Successive Onset of Molecular, Cellular and Tissue-Specific Responses in Midgut Gland of Littorina littorea Exposed to Sub-Lethal Cadmium Concentrations

 ,
, 
Abstract
:
1. Introduction
2. Results
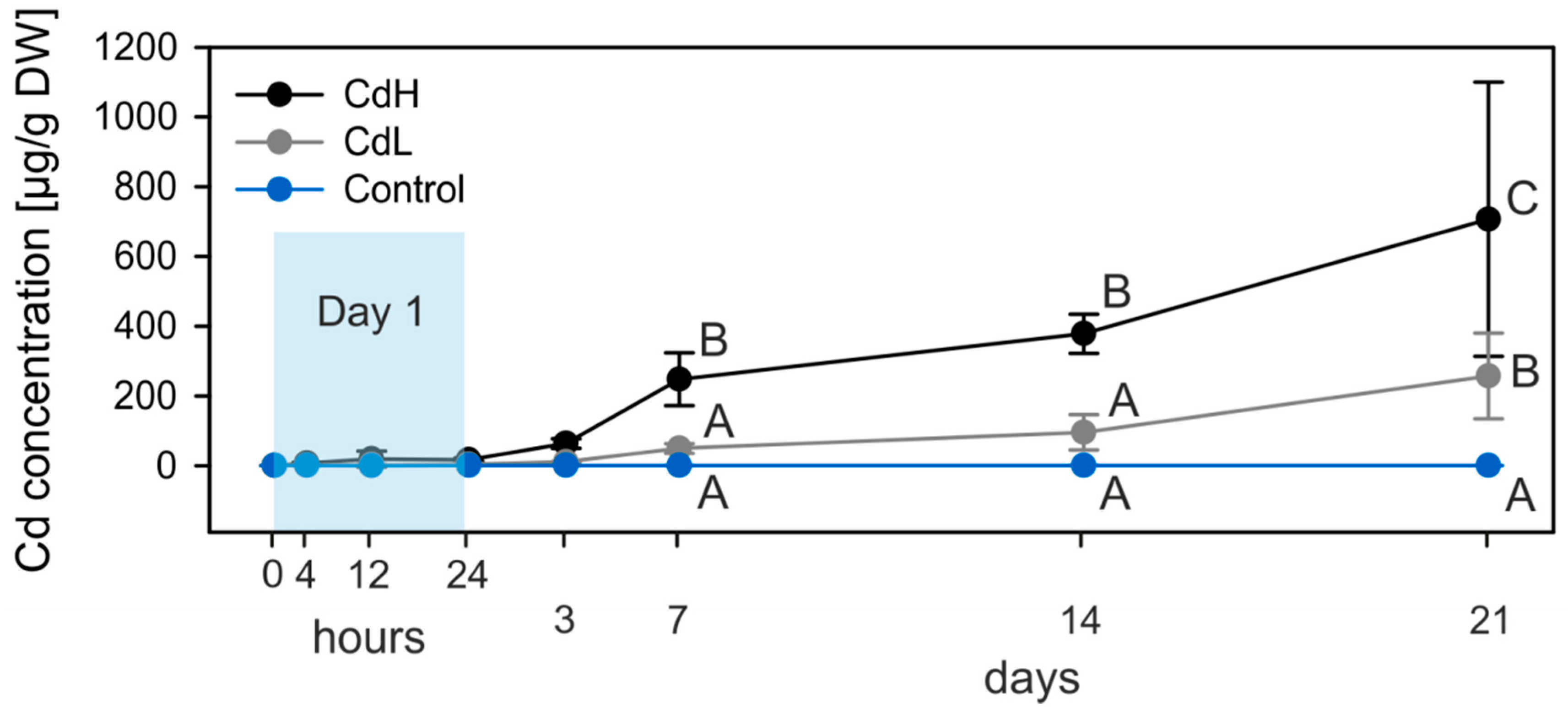
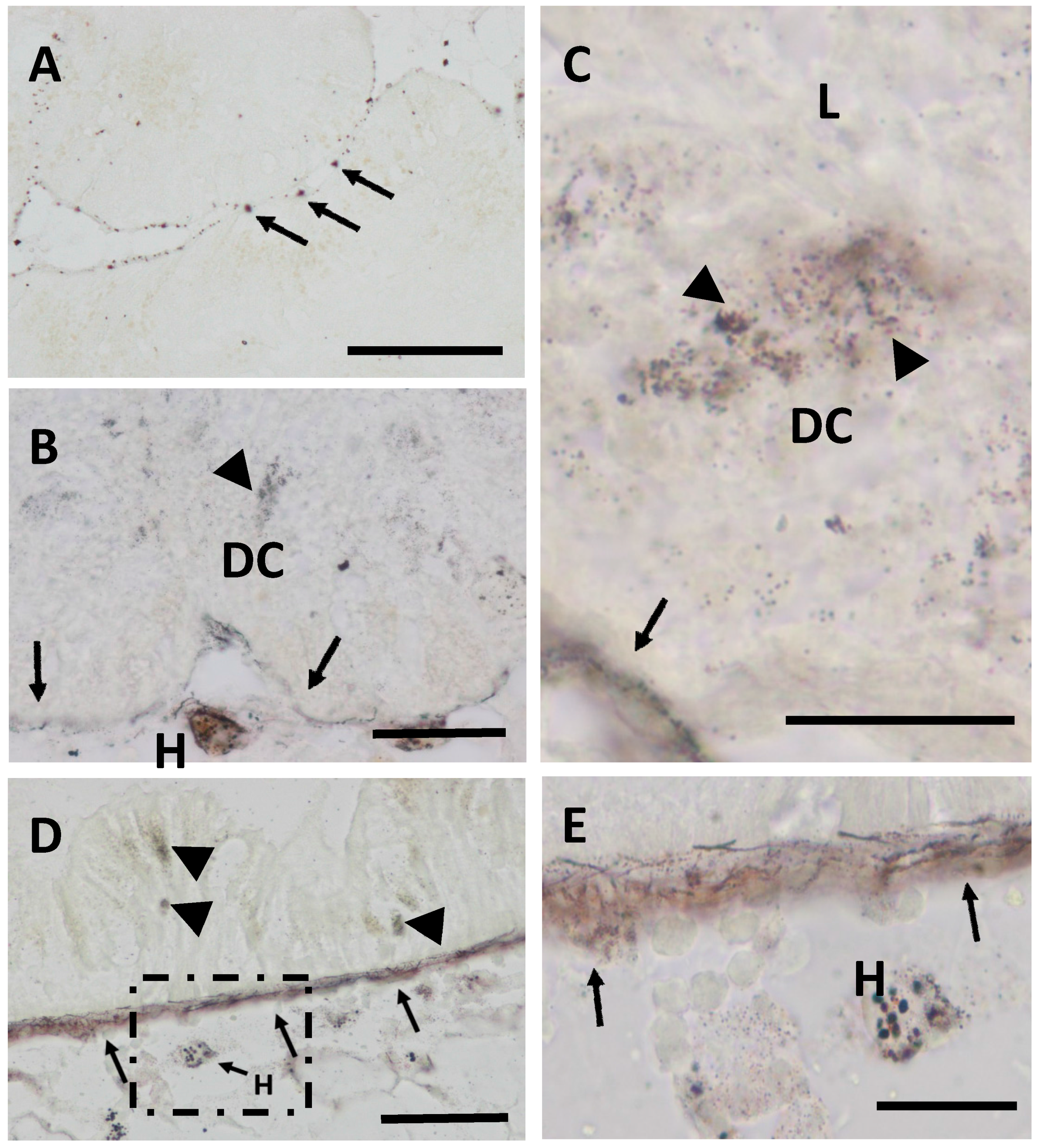
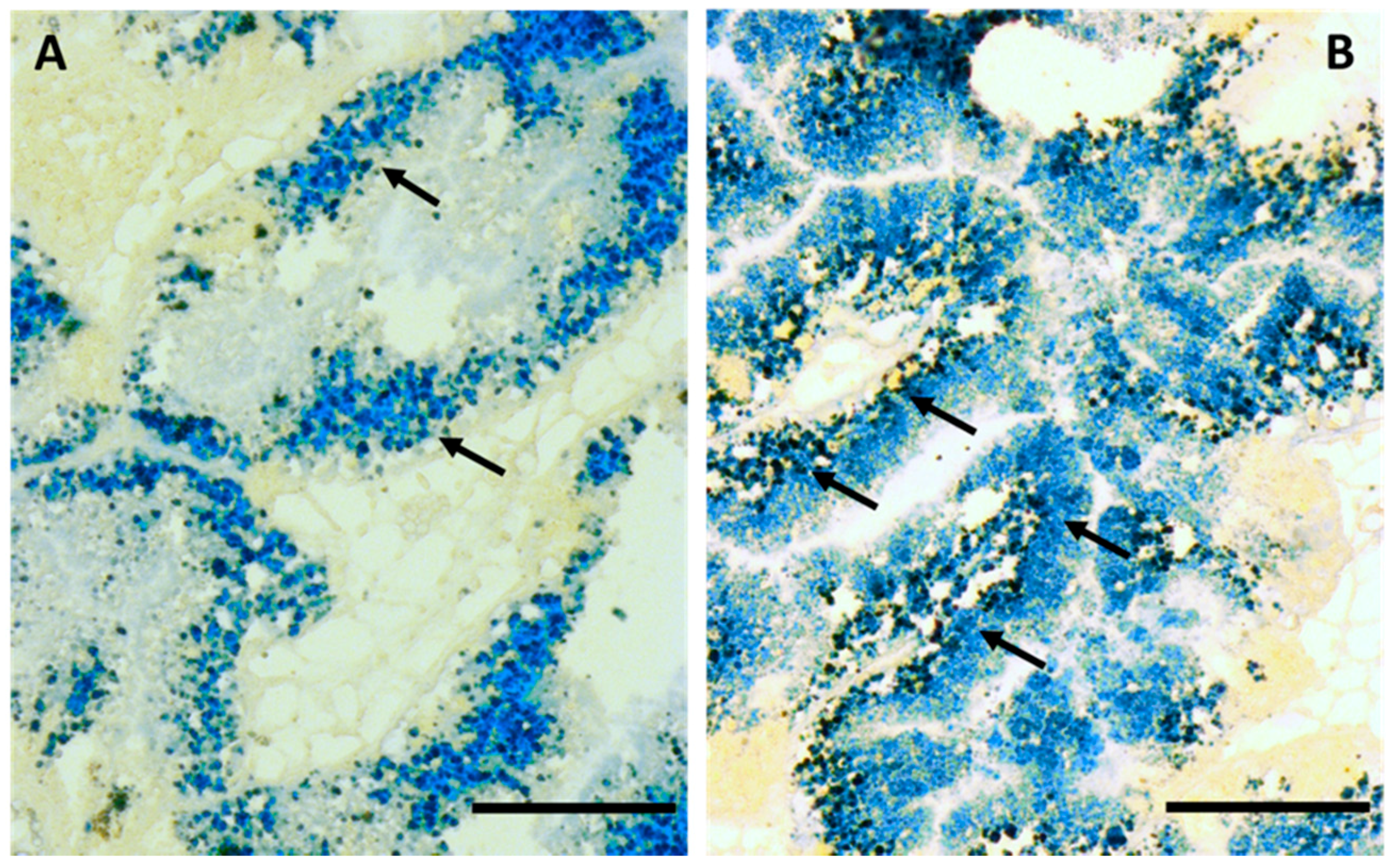
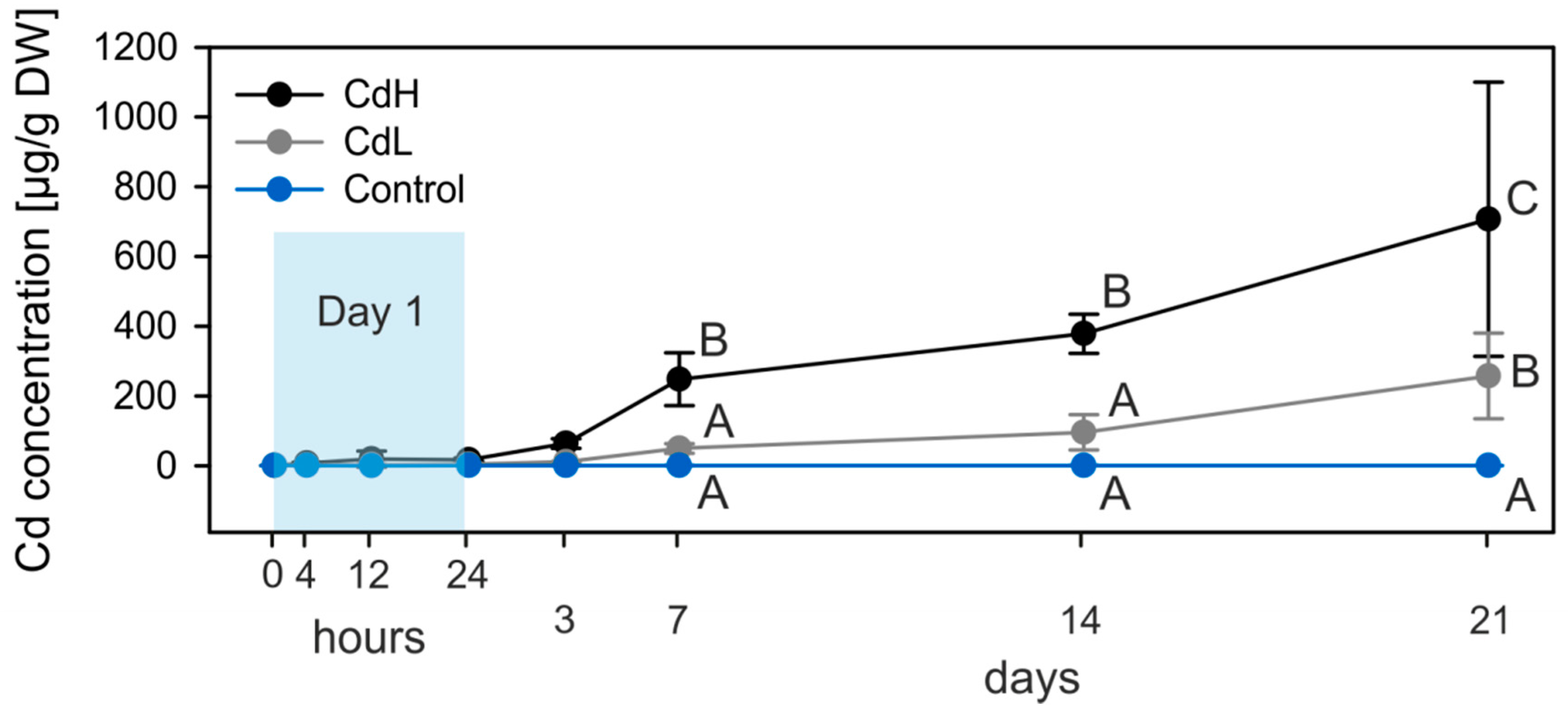
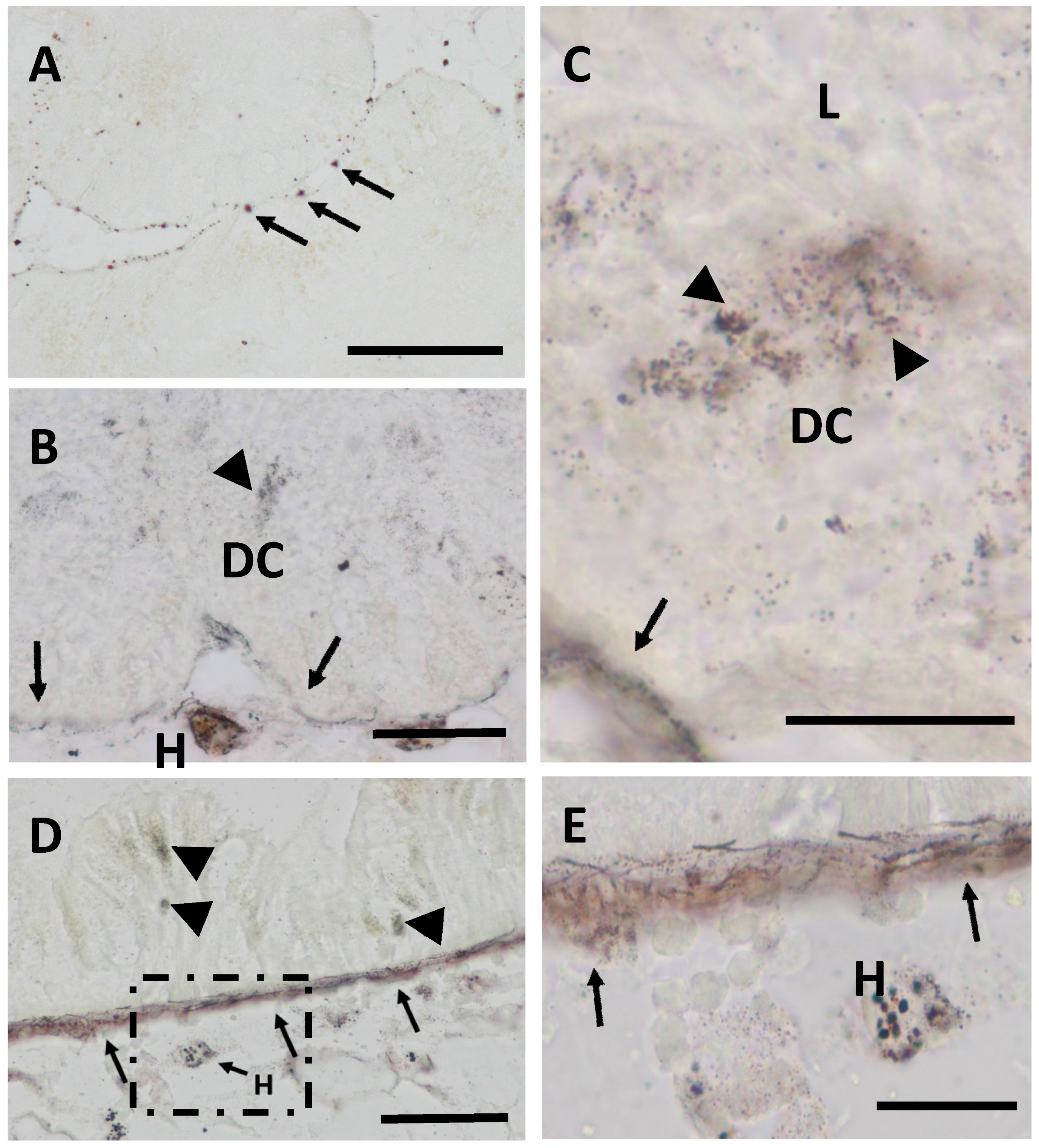
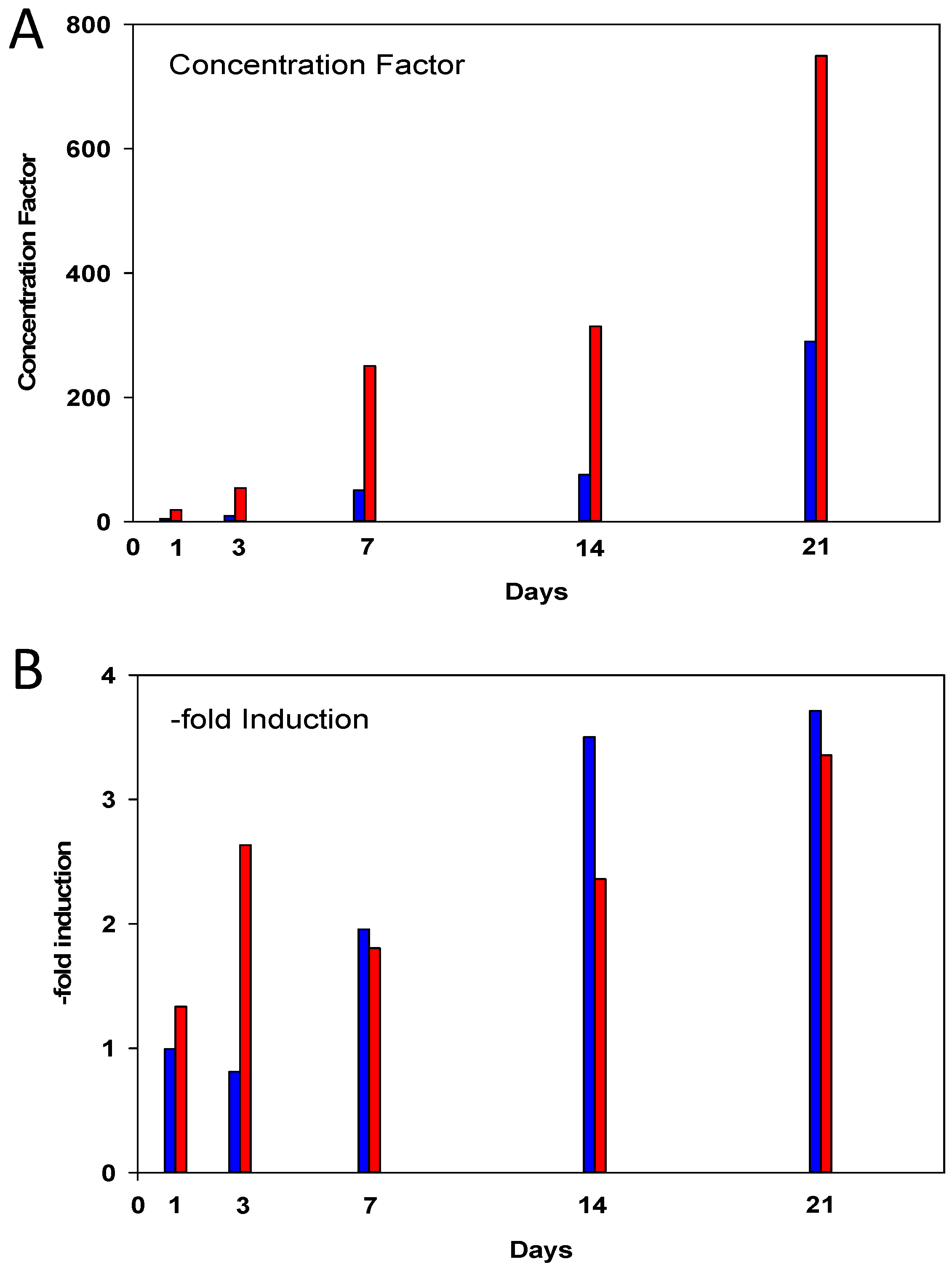
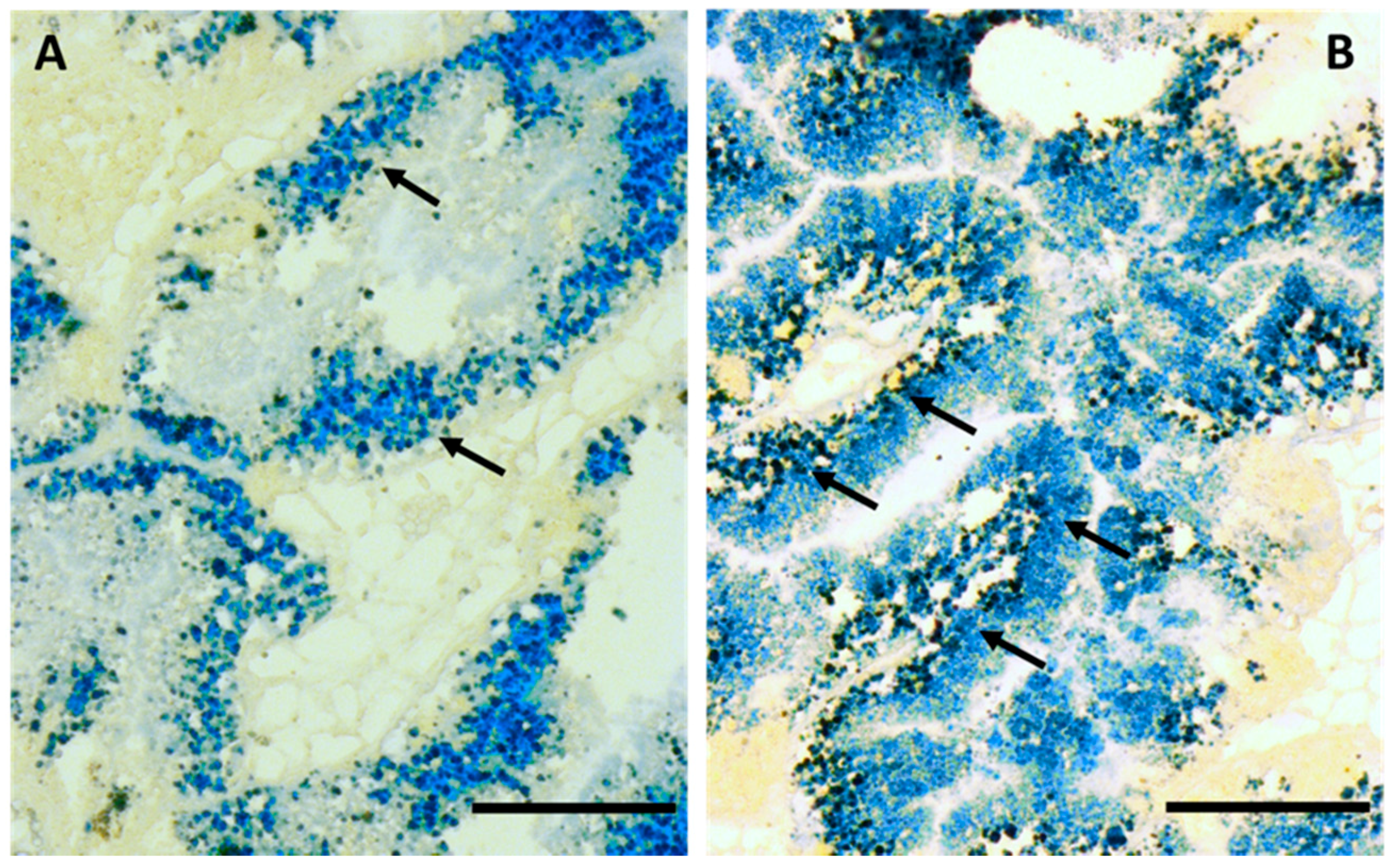
