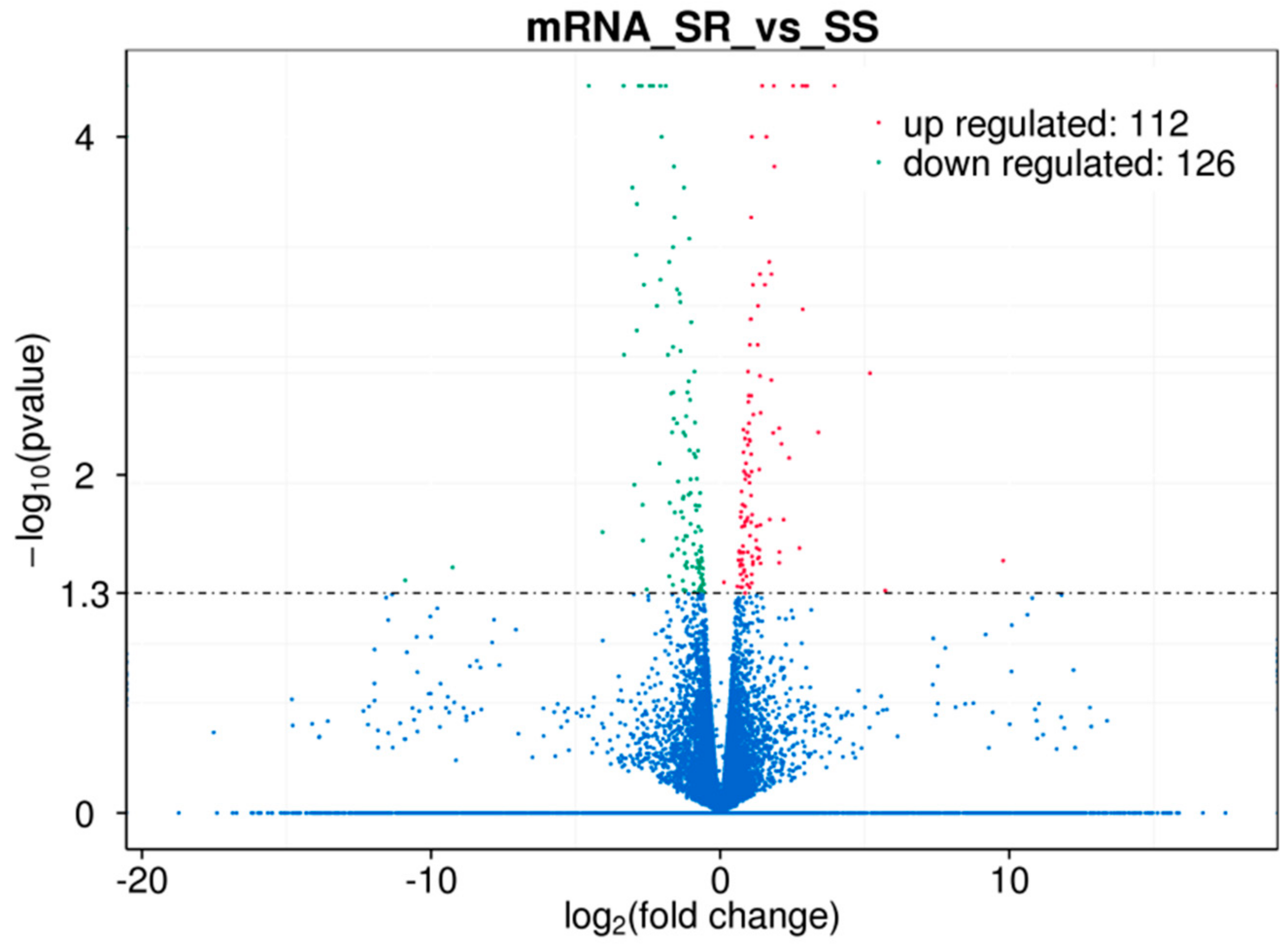
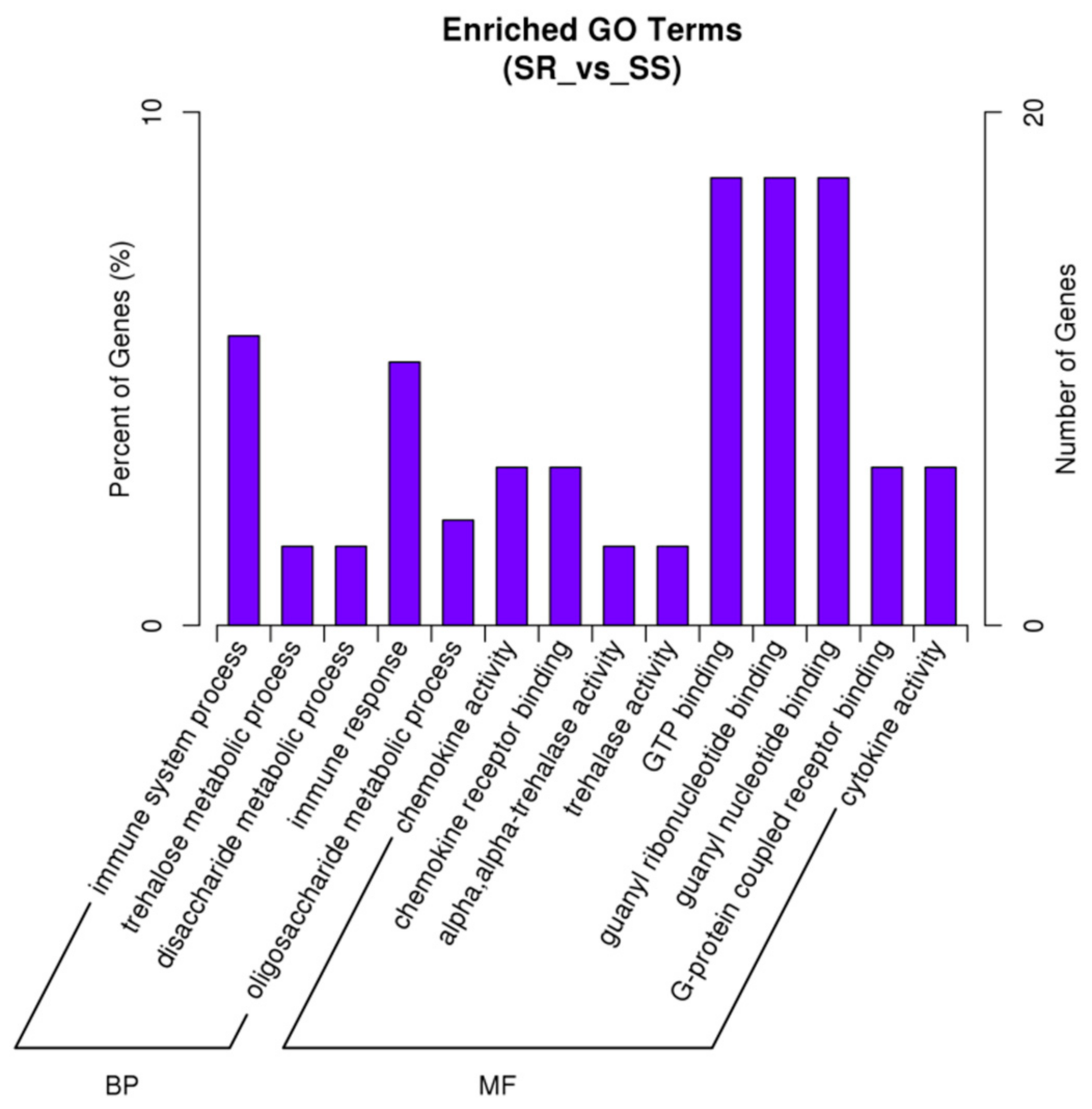
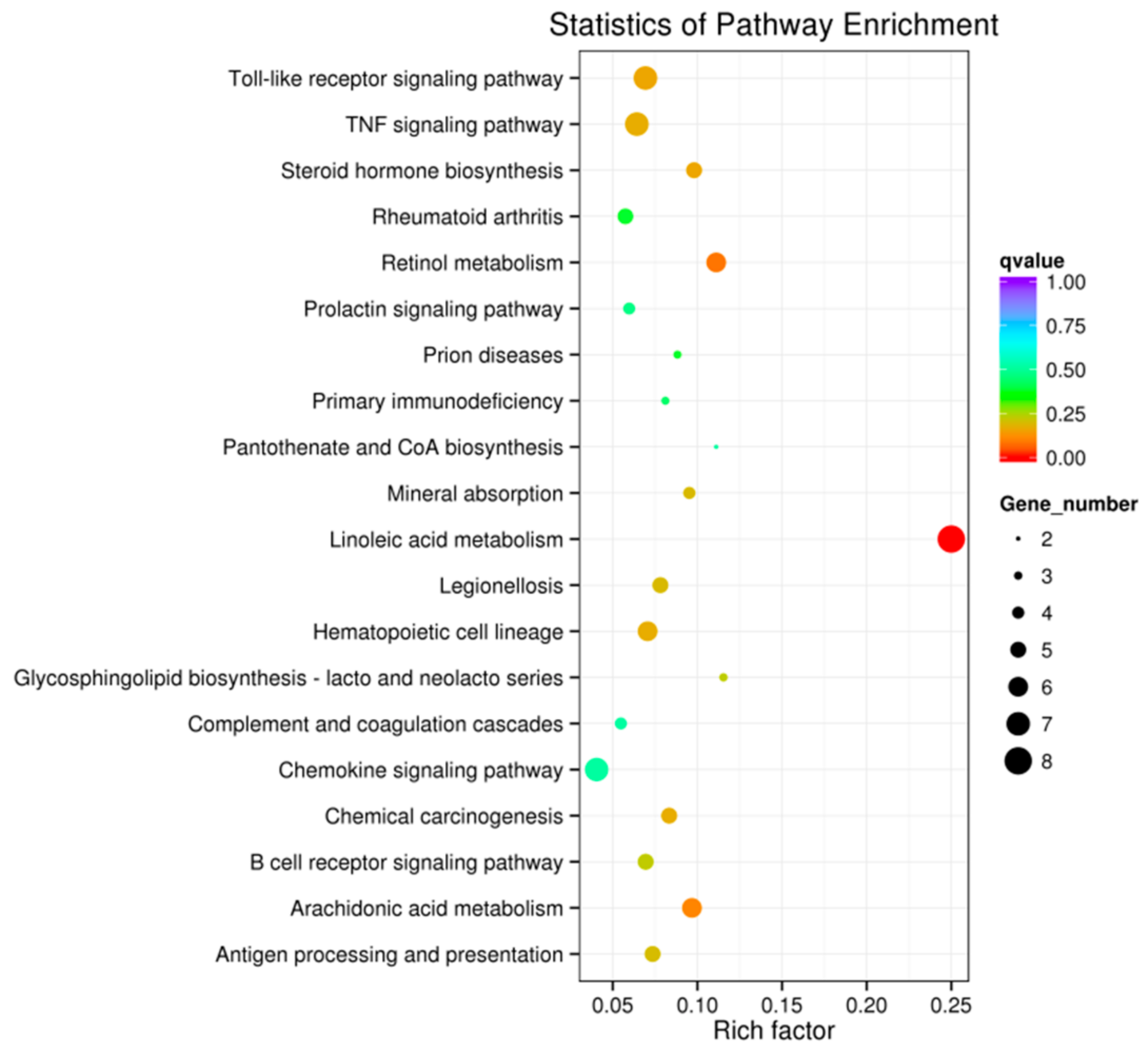
2.1. FUT2 Was Identified as a Host Gene Related to E. coli F18 Resistance Based on an RNA-Seq Analysis
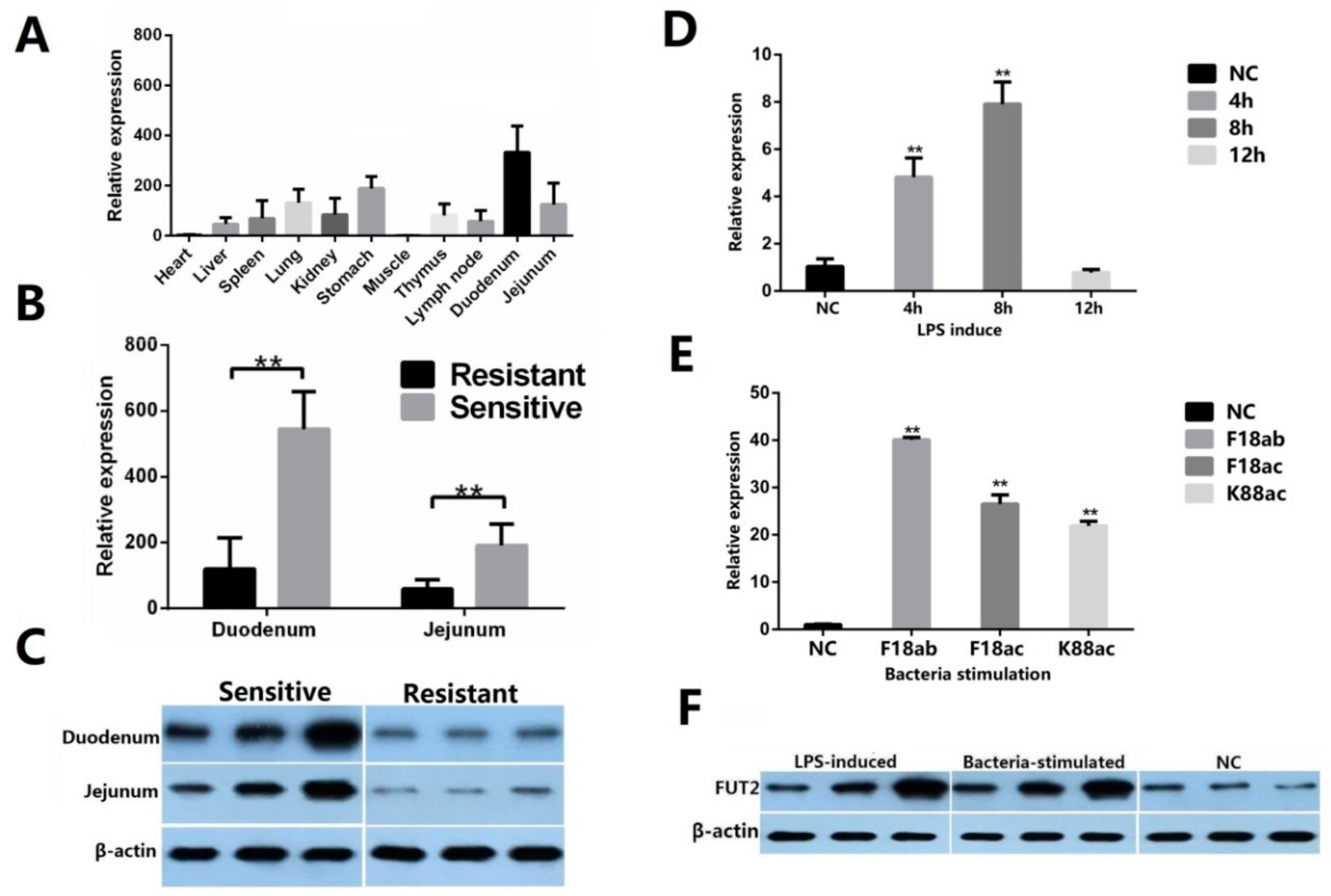
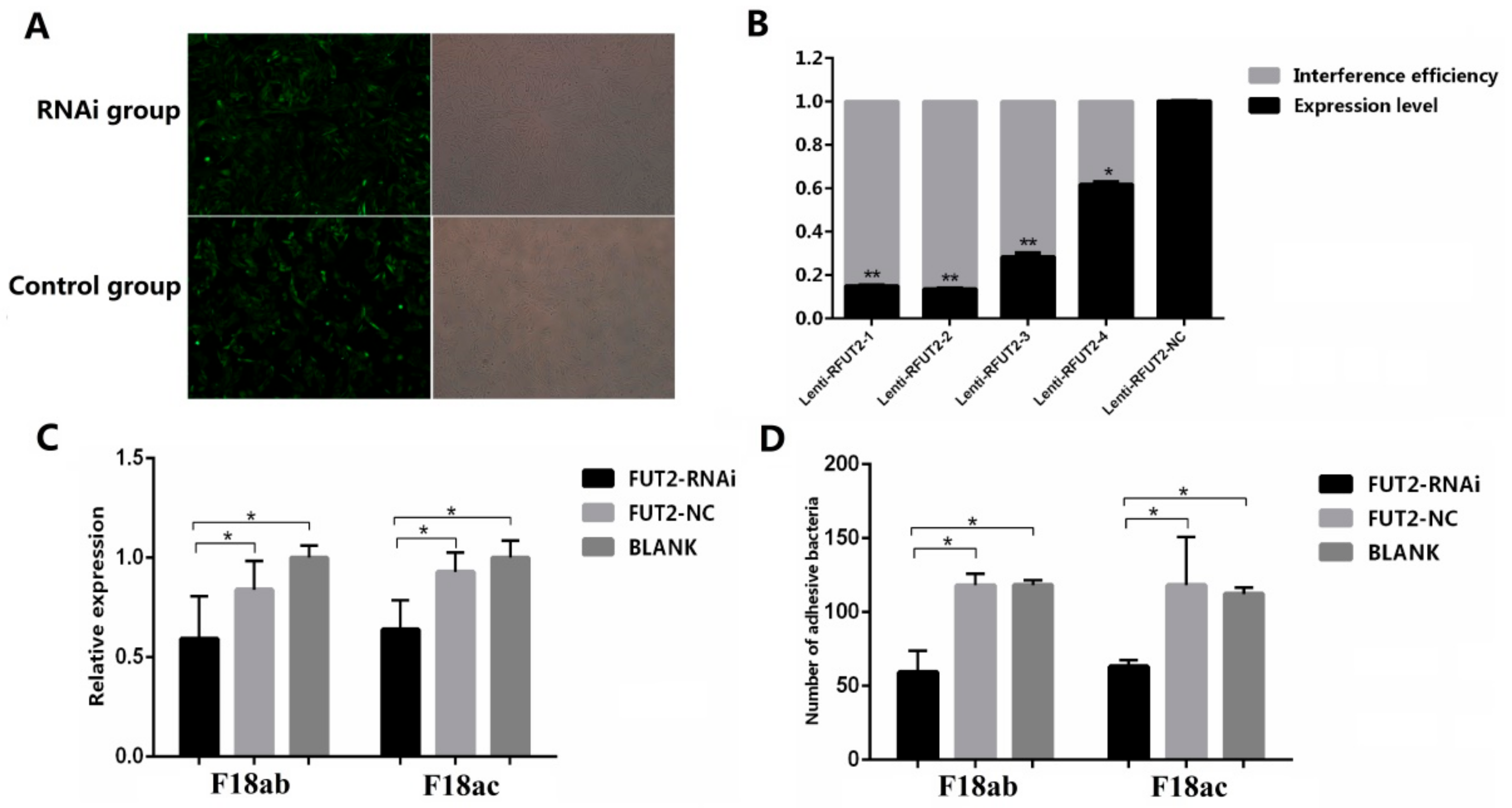
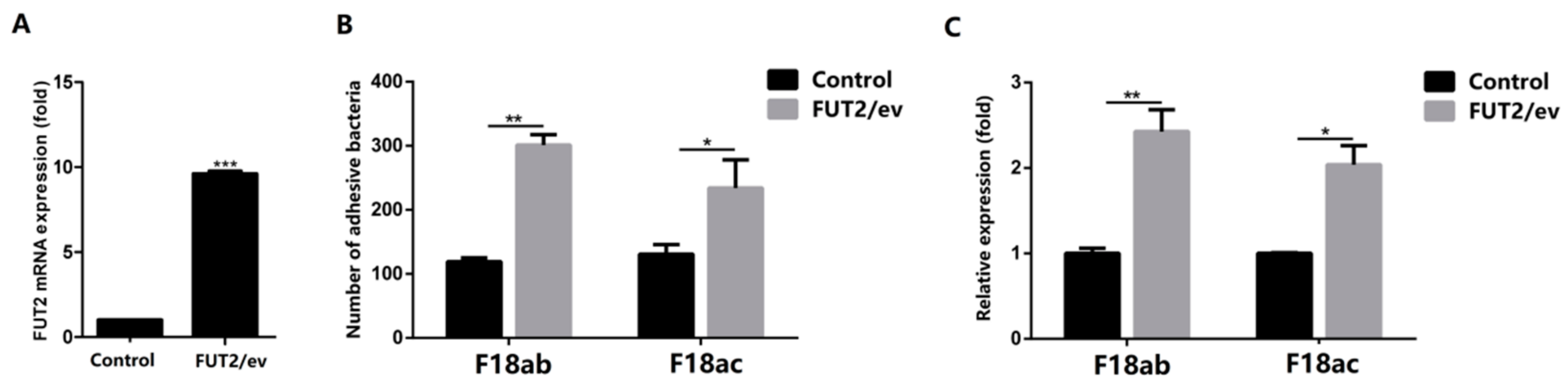
2.2. Decreased Expression of FUT2 Contributes to Enhancing E. coli F18 Resistance in Piglets
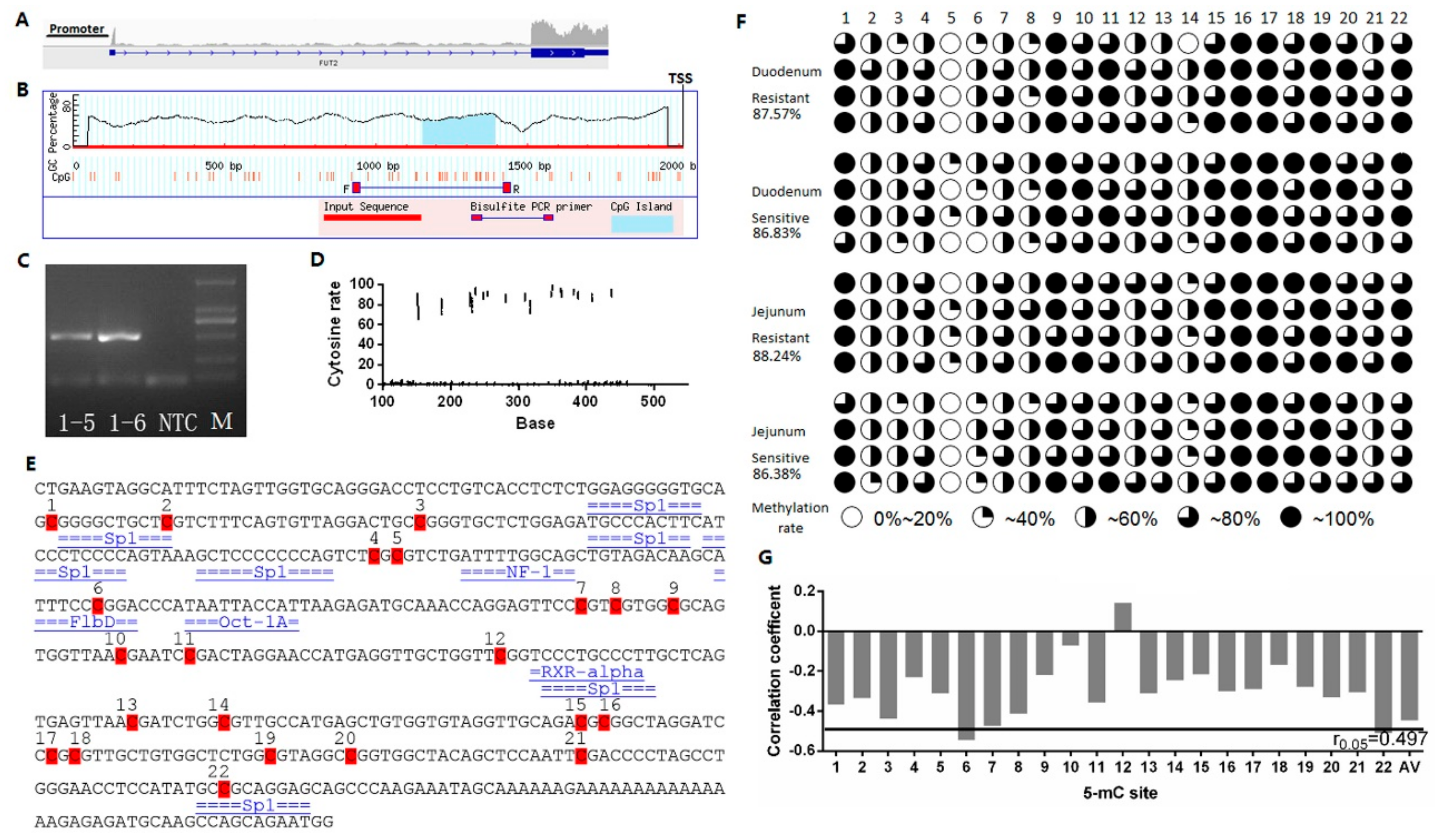
2.3. Effect of FUT2 Promoter Methylation Level on Gene Expression
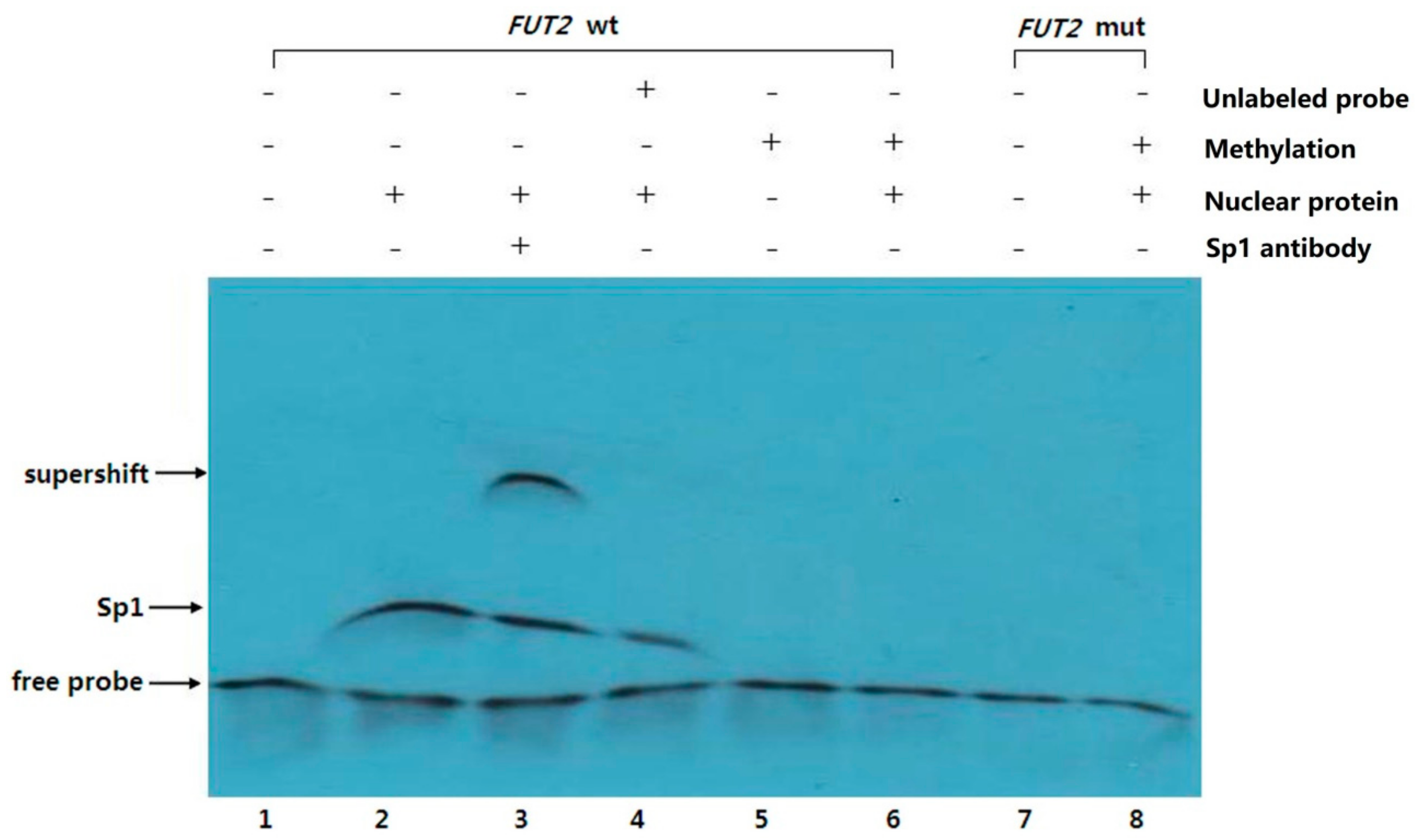
2.4. EMSA Analysis of Sp1 Binding to the FUT2 Gene Promoter
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Transcriptome Sequencing and Data Analysis
4.2.1. cDNA Library Preparation and Sequencing
4.2.2. Identification of Differential Expression Genes (DEGs)
4.2.3. Gene Function Annotation
4.3. Cell Culture, LPS-Induction and E. coli Stimulation in IPEC-J2 Cells
4.4. RNA Interference and Overexpression of FUT2 Gene in IPEC-J2 Cell and its Effects on E. coli F18 Adhesion In Vitro
4.5. qRT-PCR Analysis
4.6. Western Blot Analysis
4.7. Bioinformatics Analysis of the FUT2 Promoter
4.8. Methylation Sequencing of FUT2 Promoter
4.9. Electrophoretic Mobility Shift Assay
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| E. coli | Escherichia coli |
| LPS | Lipopolysaccharide |
| PWD | Post-weaning diarrhea |
| FUT2 | α(1,2) fucosyltransferase 2 |
| Sp1 | Specificity protein 1 |
| EMSA | Electrophoretic mobility shift assays |
| NGS | Next-generation sequencing |
| RNA-seq | RNA sequencing |
| BSAS | Bisulfite amplicon sequencing |
| DEGs | Differential expression genes |
| TFBS | Transcription factor binding site |
| IGV | Integrative Genomics Viewer |
