Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
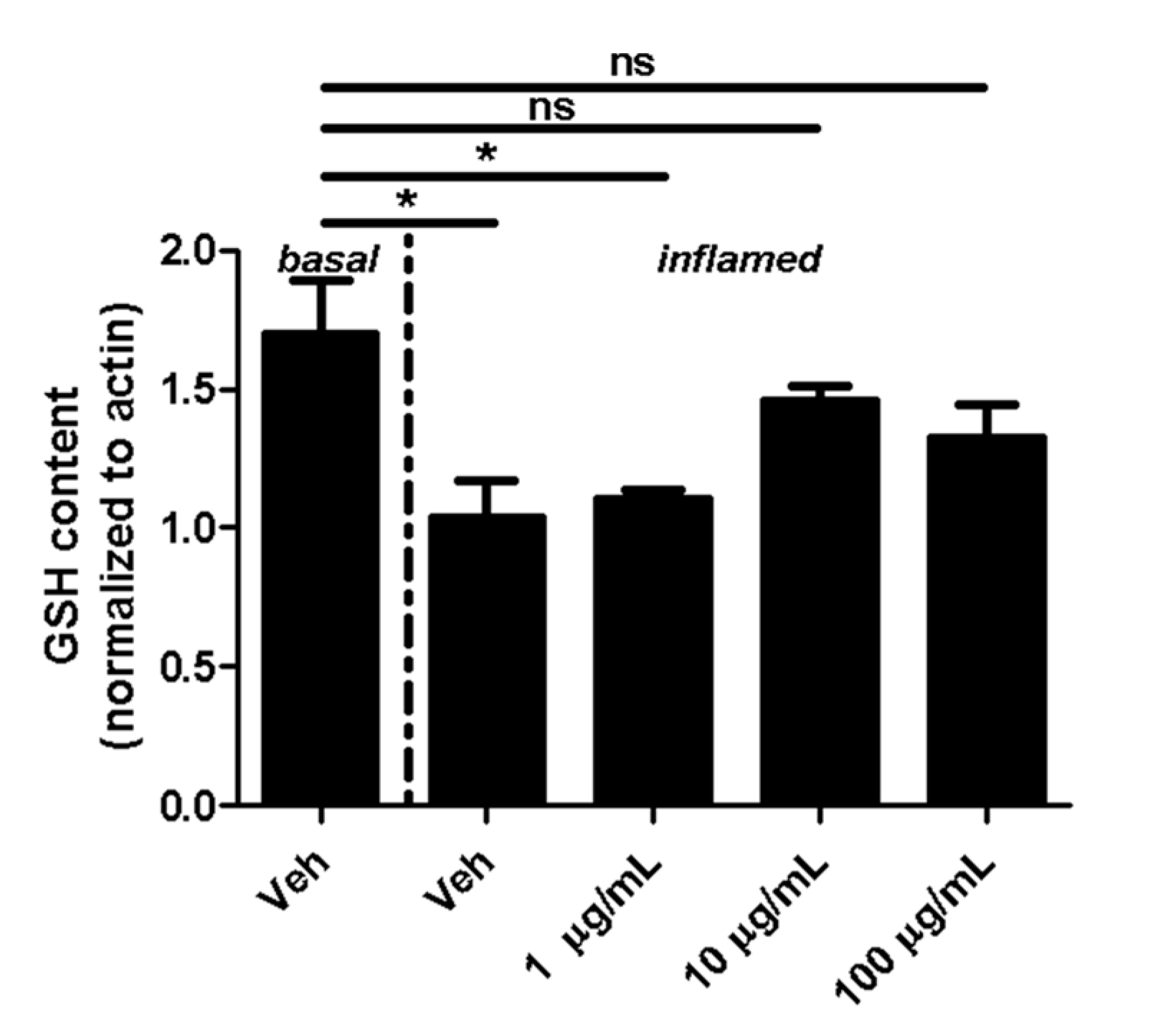
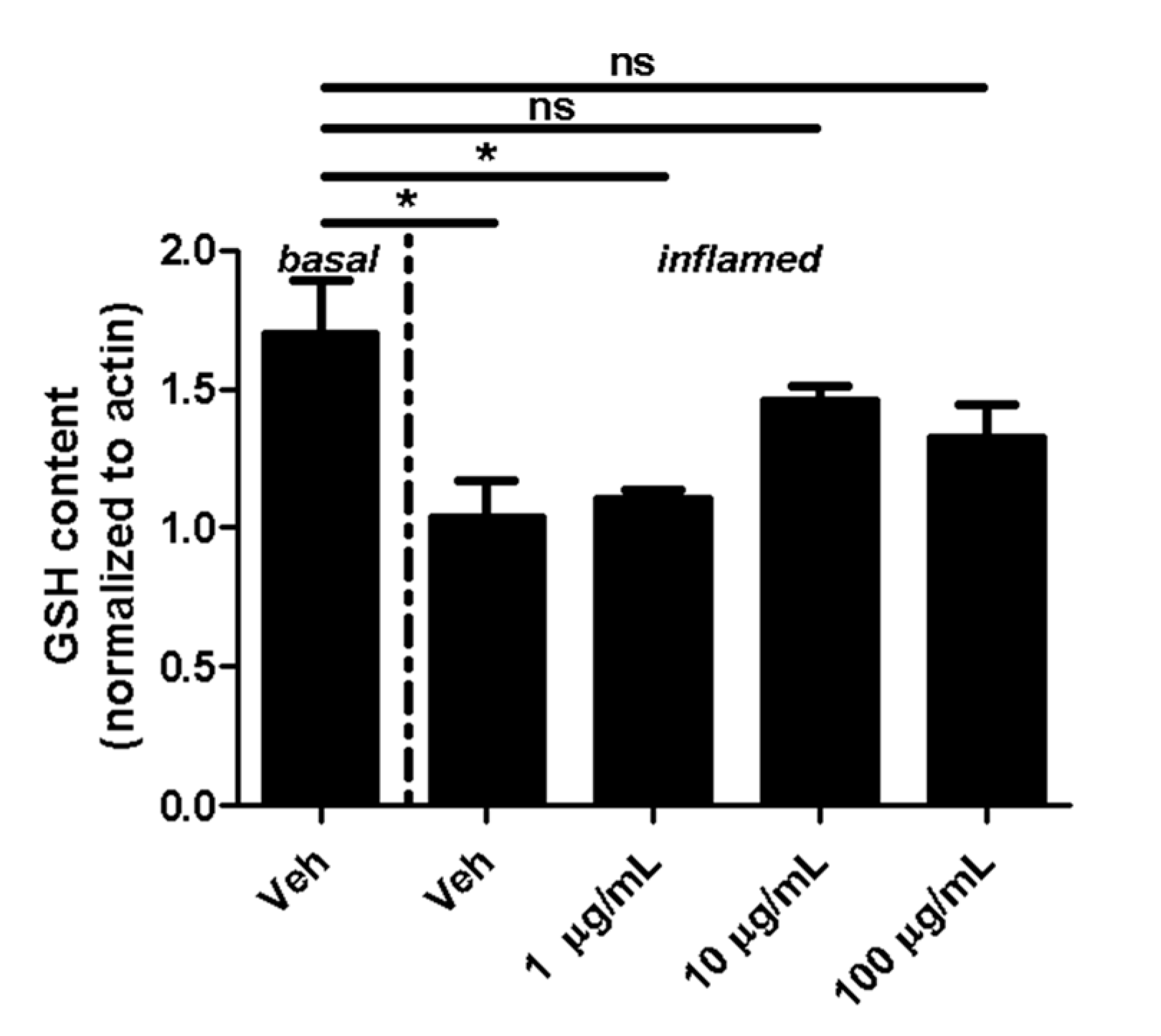
