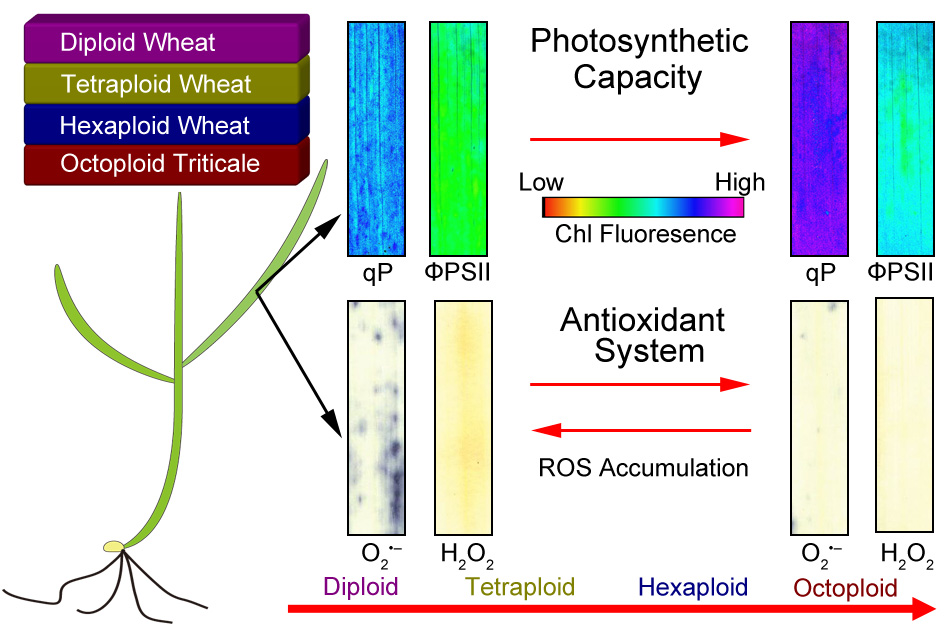
Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale


 ,
, 
Abstract

1. Introduction
2. Results
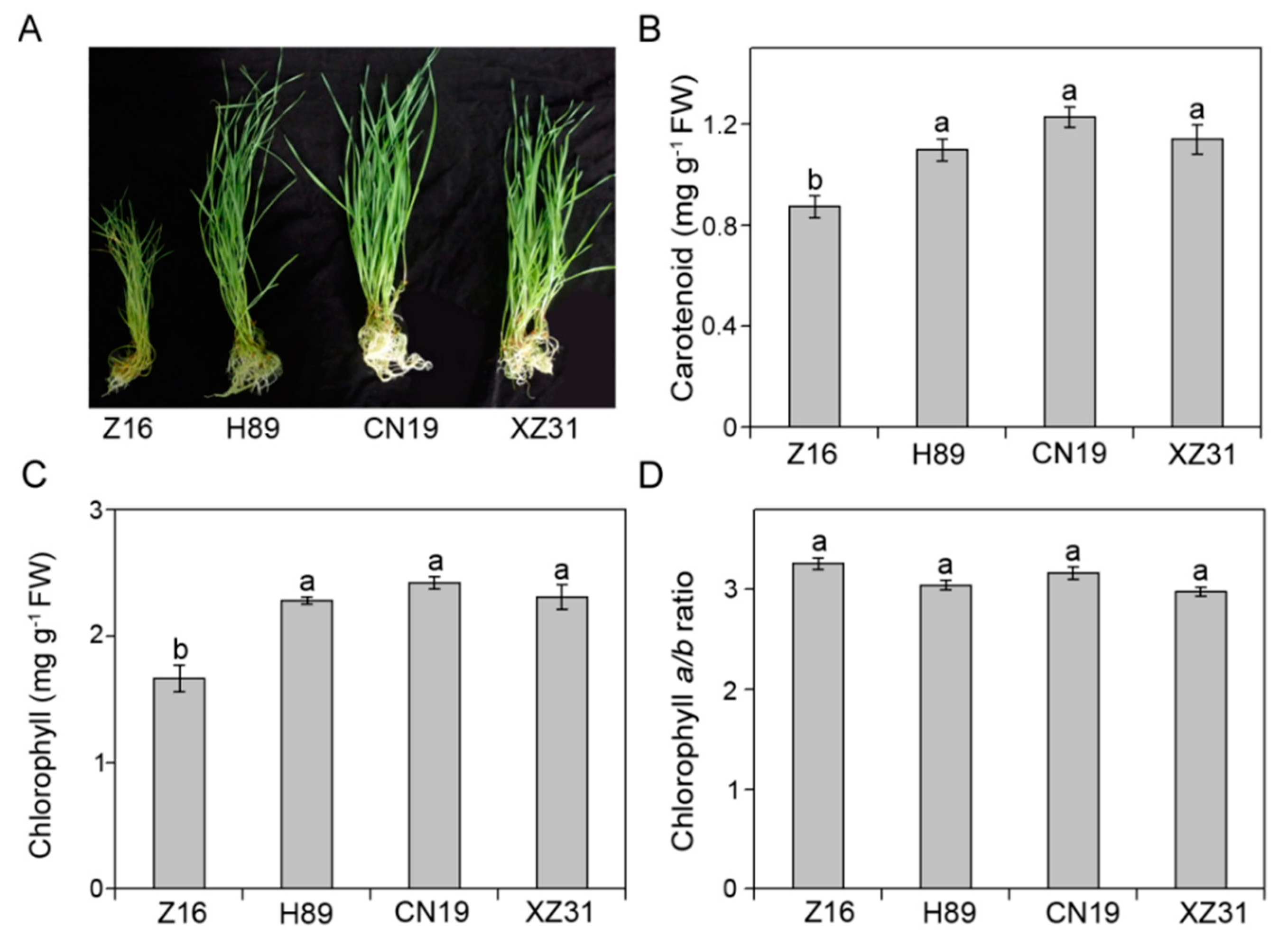
2.1. High Ploidy Levels Are Associated with Better Growth
2.2. High Ploidy Levels Are Associated with Higher Pigment Content, Proline Content, Total Protein, Soluble Sugar, But Lower Stomatal Density
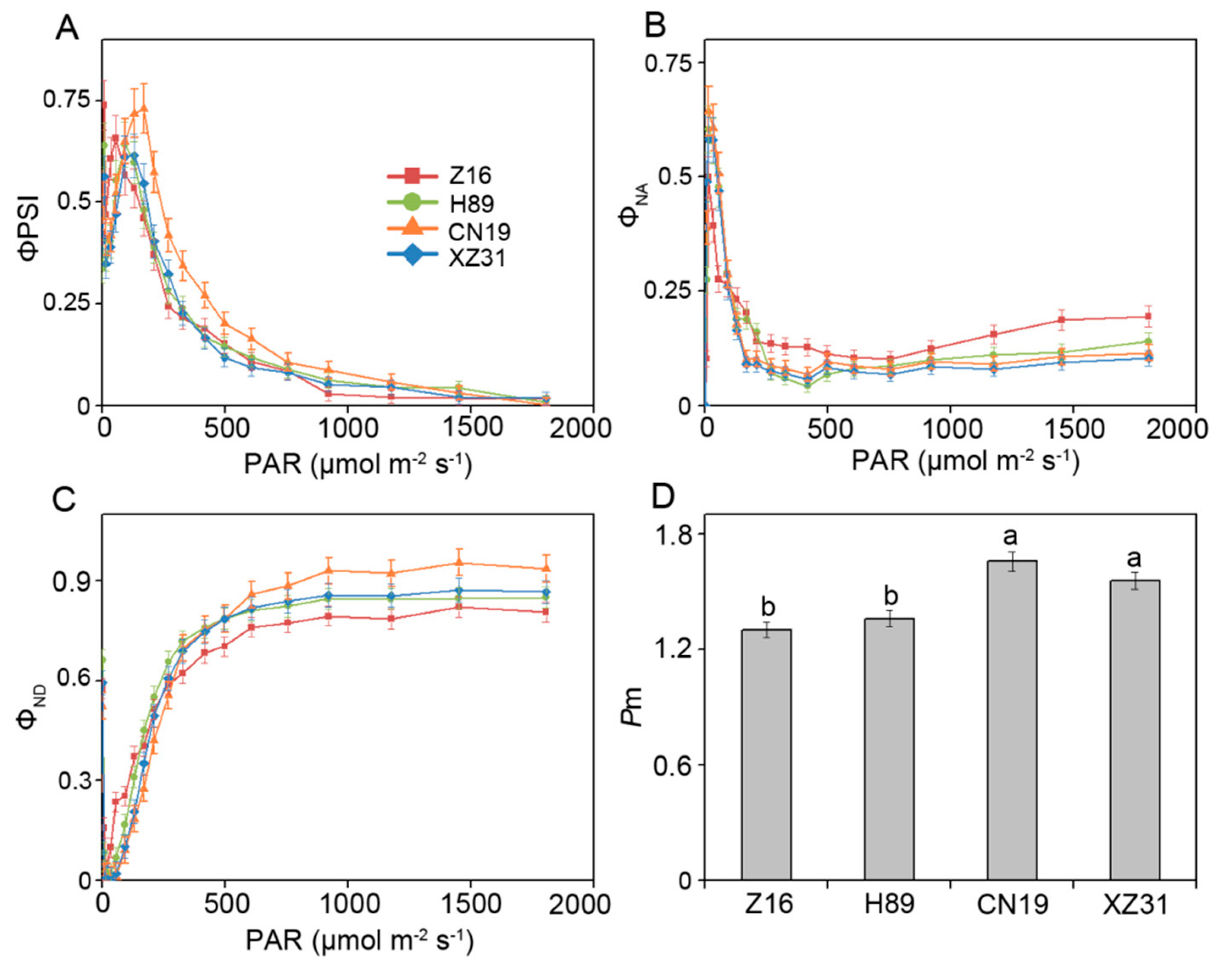
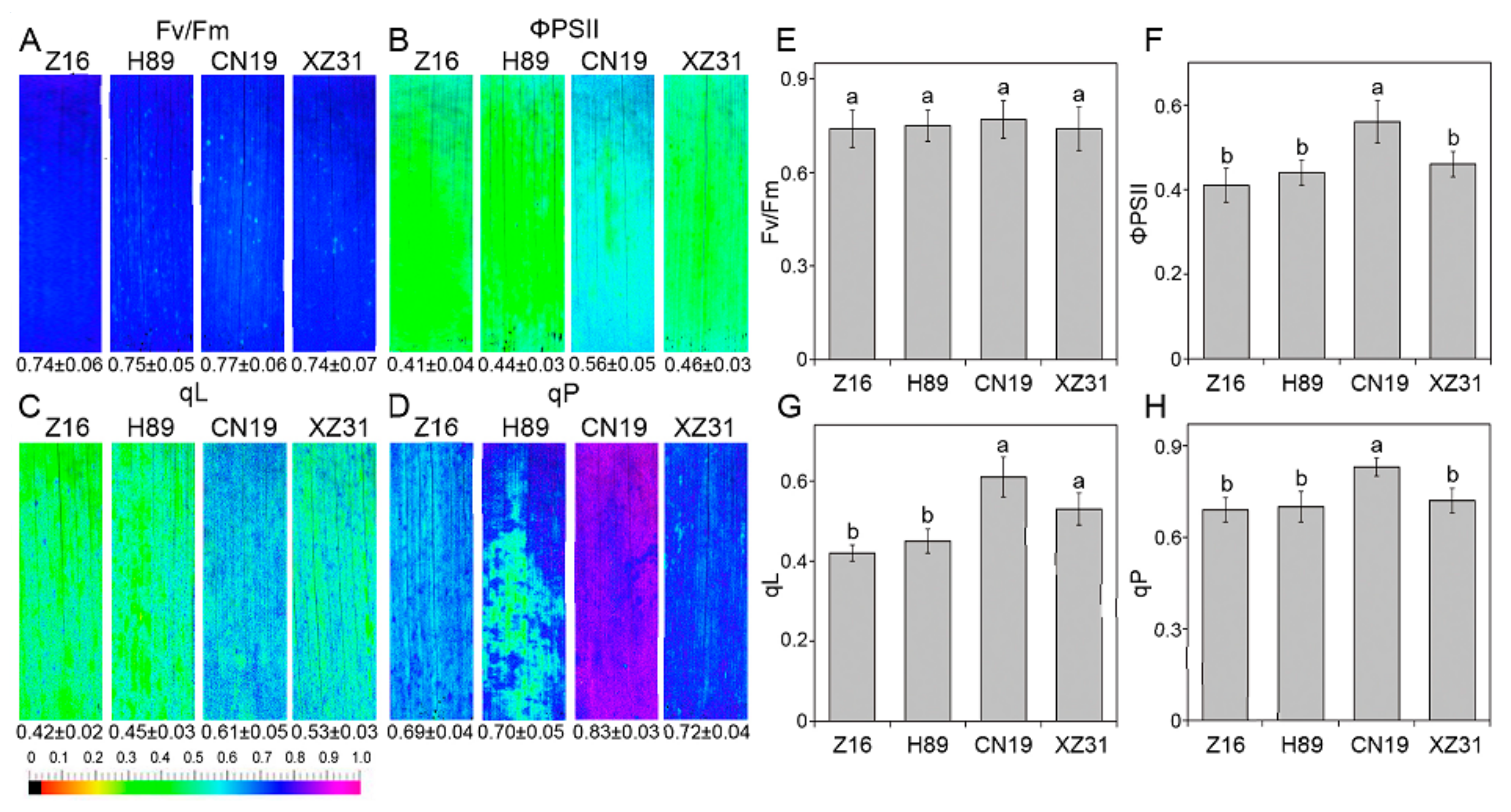
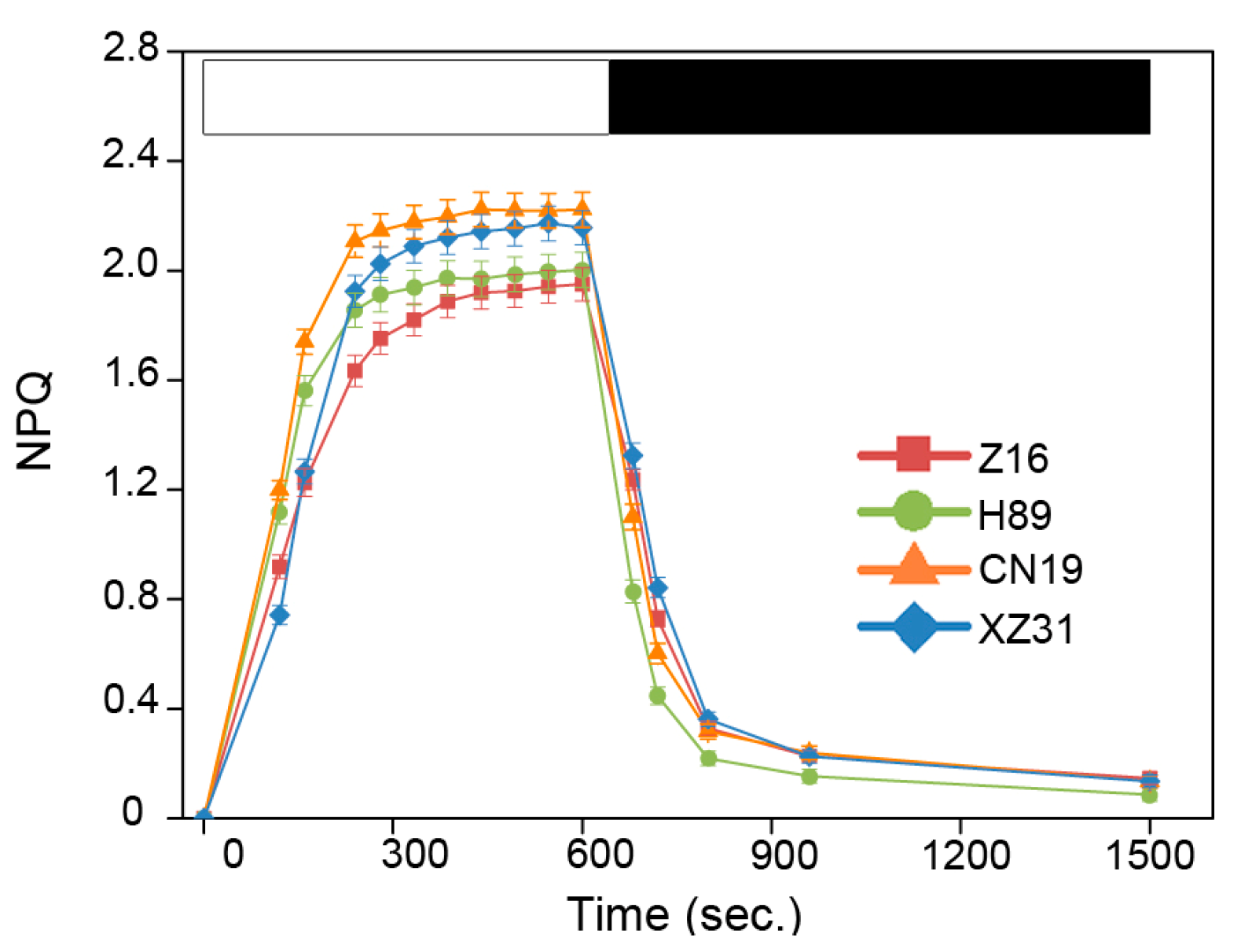
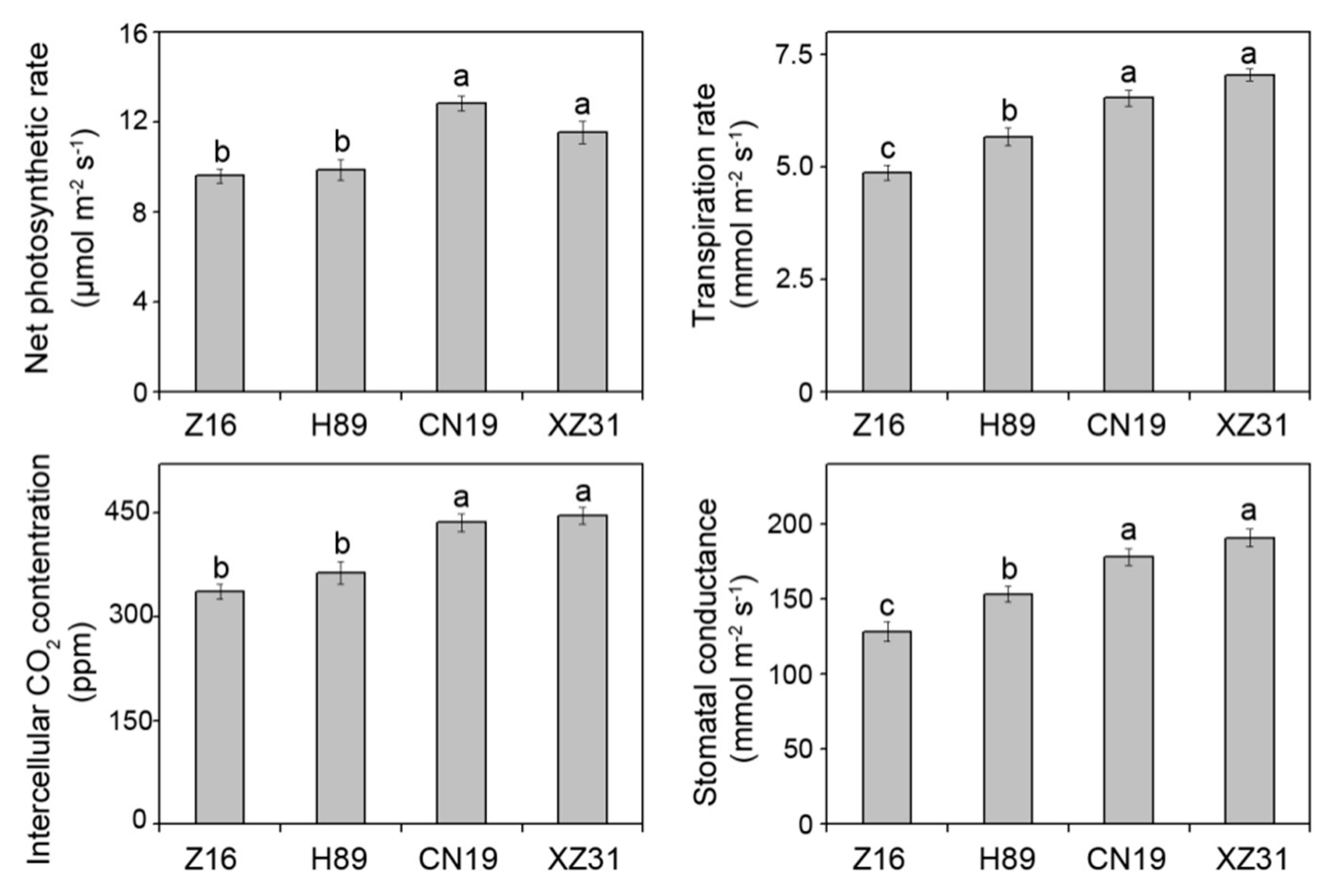
2.3. High Ploidy Levels Are Associated with Higher Photosynthetic Capacities
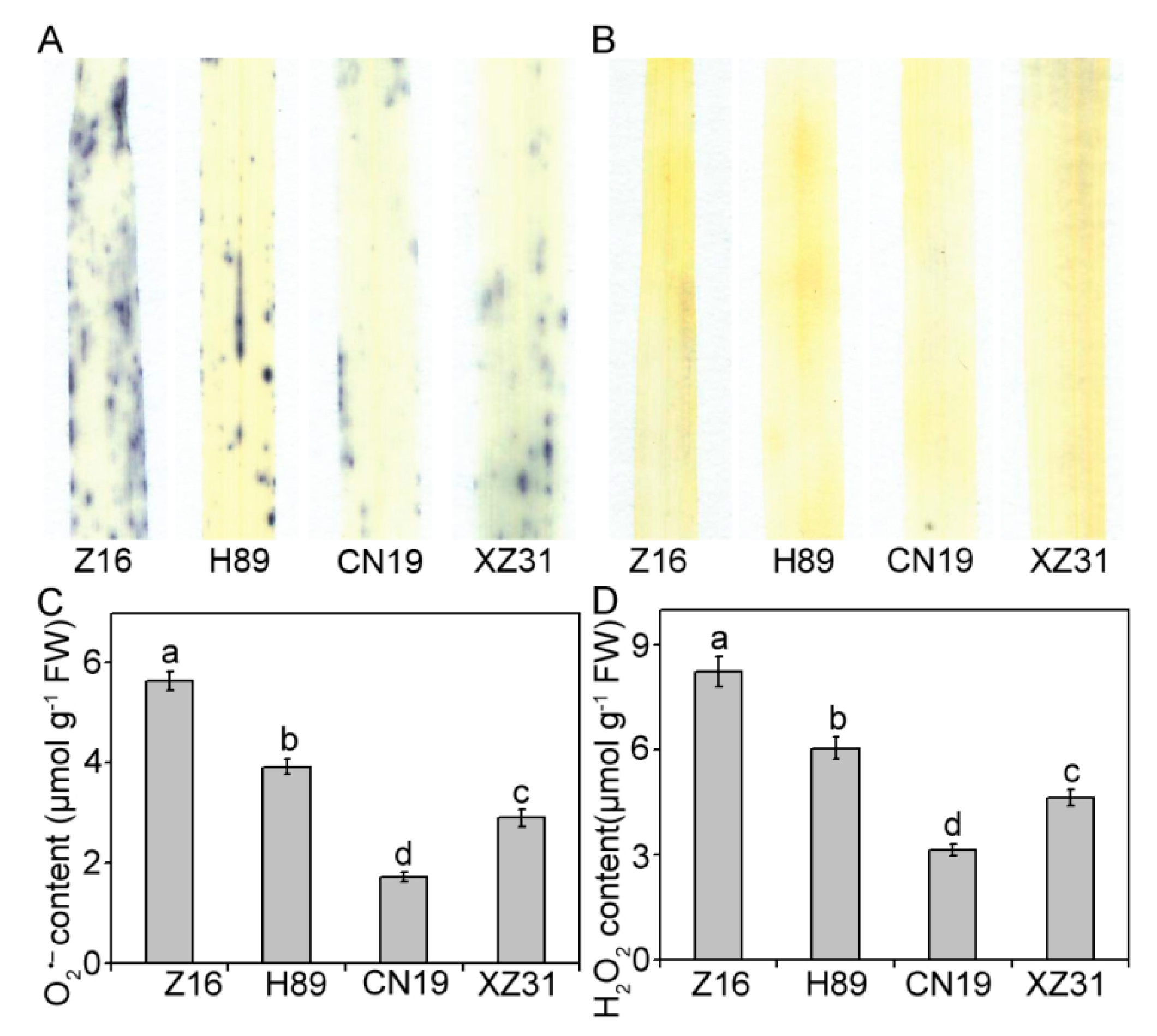
2.4. High Ploidy Levels Are Associated with Lower ROS Accumulation
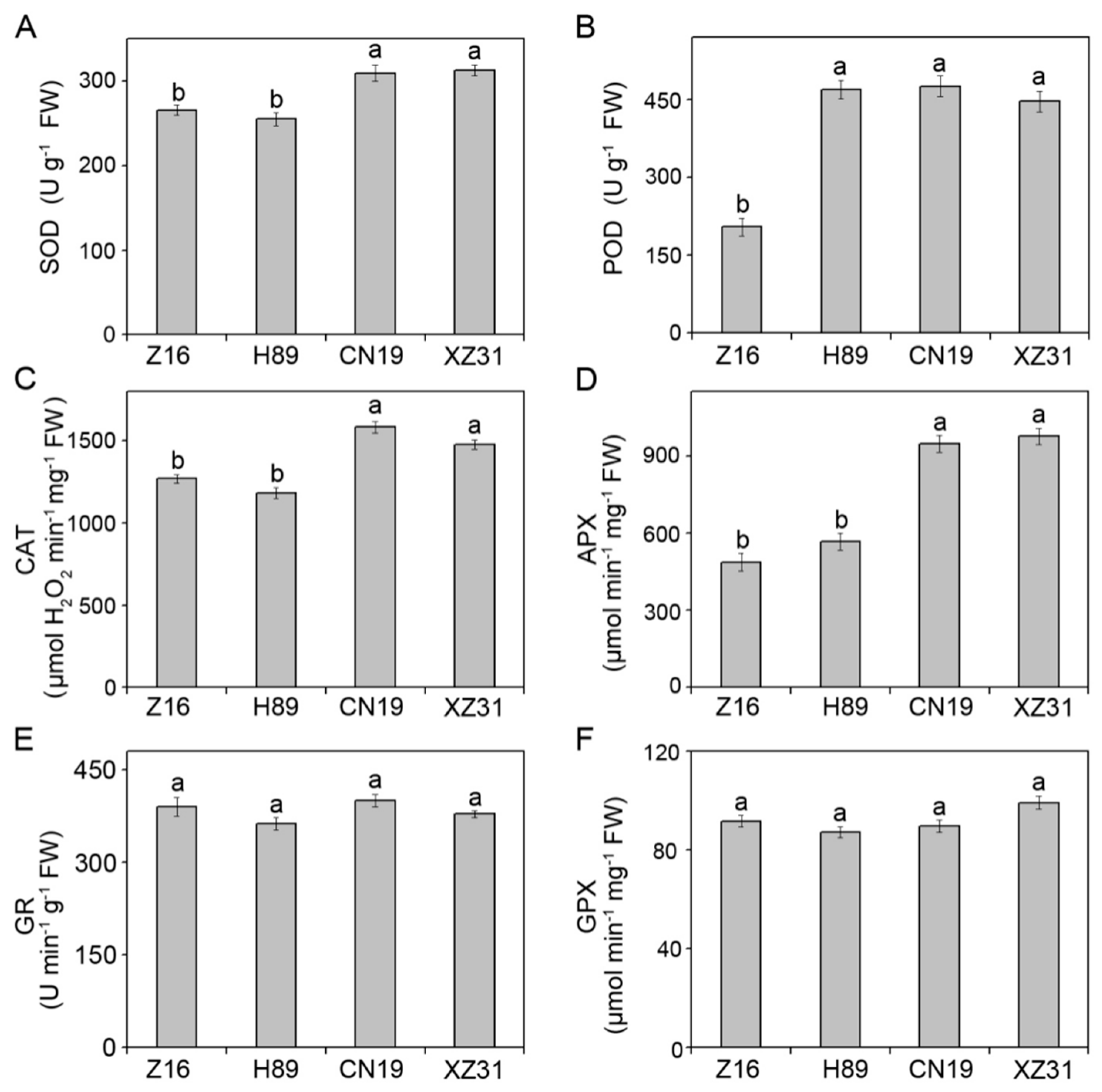
2.5. High Ploidy Levels Are Associated with Higher Enzymatic and Non-Enzymatic Antioxidant Activities
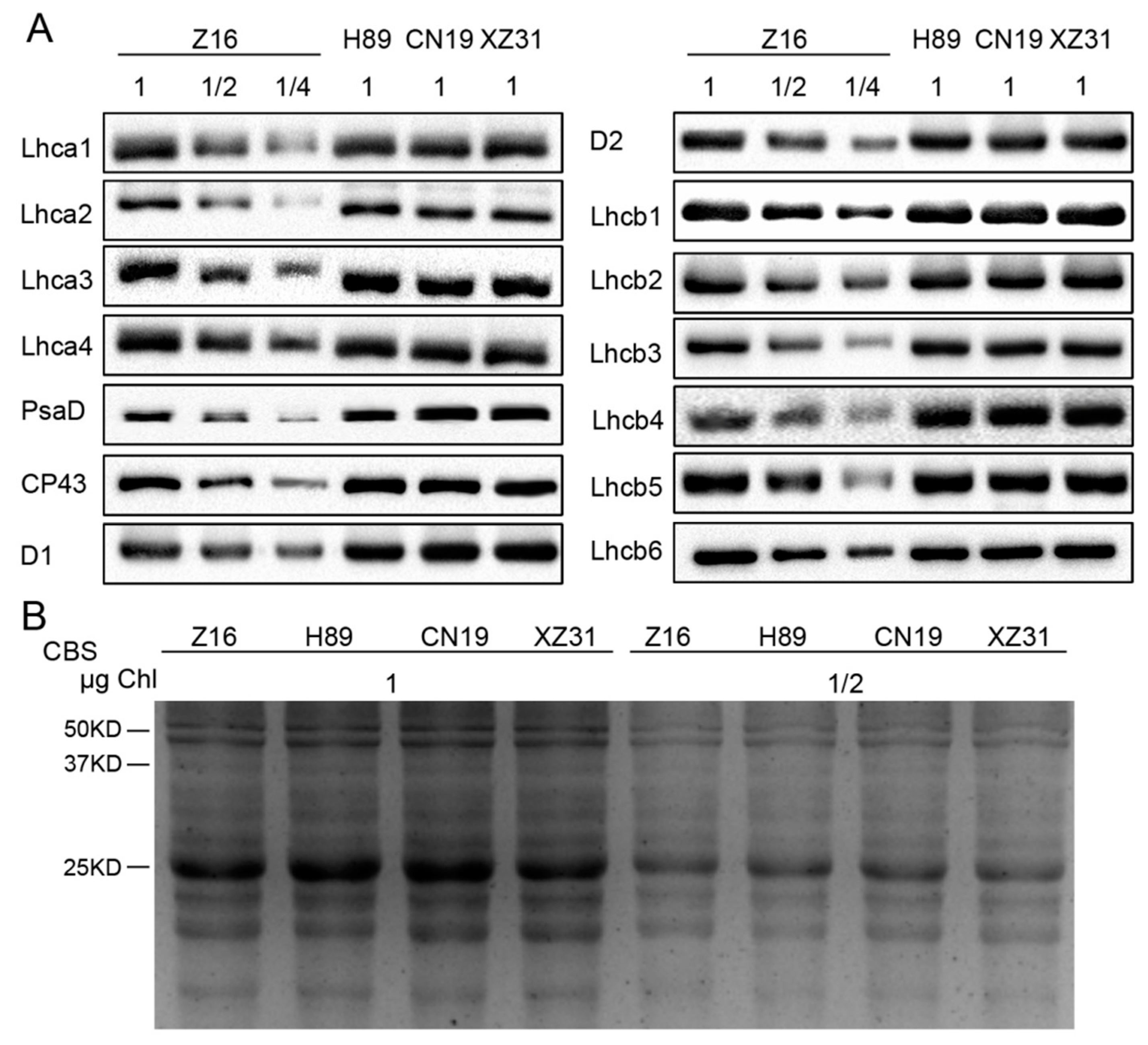
2.6. Ploidy Levels Are Not Associated with Thylakoid Membrane Protein Contents
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
4.2. Seed Germination, Seedling Growth, 1000-Grain Weight, and Stomata
4.3. Pigment Contents, Osmotic Regulators, and Total Protein Content
4.4. Measurements of Leaf Chlorophyll Fluorescence and Gas Exchange
4.5. Measurement of Reactive Oxygen Species (ROS)
4.6. Measurement of Antioxidative Enzymes and Non-Enzymatic Antioxidants
4.7. Immunoblot Analysis
4.8. Statistical Analysis
5. Conclusions and Perspective
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| PSII | Photosystem II |
| Chl | Chlorophyll |
| ROS | Reactive oxygen species |
| Pn | Net photosynthetic rate |
| NPQ | Non-photochemical quenching |
| Fv/Fm | Maximum quantum yield of PSII |
| ΦPSII | Effective quantum yield of PSII |
| Qp | Photochemical quenching |
| Ql | Fraction of PSII centers that are open |
References
- Chen, Y.E.; Su, Y.Q.; Zhang, C.M.; Ma, J.; Mao, H.T.; Yang, Z.H.; Yuan, M.; Zhang, Z.W.; Yuan, S.; Zhang, H.Y. Comparison of photosynthetic characteristics and antioxidant systems in different wheat strains. J. Plant Growth Regul. 2017, 1–13. [Google Scholar] [CrossRef]