| TSS | Transcriptional start site |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
References
- Boldin, B. Persistence and spread of gastro-intestinal infections: The case of enterotoxigenic Escherichia coli in piglets. Bull. Math. Biol. 2008, 70, 2077–2101. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Pan, Z.Y.; Wang, X.G.; Hua, J.D.; Wu, S.L.; Bao, W.B. Polymorphism of FUT1 gene and its relationship with growth and development in foundation colony of sutai ETEC F18+ resistant breeding pigs. China Anim. Husb. Vet. Med. 2010, 37, 158–161. (In Chinese) [Google Scholar]
- Wu, Z.C.; Liu, Y.; Dong, W.H.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. CD14 in the TLRs signaling pathway is associated with the resistance to E. coli F18 in Chinese domestic weaned piglets. Sci. Rep. 2016, 6, 24611. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.W.; Sun, L.; Qin, W.Y.; Gan, L.N.; Bao, W.B.; Wu, S.L. Developmental expression of LTβR and differential expression in Escherichia coli F18 resistant/sensitive piglets. Genet. Mol. Res. 2016, 15, gmr.15038377. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gan, L.N.; Qin, W.Y.; Sun, S.Y.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Differential expression of Toll-like receptor 4 signaling pathway genes in Escherichia coli F18-resistant and -sensitive Meishan piglets. Pol. J. Vet. Sci. 2016, 19, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Dong, W.H.; Huo, Y.J.; Huang, X.G.; Wu, S.L.; Bao, W.B. Dynamic changes in TAP1 expression levels in newborn to weaning piglets, and its association with Escherichia coli F18 resistance. Genet. Mol. Res. 2014, 13, 3686–3692. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zi, C.; Pan, Z.Y.; Zhu, J.; Du, Z.D.; Zhu, G.Q.; Huang, X.G.; Bao, W.B.; Wu, S.L. Investigation of the relationship between SLA-1 and SLA-3 gene expression and susceptibility to Escherichia coli F18 in post-weaning pigs. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, E.; Neuenschwander, S.; Fries, R.; Dinter, A.; Bertschinger, H.U.; Stranzinger, G.; Vögeli, P. A DNA polymorphism influencing α(1,2) fucosyltransferase activity of the pig FUT1 enzyme determines susceptibility of small intestinal epithelium to Escherichia coli F18 adhesion. Immunogenetics 2000, 52, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.S.; Huang, S.Q.; Liu, X.C.; He, C.Q.; Jiang, J. Polymorphism of E. coli F18 receptor gene in different pig breeds. Acta Genet. Sin. 2003, 30, 221–224. [Google Scholar] [PubMed]
- Yan, X.M.; Guo, Y.M.; Ding, N.S.; Ren, J. Study on the genetic variation of α1-fucosytransferase gene in different pig breeds. Chin. J. Anim. Sci. 2004, 40, 8–10. (In Chinese) [Google Scholar]
- Bao, W.B.; Wu, S.L.; Musa, H.H.; Zhu, G.Q.; Chen, G.H. Genetic variation at the α-1 fucosyltransferase (FUT1) gene in Asian wild boar and Chinese and Western commercial pig breeds. J. Anim. Breed. Genet. 2008, 125, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Coddens, A.; Diswall, M.; Angström, J.; Breimer, M.E.; Goddeeris, B.; Cox, E.; Teneberg, S. Recognition of blood group ABH type 1 determinants by the FedF adhesin of F18-fimbriated Escherichia coli. J. Biol. Chem. 2009, 284, 9713–9726. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, E.; Fries, R.; Vogeli, P.; Masabanda, J.; Wigger, G.; Stricker, C.; Neuenschwander, S.; Bertschinger, H.U.; Stranzinger, G. Two α (1,2) fucosyltransferase genes on porcine chromosome 6q11 are closely linked to the blood group inhibitor (S) and Escherichia coli F18 receptor (ECF18R) Loci. Mamm. Genome 1997, 8, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Vogeli, P.; Meijerink, E.; Fries, R.; Stricker, C.; Bertschinger, H.U. A molecular test for the detection of E. coli F18 receptors: A breakthrough in the struggle against oedema disease and postweaning diarrhoea in Swine. Schweiz Arch. Tierheilkd. 