2.1. Cd Accumulation and Black Silver Deposits (BSD) in the Midgut Gland
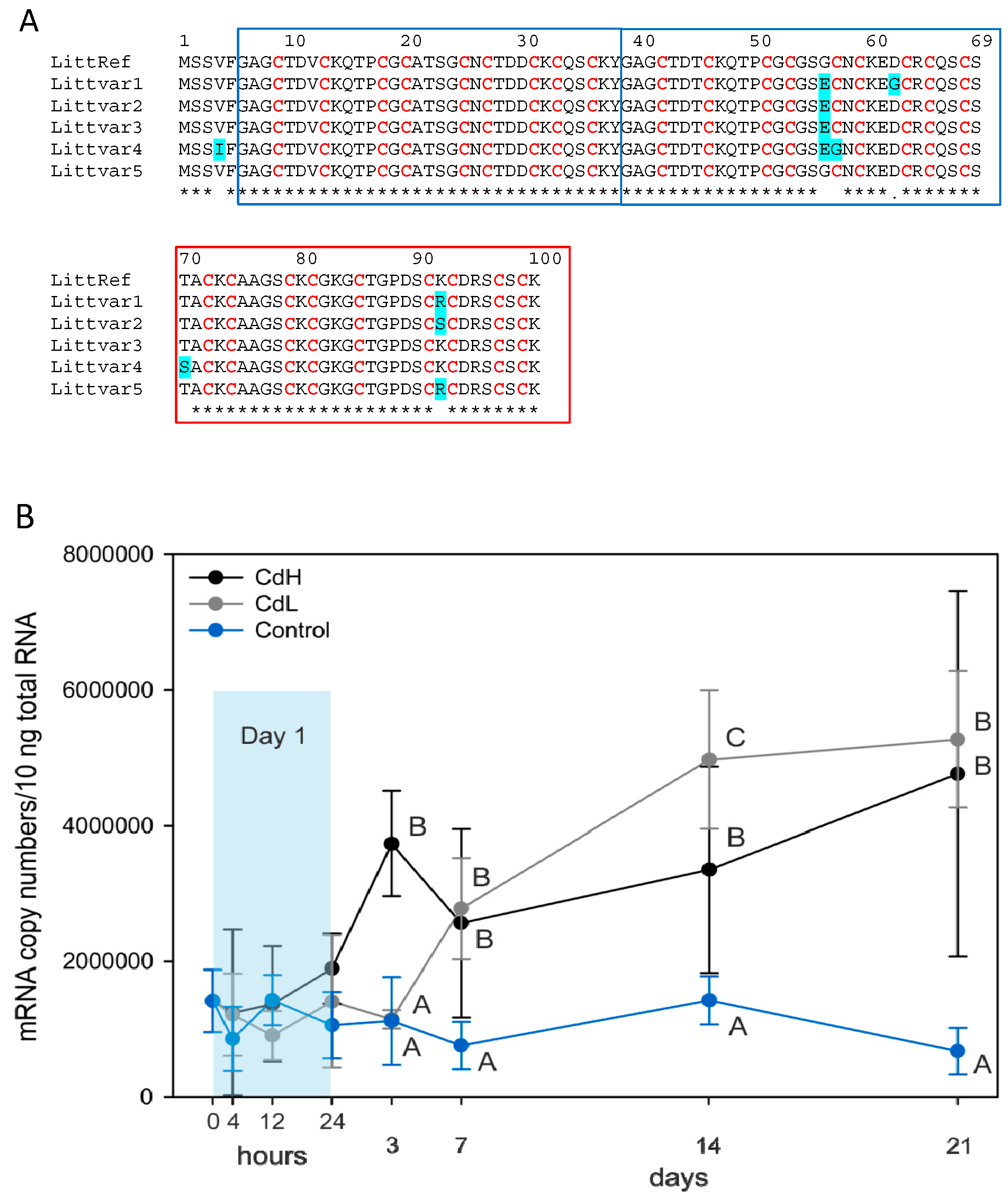
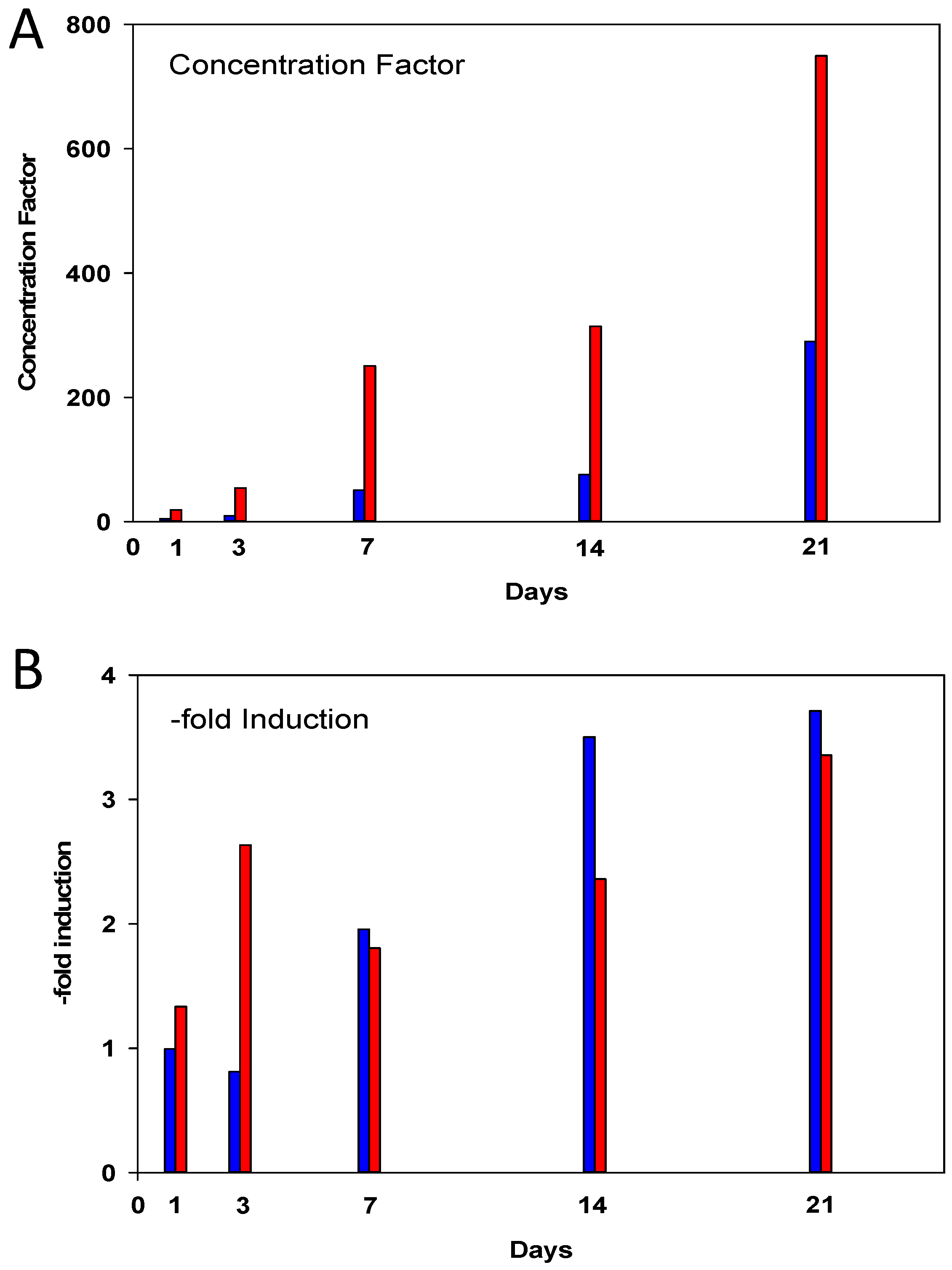
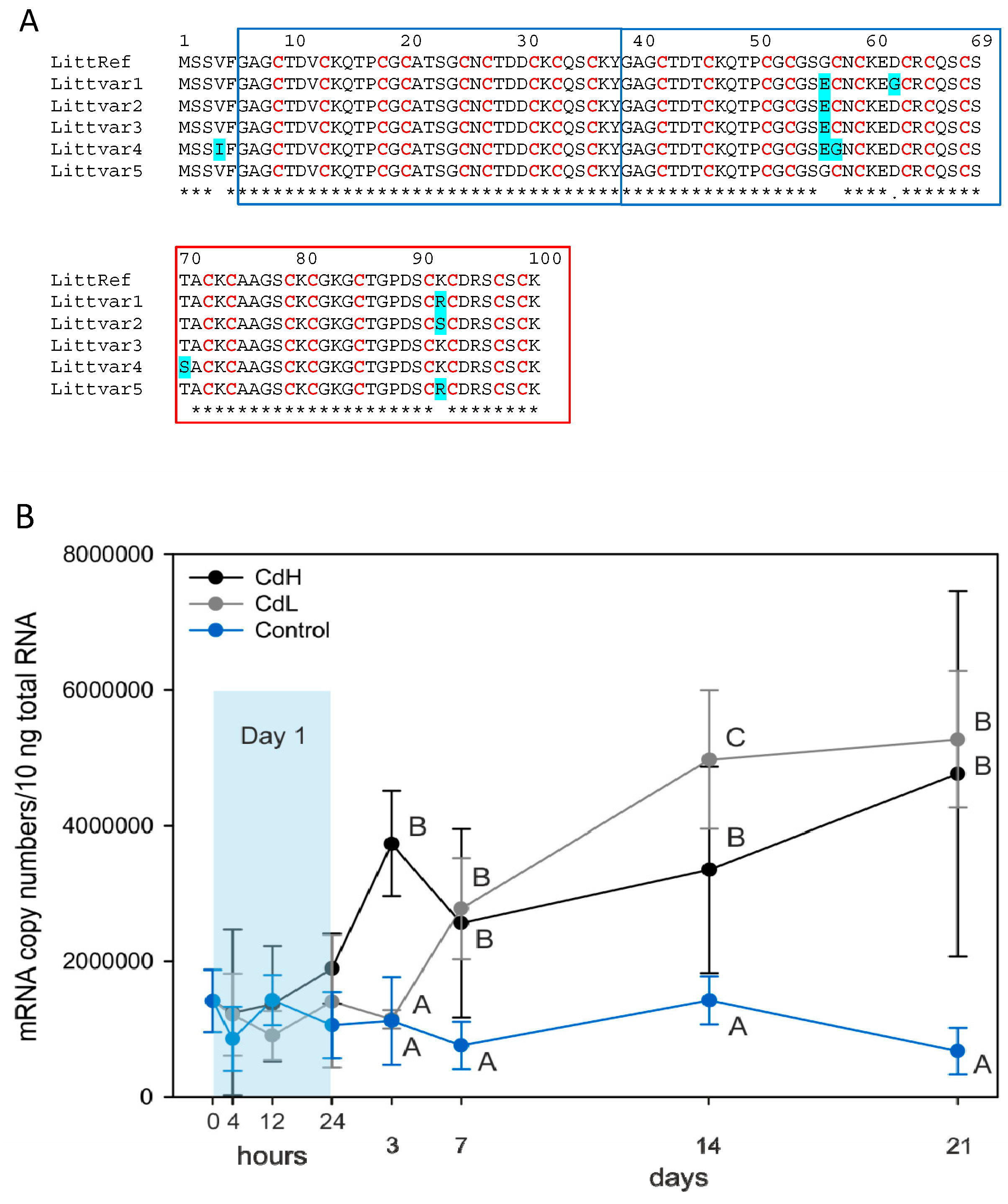
2.2. Increase of Metallothionein mRNA Concentration
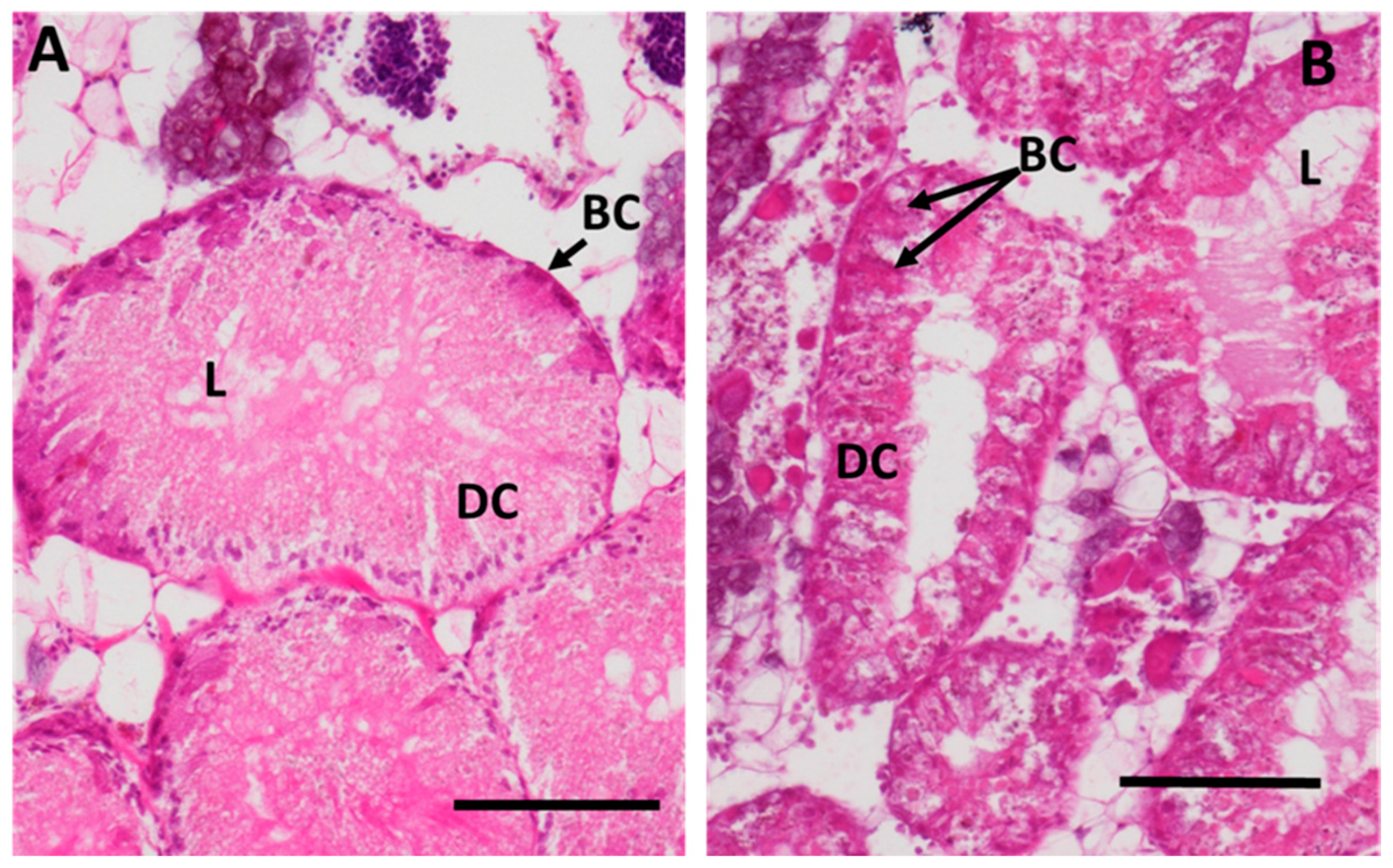
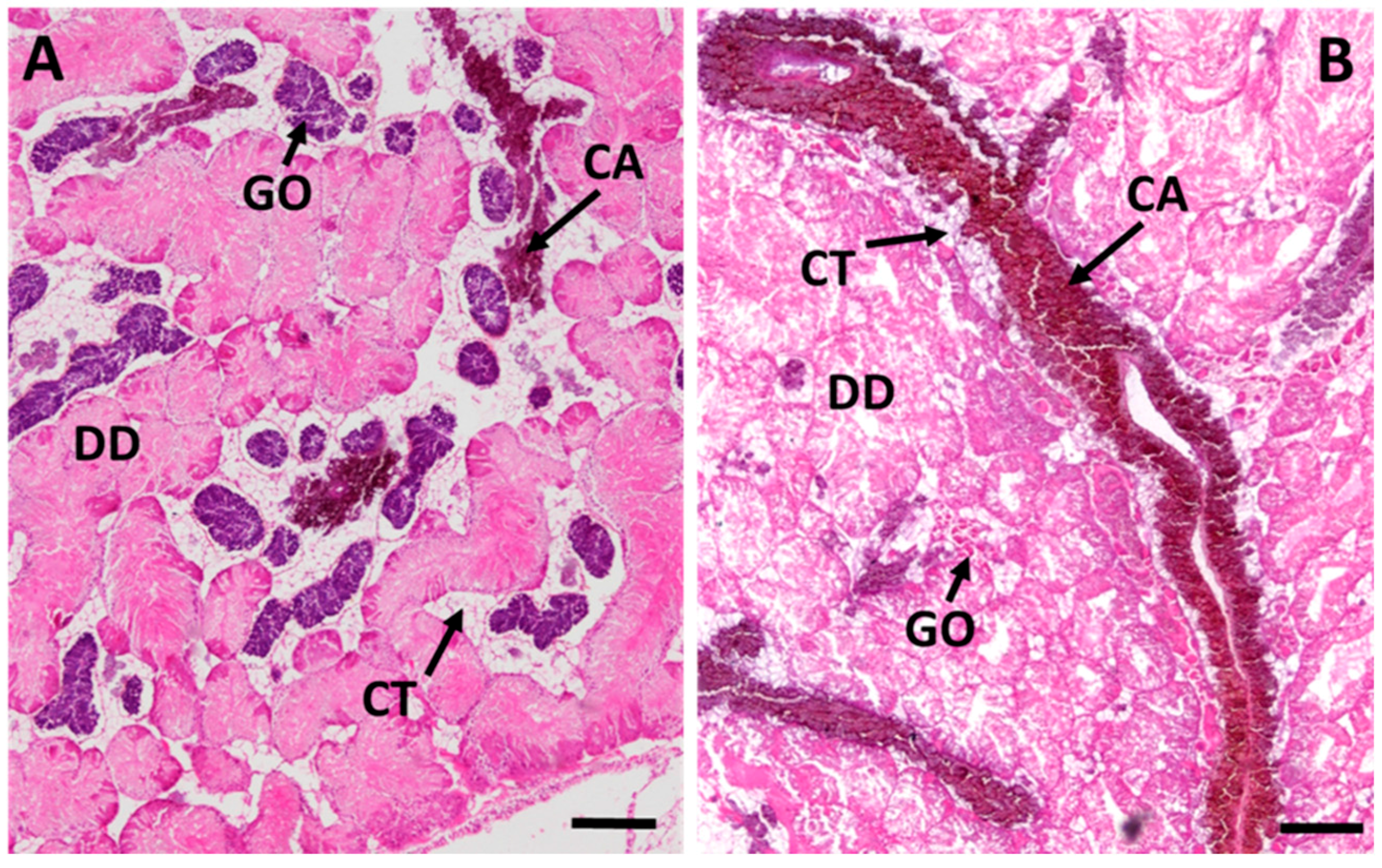
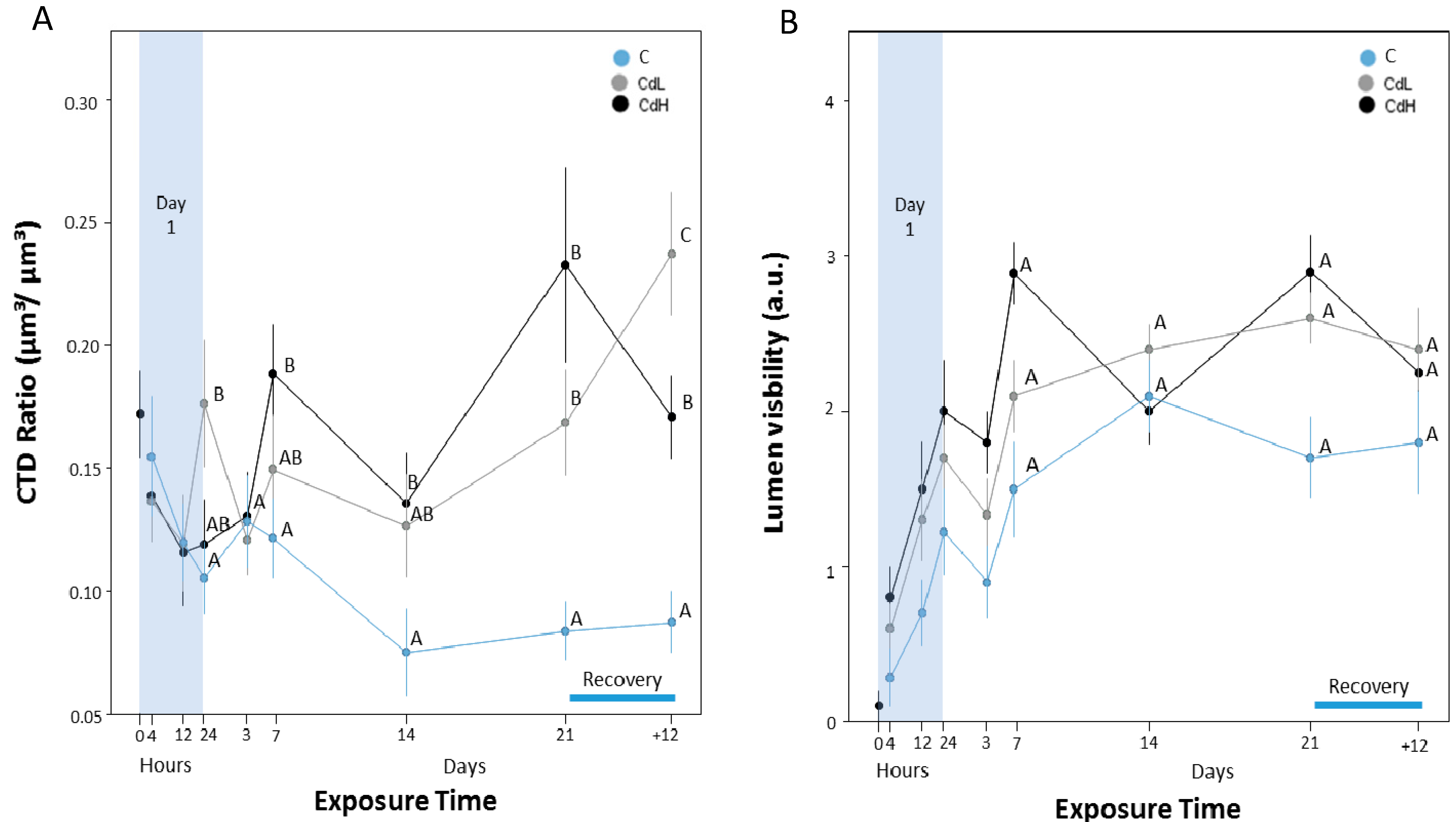
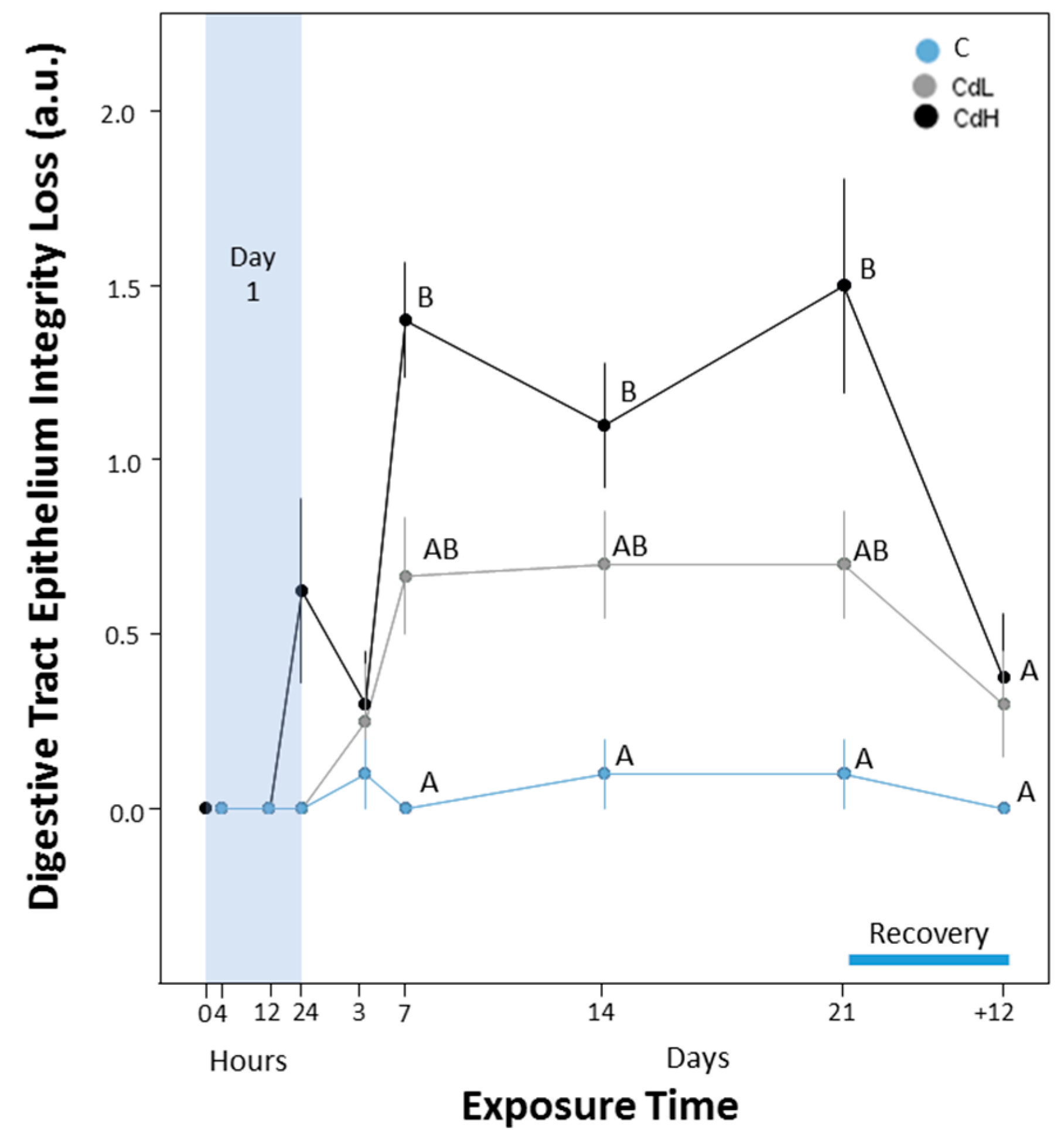
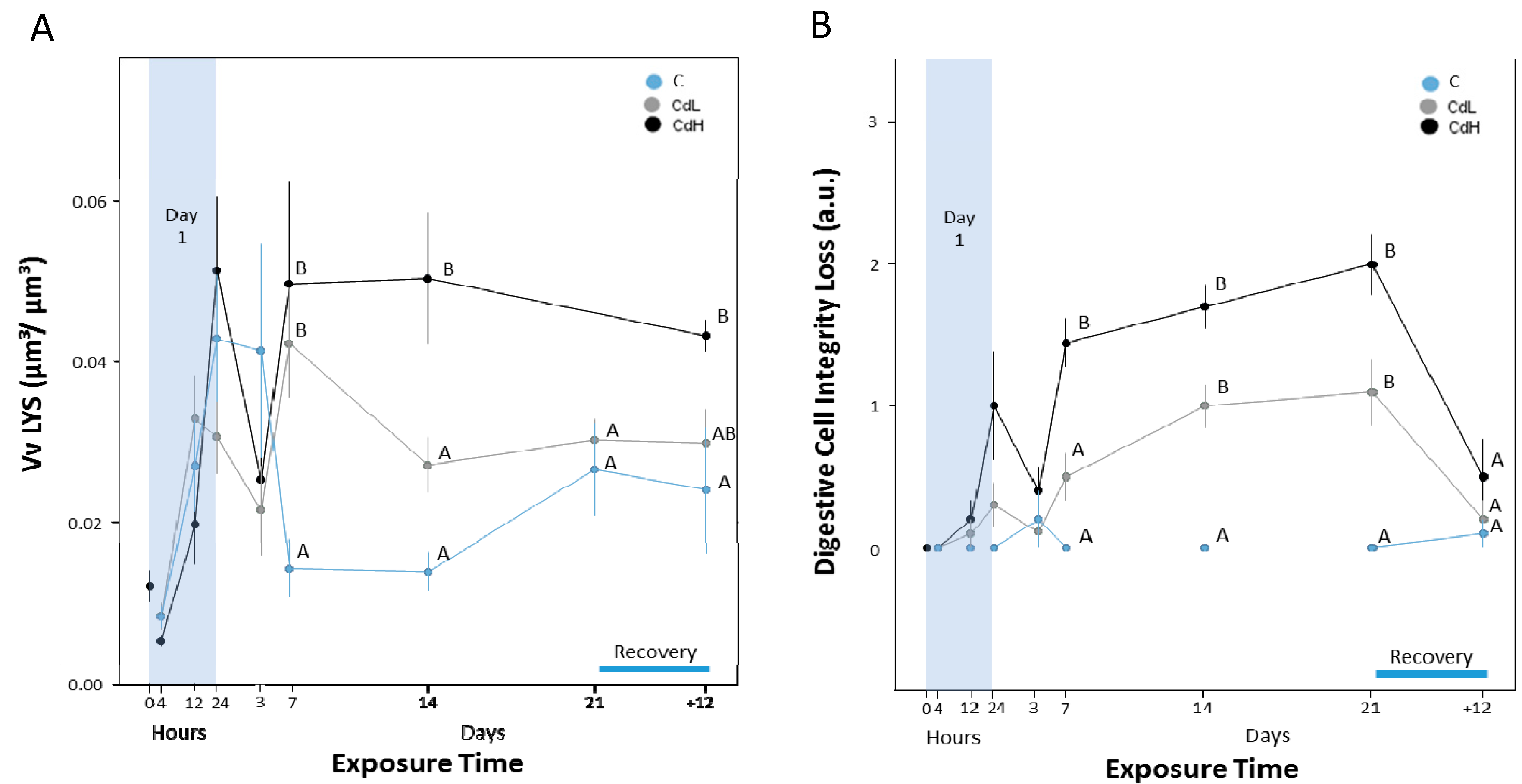
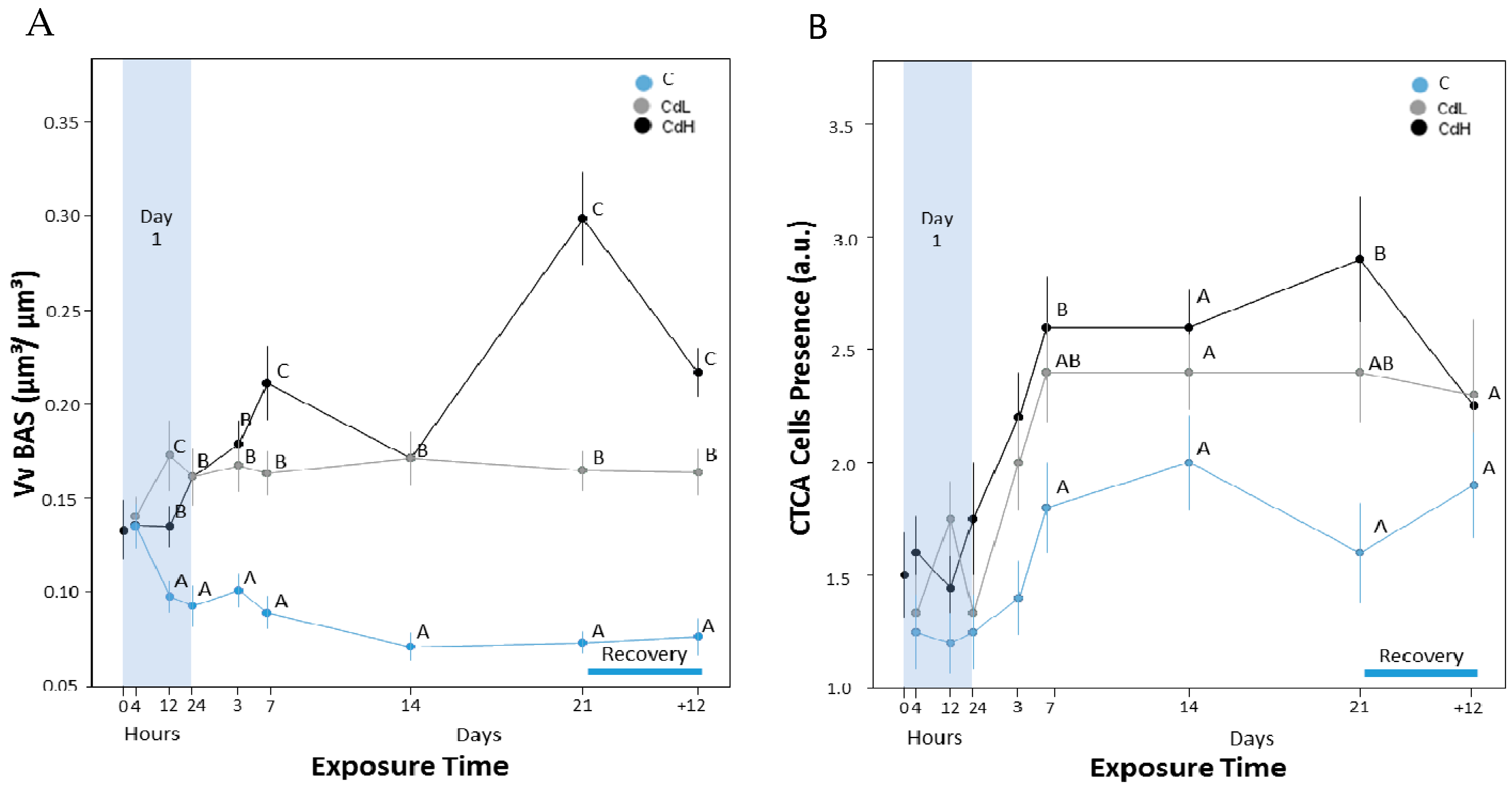
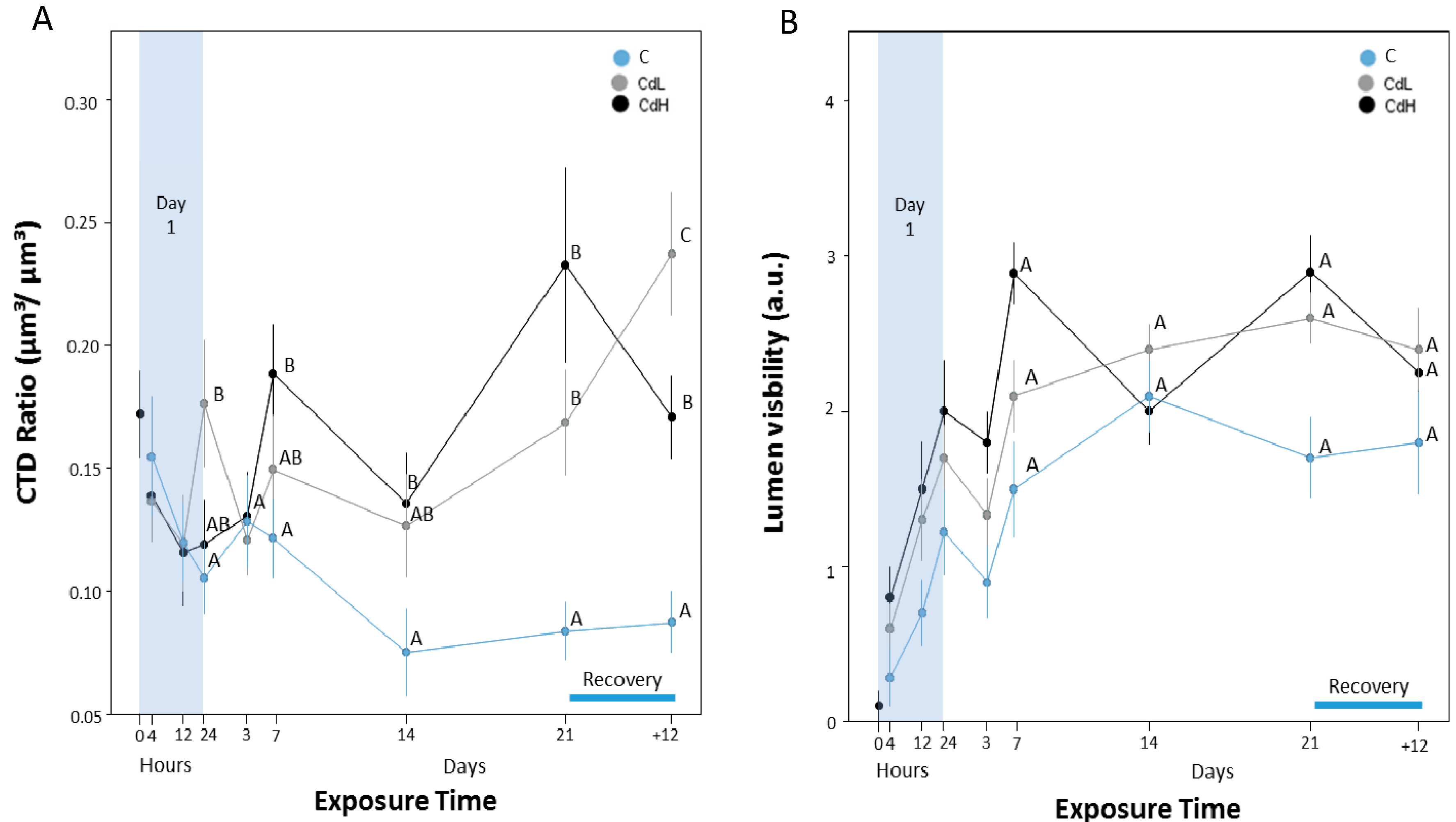