2.1. Fullerenols Modulate Glutathione (GSH) Content in Inflamed MBMEC
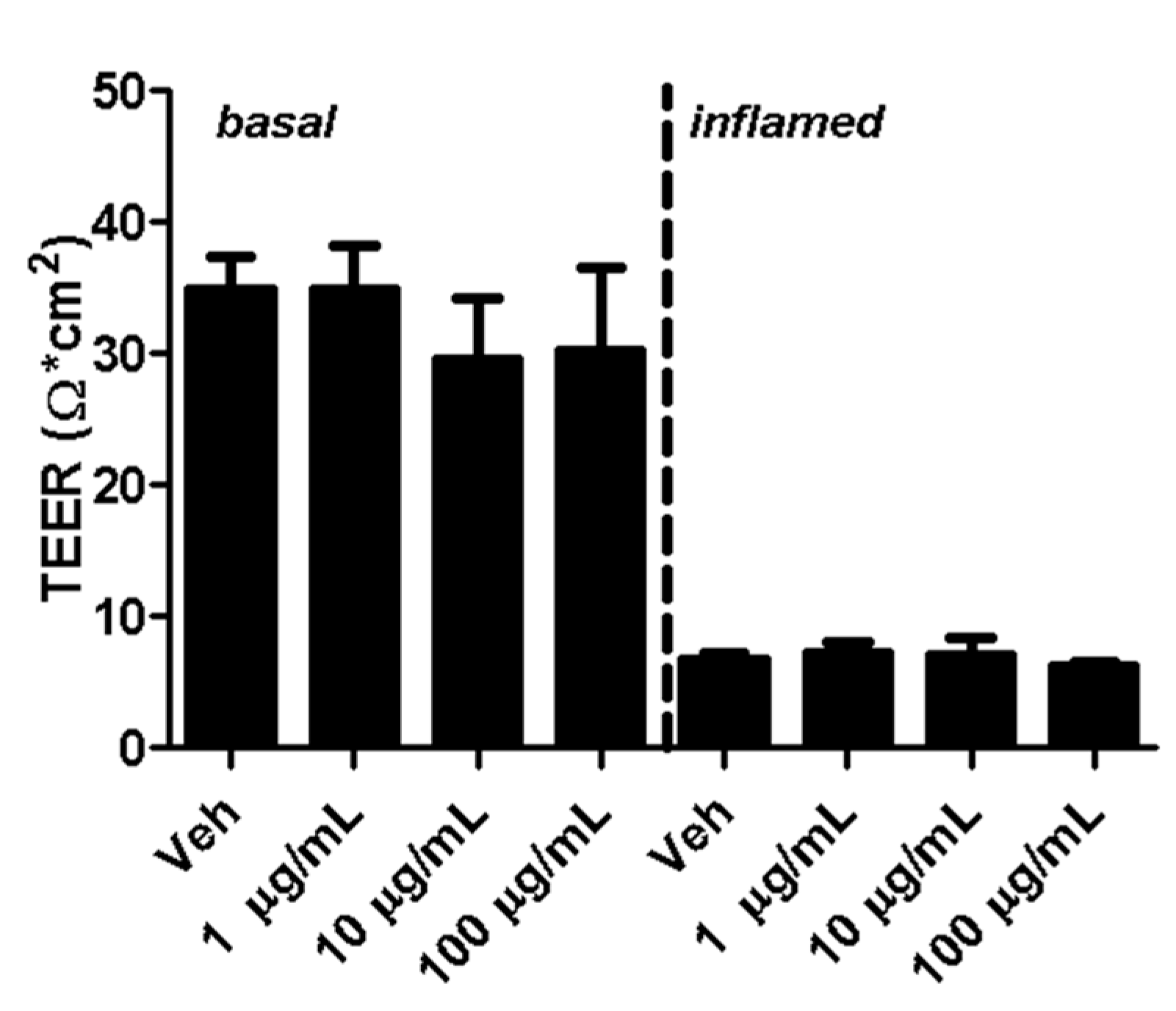
