Harnessing Invariant NKT Cells to Improve Influenza Vaccines: A Pig Perspective


Abstract
1. Introduction
2. Challenges Facing the Development of Effective Swine Influenza Vaccines
3. Current Understanding of the Porcine iNKT Cell–CD1d System
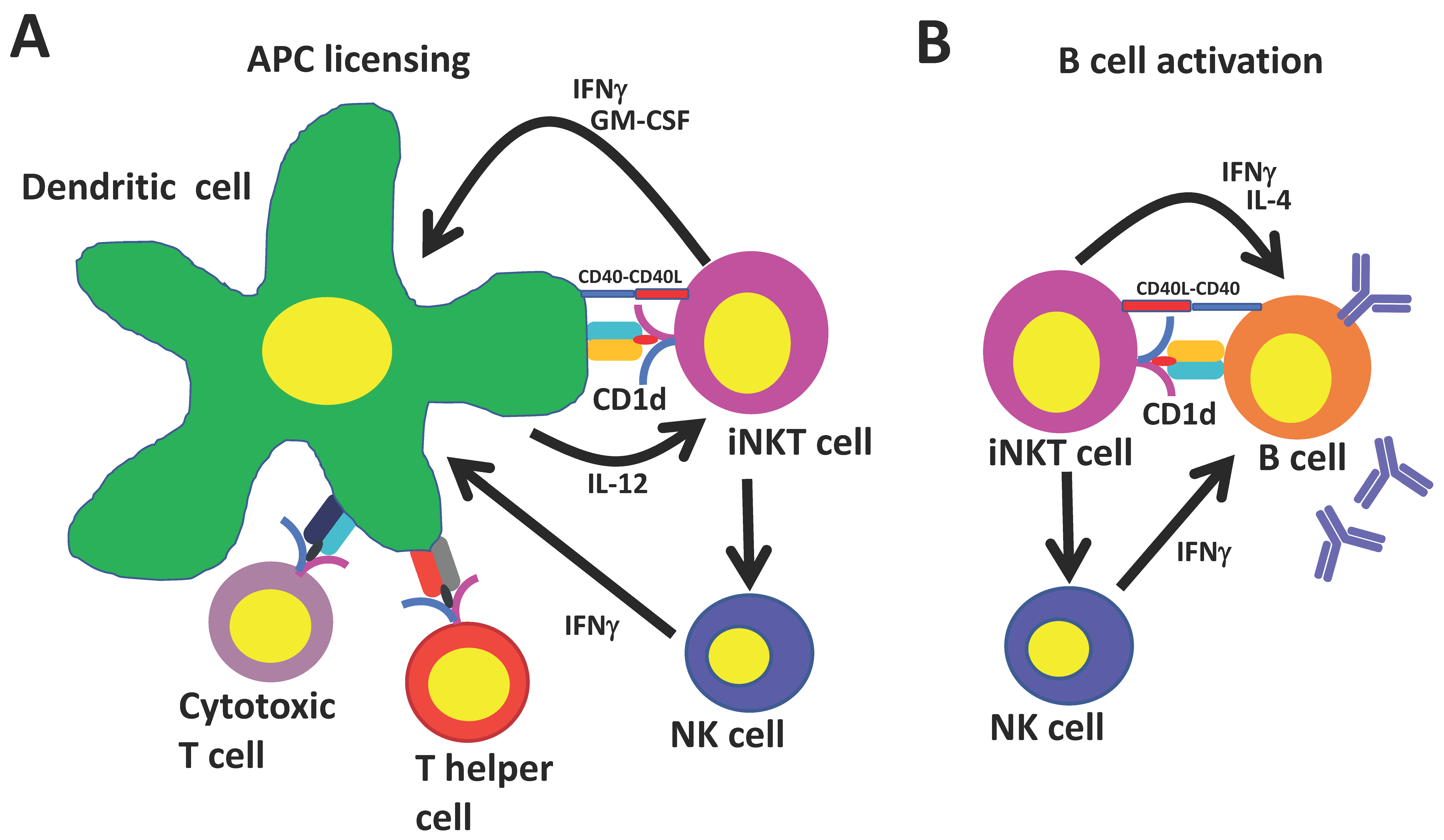
4. Universal Helper T Cell Functions of iNKT Cells
5. Adjuvanting Influenza Vaccines with iNKT Cell Agonists—A Brief History
6. Use of iNKT Cell Agonists as Adjuvants in Swine
7. Challenges to Applying iNKT Cell Agonists in Swine
8. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| iNKT | Invariant natural killer T |
| TCR | T cell receptor |
| MHC | Major histocompatibility complex |
| MAIT | Mucosal-associated invariant T |
| α-GalCer | α-galactosylceramide |
| DC | Dendritic cell |
| APC | Antigen-presenting cell |
| PRR | Pattern recognition receptor |
| PAMP | Pathogen-associated molecule pattern |
| IAV | Influenza A viruses |
| IAV-S | Swine influenza A virus |
| NP | Nucleoprotein |
| M1 or M2 | Matrix protein 1 or 2 |
| LAIV | Live attenuated influenza virus |
| TLR | Toll-like receptor |
| ILC | Innate lymphoid cell |
| IFNγ | Interferon gamma |
| BALF | Bronchoalveolar lavage fluid |
| TBLN | Tracheobronchial lymph node |
| CLN | Cervical lymph nodes |
| MLN | Mesenteric lymph nodes |
| PLZF | Promyelocytic leukaemia zinc finger protein |
| PMA | Phorbol myristate acetate |
| IL | Interleukin |
| TGF-β | Transforming growth factor beta |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IgG | Immunoglobulin G |
| PR8 HA | H1N1 strain A/Puerto Rico/8/34 hemagglutinin |
| i.n. | Intranasal |
| NALT | Nasal-associated lymphoid tissue |
| CTL | Cytotoxic T lymphocyte |
| SLP | Synthetic long-peptides |
| OVA | Ovalbumin |
| i.p. | Intraperitoneal |
| s.c. | Subcutaneous |
| i.m. | Intramuscular |
| i.v. | Intraveneous |
| CFA | Complete Freund’s Adjuvant |
| IDO | Indoleamine 2,3-dioxygenase |
| HEL | Hen egg lysozyme |
| UV | Ultraviolet |
| VAERD | Vaccine-associated enhanced respiratory disease |
References
- Greenwood, B. The contribution of vaccination to global health: Past, present and future. Philos Trans. R Soc. Lond B Biol. Sci. 2014, 369, 20130433. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.L.; Khader, S.A. Novel vaccine approaches for protection against intracellular pathogens. Curr. Opin. Immunol. 2014, 28, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hong, H.; Li, D.; Ma, S.; Di, Y.; Stoten, A.; Haig, N.; Di Gleria, K.; Yu, Z.; Xu, X.N.; et al. Comparing pooled peptides with intact protein for accessing cross-presentation pathways for protective CD8+ and CD4+ T cells. J. Biol. Chem. 2009, 284, 9184–9191. [Google Scholar] [CrossRef] [PubMed]
- Looringh van Beeck, F.A.; Reinink, P.; Hermsen, R.; Zajonc, D.M.; Laven, M.J.; Fun, A.; Troskie, M.; Schoemaker, N.J.; Morar, D.; Lenstra, J.A.; et al. Functional CD1d and/or NKT cell invariant chain transcript in horse, pig, African elephant and guinea pig, but not in ruminants. Mol. Immunol. 2009, 46, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Stankovic, S.; Baxter, A.G. Raising the NKT cell family. Nat. Immunol. 2010, 11, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Kawano, T.; Cui, J.; Koezuka, Y.; Toura, I.; Kaneko, Y.; Motoki, K.; Ueno, H.; Nakagawa, R.; Sato, H.; Kondo, E.; et al. CD1d-restricted and TCR-mediated activation of vα14 NKT cells by glycosylceramides. Science 1997, 278, 1626–1629. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.L.; Naidenko, O.V.; Gapin, L.; Nakayama, T.; Taniguchi, M.; Wang, C.R.; Koezuka, Y.; Kronenberg, M. Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. J. Exp. Med. 2000, 192, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.L.; Mallevaey, T.; Scott-Browne, J.; Gapin, L. CD1d-restricted iNKT cells, the ‘Swiss-Army knife’ of the immune system. Curr. Opin. Immunol. 2008, 20, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Carnaud, C.; Lee, D.; Donnars, O.; Park, S.H.; Beavis, A.; Koezuka, Y.; Bendelac, A. Cutting edge: Cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells. J. Immunol. 1999, 163, 4647–4650. [Google Scholar] [PubMed]