2.3. Cd-Induced Response Patterns at the Cellular and Tissue Levels
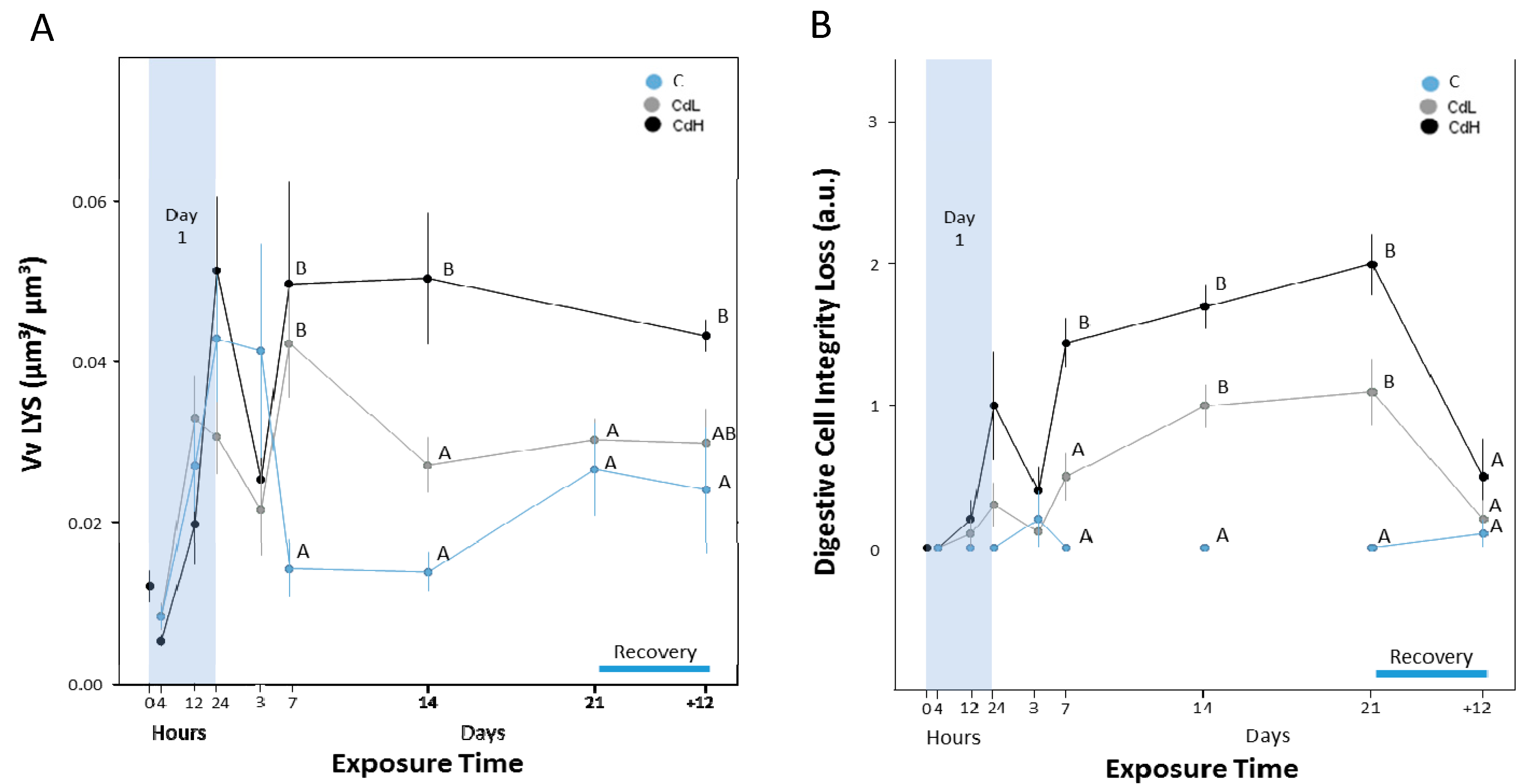
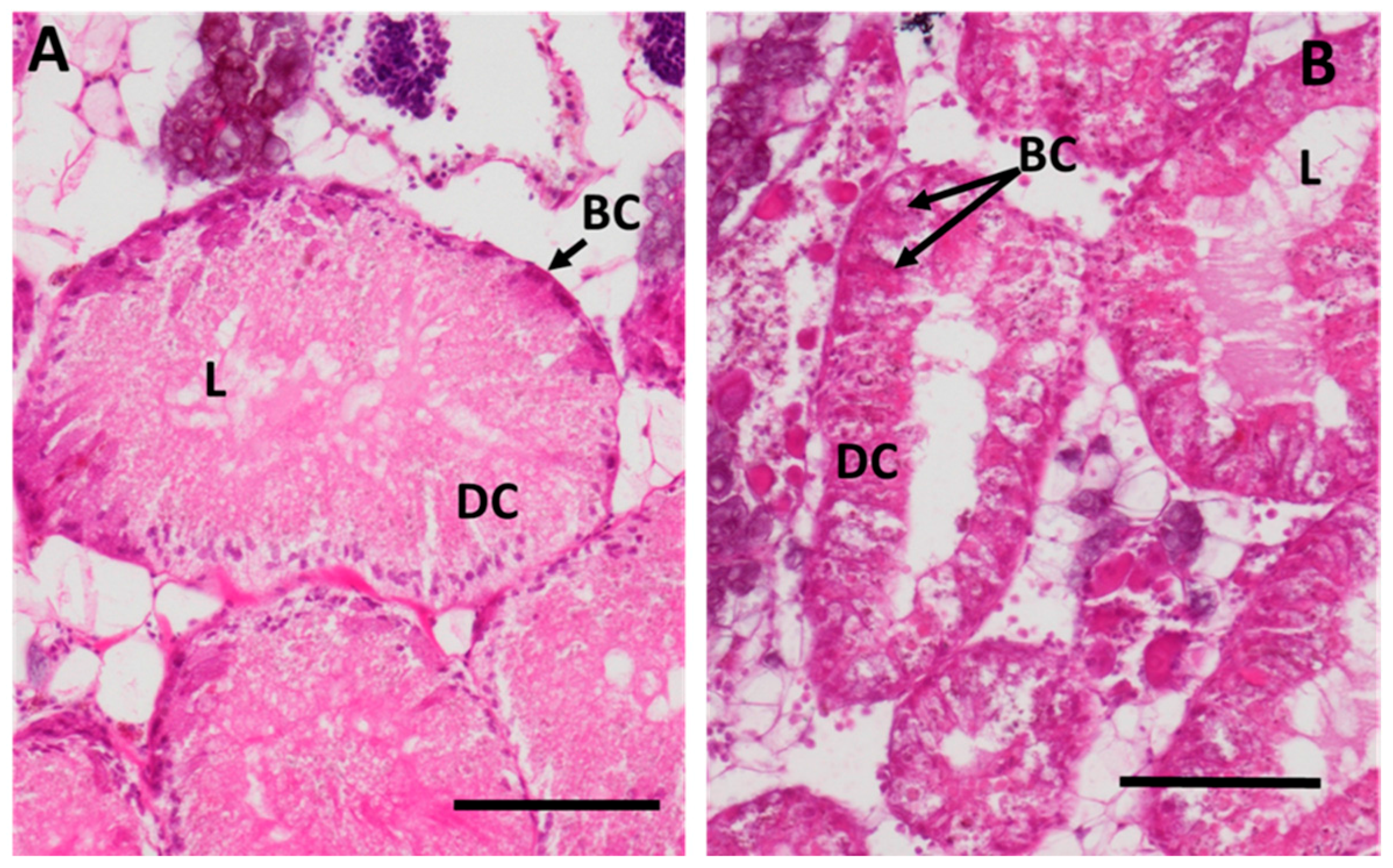
2.3.1. Digestive Cells
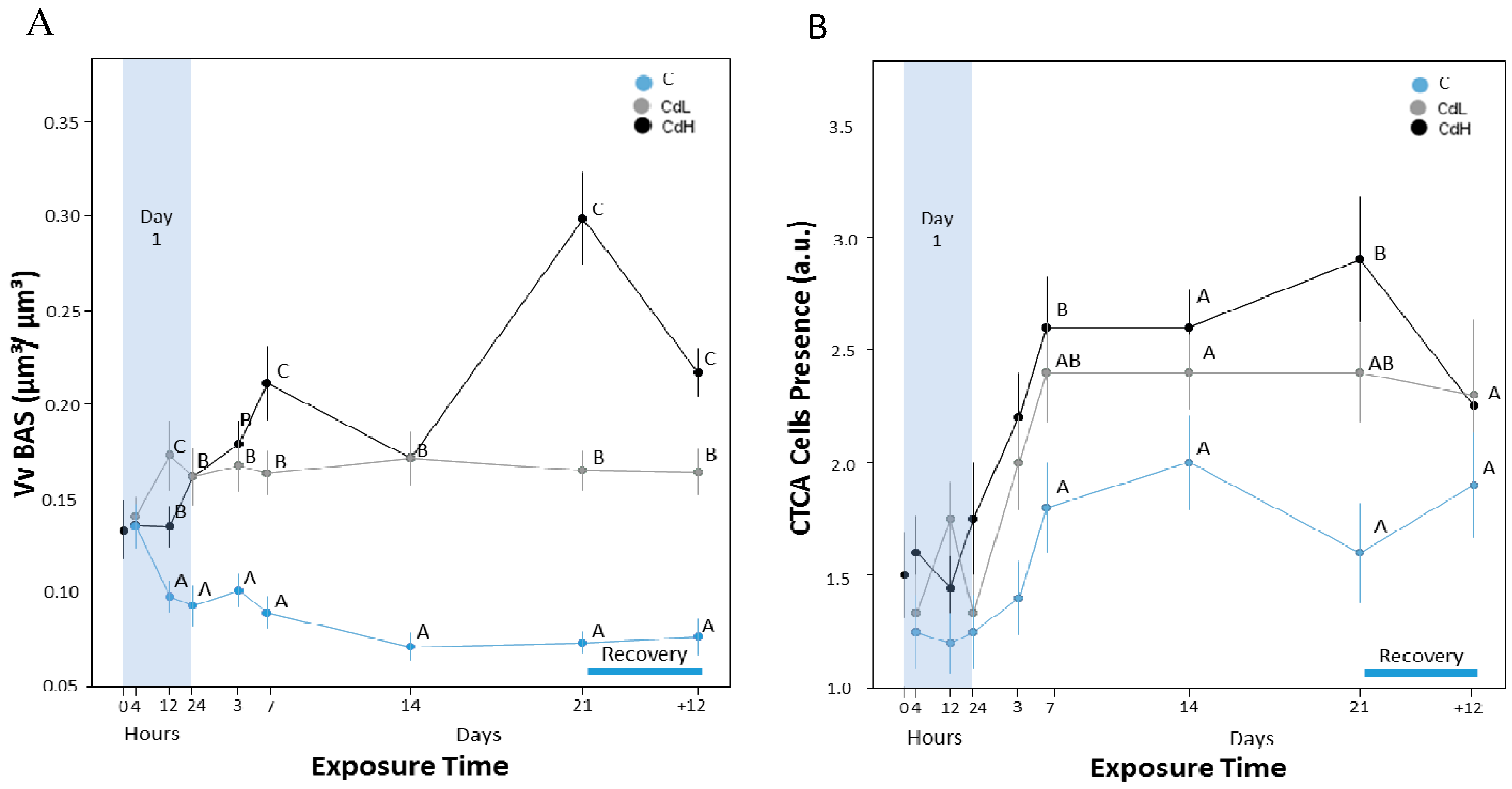
2.3.2. Basophilic Cells
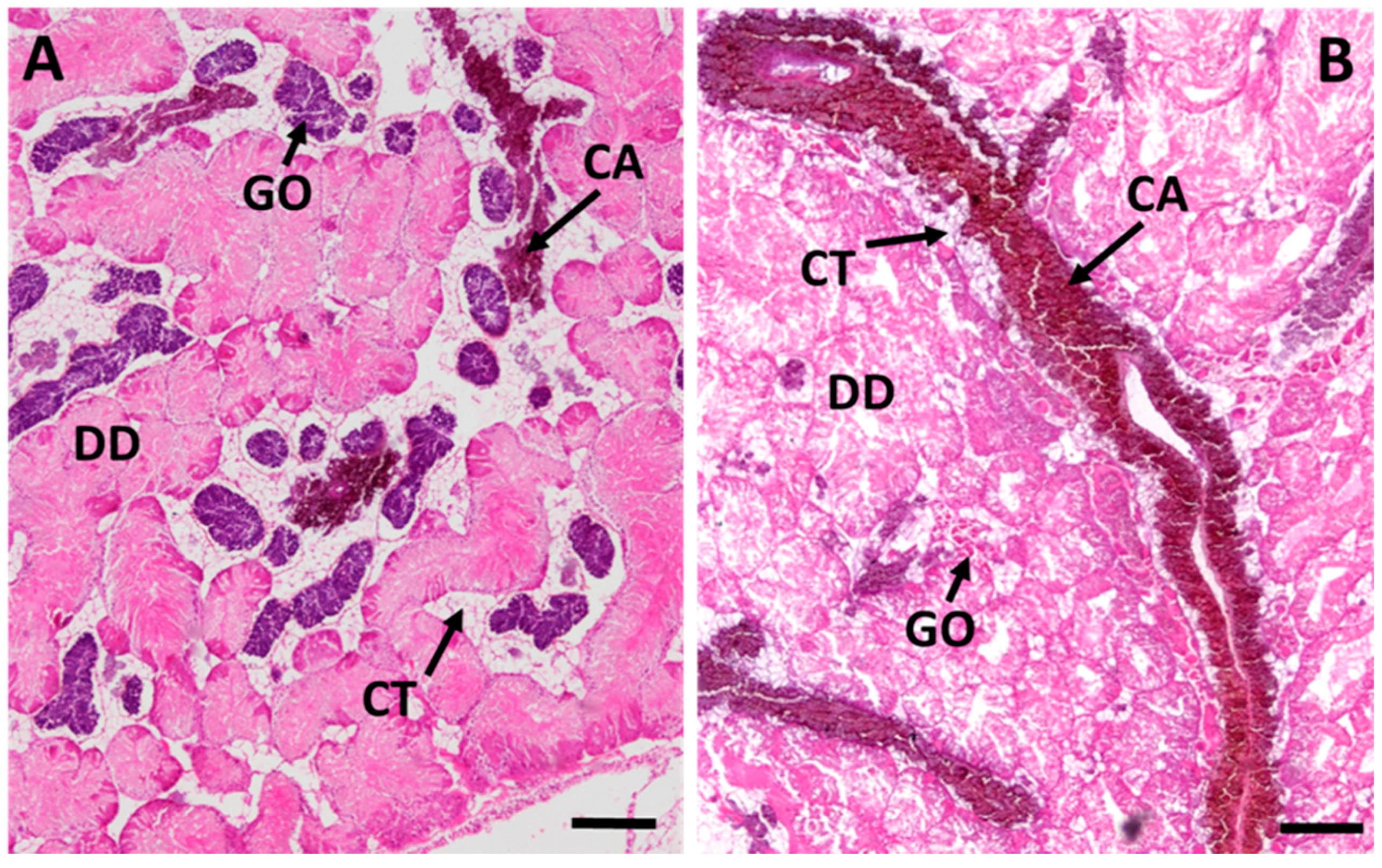
2.3.3. Connective Tissue Calcium Cells
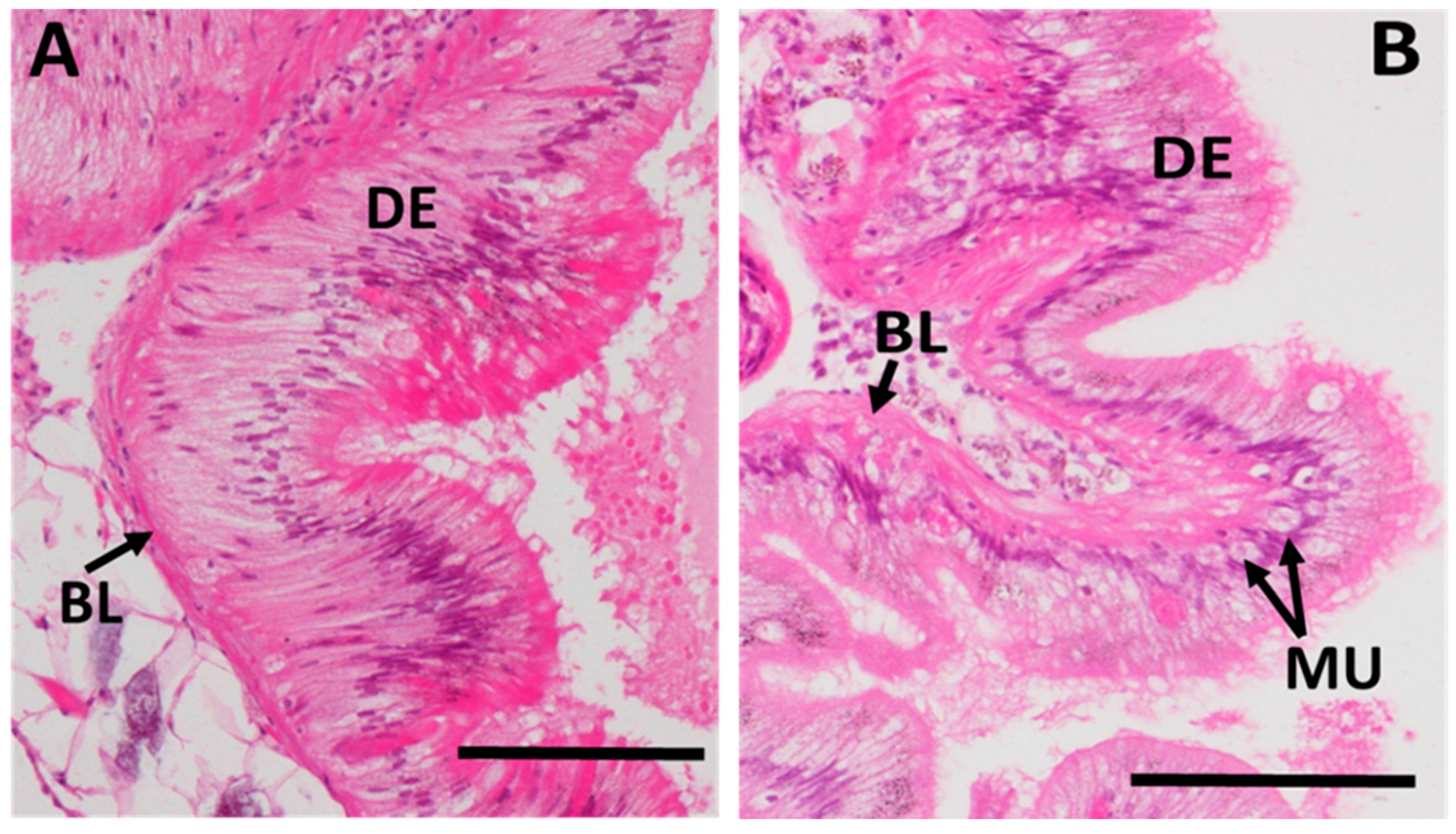
2.3.4. Midgut Gland Structure and Integrity
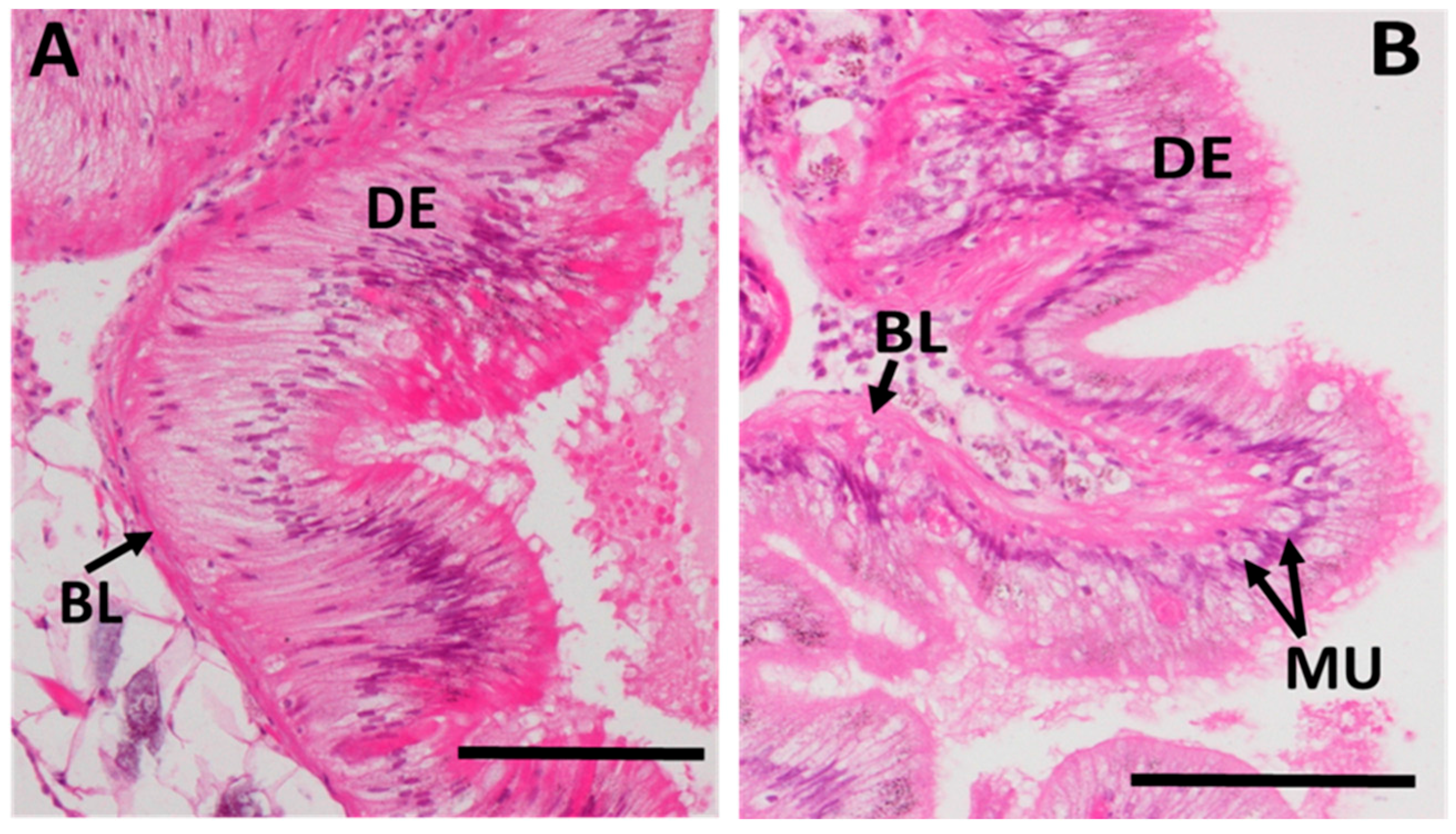
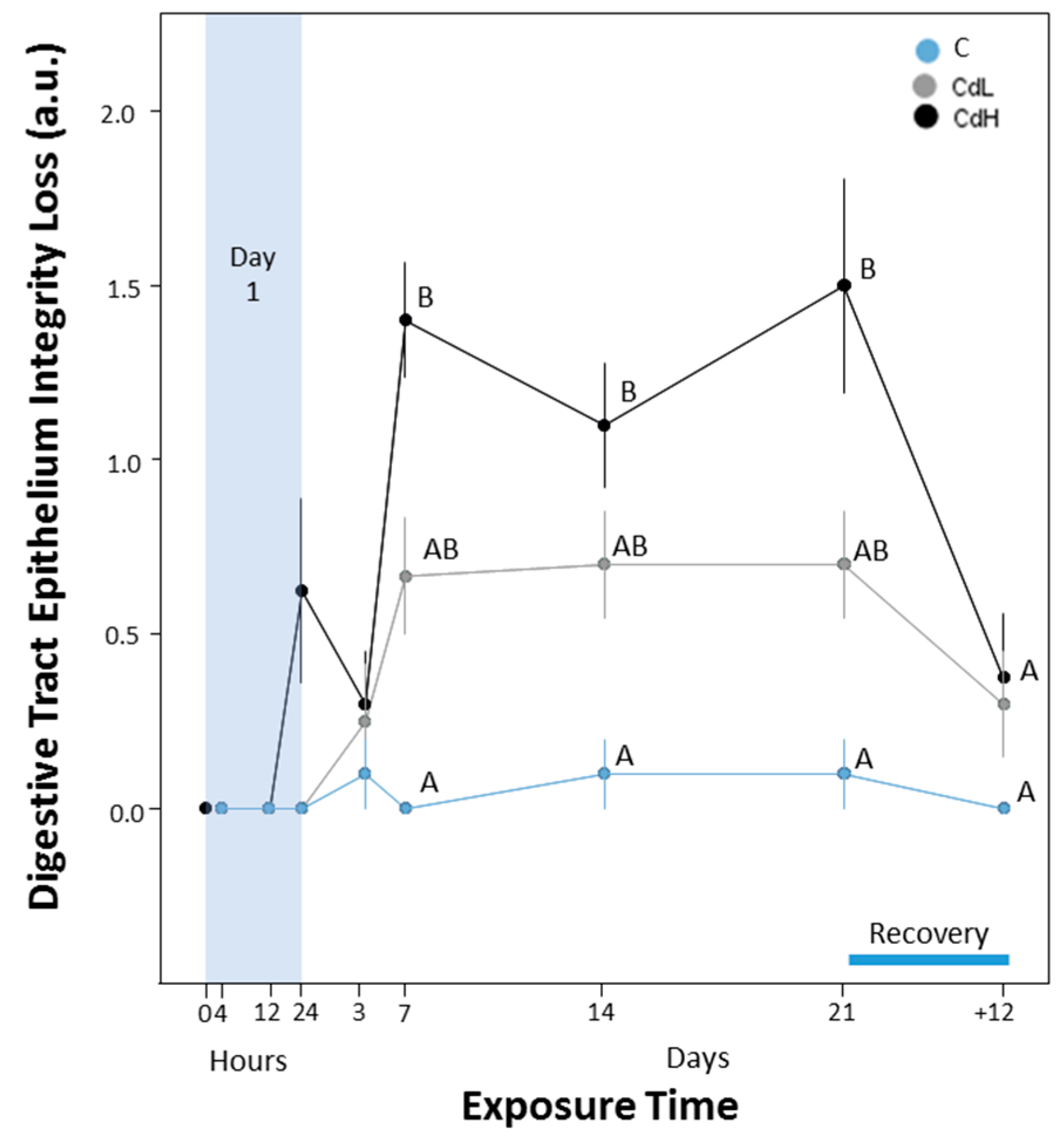