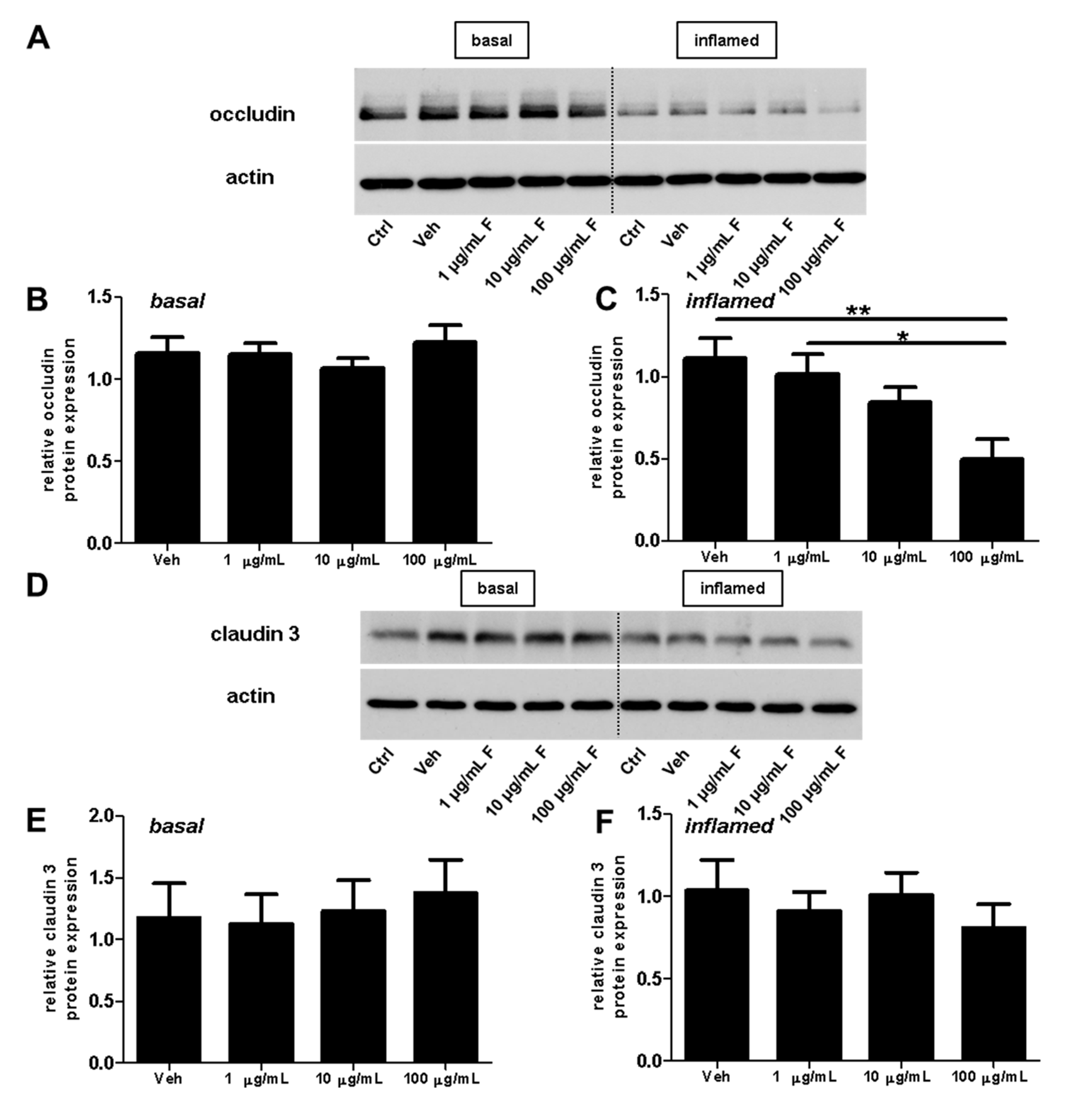
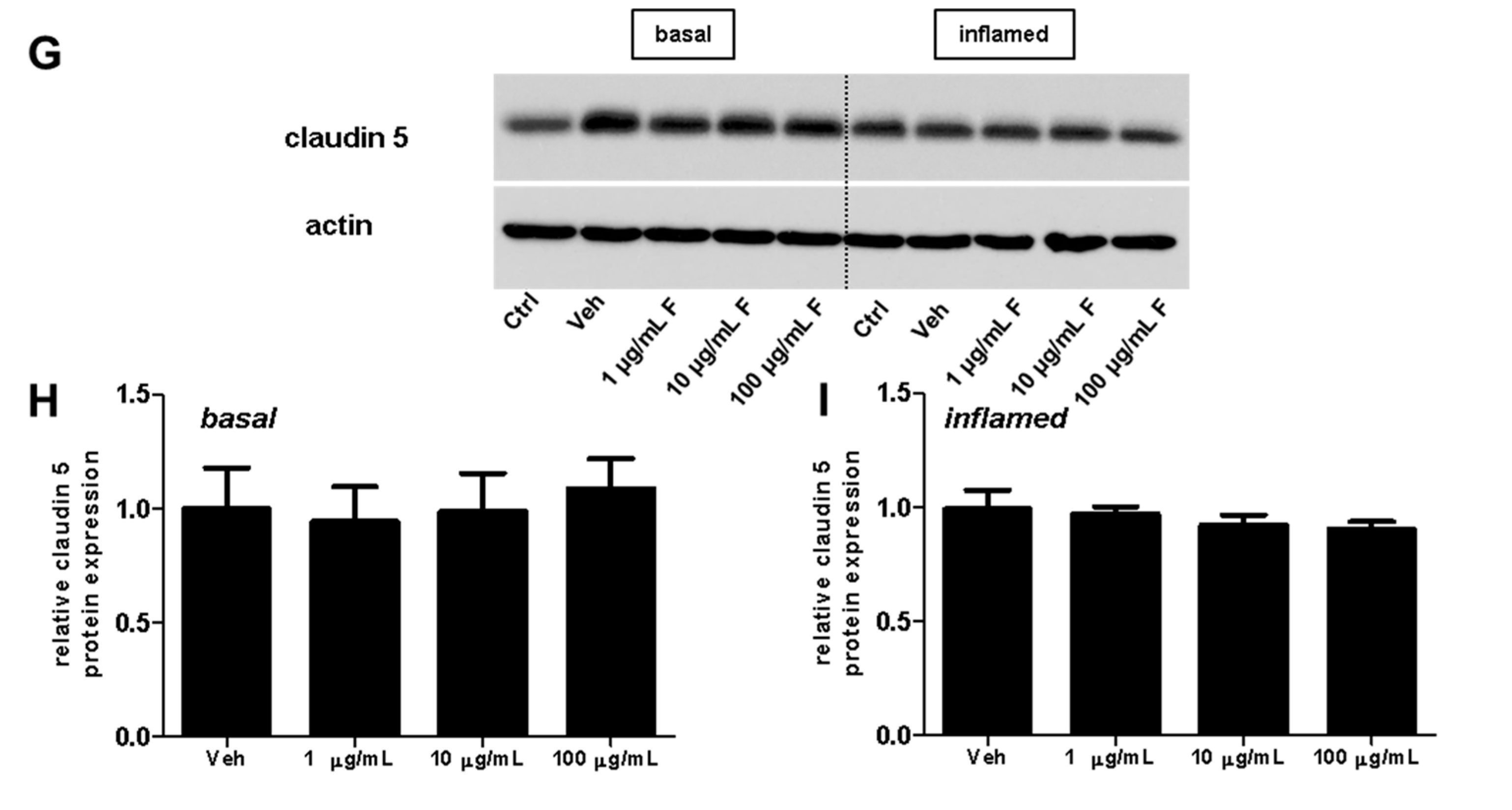
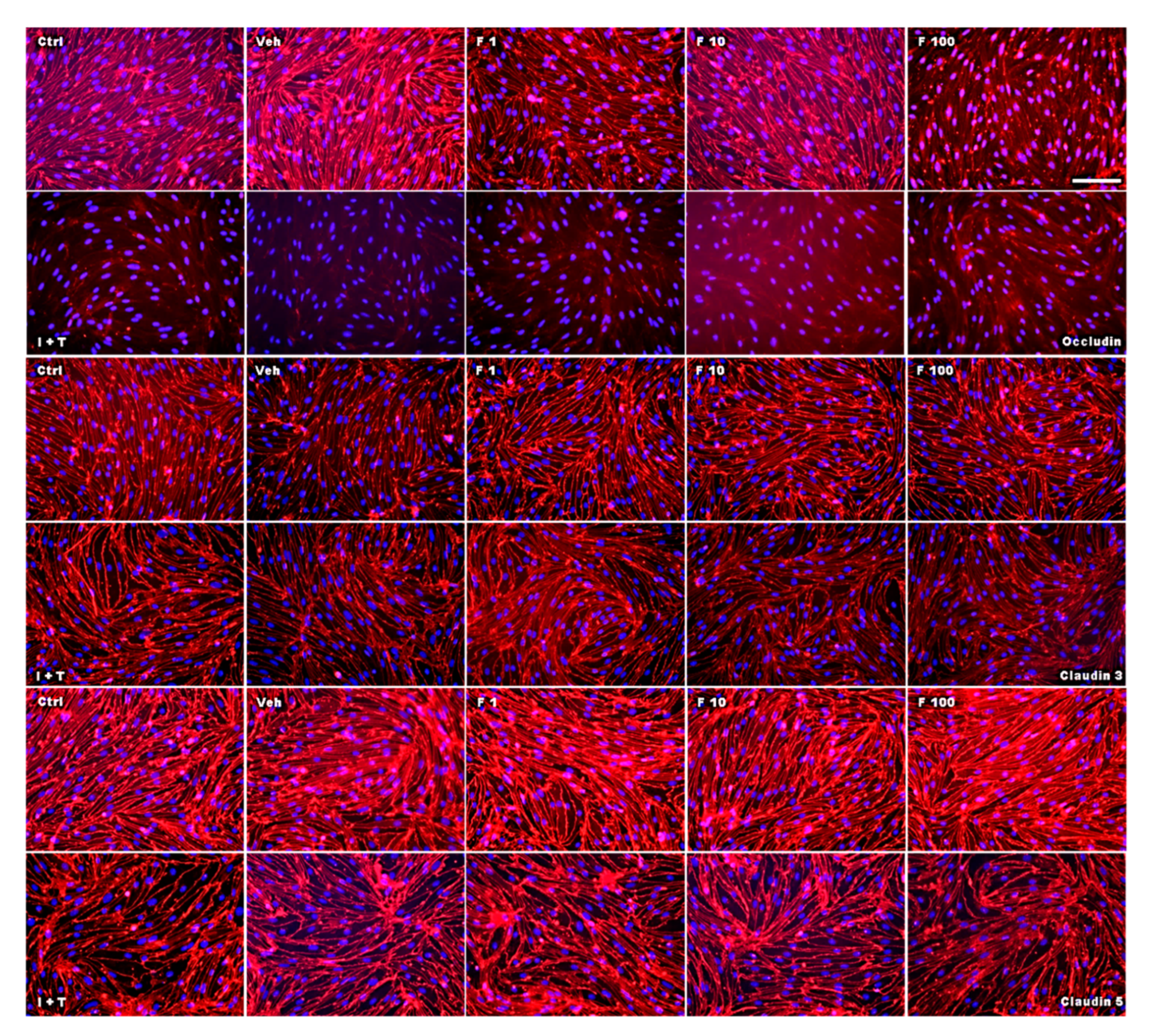