- Brigl, M.; Tatituri, R.V.; Watts, G.F.; Bhowruth, V.; Leadbetter, E.A.; Barton, N.; Cohen, N.R.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Innate and cytokine-driven signals, rather than microbial antigens, dominate in natural killer T cell activation during microbial infection. J. Exp. Med. 2011, 208, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Leadbetter, E.A.; Brigl, M.; Illarionov, P.; Cohen, N.; Luteran, M.C.; Pillai, S.; Besra, G.S.; Brenner, M.B. NK T cells provide lipid antigen-specific cognate help for B cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8339–8344. [Google Scholar] [CrossRef] [PubMed]
- Hermans, I.F.; Silk, J.D.; Gileadi, U.; Salio, M.; Mathew, B.; Ritter, G.; Schmidt, R.; Harris, A.L.; Old, L.; Cerundolo, V. NKT cells enhance CD4+ and CD8+ T cell responses to soluble antigen in vivo through direct interaction with dendritic cells. J. Immunol. 2003, 171, 5140–5147. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L.; Parekh, V.V.; Wu, L. The Response of CD1d-Restricted Invariant NKT Cells to Microbial Pathogens and Their Products. Front. Immunol. 2015, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Behar, S.M.; Porcelli, S.A. CD1-restricted T cells in host defense to infectious diseases. Curr. Top. Microbiol. Immunol. 2007, 314, 215–250. [Google Scholar] [PubMed]
- Tupin, E.; Kinjo, Y.; Kronenberg, M. The unique role of natural killer T cells in the response to microorganisms. Nat. Rev. Microbiol. 2007, 5, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.J.; Brigl, M.; Brenner, M.B. Invariant natural killer T cells: An innate activation scheme linked to diverse effector functions. Nat. Rev. Immunol. 2013, 13, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L.; Parekh, V.V.; Wu, L. Invariant NK T cells: Potential for immunotherapeutic targeting with glycolipid antigens. Immunotherapy 2011, 3, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Speir, M.; Hermans, I.F.; Weinkove, R. Engaging Natural Killer T Cells as ‘Universal Helpers’ for Vaccination. Drugs 2017, 77, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Artiaga, B.L.; Whitener, R.L.; Staples, C.R.; Driver, J.P. Adjuvant effects of therapeutic glycolipids administered to a cohort of NKT cell-diverse pigs. Vet. Immunol. Immunopathol. 2014, 162, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mair, K.H.; Sedlak, C.; Käser, T.; Pasternak, A.; Levast, B.; Gerner, W.; Saalmüller, A.; Summerfield, A.; Gerdts, V.; Wilson, H.L.; et al. The porcine innate immune system: An update. Dev. Comp. Immunol. 2014, 45, 321–343. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Sinkora, M.; Wertz, N.; Holtmeier, W.; Lemke, C.D. Development of the neonatal B and T cell repertoire in swine: Implications for comparative and veterinary immunology. Vet. Res. 2006, 37, 417–441. [Google Scholar] [CrossRef] [PubMed]
- Helke, K.L.; Swindle, M.M. Animal models of toxicology testing: The role of pigs. Expert Opin. Drug. Metab. Toxicol. 2013, 9, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Awada, L.; Brown, I.; Chen, H.; Claes, F.; Dauphin, G.; Donis, R.; Culhane, M.; Hamilton, K.; Lewis, N.; et al. Review of influenza A virus in swine worldwide: A call for increased surveillance and research. Zoonoses Public Health 2014, 61, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: A modelling study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef]
- Van Reeth, K.; van Gucht, S.; Pensaert, M. Investigations of the efficacy of European H1N1- and H3N2-based swine influenza vaccines against the novel H1N2 subtype. Vet. Rec. 2003, 153, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Lager, K.M.; Janke, B.H.; Gramer, M.R.; Richt, J.A. Failure of protection and enhanced pneumonia with a US H1N2 swine influenza virus in pigs vaccinated with an inactivated classical swine H1N1 vaccine. Vet. Microbiol. 