1997, 139, 479–484. [Google Scholar] [PubMed]
- Wang, J.; Yin, X.M.; Sun, L.; Zi, C.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Correlation between BPI Gene upstream CpG island methylation and mRNA expression in piglets. Int. J. Mol. Sci. 2014, 15, 10989–10998. [Google Scholar] [CrossRef] [PubMed]
- Aslani, S.; Mahmoudi, M.; Garshasbi, M.; Jamshidi, A.R.; Karami, J.; Nicknam, M.H. Evaluation of DNMT1 gene expression profile and methylation of its promoter region in patients with ankylosing spondylitis. Clin. Rheumatol. 2016, 35, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Chu, Y.; Kordower, J.H.; Li, B.; Cao, H.; Huang, L.; Nishida, M.; Song, L.; Wang, D.; Federoff, H.J. PGC-1α promoter methylation in Parkinson’s disease. PLoS ONE 2015, 10, e0134087. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, J.; Yin, X.; Sun, S.Y.; Zi, C.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Identification of a 5-methylcytosine site that may regulate C/EBPβ binding and determine tissue-specific expression of the BPI Gene in piglets. Sci. Rep. 2016, 6, 28506. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.H.; Yin, X.M.; Sun, L.; Wang, J.; Sun, S.Y.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Age-associated methylation change of TAP1 promoter in piglet. Gene 2015, 573, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Laurent, L.; Wong, E.; Li, G.; Huynh, T.; Tsirigos, A.; Ong, C.T.; Low, H.M.; Kin Sung, K.W.; Rigoutsos, I.; Loring, J.; et al. Dynamic changes in the human methylome during differentiation. Genome Res. 2010, 20, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Franca, L.T.; Carrilho, E.; Kist, T.B. A review of DNA sequencing techniques. Q. Rev. Biophys. 2002, 35, 169–200. [Google Scholar] [CrossRef] [PubMed]