2.3.5. Integrity of Digestive Tract Epithelium
3. Discussion
3.1. Cellular Cadmium Accumulation and the Significance of MT mRNA Increase
3.2. Cadmium-Induced Cellular Response Patterns in the Midgut Gland
3.2.1. Digestive Cells
3.2.2. Basophilic Cells
3.2.3. Connective Tissue Calcium Cells
3.2.4. Cd-Induced Loss of Tissue Structure and Integrity
3.2.5. The Biomarker Potential of Assessed Parameters
4. Materials and Methods
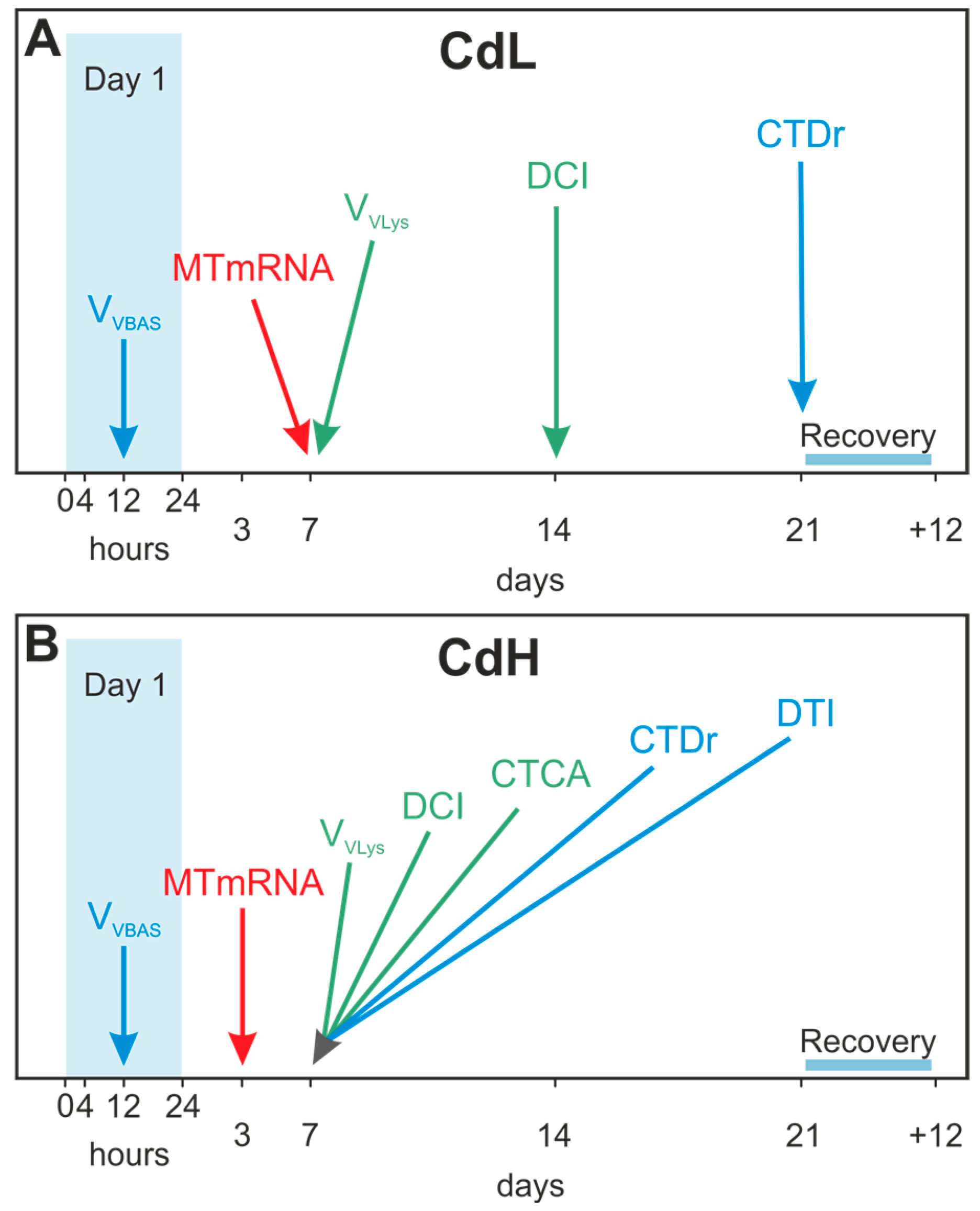
4.1. Experimental Set-Up
4.2. mRNA Isolation, Allelic Variant Screening and Quantitative Real-Time PCR of the Reference Gene
4.3. Metal Analysis
4.4. Sample Processing for Microscopy
4.5. Histopathology
4.6. Autometallography
4.7. Stereology of Digestive Cell Lysosomes
4.8 Lipofuscin Determination
4.9. Biometry
4.10. Statistics
5. Conclusions
- The exposure of periwinkles (Littorina littorea) to sub-lethal Cd concentrations (Cd Low, 0.25 and Cd High, 1 mg Cd/L) over 21 days provoked the successive induction of molecular, cellular and tissue-specific response reactions in the midgut gland and digestive tract of metal-exposed winkles. The assessed parameters were: increase of MT mRNA concentration, volume density of digestive cell lysosomes and lipofuscin formation in the midgut gland tubular epithelium, tubular digestive cell integrity, volume density of tubular basophilic cells, presence of connective tissue calcium cells, as well as tissue integrity of the midgut gland/gonad complex and of the digestive tract.