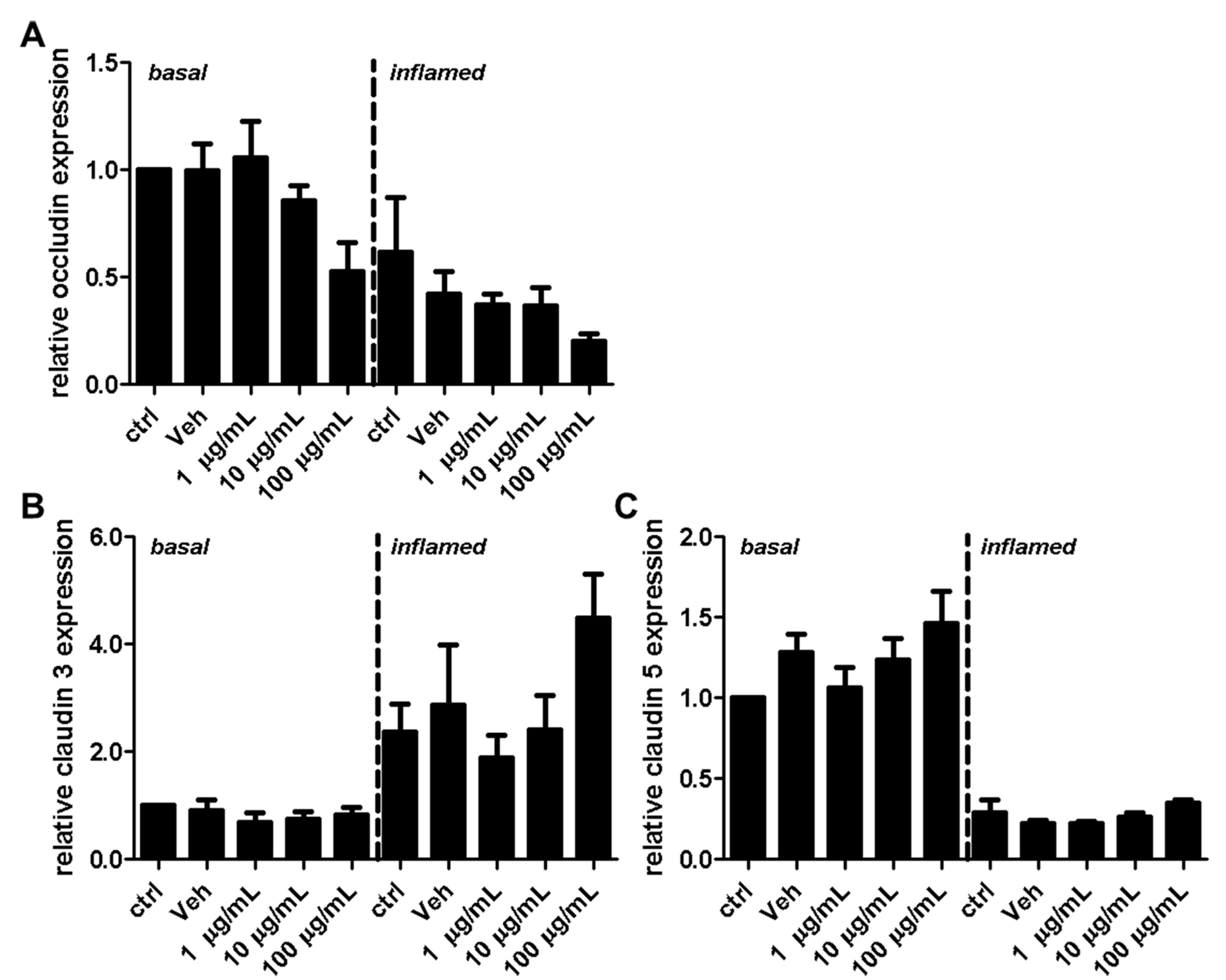
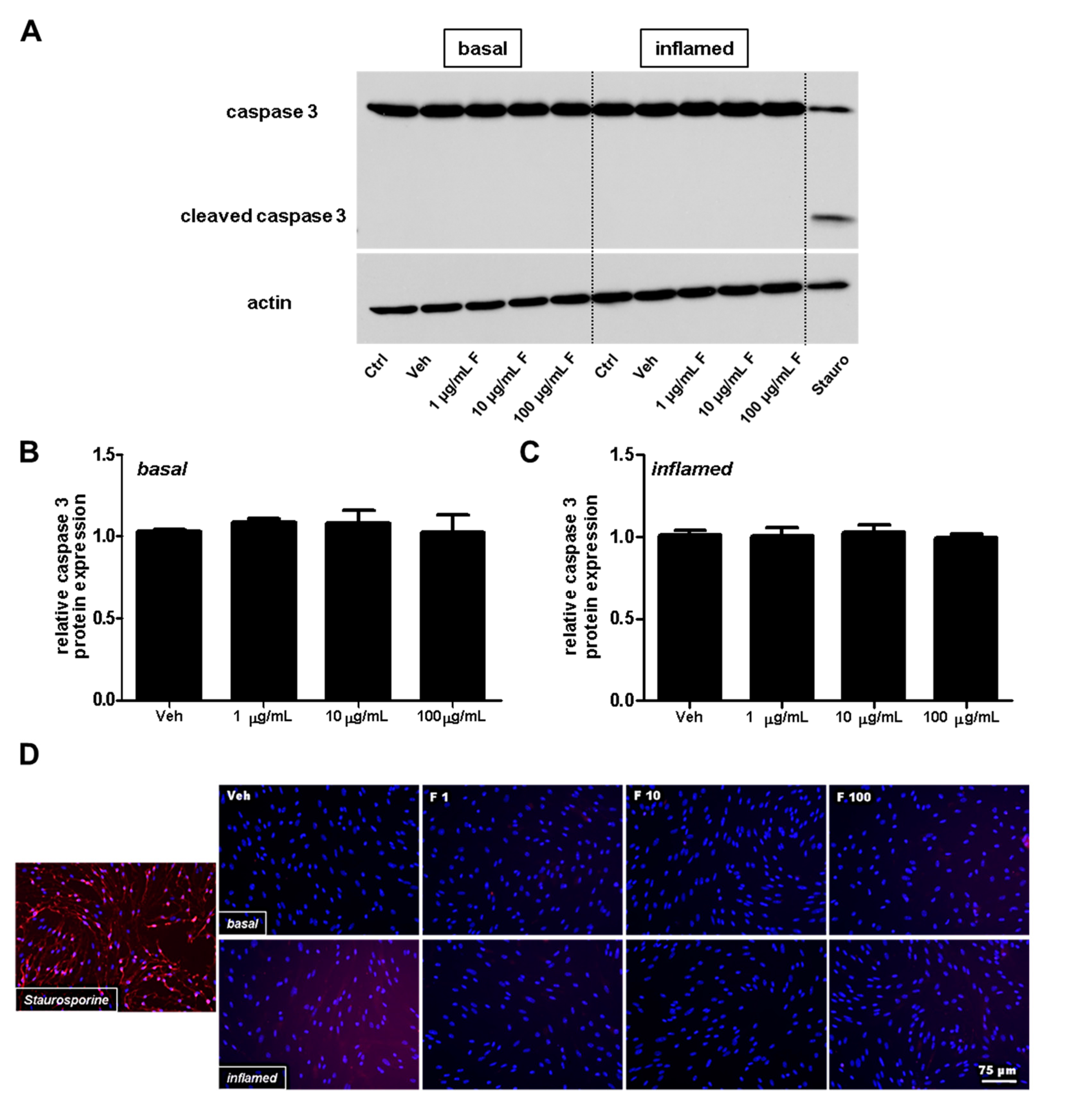
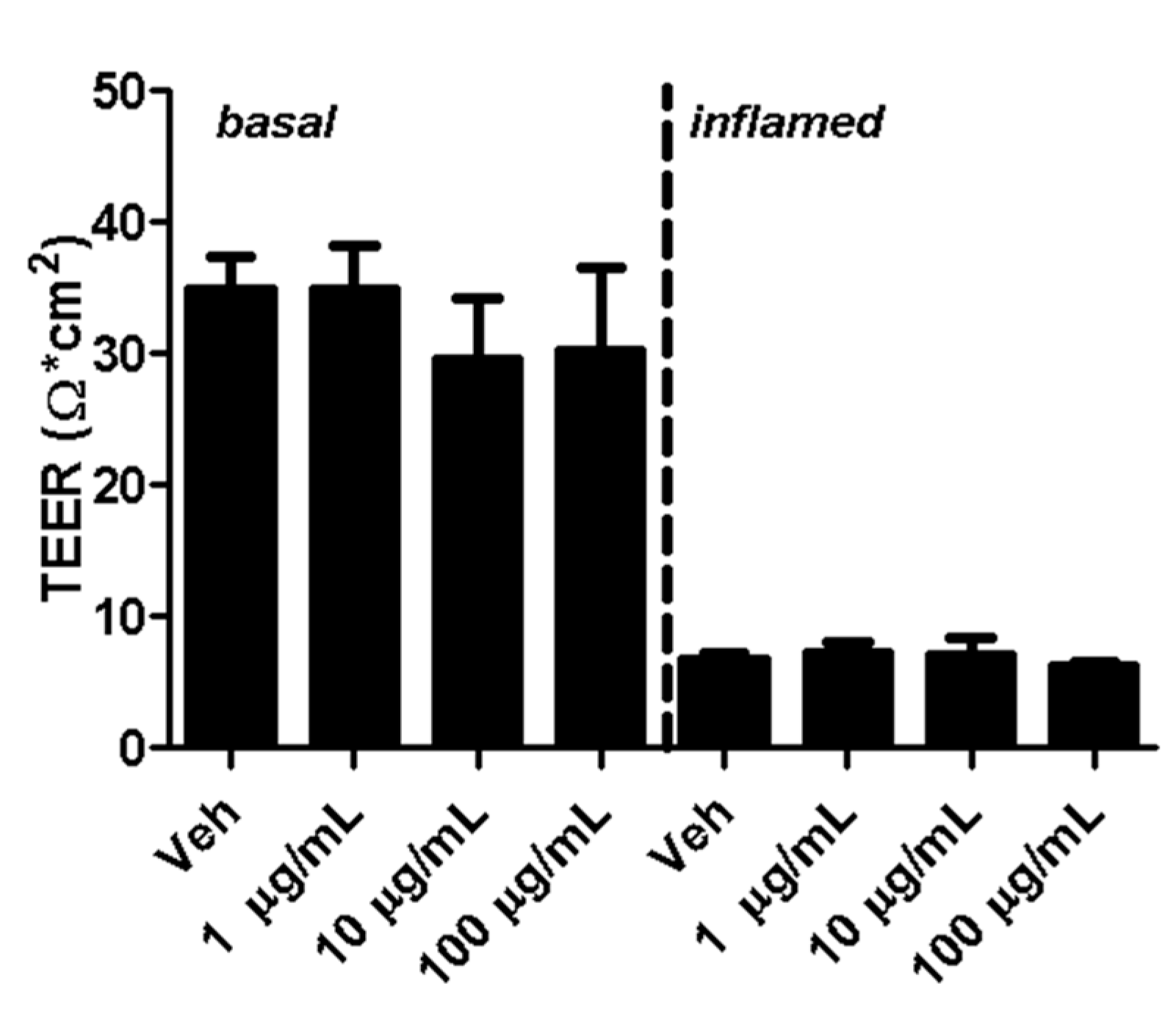
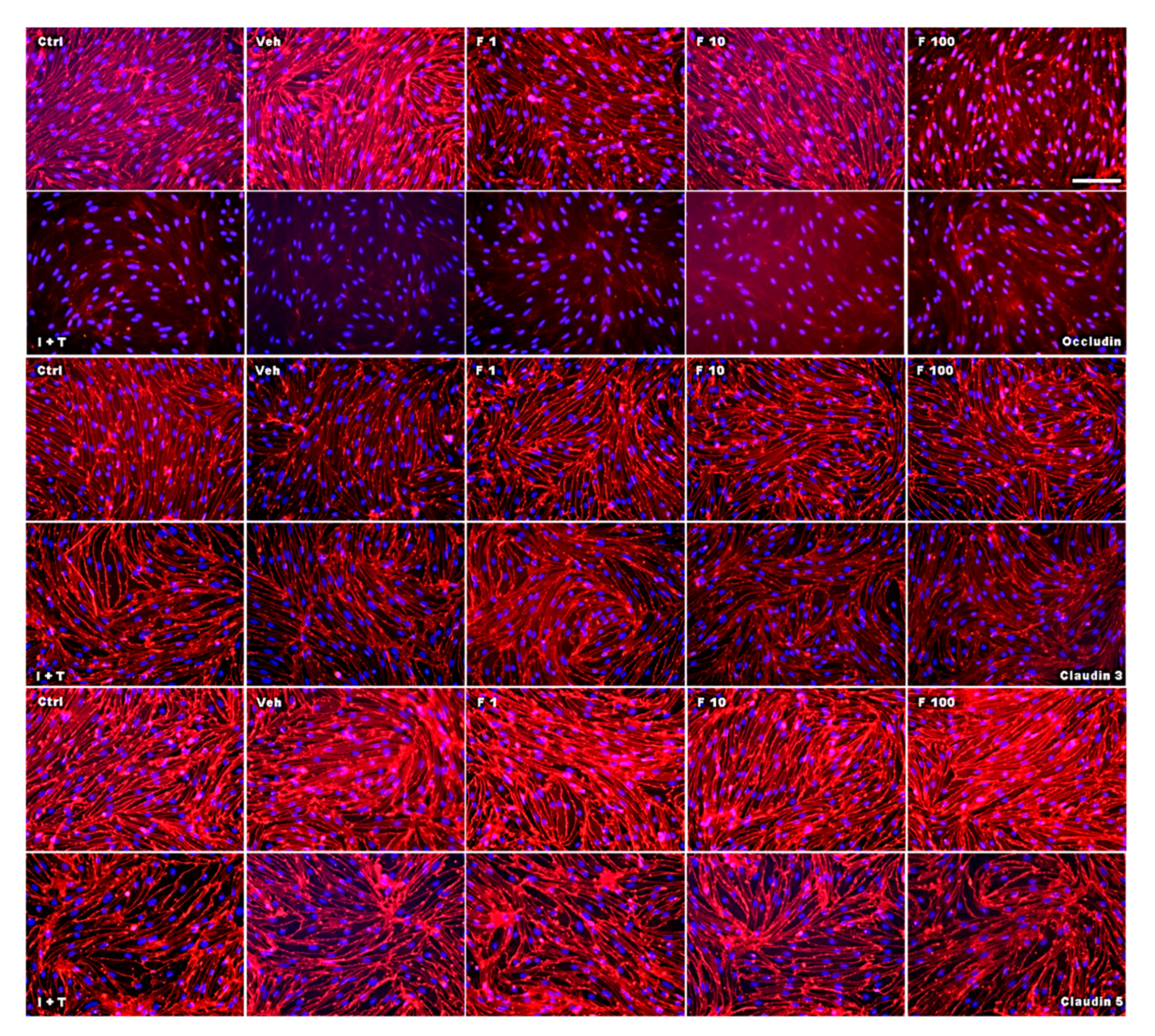