- Wang, J.Y.; Turner, N.C.; Liu, Y.X.; Siddique, K.H.M.; Xiong, Y.C. Effects of drought stress on morphological, physiological and biochemical characteristics of wheat species differing in ploidy level. Funct. Plant Biol. 2017, 44, 219–234. [Google Scholar] [CrossRef]
- Tanno, K.; Willcox, G. How fast was wild wheat domesticated? Science 2006, 311, 1886. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, P.; Raskina, O.; Ma, X.F.; Nevo, E. Wheat evolution, domestication, and improvement. In Wheat Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 5–30. [Google Scholar]
- Brestic, M.; Zivcak, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.H.; Li, X.N.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Chantret, N.; Salse, J.; Sabot, F.; Rahman, S.; Bellec, A.; Laubin, B.; Dubois, I.; Dossat, C.; Sourdille, P.; Joudrier, P.; et al. Molecular basis of evolutionary events that shaped the Hardness locus in diploid and polyploid wheat species (Triticum and Aegilops). Plant Cell 2005, 17, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Dou, Q.W.; Tanaka, H.; Nakata, N.; Tsujimoto, H. Molecular cytogenetic analyses of hexaploid lines spontaneously appearing in octoploid Triticale. Theor. Appl. Genet. 2006, 114, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Kobayashi, S.; Furuta, Y. Effect of genome and ploidy on photosynthesis of wheat. Euphytica 1997, 94, 303–309. [Google Scholar] [CrossRef]
- Li, M.S.; Wang, C.Y.; Song, J.Q.; Chi, Y.G.; Wang, X.F.; Wu, Y.F. Evolutional trends of leaf stomatal and photosynthetic characteristics in wheat evolutions. Acta Ecol. Sin. 2008, 28, 5385–5391. [Google Scholar]
- Bilgrami, S.S.; Houshmand, S.A.; Khodambashi, M.; Zandi, P.; Siavoshi, M.; Khademi, S.; Navabpour, S.; Nasiri-Dehbaneh, M.; Amoue, H.; Tadayyon, M.R.; et al. Photosynthetic performance in ploidy levels and amphyploids of wheat during developmental stages. J. Anim. Plant Sci. 2015, 25, 1633–1643. [Google Scholar]