- Stamps, B.W.; Corsetti, F.A.; Spear, J.R.; Stevenson, B.S. Draft genome of a novel Chlorobi member assembled by tetranucleotide binning of a hot spring metagenome. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Masser, D.R.; Berg, A.S.; Freeman, W.M. Focused, high accuracy 5-methylcytosine quantitation with base resolution by benchtop next-generation sequencing. Epigenet. Chromatin 2013, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Macleod, D.; Charlton, J.; Mullins, J.; Bird, A.P. Sp1 sites in the mouse aprt gene promoter are required to prevent methylation of the CpG island. Genes Dev. 1994, 8, 2282–2292. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.J.; Harrison, J.; Molloy, P.L. Sp1 binding is inhibited by mCpmCpG methylation. Gene 1997, 195, 67–71. [Google Scholar] [CrossRef]
- Chuang, J.Y.; Chang, W.C.; Hung, J.J. Hydrogen peroxide induces Sp1 methylation and thereby suppresses cyclin B1 via recruitment of Suv39H1 and HDAC1 in cancer cells. Free Radic Biol. Med. 2011, 51, 2309–2318. [Google Scholar] [CrossRef] [PubMed]
- Benin, A.M.; Ducher-Suchaux, M.F. Relationship between virulence and adherence of various enterotoxigenic Escherichia coli: Strains to isolated intestinal epithelial cells from Chinese Meishan and European large white pigs. Am. J. Vet. Res. 1991, 52, 45–49. [Google Scholar]