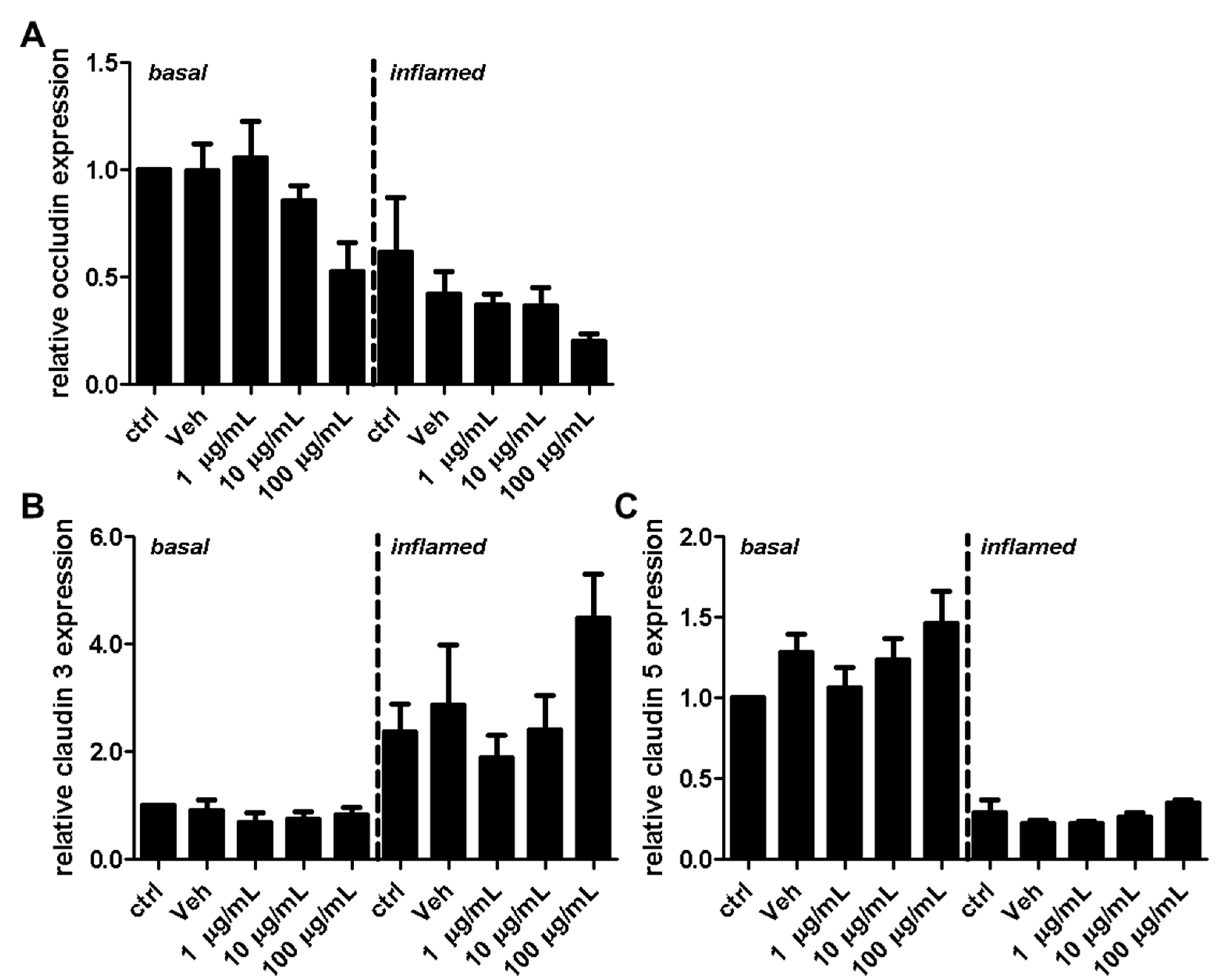
2.2. Fullerenols Do Not Modulate MBMEC Barrier Functions