2008, 126, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Lager, K.M.; Faaberg, K.S.; Harland, M.; Zanella, E.L.; Ciacci-Zanella, J.R.; Kehrli, M.E.; Janke, B.H.; Klimov, A. Experimental inoculation of pigs with pandemic H1N1 2009 virus and HI cross-reactivity with contemporary swine influenza virus antisera. Influenza Other Respir. Viruses 2010, 4, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Hoft, D.F.; Babusis, E.; Worku, S.; Spencer, C.T.; Lottenbach, K.; Truscott, S.M.; Abate, G.; Sakala, I.G.; Edwards, K.M.; Creech, C.B.; et al. Live and inactivated influenza vaccines induce similar humoral responses, but only live vaccines induce diverse T-cell responses in young children. J. Infect. Dis. 2011, 204, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Tosh, P.K.; Boyce, T.G.; Poland, G.A. Flu myths: Dispelling the myths associated with live attenuated influenza vaccine. Mayo Clin. Proc. 2008, 83, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lanthier, P.A.; Huston, G.E.; Moquin, A.; Eaton, S.M.; Szaba, F.M.; Kummer, L.W.; Tighe, M.P.; Kohlmeier, J.E.; Blair, P.J.; Broderick, M.; et al. Live attenuated influenza vaccine (LAIV) impacts innate and adaptive immune responses. Vaccine 2011, 29, 7849–7856. [Google Scholar] [CrossRef] [PubMed]
- Pena, L.; Vincent, A.L.; Loving, C.L.; Henningson, J.N.; Lager, K.M.; Li, W.; Perez, D.R. Strain-dependent effects of PB1-F2 of triple-reassortant H3N2 influenza viruses in swine. J. Gen. Virol. 2012, 93, 2204–2214. [Google Scholar] [CrossRef] [PubMed]
- Solórzano, A.; Webby, R.J.; Lager, K.M.; Janke, B.H.; García-Sastre, A.; Richt, J.A. Mutations in the NS1 protein of swine influenza virus impair anti-interferon activity and confer attenuation in pigs. J. Virol. 2005, 79, 7535–7543. [Google Scholar] [CrossRef] [PubMed]
- Masic, A.; Babiuk, L.A.; Zhou, Y. Reverse genetics-generated elastase-dependent swine influenza viruses are attenuated in pigs. J. Gen. Virol. 2009, 90, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Sandbulte, M.R.; Spickler, A.R.; Zaabel, P.K.; Roth, J.A. Optimal Use of Vaccines for Control of Influenza A Virus in Swine. Vaccines 2015, 3, 22–73. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; D’Oro, U.; Wack, A. Immunology of TLR-independent vaccine adjuvants. Curr. Opin. Immunol. 2009, 21, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Kool, M.; Willart, M.A.; Hammad, H. Mechanism of action of clinically approved adjuvants. Curr. Opin. Immunol. 2009, 21, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanyam, P.; Webb, T.J. Boosting the Immune Response: The Use of iNKT cell ligands as vaccine adjuvants. Front. Biol. 2012, 7, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Kharkwal, S.S.; Arora, P.; Porcelli, S.A. Glycolipid activators of invariant NKT cells as vaccine adjuvants. Immunogenetics 2016, 68, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.R.; Brennan, P.J.; Shay, T.; Watts, G.F.; Brigl, M.; Kang, J.; Brenner, M.B.; Consortium, I.P. Shared and distinct transcriptional programs underlie the hybrid nature of iNKT cells. Nat. Immunol. 2013, 14, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Engel, I.; Seumois, G.; Chavez, L.; Samaniego-Castruita, D.; White, B.; Chawla, A.; Mock, D.; Vijayanand, P.; Kronenberg, M. Innate-like functions of natural killer T cell subsets result from highly divergent gene programs. Nat. Immunol. 2016, 17, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Tsaih, S.W.; Serreze, D.V. Genetic control of murine invariant natural killer T-cell development dynamically differs dependent on the examined tissue type. Genes Immun. 2012, 13, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.T.; Benlagha, K.; Teyton, L.; Bendelac, A. Distinct functional lineages of human Vα24 natural killer T cells. J. Exp. Med. 2002, 195, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Gumperz, J.E.; Miyake, S.; Yamamura, T.; Brenner, M.B. Functionally distinct subsets of CD1d-restricted natural killer T cells revealed by CD1d tetramer staining. J. Exp. Med. 2002, 195, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Benlagha, K.; Weiss, A.; Beavis, A.; Teyton, L.; Bendelac, A. In vivo identification of glycolipid antigen-specific T cells using fluorescent CD1d tetramers. J. Exp. Med. 2000, 191, 1895–1903. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.T.; Johansson, C.; Olivares-Villagómez, D.; Singh, A.K.; Stanic, A.K.; Wang, C.R.; Joyce, S.; Wick, M.J.; van Kaer, L. The response of natural killer T cells to glycolipid antigens is characterized by surface receptor down-modulation and expansion. Proc. Natl. Acad. Sci. USA 2003, 100, 10913–10918. [Google Scholar] [CrossRef] [PubMed]