- Coddens, A.; Verdonck, F.; Tiels, P.; Rasschaert, K.; Goddeeris, B.M.; Cox, E. The age-dependent expression of the F18+ E. coli receptor on porcine gut epithelial cells is positively correlated with the presence of histo-blood group antigens. Vet. Microbiol. 2007, 122, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Deprez, P.; Van den Hende, C.; Muylle, E.; Oyaert, W. The influence of the administration of sow’s milk on the postweaning excretion of hemolytic E. coli in the pig. Vet. Res. Commun. 1986, 10, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Oriol, R.; Le Pendu, J.; Mollicone, R. Genetics of ABO, H, Lewis, X and related antigens. Vox Sang. 1986, 51, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Barrera, V.; Peinado, M.A. Evaluation of single CpG sites as proxies of CpG island methylation states at the genome scale. Nucleic Acids Res. 2012, 40, 11490–11498. [Google Scholar] [CrossRef] [PubMed]
- Mikeska, T.; Bock, C.; El-Maarri, O.; Hübner, A.; Ehrentraut, D.; Schramm, J.; Felsberg, J.; Kahl, P.; Büttner, R.; Pietsch, T.; et al. Optimization of quantitative MGMT promoter methylation analysis using pyrosequencing and combined bisulfite restriction analysis. J. Mol. Diagn. 2007, 9, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.L.; Yu, Y.; Yuan, Z.F.; Yang, J.; Ma, P.P.; Li, D.C.; Yu, S.K.; An, F.; Feng, X.J.; Zhang, Y. Comparative analysis on content and distribution of CpG sites in milk production traits and mastitis-related genes in dairy cattle. Yi Chuan 2012, 34, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Meilinger, D.; Fellinger, K.; Bultmann, S.; Rothbauer, U.; Bonapace, I.M.; Klinkert, W.E.; Spada, F.; Leonhardt, H. Np95 interacts with de novo DNA methyltransferases, Dnmt3a and Dnmt3b, and mediates epigenetic silencing of the viral CMV promoter in embryonic stem cells. EMBO Rep. 2009, 10, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.P.; Leonard, W.J. CREB/ATF-dependent T cell receptor-induced FoxP3 gene expression: A