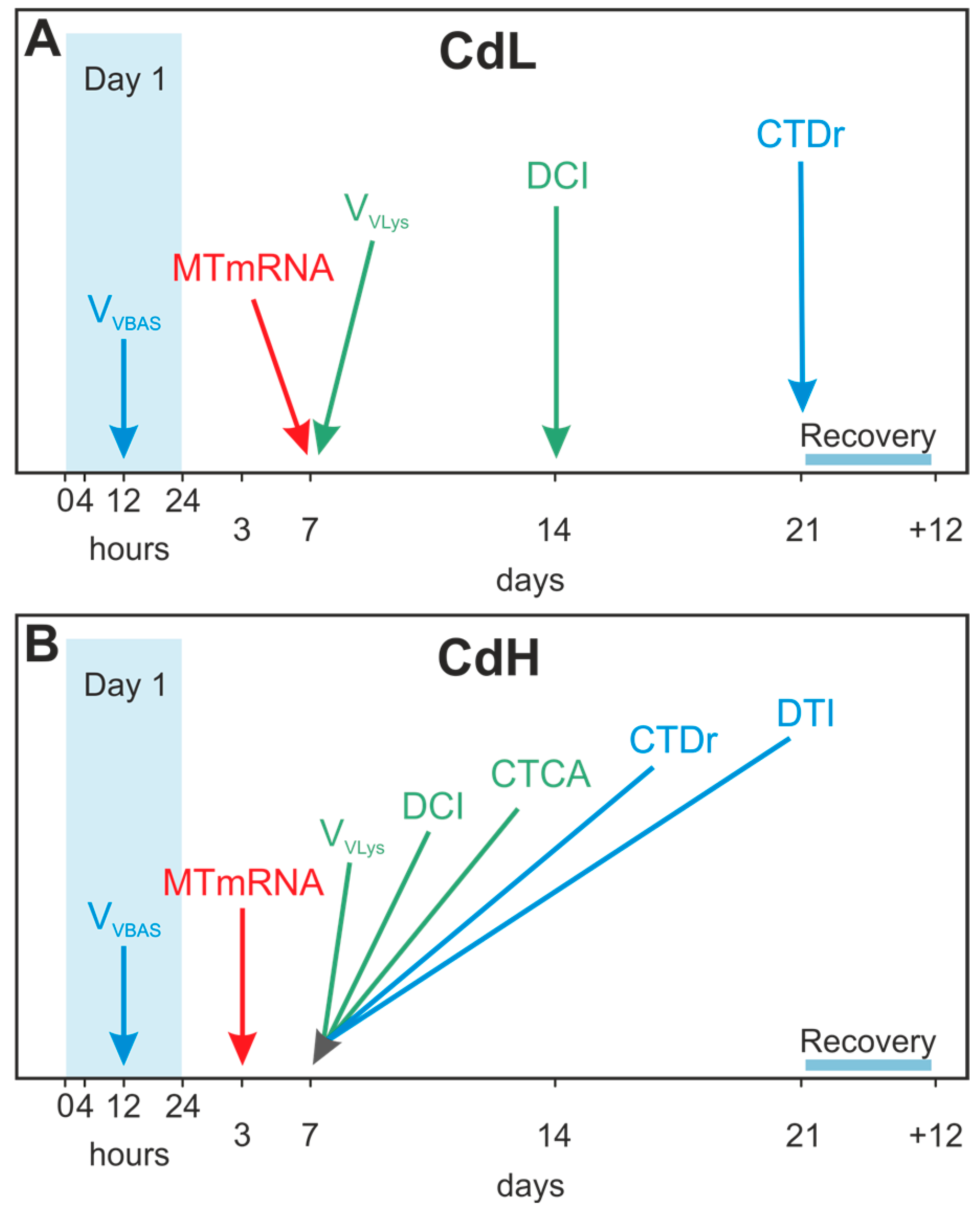
- The beginning of alterations of the assessed parameters followed distinct concentration-dependent and time-dependent patterns, where the timeframe for the onset of the different response reactions became narrower at higher Cd concentrations (CdH) compared to lower exposure concentrations (CdL).
- Interestingly, the first parameter that showed distinct reactions at both Cd concentrations (CdL and CdH) was the volume density of basophilic cells (VVBAS) in the periwinkle’s midgut gland, with significantly increased values after only 12 h of exposure. This proves that Cd-stressed periwinkles respond to the intake of the metal by a very early re-modelling of the cellular composition in the tubular tissue of the midgut gland.
- The increase of the MT mRNA concentration follows next, prior to or together with the activation of the lysosomal compartment of tubular digestive cells, suggesting that cytosolic inactivation of Cd by binding to MT and lysosomal Cd accumulation may be complementary processes of detoxification which interfere with each other. At the beginning of Cd exposure, most of the metal may first be sequestered by constitutive and de novo synthesized MT. At this stage of Cd accumulation, the MT-associated Cd pool may prevail. Due to ongoing MT turnover and degradation (Bebianno and Langston 1998), however, a certain part of liberated Cd ions would be bound to de novo synthesized MT, whereas a minor proportion of the metal (perhaps still attached to half-degraded and denatured MT) would end up for final storage in digestive cell lysosomes. Overall, the near concomitance of increasing MT mRNA transcription with enormous alterations at the cellular and tissue-specific levels reinforces the impression that the increasing MT mRNA concentrations in the midgut gland of periwinkles apparently serve the purpose of protecting Littorina littorea from potentially adverse effects induced by Cd2+ ions entering the cells or leaking out from impaired cellular structures.
- An important response strategy regarding Cd stress in Littorina littorea is, apart from Cd sequestration by MT and lysosomes, the re-modelling of midgut gland tubules by cell replacement, where digestive cells die at a faster rate than they proliferate, implicating an increase of the number and volume density of basophilic cells, which prevail in tubules of Cd-stressed individuals. In addition to this, there is also a Cd-induced increase of the number of connective tissue calcium cells.
- The specific Cd-induced molecular and cellular alterations in metal-stressed Littorina littorea may themselves be applied as biomarkers in environmental monitoring. Taken together, the time patterns of the different molecular, cellular and tissue-specific response reactions to Cd exposure in periwinkles may well provide a valuable biomarker battery approach.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hamada, T.; Tanimoto, A.; Sasaguri, Y. Apoptosis induced by cadmium. Apoptosis 1997, 2, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Gatzidou, E.; Theocharis, S. DNA repair systems as targets of cadmium toxicity. Toxicol. Appl. Pharmacol. 2006, 213, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Waisberg, M.; Joseph, P.; Hale, B.; Beyersmann, D. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology 2003, 192, 95–117. [Google Scholar] [CrossRef]
- Liu, J.; Qu, W.; Kadiiska, M.B. Role of oxidative stress in cadmium toxicity and carcinogenesis. Toxicol. Appl. Pharmacol. 2009, 238, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Koutsogiannaki, S.; Franzellitti, S.; Fabbri, E.; Kaloyianni, M. Oxidative stress parameters induced by exposure to either cadmium or 17 β-estradiol on Mytilus galloprovincialis hemocytes. The role of signaling molecules. Aquat. Toxicol. 2014, 146, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Henson, M.C.; Chedrese, P.J. Endocrine Disruption by Cadmium, a Common Environmental Toxicant with Paradoxical Effects on Reproduction. Exp. Biol. Med. 2004, 229, 383–392. [Google Scholar] [CrossRef]
- Byrne, C.; Divekar, S.D.; Storchan, G.B.; Parodi, D.A.; Martin, M.B. Cadmium—A metallohormone? Toxicol. Appl. Pharmacol. 2009, 238, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Ronald, E. Boron hazards to fish, wildlife, and invertebrates: A synoptic review. Contam. Hazard. Rev. 1990, 85, 1–20. [Google Scholar]
- Zhu, J.Y.; Huang, H.Q.; Bao, X.D.; Lin, Q.M.; Cai, Z. Acute toxicity profile of cadmium revealed by proteomics in brain tissue of Paralichthys olivaceus: Potential role of transferrin in cadmium toxicity. Aquat. Toxicol. 2006, 78, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Shadab, G.G. Cytogenetic evaluation of cadmium chloride on Channa punctatus. J. Environ. Biol. 2012, 33, 663–666. [Google Scholar] [PubMed]
- Marigomez, J.A.; Ireland, M.P. Accumulation, distribution and loss of cadmium in the marine prosobranch Littorina littorea (L.). Sci. Total Environ. 1989, 78, 1–12. [Google Scholar] [CrossRef]
- Edwards, J.W.; Edyvane, K.S.; Boxall, V.A.; Hamann, M.; Soole, K.L. Metal levels in seston and marine fish flesh near industrial and metropolitan centres in South Australia. Mar. Pollut. Bull. 2001, 42, 389–396. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Costa, A.C.S.; Korn, M.G.A. Preliminary evaluation of the cadmium concentration in seawater of the Salvador City, Brazil. Microchem. J. 2004, 78, 77–83. [Google Scholar] [CrossRef]
- Komjarova, I.; Blust, R. Comparison of liquid-liquid extraction, solid-phase extraction and co-precipitation preconcentration methods for the determination of cadmium, copper, nickel, lead and zinc in seawater. Anal. Chim. Acta 2006, 576, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Bargagli, R.; Nelli, L.; Ancora, S.; Focardi, S. Elevated cadmium accumulation in marine organisms from terra nova bay (antarctica). Polar Biol. 1996, 16, 513–520. [Google Scholar] [CrossRef]
- Dietz, R.; Riget, F.; Cleemann, M.; Aarkrog, A.; Johansen, P.; Hansen, J.C. Comparison of contaminants from different trophic levels and ecosystems. Sci. Total Environ. 2000, 245, 221–231. [Google Scholar] [CrossRef]
- Dural, M.; Göksu, M.Z.L.; Özak, A.A. Investigation of heavy metal levels in economically important fish species captured from the Tuzla lagoon. Food Chem. 2007, 102, 415–421. [Google Scholar] [CrossRef]
- Bille, L.; Binato, G.; Cappa, V.; Toson, M.; Dalla Pozza, M.; Arcangeli, G.; Ricci, A.; Angeletti, R.; Piro, R. Lead, mercury and cadmium levels in edible marine molluscs and echinoderms from the Veneto Region (north-western Adriatic Sea—Italy). Food Control. 2014, 50, 362–370. [Google Scholar] [CrossRef]
- Jitar, O.; Teodosiu, C.; Oros, A.; Plavan, G.; Nicoara, M. Bioaccumulation of heavy metals in marine organisms from the Romanian sector of the Black Sea. New Biotechnol. 2015, 32, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Bryan, G.W. Brown seaweed, fucus vesiculosus, and the gastropod, Littorina littoralis, as indicators of trace-metal availability in estuaries. Sci. Total Environ. 1983, 28, 91–104. [Google Scholar] [CrossRef]
- Soto, M. Simultaneous Quantification of Bioavailable Heavy Metals in Molluscs by Means of Cellular and Tissue Analysis. Implications for Monitoring Metal Pollution in Water Quality Assessment. Ph.D. Thesis, University of the Basque Country, Basque, Spain, 1995. [Google Scholar]
- Regoli, F.; Nigro, M.; Orlando, E. Lysosomal and antioxidant responses to metals in the Antarctic scallop Adamussium colbecki. Aquat. Toxicol. 1998, 40, 375–392. [Google Scholar] [CrossRef]
- Soto, M.; Zaldibar, B.; Cancio, I.; Taylor, M.G.; Turner, M.; Morgan, A.J.; Marigómez, I. Subcellular distribution of cadmium and its cellular ligands in mussel digestive gland cells as revealed by combined autometallography and X-ray microprobe analysis. Histochem. J. 2002, 34, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Marigomez, J.A.; Cajaraville, M.P.; Angulo, E. Cellular cadmium distribution in the common winkle, Littorina littorea (L.) determined by X-ray microprobe analysis and histochemistry. Histochemistry 1990, 94, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Nott, J.A.; Bebianno, M.J.; Langston, W.J.; Ryan, K.P. Cadmium in the gastropod Littorina littorea. J. Mar. Biol. Assoc. U. K. 1993, 73, 655–665. [Google Scholar] [CrossRef]
- Klekowski, R. The influence of low salinity and desiccation on the survival, osmoregulation and water balance of Littorina littorea (L.) (Prosobranchia). Pol. Arch. Hydrobiol. 1963, 11, 241–250. [Google Scholar]
- Churchill, T.A.; Storey, K.B. Metabolic responses to freezing and anoxia by the periwinkle Littorina littorea. J. Therm. Biol. 1996, 21, 57–63. [Google Scholar] [CrossRef]
- Russell, E.L.; Storey, K.B. Anoxia and freezing exposures stimulate covalent modification of enzymes of carbohydrate metabolism in Littorina littorea. J. Comp. Physiol. B 1995, 165, 132–142. [Google Scholar] [CrossRef]
- Storey, K.B.; Lant, B.; Anozie, O.O.; Storey, J.M. Metabolic mechanisms for anoxia tolerance and freezing survival in the intertidal gastropod, Littorina littorea. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 165, 448–459. [Google Scholar] [CrossRef] [PubMed]
- English, T.E.; Storey, K.B. Freezing and anoxia stresses induce expression of metallothionein in the foot muscle and hepatopancreas of the marine gastropod Littorina littorea. J. Exp. Biol. 2003, 206, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- Zaldibar, B.; Cancio, I.; Marigomez, I. Reversible alterations in epithelial cell turnover in digestive gland of winkles (Littorina littorea) exposed to cadmium and their implications for biomarker measurements. Aquat. Toxicol. 2007, 81, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Marigomez, I. Aportaciones Cito-Histologicas a la Evaluación Ecotoxicologica de Niveles Subletales de Cadmio en el Medio Marino: Estudios de Laboratorio en el Gasterópodo Prosobranquio Littorina littorea (L.). Ph.D. Thesis, University of the Basque Country, Basque, Spain, 1989. [Google Scholar]
- Marigomez, I.; Cajaraville, M.P.; Angulo, E. Histopathology of the digestive gland-gonad complex of the marine prosobranch Littorina littorea exposed to cadmium. Dis. Aquat. Organ. 1990, 9, 229–238. [Google Scholar] [CrossRef]
- Marigomez, I.; Soto, M.; Cajaraville, M.P.; Angulo, E.; Giamberini, L. Cellular and subcellular distribution of metals in molluscs. Microsc. Res. Tech. 2002, 56, 358–392. [Google Scholar] [CrossRef] [PubMed]
- Bebianno, M. Cadmium and metallothionein turnover in different tissues of the gastropod Littorina littorea. Talanta 1998, 46, 301–313. [Google Scholar] [CrossRef]
- Baumann, C.; Beil, A.; Jurt, S.; Niederwanger, M.; Palacios, O.; Capdevila, M.; Atrian, S.; Dallinger, R.; Zerbe, O. Structural Adaptation of a Protein to Increased Metal Stress: NMR Structure of a Marine Snail Metallothionein with an Additional Domain. Angew. Chem. Int. Ed. Engl. 2017, 56, 4617–4622. [Google Scholar] [CrossRef] [PubMed]
- Palacios, Ò.; Jiménez-Marti, E.; Niederwanger, M.; Gil-Moreno, S.; Zerbe, O.; Atrian, S.; Dallinger, R.; Capdevila, M. Analysis of metal-binding features of the wild type and two domain-truncated mutant variants of Littorina littorea metallothionein reveals its Cd-specific character. Int. J. Mol. Sci. 2017, 18, 1452. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.M.; Marigomez, J.A.; Angulo, E. Quantitative alterations in the structure of the digestive cell of Littorina littorea on exposure to cadmium. Mar. Biol. 1989, 103, 547–553. [Google Scholar] [CrossRef]
- Marigómez, I.; Izagirre, U.; Lekube, X. Lysosomal enlargement in digestive cells of mussels exposed to cadmium, benzo[a]pyrene and their combination. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 141, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. The distribution of some hydrolytic enzymes in the cells of the digestive gland of certain lamellibranchs and gastropods. J. Zool. 1969, 158, 277–291. [Google Scholar] [CrossRef]
- Robinson, W.E. Assessment of bivalve intracellular digestion based on direct measurements. J. Moll. Stud. 1983, 49, 1–8. [Google Scholar] [CrossRef]
- Moore, M.N.; Pipe, R.K.; Farrar, S.V. Lysosomal and microsomal responses to environmental factors in Littorina littorea from Sullom Voe. Mar. Pollut. Bull. 1982, 13, 340–345. [Google Scholar] [CrossRef]
- Cajaraville, M.P.; Marigómez, J.A.; Angulo, E. Short-term toxic effects of 1-naphthol on the digestive gland-gonad complex of the marine prosobranch Littorina littorea (L): A light microscopic study. Arch. Environ. Contam. Toxicol. 1990, 19, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Marigomez, I.; Soto, M.; Etxeberria, M.; Angulo, E. Effects of size, sex, reproduction, and trematode infestation on the quantitative structure of digestive tubules in stressed winkles. In Zoologische Jahrbücher. Abteilung für Anatomie und Ontogenie der Tiere; Fischer, G., Ed.; Fischer: Jena, Germany, 1993; Volume 123, pp. 319–336. [Google Scholar]