- Del Blanco, I.A.; Rajaram, S.; Kronstad, W.E.; Reynolds, M.P. Physiological performance of synthetic hexaploid wheat-derived populations. Crop Sci. 2000, 40, 1257–1263. [Google Scholar] [CrossRef]
- Nygren, J.; Shad, N.; Kvarnheden, A.; Westerbergh, A. Variation in susceptibility to wheat dwarf virus among wild and domesticated wheat. PLoS ONE 2015, 10, e0121580. [Google Scholar] [CrossRef] [PubMed]
- Austin, R.B.; Morgan, C.L.; Ford, M.A.; Bhagwat, S.G. Flag leaf photosynthesis of Triticum aestivum and related diploid and tetraploid Species. Ann. Bot. 1982, 49, 177–189. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li, M.S.; Song, J.Q.; Chi, Y.G.; Wang, X.F.; Wu, Y.F. Differences in stomatal and photosynthetic characteristics of five diploidy wheat species. Acta Ecol. Sin. 2008, 28, 3277–3283. [Google Scholar]
- Zhang, S.Q.; Shan, L. Difference of water use efficiency of diploidy wheat species with different chromosome set and its relationship with root system growth. Acta Agron. Sin. 2003, 29, 569–573. [Google Scholar]
- Planchon, C.; Fesquet, J. Effect of the D genome and of selection on photosynthesis in wheat. Theor. Appl. Genet. 1982, 61, 359–365. [Google Scholar] [PubMed]
- Austin, R.B.; Ford, M.A.; Miller, T.E.; Morgan, C.L.; Parry, M.A.J. Variation in photosynthetic characteristics among Triticum species and attempts to exploit it in breeding. In Progress in Photosynthesis Research; Biggens, J., Ed.; Springer: Dordrecht, The Netherlands, 1987; pp. 361–368. [Google Scholar]
- Huang, M.L.; Deng, X.P.; Zhou, S.L.; Zhao, Y.Z. Grain yield and water use efficiency of diploid, tetraploid and hexaploid wheats. Acta Ecol. Sin. 2007, 27, 1113–1121. [Google Scholar]
- Halloran, G.M.; Pennell, A.L. Grain size and seedling growth of wheat at different ploidy levels. Ann. Bot. 1982, 49, 103–113. [Google Scholar] [CrossRef]