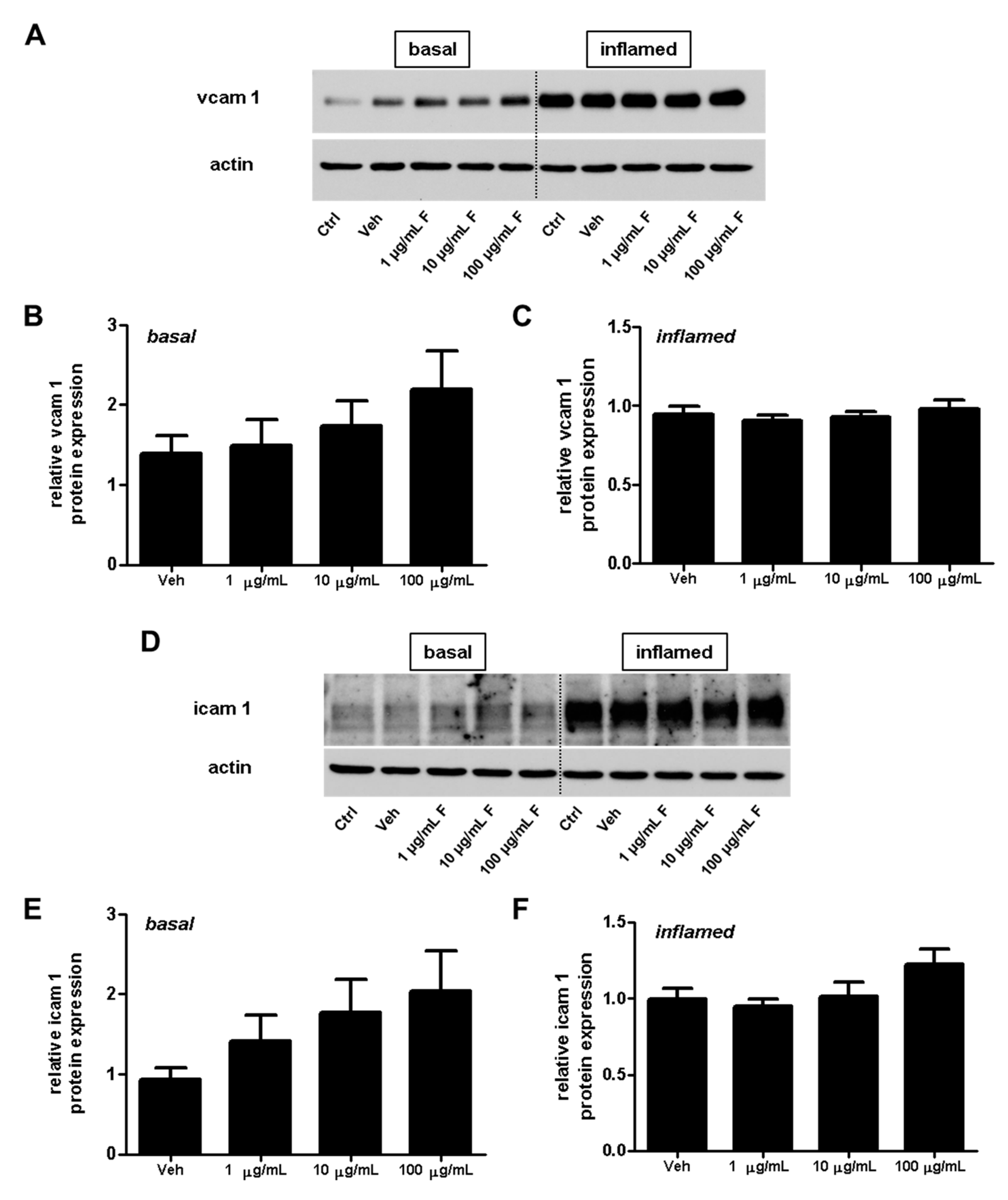
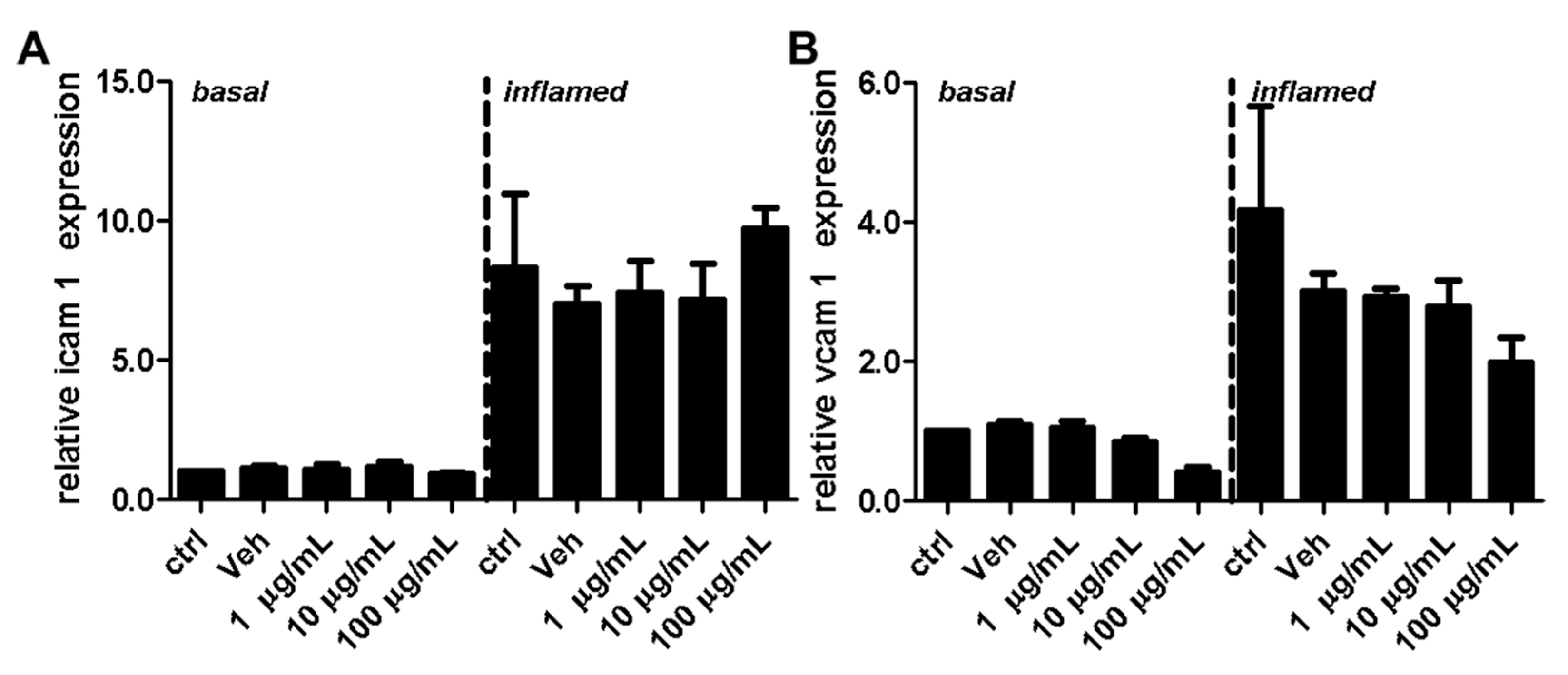
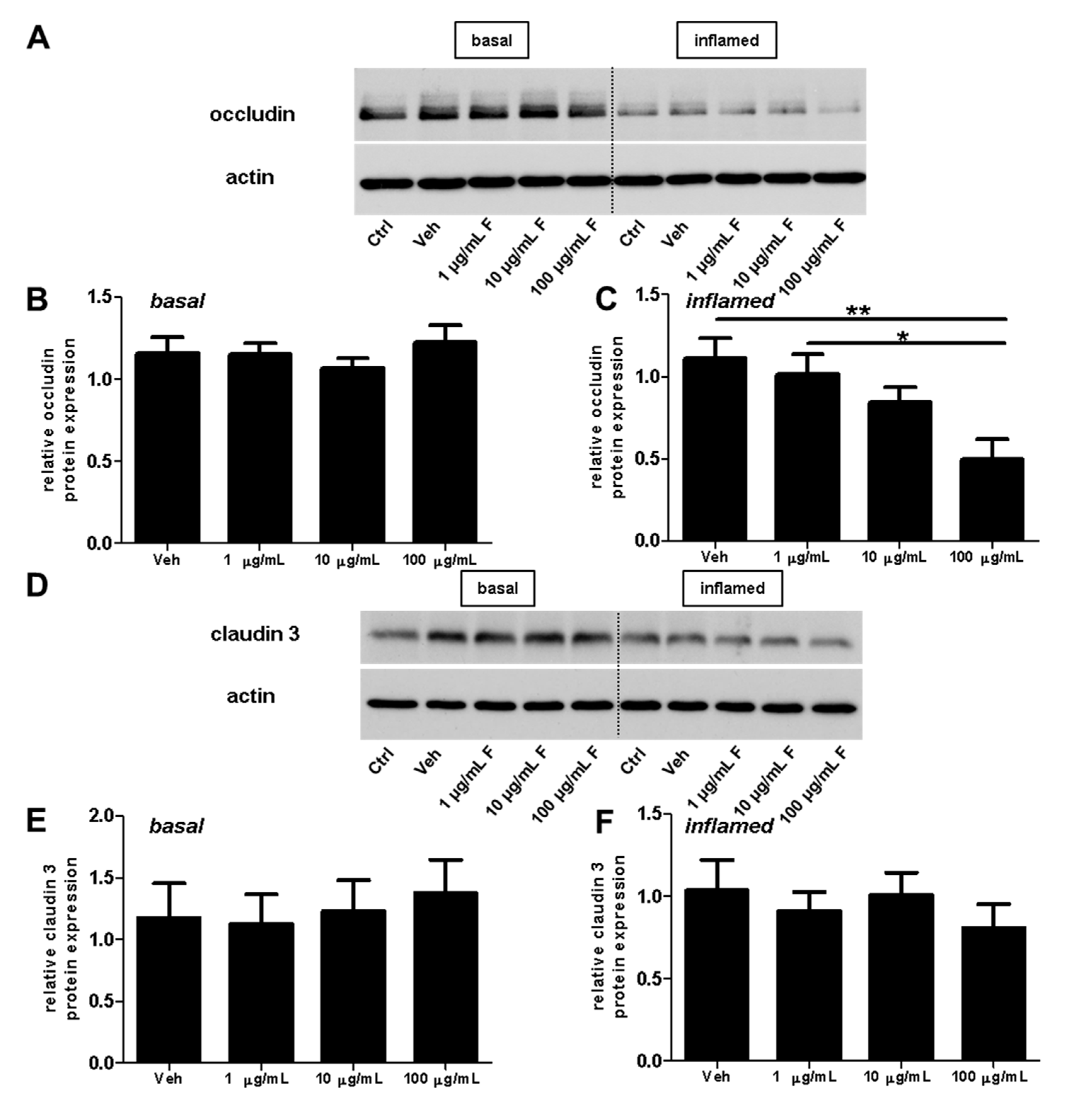
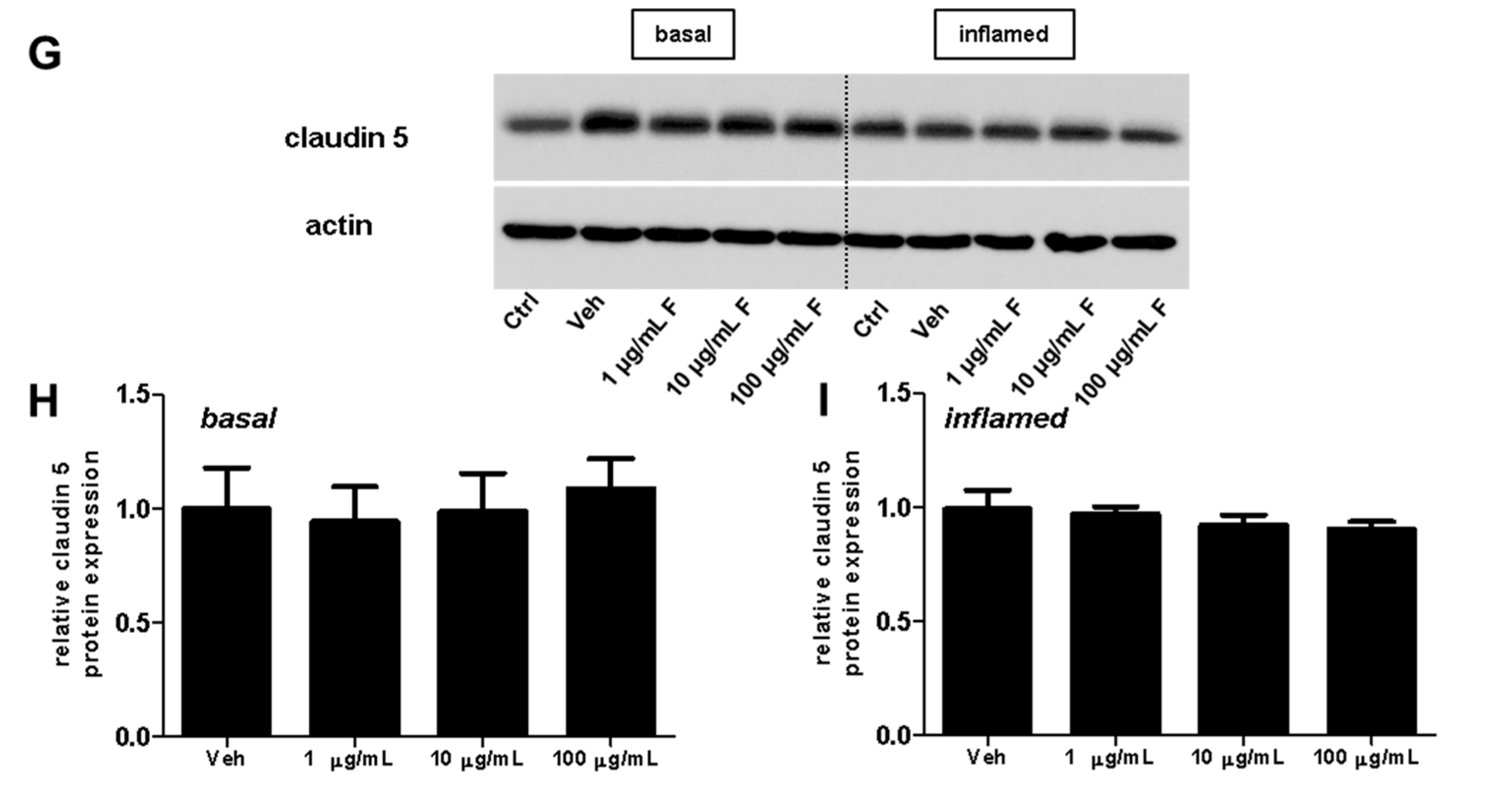