- Marigomez, I.; Cajaraville, M.P.; Soto, M.; Lekube, X. Cell-type replacement, a successful strategy of molluscs to adapt to chronic exposure to pollutants. Cuad. Investig. Biol. 1998, 20, 411–414. [Google Scholar]
- Gil, J.M.; Marigomez, J.A.; Angulo, E. Histophysiology of polysaccharide and lipid reserves in various tissues of Littorina littorea exposed to sublethal concentrations of cadmium. Comp. Biochem. Physiol. Part C Comp. 1989, 94, 641–648. [Google Scholar] [CrossRef]
- Cajaraville, M.P.; Bebianno, M.J.; Blasco, J.; Porte, C.; Sarasquete, C.; Viarengo, A. The use of biomarkers to assess the impact of pollution in coastal environments of the Iberian Peninsula: A practical approach. Sci. Total Environ. 2000, 247, 295–311. [Google Scholar] [CrossRef]
- Kwamla Atupra, D. Determination of the Toxicity of Cadmium in Three Populations of Littorina littorea Taken from a Pollution and Salinity Gradient in the Scheldt Estuary (Mollusca: Gastropoda). Master’s Thesis, Vrije Universiteit Brussel, Brussel, Belgium, 2001. [Google Scholar]
- De Wolf, H.; Backeljau, T.; Blust, R. Sensitivity to cadmium along a salinity gradient in populations of the periwinkle, Littorina littorea, using time-to-death analysis. Aquat. Toxicol. 2004, 66, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Danscher, G. Autometallography. A new technique for light and electron microscopic visualization of metals in biological tissues (gold, silver, metal sulphides and metal selenides). Histochemistry 1984, 81, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.; Cajaraville, M.P.; Angulo, E.; Marigómez, I. Autometallographic localization of protein-bound copper and zinc in the common winkle, Littorina littorea: A light microscopical study. Histochem. J. 1996, 28, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Kägi, J.H.R. Evolution, structure and chemical activity of class I metallothionein: An overview. In Metallothionein; Suzuki, K.T., Imura, N., Kimura, M., Eds.; Birkhauser Verlag: Basel, Switzerland, 1993; Volume III, pp. 29–55. [Google Scholar]
- Dallinger, R.; Wang, Y.; Berger, B.; Mackay, E.A.; Kägi, J.H.R. Spectroscopic characterization of metallothionein from the terrestrial snail, Helix pomatia. Eur. J. Biochem. 2001, 268, 4126–4133. [Google Scholar] [CrossRef] [PubMed]
- Bebianno, M.J.; Langston, W.J.; Simkiss, K. Metallothionein induction in Littorina-littorea (Mollusca, Prosobranchia) on exposure to cadmium. J. Mar. Biol. Assoc. U. K. 1992, 72, 329–342. [Google Scholar] [CrossRef]
- Carginale, V.; Scudiero, R.; Capasso, C.; Capasso, A.; Kille, P.; di Prisco, G.; Parisi, E. Cadmium-induced differential accumulation of metallothionein isoforms in the Antarctic icefish, which exhibits no basal metallothionein protein but high endogenous mRNA levels. Biochem. J. 1998, 332, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Tom, M.; Chen, N.; Segev, M.; Herut, B.; Rinkevich, B. Quantifying fish metallothionein transcript by real time PCR for its utilization as an environmental biomarker. Mar. Pollut. Bull. 2004, 48, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Dondero, F.; Piacentini, L.; Banni, M.; Rebelo, M.; Burlando, B.; Viarengo, A. Quantitative PCR analysis of two molluscan metallothionein genes unveils differential expression and regulation. Gene 2005, 345, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.M.Y.; Taylor, A.C.; Furness, R.W. Temperature-dependent physiological responses of the dogwhelk Nucella lapillus to cadmium exposure. J. Mar. Biol. Assoc. U. K. 2000, 80, S0025315400002472. [Google Scholar] [CrossRef]
- Babczynska, A.; Wilczek, G.; Wilczek, P.; Szulinska, E.; Witas, I. Metallothioneins and energy budget indices in cadmium and copper exposed spiders Agelena labyrinthica in relation to their developmental stage, gender and origin. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Liu, J.; Choudhuri, S. METALLOTHIONEIN: An Intracellular Protein to Protect Against Cadmium Toxicity. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 267–294. [Google Scholar] [CrossRef] [PubMed]
- Pipe, R.K.; Moore, M.N. An ultrastructural study on the effects of phenanthrene on lysosomal membranes and distribution of the lysosomal enzyme β-glucuronidase in digestive cells of the periwinkle Littorina littorea. Aquat. Toxicol. 1986, 8, 65–76. [Google Scholar] [CrossRef]
- Cajaraville, M.P.; Marigomez, J.A.; Angulo, E. A stereological survey of lysosomal structure alterations in Littorina littorea exposed to 1-naphthol. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1989, 93, 231–237. [Google Scholar] [CrossRef]
- Lowe, D.M.; Moore, M.N.; Readman, J.W. Pathological reactions and recovery of hepatopancreatic digestive cells from the marine snail Littorina littorea following exposure to a polycyclic aromatic hydrocarbon. Mar. Environ. Res. 2006, 61, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Ugarteburu, G.; Saez, V.; McQuaid, C.D.; Angulo, E. Validation of a planimetric procedure to quantify stress in Littorina littorea (Gastropoda: Mollusca): Is it independent of the reproductive cycle? In Advances in Littorinid Biology, Proceedings of the Fourth International Symposium on Littorinid Biology, Roscoff, France, 19–25 September 1993; Mill, P.J., McQuaid, C.D., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1995; pp. 37–44. [Google Scholar]
- Marigómez, I.; Soto, M.; Angulo, E. Seasonal variability in the quantitative structure of the digestive tubules of Littorina littorea. Aquat. Living Resour. 1992, 5, 299–305. [Google Scholar] [CrossRef]
- Beyersmann, D.; Hechtenberg, S. Cadmium, gene regulation, and cellular signalling in mammalian cells. Toxicol. Appl. Pharmacol. 1997, 144, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Zaldibar, B.; Cancio, I.; Soto, M.; Marigomez, I. Digestive cell turnover in digestive gland epithelium of slugs experimentally exposed to a mixture of cadmium and kerosene. Chemosphere 2007, 70, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Chabicovsky, M.; Klepal, W.; Dallinger, R. Mechanisms of cadmium toxicity in terrestrial pulmonates: Programmed cell death and metallothionein overload. Environ. Toxicol. Chem. 2004, 23, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Triebskorn, R.; Köhler, H.R. The impact of heavy metals on the grey garden slug, Deroceras reticulatum (Muller): Metal storage, cellular effects and semi-quantitative evaluation of metal toxicity. Environ. Pollut. 1996, 93, 327–343. [Google Scholar] [CrossRef]
- Hödl, E.; Felder, E.; Chabicovsky, M.; Dallinger, R. Cadmium stress stimulates tissue turnover in Helix pomatia: Increasing cell proliferation from metal tolerance to exhaustion in molluscan midgut gland. Cell. Tissue Res. 2010, 341, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Marigómez, J.A.; Sáez, V.; Cajaraville, M.P.; Angulo, E. A planimetric study of the mean epithelial thickness (MET) of the molluscan digestive gland over the tidal cycle and under environmental stress conditions. Helgoländer Meeresuntersuchungen 1990, 44, 81–94. [Google Scholar] [CrossRef]
- Cajaraville, M.P.; Robledo, Y.; Etxeberria, M.; Marigomez, I. Cellular biomarkers as useful tools in the biological monitoring of pollution: Molluscan digestive lysosomes. In Cell Biology in Environmental Toxicology; University of the Basque Country Press Service: Bilbao, Spain, 1995; pp. 29–55. [Google Scholar]
- Lowe, D.M.; Clarke, K.R. Contaminant-induced changes in the structure of the digestive epithelium of Mytilus edulis. Aquat. Toxicol. 1989, 15, 345–358. [Google Scholar] [CrossRef]
- Marigómez, I.; Soto, M.; Kortabitarte, M. Tissue-level biomarkers and biological effect of mercury on sentinel slugs Arion ater. Arch. Environ. Contam. Toxicol. 1996, 31, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.Z.; Nott, J.A. The role of intracellular biomineralized granules in the regulation and detoxification of metals in gastropods with special reference to the marine prosobranch Littorina littorea. Aquat. Toxicol. 1981, 1, 239–256. [Google Scholar] [CrossRef]
- Chmielowska-Bak, J.; Izbiańska, K.; Deckert, J. The toxic Doppelganger: On the ionic and molecular mimicry of cadmium. Acta Biochim. Pol. 2013, 60, 369–374. [Google Scholar] [PubMed]
- Brooks, S.; Harman, C.; Soto, M.; Cancio, I.; Glette, T.; Marigomez, I. Integrated coastal monitoring of a gas processing plant using native and caged mussels. Sci. Total Environ. 2012, 426, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Rementeria, A.; Mikolaczyk, M.; Lanceleur, L.; Blanc, G.; Soto, M.; Schäfer, J.; Zaldibar, B. Assessment of the effects of Cu and Ag in oysters Crassostrea gigas (Thunberg, 1793) using a battery of cell and tissue level biomarkers. Mar. Environ. Res. 2016, 122, 11–22. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.F.; Shugart, L.R. Biological markers of environmental contamination. In Biomarkers of Environmental Contamination; McCarthy, J.F., Shugart, L.R., Eds.; Lewis Publishers: Chelsea, MI, USA, 1990; Volume 3, pp. 3–14. [Google Scholar]
- Izagirre, U.; Ruiz, P.; Marigomez, I. Time-course study of the early lysosomal responses to pollutants in mussel digestive cells using acid phosphatase as lysosomal marker enzyme. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R. Stereological Methods. Vol 1: Practical Methods for Biological Morphometr; Academic Press: New York, NY, USA, 1979; p. 415. [Google Scholar]
- Moore, M.N. Cytochemical demonstration of latency of lysosomal hydrolases in digestive cells of the common mussel, mytilus edulis, and changes induced by thermal stress. Cell. Tissue Res. 1976, 175, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Lowe, D.M.; Moore, M.N.; Clarke, K.R. Effects of oil on digestive cells in mussels: Quantitative alterations in cellular and lysosomal structure. Aquat. Toxicol. 1981, 1, 213–226. [Google Scholar] [CrossRef]
- Etxeberria, M.; Sastre, I.; Cajaraville, M.P.; Marigomez, I. Digestive lysosome enlargement induced by experimental exposure to metals (Cu, Cd, and Zn) in mussels collected from a zinc-polluted site. Arch. Environ. Contam. Toxicol. 1994, 27, 338–345. [Google Scholar] [CrossRef]
- Pearse, A.G.E. Pigments and pigment precursors. In Histochemistry. Theoretical and Applied. Vol. 2: Analytical Technology AGE Pearse, 4th ed.; Churchill Livingstone: Edinburgh, UK, 1985; pp. 874–928. [Google Scholar]
- Regoli, F. Lysosomal responses as a sensitive stress index in biomonitoring heavy metal pollution. Mar. Ecol. Prog. Ser. 1992, 84, 63–69. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endpoint | Score | ||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| Digestive cell integrity loss | Control integrity | Slightly lower cells | Lower cells slightly vacuolated epithelium | Low cells Vacuolated epithelium | Disintegrated tissue |
| Integrity loss of digestive tract epithelium | Control integrity | Slightly lower cells | Slightly lower cells slightly vacuolated epithelium | Slightly lower cells vacuolated epithelium | Disintegrated tissue |
| Lumen visibility of midgut gland tubules | <10% of the area of the tubule | <30% of the area of the tubule | <40% of the area of the tubule | <50% of the area of the tubule | >50% of the area of the tubule |
| Presence of Ca cells in the midgut gland connective tissue | <30% of the connective tissue | <45% of the connective tissue | <60% of the connective tissue | <85% of the connective tissue | >85% of the connective tissue |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito, D.; Niederwanger, M.; Izagirre, U.; Dallinger, R.; Soto, M. Successive Onset of Molecular, Cellular and Tissue-Specific Responses in Midgut Gland of Littorina littorea Exposed to Sub-Lethal Cadmium Concentrations. Int. J. Mol. Sci. 2017, 18, 1815. https://doi.org/10.3390/ijms18081815
Benito D, Niederwanger M, Izagirre U, Dallinger R, Soto M. Successive Onset of Molecular, Cellular and Tissue-Specific Responses in Midgut Gland of Littorina littorea Exposed to Sub-Lethal Cadmium Concentrations. International Journal of Molecular Sciences. 2017; 18(8):1815. https://doi.org/10.3390/ijms18081815
Chicago/Turabian StyleBenito, Denis, Michael Niederwanger, Urtzi Izagirre, Reinhard Dallinger, and Manu Soto. 2017. "Successive Onset of Molecular, Cellular and Tissue-Specific Responses in Midgut Gland of Littorina littorea Exposed to Sub-Lethal Cadmium Concentrations" International Journal of Molecular Sciences 18, no. 8: 1815. https://doi.org/10.3390/ijms18081815
APA StyleBenito, D., Niederwanger, M., Izagirre, U., Dallinger, R., & Soto, M. (2017). Successive Onset of Molecular, Cellular and Tissue-Specific Responses in Midgut Gland of Littorina littorea Exposed to Sub-Lethal Cadmium Concentrations. International Journal of Molecular Sciences, 18(8), 1815. https://doi.org/10.3390/ijms18081815