- Hejnák, V.; Hniličková, H.; Hnilička, F.; Andr, J. Gas exchange and Triticum sp. with different ploidy in relation to irradiance. Plant Soil Environ. 2016, 62, 47–52. [Google Scholar] [CrossRef]
- Schreiber, U.; Klughammer, C.; Neubauer, C. Measuring P700 absorbance changes around 830 nm with a new type of pulse modulation system. Z. Naturforsch. 1988, 43, 686–698. [Google Scholar] [CrossRef]
- Jensen, P.E.; Gilpin, M.; Knoetzel, J.; Scheller, H.V. The PSI-K subunit of photosystem I is involved in the interaction between light-harvesting complex I and the photosystem I reaction center core. J. Biol. Chem. 2000, 275, 24701–24708. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Soltis, P.S. Molecular data and the dynamic nature of polyploidy. Crit. Rev. Plant Sci. 1993, 12, 243–273. [Google Scholar] [CrossRef]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J. Polyploidy and ecological adaptation in wild yarrow. Proc. Natl. Acad. Sci. USA 2011, 108, 7096–7101. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alfocea, F.; Larher, F. Sucrose and proline accumulation and sugar efflux in tomato leaf discs affected by NaCl and polyethylene glycol 6000 iso-osmotic stresses. Plant Sci. 1995, 107, 9–15. [Google Scholar] [CrossRef]
- Singh, T.N.; Paleg, I.G.; Aspinall, D. Stress Metabolism III. Variations in response to water deficit in the barley plant. Aust. J. Biol. Sci. 1973, 26, 65–76. [Google Scholar]
- Monneveux, P.; Nemmar, M. Contribution to the study of drought resistance in bread wheat (Triticum aestivum L.) and durum wheat (Triticum durum Desf.): Study of proline accumulation during development. Agronomie 1986, 6, 583–590. [Google Scholar] [CrossRef]
- Sonoike, K. Photoinhibition of photosystem I. Physiol. Plant. 