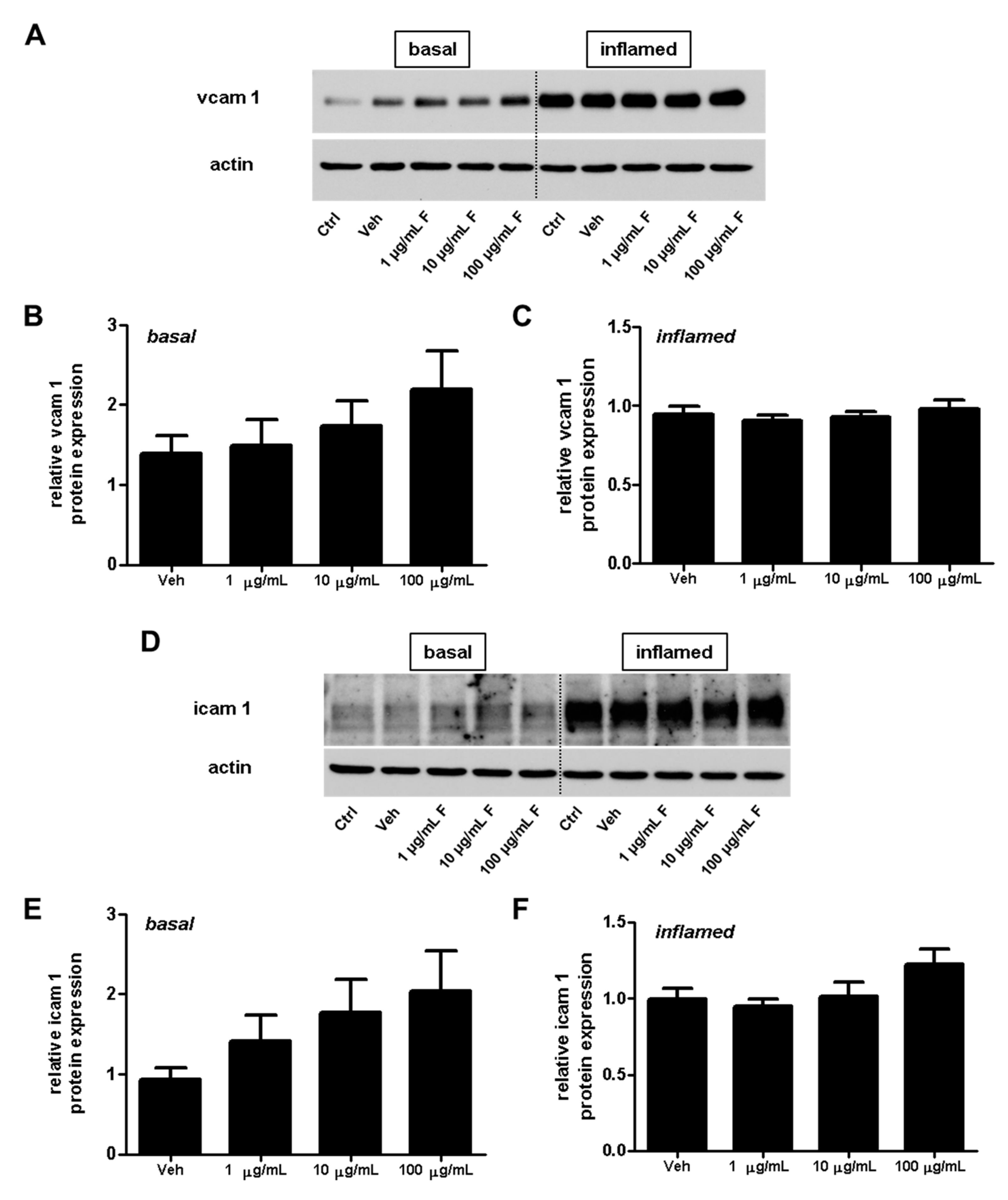
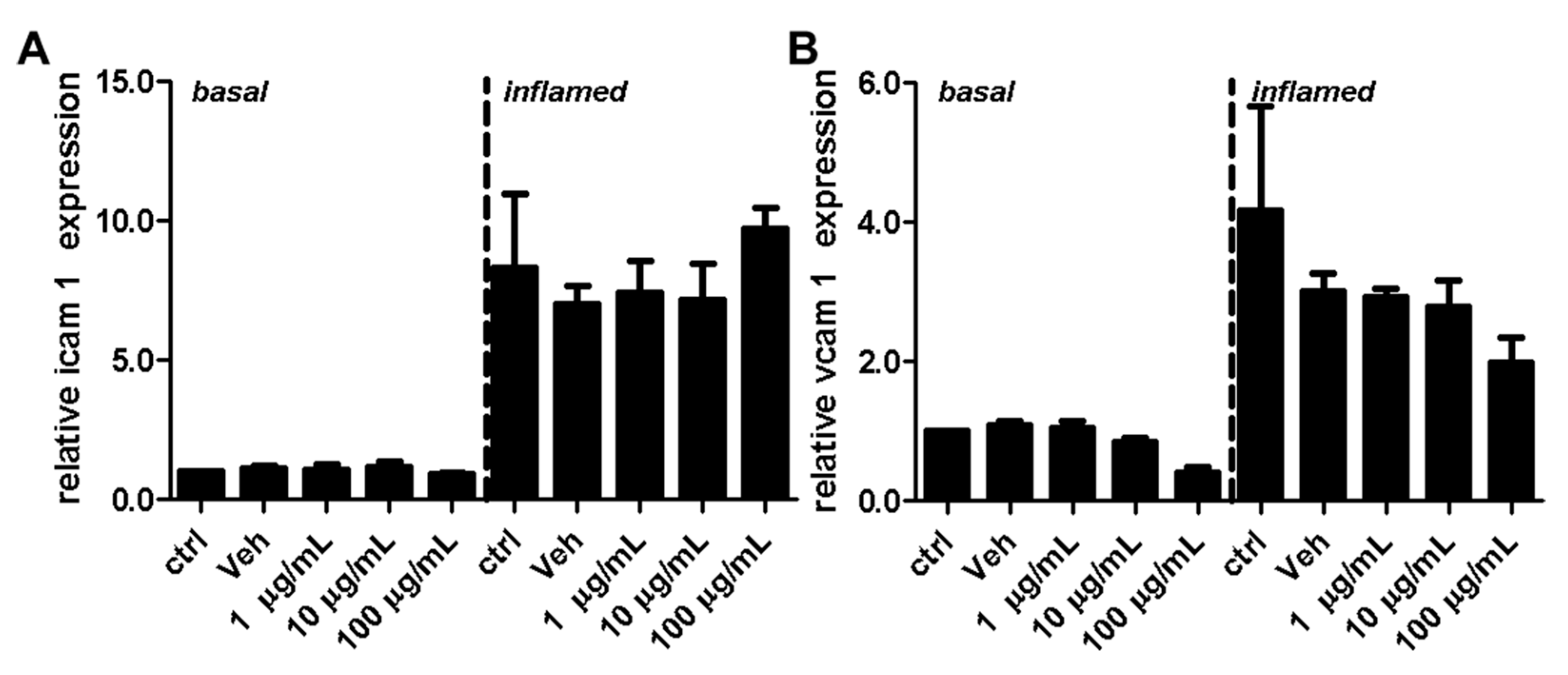
2.3. Fullerenols Do Not Modulate