- Laloux, V.; Beaudoin, L.; Jeske, D.; Carnaud, C.; Lehuen, A. NK T cell-induced protection against diabetes in Vα14-Jα281 transgenic nonobese diabetic mice is associated with a Th2 shift circumscribed regionally to the islets and functionally to islet autoantigen. J. Immunol. 2001, 166, 3749–3756. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.G. Natural killer T cells within the liver: Conductors of the hepatic immune orchestra. Dig. Dis. 2010, 28, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Berzins, S.P.; Smyth, M.J.; Baxter, A.G. Presumed guilty: Natural killer T cell defects and human disease. Nat. Rev. Immunol. 2011, 11, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Artiaga, B.L.; Hackmann, T.J.; Samuel, M.S.; Walters, E.M.; Salek-Ardakani, S.; Driver, J.P. Targeted disruption of CD1d prevents NKT cell development in pigs. Mamm. Genome 2015, 26, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Eguchi-Ogawa, T.; Morozumi, T.; Tanaka, M.; Shinkai, H.; Okumura, N.; Suzuki, K.; Awata, T.; Uenishi, H. Analysis of the genomic structure of the porcine CD1 gene cluster. Genomics 2007, 89, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L.; Wu, L.; Joyce, S. Mechanisms and Consequences of Antigen Presentation by CD1. Trends Immunol. 2016, 37, 738–754. [Google Scholar] [CrossRef] [PubMed]
- Kasmar, A.; van Rhijn, I.; Moody, D.B. The evolved functions of CD1 during infection. Curr. Opin. Immunol. 2009, 21, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Thierry, A.; Robin, A.; Giraud, S.; Minouflet, S.; Barra, A.; Bridoux, F.; Hauet, T.; Touchard, G.; Herbelin, A.; Gombert, J.M. Identification of invariant natural killer T cells in porcine peripheral blood. Vet. Immunol. Immunopathol. 2012, 149, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, V.; Manickam, C.; Dhakal, S.; Binjawadagi, B.; Ouyang, K.; Hiremath, J.; Khatri, M.; Hague, J.G.; Lee, C.W.; Renukaradhya, G.J. Adjuvant effects of invariant NKT cell ligand potentiates the innate and adaptive immunity to an inactivated H1N1 swine influenza virus vaccine in pigs. Vet. Microbiol. 2016, 186, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Artiaga, B.L.; Yang, G.; Hackmann, T.J.; Liu, Q.; Richt, J.A.; Salek-Ardakani, S.; Castleman, W.L.; Lednicky, J.A.; Driver, J.P. Α-Galactosylceramide protects swine against influenza infection when administered as a vaccine adjuvant. Sci. Rep. 2016, 6, 23593. [Google Scholar] [CrossRef] [PubMed]
- Artiaga, B.L.; Yang, G.; Hutchinson, T.E.; Loeb, J.C.; Richt, J.A.; Lednicky, J.A.; Salek-Ardakani, S.; Driver, J.P. Rapid control of pandemic H1N1 influenza by targeting NKT-cells. Sci. Rep. 2016, 6, 37999. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Artiaga, B.L.; Lewis, S.T.; Driver, J.P. Characterizing porcine invariant natural killer T cells: A comparative study with NK cells and T cells. Dev. Comp. Immunol. 2017, 76, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Renukaradhya, G.J.; Manickam, C.; Khatri, M.; Rauf, A.; Li, X.; Tsuji, M.; Rajashekara, G.; Dwivedi, V. Functional invariant NKT cells in pig lungs regulate the airway hyperreactivity: A potential animal model. J. Clin. Immunol. 2011, 31, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Saalmüller, A.; Reddehase, M.J.; Bühring, H.J.; Jonjić, S.; Koszinowski, U.H. Simultaneous expression of CD4 and CD8 antigens by a substantial proportion of resting porcine T lymphocytes. Eur. J. Immunol. 1987, 17, 1297–1301. [Google Scholar] [CrossRef] [PubMed]
- Gerner, W.; Käser, T.; Saalmüller, A. Porcine T lymphocytes and NK cells—An update. Dev. Comp. Immunol. 2009, 33, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, A.; Rziha, H.J.; Saalmüller, A. Functional characterization of porcine CD4+CD8+ extrathymic T lymphocytes. Cell. Immunol. 1996, 168, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Ober, B.T.; Summerfield, A.; Mattlinger, C.; Wiesmüller, K.H.; Jung, G.; Pfaff, E.; Saalmüller, A.; Rziha, H.J. Vaccine-induced, pseudorabies virus-specific, extrathymic CD4+CD8+ memory T-helper cells in swine. J. Virol. 1998, 72, 4866–4873. [Google Scholar] [PubMed]