2011, 142, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Brestic, M.; Zivcak, M.; Kunderlikova, K.; Allakhverdiev, S.I. High temperature speciically afects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 2016, 130, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Miyake, C.; Miyata, M.; Shinzaki, Y.; Tomizawa, K. CO2 response of cyclic electron flow around PSI (CEF-PSI) in tobacco leaves–relative electron fluxes through PSI and PSII determine the magnitude of non-photochemical quenching (NPQ) of Chl fluorescence. Plant Cell Physiol. 2005, 46, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2017, 132, 13–66. [Google Scholar] [CrossRef] [PubMed]
- Graßes, T.; Pesaresi, P.; Schiavon, F.; Varotto, C.; Salamini, F.; Jahns, P.; Leister, D. The role of ∆pH-dependent dissipation of excitation energy in protecting photosystem II against light-induced damage in Arabidopsis thaliana. Plant Physiol. Biochem. 2002, 40, 41–49. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; von Caemmerer, S.; Condon, A.G.; Zwart, A.B.; Munns, R. Genetic variation in tolerance to the osmotic stress component of salinity stress in durum wheat. Funct. Plant Biol. 2008, 35, 111–123. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.; Jajoo, A.; Poudyal, R.S.; Timilsina, R.; Park, Y.S.; Aro, E.M.; Nam, H.G.; Lee, C.H. Towards a critical understanding of the photosystem II repair mechanism and its regulation during stress conditions. FEBS Lett. 2013, 587, 3372–3381. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhao, J.D.; Warren, P.V.; Warden, J.T.; Bryant, D.A.; Golbeck, J.H. PsaD is required for the stable binding of PsaC to the photosystem I core protein of Synechococcus sp. PCC 6301. Biochemistry 1991, 30, 7863–7872. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Zhao, Z.Y.; Zhang, H.Y.; Zeng, X.Y.; Yuan, S. The significance of CP29 reversible phosphorylation in thylakoids of higher plants under environmental stresses. J. Exp. Bot. 2013, 64, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Munzuroglu, O.; Geckil, H. Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Arch. Environ. Contam. Toxicol. 