role for DNA methylation. J. Exp. Med. 2007, 204, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Perini, G.; Diolaiti, D.; Porro, A.; Della Valle, G. In vivo transcriptional regulation of N-Myc target genes is controlled by E-box methylation. Proc. Natl. Acad. Sci. USA 2005, 102, 12117–12122. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, G.C.; Lawe, D.; Ziff, E.B. Association of Myn, the murine homolog of max, with c-Myc stimulates methylation-sensitive DNA binding and ras cotransformation. Cell 1991, 65, 395–407. [Google Scholar] [CrossRef]
- Aoki, M.; Terada, T.; Kajiwara, M.; Ogasawara, K.; Ikai, I.; Ogawa, O.; Katsura, T.; Inui, K. Kidney-specific expression of human organic cation transporter 2 (OCT2/SLC22A2) is regulated by DNA methylation. Am. J. Physiol. Renal. Physiol. 2008, 295, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.C.; Felsenfeld, G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 2000, 405, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Uhm, T.G.; Lee, S.K.; Kim, B.S.; Kang, J.H.; Park, C.S.; Rhim, T.Y.; Chang, H.S.; Kim, D.J.; Chung, I.Y. CpG methylation at GATA elements in the regulatory region of CCR3 positively correlates with CCR3 transcription. Exp. Mol. Med. 2012, 44, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Comb, M.; Goodman, H.M. CpG methylation inhibits proenkephalin gene expression and binding of the transcription factor AP-2. Nucleic Acids Res. 1990, 18, 3975–3982. [Google Scholar] [CrossRef] [PubMed]
- Isomura, H.; Stinski, M.F.; Kudoh, A.; Daikoku, T.; Shirata, N.; Tsurumi, T. Two Sp1/Sp3 binding sites in the major immediate-early proximal enhancer of human cytomegalovirus have a significant role in viral replication. J. Virol. 2005, 79, 9597–9607. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.G.; Srinivasan, K.; Dai, Z.; Duan, W.; Druhan, L.J.; Ding, H.; Yee, L.; Villalona-Calero, M.A.; Plass, C.; Otterson, G.A. Methylation of adjacent CpG sites affects Sp1/Sp3 binding and activity in the p21 (Cip1) promoter. Mol. Cell. Biol. 2003, 23, 4056–4065. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xia, R.; Yin, X.; Huo, Y.; Zhu, G.; Wu, S.; Bao, W. Proteomic analysis of duodenal tissue from Escherichia coli F18-resistant and -susceptible weaned piglets. PLoS ONE 2015, 10, e0127164. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.H.; Gan, L.N.; Qin, W.Y.; Zi, C.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Use of fluorescence quantitative polymerase chain reaction (PCR) for the detection of Escherichia coli adhesion to pig intestinal epithelial cells. Pol. J. Vet. Sci. 2016, 19, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Feng, H.; Cao, Y.; Huang, Y.; Dai, C.; Wu, S.; Bao, W. New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets. Int. J. Mol. Sci. 2018, 19, 3301. https://doi.org/10.3390/ijms19113301
Wu Z, Feng H, Cao Y, Huang Y, Dai C, Wu S, Bao W. New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets. International Journal of Molecular Sciences. 2018; 19(11):3301. https://doi.org/10.3390/ijms19113301
Chicago/Turabian StyleWu, Zhengchang, Haiyue Feng, Yue Cao, Yanjie Huang, Chaohui Dai, Shenglong Wu, and Wenbin Bao. 2018. "New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets" International Journal of Molecular Sciences 19, no. 11: 3301. https://doi.org/10.3390/ijms19113301
APA StyleWu, Z., Feng, H., Cao, Y., Huang, Y., Dai, C., Wu, S., & Bao, W. (2018). New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets. International Journal of Molecular Sciences, 19(11), 3301. https://doi.org/10.3390/ijms19113301