- Zuckermann, F.A. Extrathymic CD4/CD8 double positive T cells. Vet. Immunol. Immunopathol. 1999, 72, 55–66. [Google Scholar] [CrossRef]
- Gerner, W.; Denyer, M.S.; Takamatsu, H.H.; Wileman, T.E.; Wiesmüller, K.H.; Pfaff, E.; Saalmüller, A. Identification of novel foot-and-mouth disease virus specific T-cell epitopes in c/c and d/d haplotype miniature swine. Virus Res. 2006, 121, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Jiang, Z.; Chen, Z.; Gu, Y.; Liu, Y.; Zhang, X.; Cao, X. Pathogen-expanded CD11b+ invariant NKT cells feedback inhibit T cell proliferation via membrane-bound TGF-β1. J. Autoimmun. 2015, 58, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, A.; Kinebuchi, M.; Chen, H.Z.; Katabami, S.; Shimizu, T.; Hashimoto, Y.; Kikuchi, K.; Sato, N. NKT cells in the rat: Organ-specific distribution of NK T cells expressing distinct Vα14 chains. J. Immunol. 2000, 164, 3140–3148. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Nieda, M.; Koezuka, Y.; Nicol, A.; Porcelli, S.A.; Ishikawa, Y.; Tadokoro, K.; Hirai, H.; Juji, T. Analysis of human Vα24+ CD4+ NKT cells activated by α-glycosylceramide-pulsed monocyte-derived dendritic cells. J. Immunol. 2000, 164, 4458–4464. [Google Scholar] [CrossRef] [PubMed]
- Benlagha, K.; Kyin, T.; Beavis, A.; Teyton, L.; Bendelac, A. A thymic precursor to the NK T cell lineage. Science 2002, 296, 553–555. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Binns, R.M. Expression and regulation of the porcine CD44 molecule. Cell. Immunol. 1993, 149, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Coquet, J.M.; Chakravarti, S.; Kyparissoudis, K.; McNab, F.W.; Pitt, L.A.; McKenzie, B.S.; Berzins, S.P.; Smyth, M.J.; Godfrey, D.I. Diverse cytokine production by NKT cell subsets and identification of an IL-17-producing CD4−NK1.1− NKT cell population. Proc. Natl. Acad. Sci. USA 2008, 105, 11287–11292. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Teixeira, L.; Resende, M.; Coffre, M.; Devergne, O.; Herbeuval, J.P.; Hermine, O.; Schneider, E.; Rogge, L.; Ruemmele, F.M.; Dy, M.; et al. Proinflammatory environment dictates the IL-17-producing capacity of human invariant NKT cells. J. Immunol. 2011, 186, 5758–5765. [Google Scholar] [CrossRef] [PubMed]
- Paget, C.; Mallevaey, T.; Speak, A.O.; Torres, D.; Fontaine, J.; Sheehan, K.C.; Capron, M.; Ryffel, B.; Faveeuw, C.; Leite de Moraes, M.; et al. Activation of invariant NKT cells by toll-like receptor 9-stimulated dendritic cells requires type I interferon and charged glycosphingolipids. Immunity 2007, 27, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Salio, M.; Speak, A.O.; Shepherd, D.; Polzella, P.; Illarionov, P.A.; Veerapen, N.; Besra, G.S.; Platt, F.M.; Cerundolo, V. Modulation of human natural killer T cell ligands on TLR-mediated antigen-presenting cell activation. Proc. Natl. Acad. Sci. USA 2007, 104, 20490–20495. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.D.; Horwitz, D.A. Activated human NK cells can stimulate resting B cells to secrete immunoglobulin. J. Immunol. 1995, 154, 5656–5664. [Google Scholar] [PubMed]
- Vomhof-DeKrey, E.E.; Yates, J.; Leadbetter, E.A. Invariant NKT cells provide innate and adaptive help for B cells. Curr. Opin. Immunol. 2014, 28, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.; Pittoni, P.; Tonti, E.; Malzone, C.; Uematsu, Y.; Tortoli, M.; Maione, D.; Volpini, G.; Finco, O.; Nuti, S.; et al. Invariant NKT cells sustain specific B cell responses and memory. Proc. Natl. Acad. Sci. USA 2007, 104, 3984–3989. [Google Scholar] [CrossRef] [PubMed]