2002, 43, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.C.; Li, F.M.; Zhang, T. Performance of wheat crops with different chromosome ploidy: Root-sourced signals, drought tolerance, and yield performance. Planta 2006, 224, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kajita, R.; Miyazaki, A.; Hokoyama, M.; Nakamura-Miura, T.; Mizuno, S.; Masuda, Y.; Irie, K.; Tanaka, Y.; Takada, S.; et al. Stomatal density is controlled by a mesophyll-derived signaling molecule. Plant Cell Physiol. 2010, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Jensen, A. Chlorophylls and carotenoids. In Handbook of Phycological Methods: Physiological and Biochemical Methods; Hellebust, A., Crargie, J.S., Eds.; Cambridge University Press: London, UK, 1978; pp. 59–70. [Google Scholar]
- Thomas, T.A. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence–a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. An improved method, using saturating light pulses, for the determination of photosystem I quantum yield via P700+-absorbance changes at 830 nm. Planta 1994, 192, 261–268. [Google Scholar] [CrossRef]
- Pietrzykowska, M.; Suorsa, M.; Semchonok, D.A.; Tikkanen, M.; Boekema, E.J.; Aro, E.M.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef] [PubMed]
- Leoni, C.; Pietrzykowska, M.; Kiss, A.Z.; Suorsa, M.; Ceci, L.R.; Aro, E.M.; Jansson, S. Very rapid phosphorylation kinetics suggest a unique role for Lhcb2 during state transitions in Arabidopsis. Plant J. 2013, 76, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Noguchi, K.; Hanba, Y.T.; Terashima, I. Effects of internal conductance on the temperature dependence of the photosynthetic rate in spinach leaves from contrasting growth temperatures. Plant Cell Physiol. 2006, 47, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Cui, J.M.; Su, Y.Q.; Yuan, S.; Yuan, M.; Zhang, H.Y. Influence of stripe rust infection on the photosynthetic characteristics and antioxidant system of susceptible and resistant wheat cultivars at the adult plant stage. Front. Plant Sci. 2015, 6, 779. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Matsuda, Y.; Yamanaka, A.; Sagisaka, S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991, 97, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y.Y.; Ge, Q.; Li, Y.L.; Sun, J.H.; Zhang, Y.; Liu, X.J. Comparative physiological responses of Solanum nigrum and Solanum torvum to cadmium stress. New Phytol. 2012, 196, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Bechtold, U.; Murphy, D.J.; Mullineaux, P.M. Arabidopsis peptide methionine sulfoxide reductase2 prevents cellular oxidative damage in long nights. Plant Cell 2004, 16, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Yuan, S.; Schröder, W.P. Comparison of methods for extracting thylakoid membranes of Arabidopsis plants. Physiol. Plant. 2016, 156, 3–12. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Z16 | H89 | CN19 | XZ31 |
|---|---|---|---|---|
| (a) Seed germination | ||||
| Germination rate (%) | 49.3 ± 0.7 c | 62.5 ± 0.6 b | 87.8 ± 0.9 a | 35.2 ± 1.1 d |
| Germination potential (%) | 39.1 ± 0.3 c | 59.2 ± 0.8 b | 83.4 ± 0.6 a | 32.5 ± 0.5 d |
| Germination index (%) | 31.1 ± 0.5 c | 49.3 ± 0.3 b | 74.7 ± 0.5 a | 27.7 ± 0.6 d |
| (b) Seedling growth at trefoil stage | ||||
| Root length (cm) | 8.2 ± 0.3 d | 14.5 ± 0.2 c | 18.2 ± 0.1 b | 19.7 ± 0.3 a |
| Root number | 5.1 ± 0.6 b | 5.6 ± 0.3 b | 5.4 ± 0.2 b | 7.2 ± 0.2 a |
| Seedling height (cm) | 3.3 ± 0.3 d | 7.4 ± 0.1 a | 4.4 ± 0.2 c | 6.3 ± 0.1 b |
| Total length of seedlings (cm) | 25.3 ± 0.3 c | 44.2 ± 0.7 b | 44.4 ± 0.5 b | 46.9 ± 0.5 a |
| Leaf area (cm2) | 2.1 ± 0.1 d | 4.8 ± 0.1 c | 8.7 ± 0.2 b | 7.3 ± 0.2 a |
| (c) 1000-grain weight (g) | 7.9 ± 0.1 d | 41.4 ± 0.7 b | 52.8 ± 0.3 a | 33.2 ± 0.3 c |
| Parameters | Z16 | H89 | CN19 | XZ31 |
|---|---|---|---|---|
| (a) Stomata of the second leaves | ||||
| Single stomatal perimeter (μm) | 135.8 ± 13.8 c | 170.3 ± 5.2 b | 205.2 ± 6.4 b | 232.4 ± 12.7 a |
| Single stomatal area (μm2) | 1174 ± 45 d | 2067 ± 38 c | 2365 ± 55 b | 2805 ± 68 a |
| Stomatal density (per mm−2 × 10−3) | 49.9 ± 2.2 a | 29.5 ± 1.6 b | 26.9 ± 1.2 bc | 24.3 ± 0.8 c |
| (b) Osmotic regulators | ||||
| Soluble sugar content (mg g−1 FW) | 83.1 ± 0.8 d | 90.6 ± 0.9 c | 115.8 ± 1.0 b | 130.3 ± 0.6 a |
| Proline content (mg g−1 FW) | 37.8 ± 0.8 b | 41.2 ± 1.6 b | 51.6 ± 2.4 a | 51.2 ± 2.0 a |
| (c) Total protein content (mg g−1 FW) | 7.8 ± 0.1 b | 12.6 ± 0.2 a | 13.3 ± 0.4 a | 12.2 ± 0.1 a |
| Species | Accession | Genome | Sources | Ploidy | Ear | Grain |
|---|---|---|---|---|---|---|
| T. monococcum | Z16 (W2n) | AA | China | Diploid (2n = 2x = 14) | Dehiscent | Hulled |
| T. dicoccum | H89 (W4n) | AABB | China | Tetraploid (2n = 4x = 28) | Indehiscent | Naked |
| T. aestivum | CN19 (W6n) | AABBDD | China | Hexaploid (2n = 6x = 42) | Indehiscent | Naked |
| Triticale | XZ31 (T8n) | AABBDDRR | China | Octaploid (2n = 8x = 56) | Indehiscent | Naked |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, H.; Chen, M.; Su, Y.; Wu, N.; Yuan, M.; Yuan, S.; Brestic, M.; Zivcak, M.; Zhang, H.; Chen, Y. Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale. Int. J. Mol. Sci. 2018, 19, 3006. https://doi.org/10.3390/ijms19103006
Mao H, Chen M, Su Y, Wu N, Yuan M, Yuan S, Brestic M, Zivcak M, Zhang H, Chen Y. Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale. International Journal of Molecular Sciences. 2018; 19(10):3006. https://doi.org/10.3390/ijms19103006
Chicago/Turabian StyleMao, Haotian, Mengying Chen, Yanqiu Su, Nan Wu, Ming Yuan, Shu Yuan, Marian Brestic, Marek Zivcak, Huaiyu Zhang, and Yanger Chen. 2018. "Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale" International Journal of Molecular Sciences 19, no. 10: 3006. https://doi.org/10.3390/ijms19103006
APA StyleMao, H., Chen, M., Su, Y., Wu, N., Yuan, M., Yuan, S., Brestic, M., Zivcak, M., Zhang, H., & Chen, Y. (2018). Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale. International Journal of Molecular Sciences, 19(10), 3006. https://doi.org/10.3390/ijms19103006