- Barral, P.; Eckl-Dorna, J.; Harwood, N.E.; De Santo, C.; Salio, M.; Illarionov, P.; Besra, G.S.; Cerundolo, V.; Batista, F.D. B cell receptor-mediated uptake of CD1d-restricted antigen augments antibody responses by recruiting invariant NKT cell help in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 8345–8350. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.A.; Exley, M.A.; Lang, M.L. The CD1d-binding glycolipid α-galactosylceramide enhances humoral immunity to T-dependent and T-independent antigen in a CD1d-dependent manner. Immunology 2006, 119, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Gaya, M.; Barral, P.; Burbage, M.; Aggarwal, S.; Montaner, B.; Warren Navia, A.; Aid, M.; Tsui, C.; Maldonado, P.; Nair, U.; et al. Initiation of Antiviral B Cell Immunity Relies on Innate Signals from Spatially Positioned NKT Cells. Cell 2017. [Google Scholar] [CrossRef] [PubMed]
- Long, H.M.; Chagoury, O.L.; Leese, A.M.; Ryan, G.B.; James, E.; Morton, L.T.; Abbott, R.J.; Sabbah, S.; Kwok, W.; Rickinson, A.B. MHC II tetramers visualize human CD4+ T cell responses to Epstein-Barr virus infection and demonstrate atypical kinetics of the nuclear antigen EBNA1 response. J. Exp. Med. 2013, 210, 933–949. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fujio, M.; Imamura, M.; Wu, D.; Vasan, S.; Wong, C.H.; Ho, D.D.; Tsuji, M. Design of a potent CD1d-binding NKT cell ligand as a vaccine adjuvant. Proc. Natl. Acad. Sci. USA 2010, 107, 13010–13015. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, M.; Persson, J.; Thörn, K.; Harandi, A.M. The mucosal adjuvant effect of α-galactosylceramide for induction of protective immunity to sexually transmitted viral infection. J. Immunol. 2009, 182, 6435–6443. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.Y.; Ko, H.J.; Chang, W.S.; Park, S.H.; Kweon, M.N.; Kang, C.Y. α-Galactosylceramide can act as a nasal vaccine adjuvant inducing protective immune responses against viral infection and tumor. J. Immunol. 2005, 175, 3309–3317. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.J.; Ko, S.Y.; Lee, K.A.; Ko, H.J.; Lee, Y.S.; Fujihashi, K.; Boyaka, P.N.; Kim, S.H.; Horimoto, T.; Kweon, M.N.; et al. A single intranasal immunization with inactivated influenza virus and α-galactosylceramide induces long-term protective immunity without redirecting antigen to the central nervous system. Vaccine 2007, 25, 5189–5198. [Google Scholar] [CrossRef] [PubMed]
- Kamijuku, H.; Nagata, Y.; Jiang, X.; Ichinohe, T.; Tashiro, T.; Mori, K.; Taniguchi, M.; Hase, K.; Ohno, H.; Shimaoka, T.; et al. Mechanism of NKT cell activation by intranasal coadministration of α-galactosylceramide, which can induce cross-protection against influenza viruses. Mucosal. Immunol. 2008, 1, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Kopecky-Bromberg, S.A.; Fraser, K.A.; Pica, N.; Carnero, E.; Moran, T.M.; Franck, R.W.; Tsuji, M.; Palese, P. α-C-galactosylceramide as an adjuvant for a live attenuated influenza virus vaccine. Vaccine 2009, 27, 3766–3774. [Google Scholar] [CrossRef] [PubMed]
- Mibayashi, M.; Martínez-Sobrido, L.; Loo, Y.M.; Cárdenas, W.B.; Gale, M.; García-Sastre, A. Inhibition of retinoic acid-inducible gene I-mediated induction of β interferon by the NS1 protein of influenza A virus. J. Virol. 2007, 81, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Ma, L.C.; Xiao, R.; Radvansky, B.; Aramini, J.; Zhao, L.; Marklund, J.; Kuo, R.L.; Twu, K.Y.; Arnold, E.; et al. Structural basis for suppression of a host antiviral response by influenza A virus. Proc. Natl. Acad. Sci. USA 2008, 105, 13093–13098. [Google Scholar] [CrossRef] [PubMed]
- Kochs, G.; García-Sastre, A.; Martínez-Sobrido, L. Multiple anti-interferon actions of the influenza A virus NS1 protein. J. Virol. 2007, 81, 7011–7021. [Google Scholar] [CrossRef] [PubMed]
- Noah, D.L.; Twu, K.Y.; Krug, R.M. Cellular antiviral responses against influenza A virus are countered at the posttranscriptional level by the viral NS1A protein via its binding to a cellular protein required for the 3′ end processing of cellular pre-mRNAS. Virology 2003, 307, 386–395. [Google Scholar] [CrossRef]
- Nemeroff, M.E.; Barabino, S.M.; Li, Y.; Keller, W.; Krug, R.M. Influenza virus NS1 protein interacts with the cellular 30 kDa subunit of CPSF and inhibits 3’end formation of cellular pre-mRNAs. Mol. Cell. 1998, 1, 991–1000. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, K.A.; Lee, J.Y.; Kang, M.H.; Song, Y.C.; Baek, D.J.; Kim, S.; Kang, C.Y. An α-GalCer analogue with branched acyl chain enhances protective immune responses in a nasal influenza vaccine. Vaccine 2011, 29, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.J.; Li, J.; Kedzierski, L.; Compton, B.J.; Hayman, C.M.; Osmond, T.L.; Tang, C.W.; Farrand, K.J.; Koay, H.F.; Almeida, C.F.D.S.; et al. Augmenting Influenza-Specific T Cell Memory Generation with a Natural Killer T Cell-Dependent Glycolipid-Peptide Vaccine. ACS Chem. Biol. 2017, 12, 2898–2905. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S. Heterosubtypic T-Cell Immunity to Influenza in Humans: Challenges for Universal T-Cell Influenza Vaccines. Front. Immunol. 2016, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Guillonneau, C.; Mintern, J.D.; Hubert, F.X.; Hurt, A.C.; Besra, G.S.; Porcelli, S.; Barr, I.G.; Doherty, P.C.; Godfrey, D.I.; Turner, S.J. Combined NKT cell activation and influenza virus vaccination boosts memory CTL generation and protective immunity. Proc. Natl. Acad. Sci. USA 2009, 106, 3330–3335. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.T.; Tsai, Y.C.; Lin, W.D.; Jan, J.T.; Lin, K.H.; Huang, J.R.; Cheng, J.Y.; Chen, M.W.; Wong, C.H.; Yu, A.L. Potent adjuvant effects of novel NKT stimulatory glycolipids on hemagglutinin based DNA vaccine for H5N1 influenza virus. Antivir. Res. 2014, 107, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Fotouhi, F.; Shaffifar, M.; Farahmand, B.; Shirian, S.; Saeidi, M.; Tabarraei, A.; Gorji, A.; Ghaemi, A. Adjuvant use of the NKT cell agonist α-galactosylceramide leads to enhancement of M2-based DNA vaccine immunogenicity and protective immunity against influenza A virus. Arch. Virol. 2017, 162, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.S.; Jegaskanda, S.; Godfrey, D.I.; Kent, S.J. In-vivo stimulation of macaque natural killer T cells with α-galactosylceramide. Clin. Exp. Immunol. 2013, 173, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Oki, S.; Chiba, A.; Yamamura, T.; Miyake, S. The clinical implication and molecular mechanism of preferential IL-4 production by modified glycolipid-stimulated NKT cells. J. Clin. Investig. 2004, 113, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Miyake, S.; Yamamura, T. A synthetic glycolipid prevents autoimmune encephalomyelitis by inducing TH2 bias of natural killer T cells. Nature 2001, 413, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Schmieg, J.; Yang, G.; Franck, R.W.; Tsuji, M. Superior protection against malaria and melanoma metastases by a C-glycoside analogue of the natural killer T cell ligand α-galactosylceramide. J. Exp. Med. 2003, 198, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.A.; Nagarajan, N.A.; Wingender, G.; Wang, J.; Scott, I.; Tsuji, M.; Franck, R.W.; Porcelli, S.A.; Zajonc, D.M.; Kronenberg, M. Mechanisms for glycolipid antigen-driven cytokine polarization by Vα14i NKT cells. J. Immunol. 2010, 184, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Giaccone, G.; Punt, C.J.; Ando, Y.; Ruijter, R.; Nishi, N.; Peters, M.; von Blomberg, B.M.; Scheper, R.J.; van der Vliet, H.J.; van den Eertwegh, A.J.; et al. A phase I study of the natural killer T-cell ligand α-galactosylceramide (KRN7000) in patients with solid tumors. Clin. Cancer Res. 2002, 8, 3702–3709. [Google Scholar] [PubMed]
- Parekh, V.V.; Wilson, M.T.; Olivares-Villagomez, D.; Singh, A.K.; Wu, L.; Wang, C.R.; Joyce, S.; van Kaer, L. Glycolipid antigen induces long-term natural killer T cell anergy in mice. J. Clin. Investig. 2005, 115, 2572–2583. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.A.; Kronenberg, M. Activation or anergy: NKT cells are stunned by α-galactosylceramide. J. Clin. Investig. 2005, 115, 2328–2329. [Google Scholar] [CrossRef] [PubMed]
- Uldrich, A.P.; Crowe, N.Y.; Kyparissoudis, K.; Pellicci, D.G.; Zhan, Y.; Lew, A.M.; Bouillet, P.; Strasser, A.; Smyth, M.J.; Godfrey, D.I. NKT cell stimulation with glycolipid antigen in vivo: Costimulation-dependent expansion, Bim-dependent contraction, and hyporesponsiveness to further antigenic challenge. J. Immunol. 2005, 175, 3092–3101. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Loving, C.L.; Manischewitz, J.; King, L.R.; Gauger, P.C.; Henningson, J.; Vincent, A.L.; Golding, H. Vaccine-induced anti-HA2 antibodies promote virus fusion and enhance influenza virus respiratory disease. Sci. Transl. Med. 2013, 5, 200ra114. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Strain | Vaccination | Details | NKT Cell Agonist (Dose per Mouse) | Ref. | |
|---|---|---|---|---|---|
| Route | Strain/Subunit | ||||
| BALB/c | i.n. | H1N1 PR8 | Immunization with PR8 HA antigen with αGC 3 times at 1-week intervals, infection with 20 LD50 PR8 2 weeks after final immunization | αGC (0.125, 0.5, 2 µg) | [85] |
| BALB/c | i.n. | H1N1 PR8 | Immunization with inactivated PR8 with αGC, infection with 20 LD50 PR8 2 weeks and 3 months after immunization | αGC (0.5 µg) | [86] |
| BALB/c | i.n./i.m. | H1N1 PR8 H1N1A/Yamagata H3N2A/Guizhou B/Ibaraki | Immunization with PR8, A/Yamagata, A/Guizhou, or B/Ibaraki HA vaccine with αGC twice at 4-week apart, infection with 40 LD50 PR8 2 weeks after second immunization | αGC (2 µg) | [87] |
| BALB/c | i.n. | H1N1 PR8 | Immunization with PR8 with αGC twice at 4-week apart, infection with 40 LD50 A/Yamagata, A/Guizhou, or B/Ibaraki 2 weeks after second immunization | αGC (2 µg) | [87] |
| BALB/c | i.n. | H5N1 NIBRG14 | Immunization with NIBRG14 inactivated vaccine with αGC twice at 4-week apart, infection with 103 PFU of A/Vietnam or A/HK483 influenza virus 2 weeks after second immunization | αGC (2 µg) | [87] |
| BALB/c | i.n. | H1N1 rNS1 1-73 | Immunization with rNS1 1-73 with different amounts of α-C-GC, infection with 100 LD50 PR8 3 weeks after immunization | α-C-GC (0.11, 0.33, 1, 2, 3, 4 µg) | [88] |
| BALB/c | i.n. | H1N1 PR8 | Immunization with inactivated PR8 with αGC or αGC analogues, infection with 5 LD50 PR8 4 weeks after immunization or 100 LD50 PR8 5 weeks after immunization | αGC (0.5 µg) KBC-007 (0.5 µg) KBC-009 (0.5 µg) | [94] |
| C57BL/6 | i.v. | H1N1 PR8-OVA257 | Immunization with SLP-conjugated vaccine PR8-OVA257 with αGC, infection with 104 PFU HKx31-OVA257 6–8 weeks after immunization | αGC (76 ng) | [95] |
| BALB/c CD1 C3H/JeJ | i.p., s.c., i.m., i.v. | HA/NA from H3N2 PNM07 | Immunization with PNM07 protein with αGC for twice at 2-week apart, analysis of protein-specific antibodies | αGC (0.1 µg) | [78] |
| C57BL/6 | i.m. | H1N1 NC20 | Immunization with NC20 protein with αGC twice at 2-week apart, infection with 100 LD50 H1N1 A/WS/33 2 weeks after second immunization | αGC (0.1 µg) | [78] |
| C57BL/6 | i.m. | H3N2 PNM07 | Immunization with H3N2 PNM07 protein with αGC twice at 0 and 2 weeks, boosted with PNM07 at 30 weeks | αGC (0.1 µg) | [78] |
| C57BL/6 | s.c. | H1N1 PR8 | Immunization with inactivated PR8 with αGC, infection with 104 PFU of live H3N2 HKx31 6 weeks after immunization | αGC (1 µg) | [97] |
| BALB/c | i.m. | pCHA5 for H5N1 | Immunization with pCHA5 with C34 with αGC for twice at 3-week apart, infection with 200 LD50 NIBRG14 2 weeks after immunization | C34 (2 µg) | [98] |
| BALB/c | i.m. | DNA vaccine encoding M2 | Immunization with DNA vaccine encoding M2 with αGC for 3 times at 2-week intervals, infection with 1 LD90 PR8 2 weeks after final immunization | αGC (1 µg) | [99] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Richt, J.A.; Driver, J.P. Harnessing Invariant NKT Cells to Improve Influenza Vaccines: A Pig Perspective. Int. J. Mol. Sci. 2018, 19, 68. https://doi.org/10.3390/ijms19010068
Yang G, Richt JA, Driver JP. Harnessing Invariant NKT Cells to Improve Influenza Vaccines: A Pig Perspective. International Journal of Molecular Sciences. 2018; 19(1):68. https://doi.org/10.3390/ijms19010068
Chicago/Turabian StyleYang, Guan, Jürgen A. Richt, and John P. Driver. 2018. "Harnessing Invariant NKT Cells to Improve Influenza Vaccines: A Pig Perspective" International Journal of Molecular Sciences 19, no. 1: 68. https://doi.org/10.3390/ijms19010068
APA StyleYang, G., Richt, J. A., & Driver, J. P. (2018). Harnessing Invariant NKT Cells to Improve Influenza Vaccines: A Pig Perspective. International Journal of Molecular Sciences, 19(1), 68. https://doi.org/10.3390/ijms19010068