the Expression of Adhesion Molecules in MBMEC
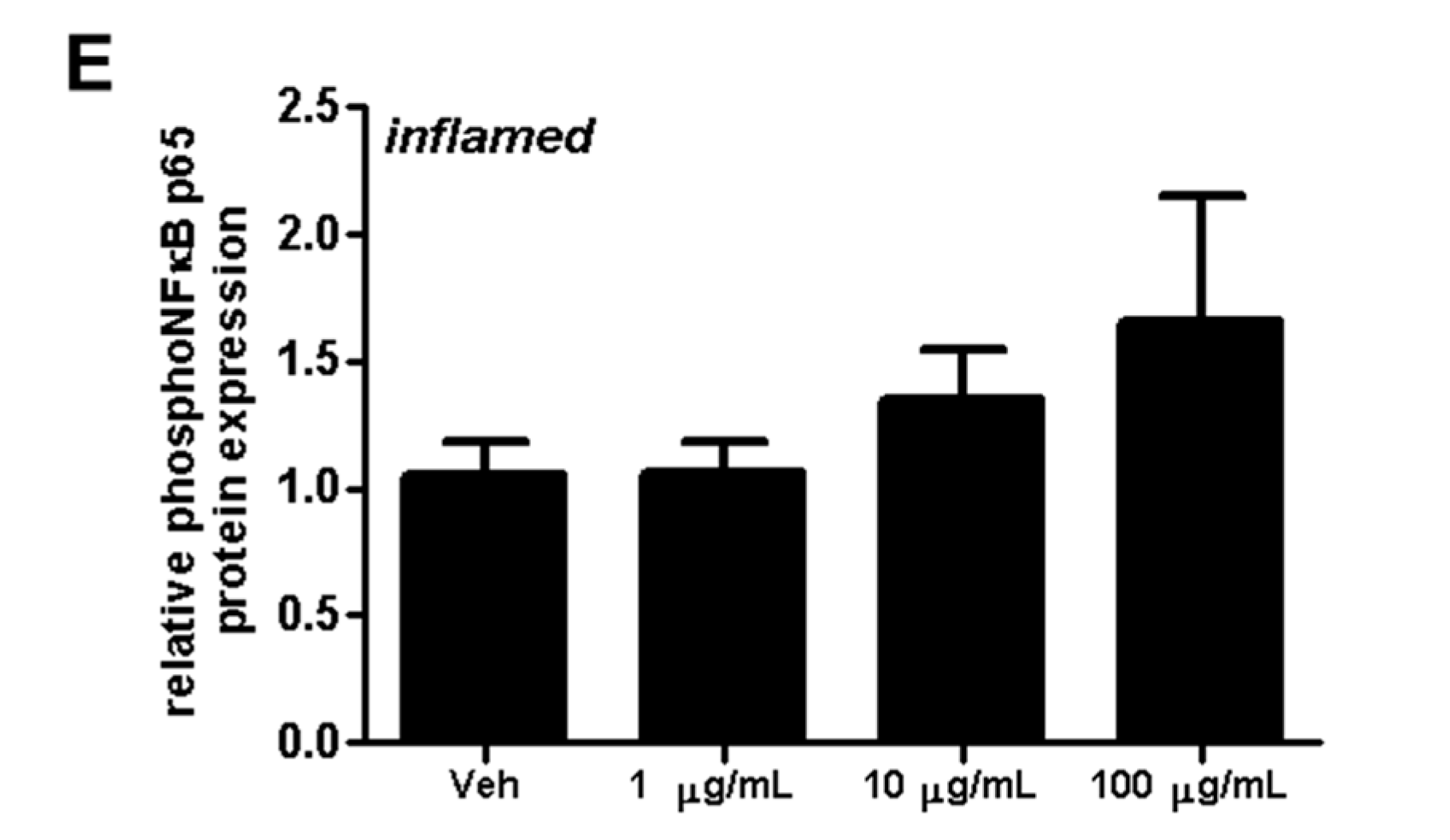
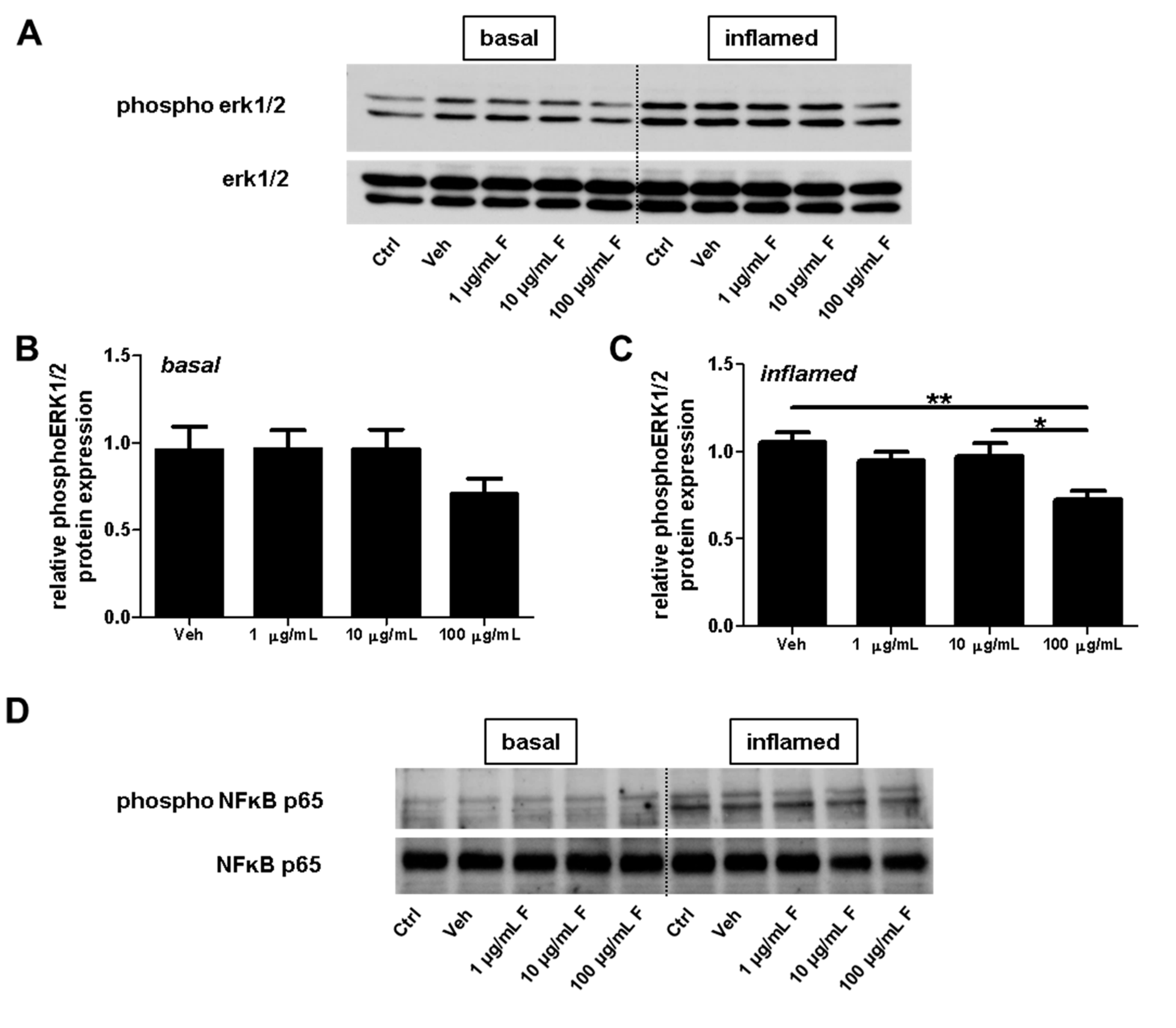
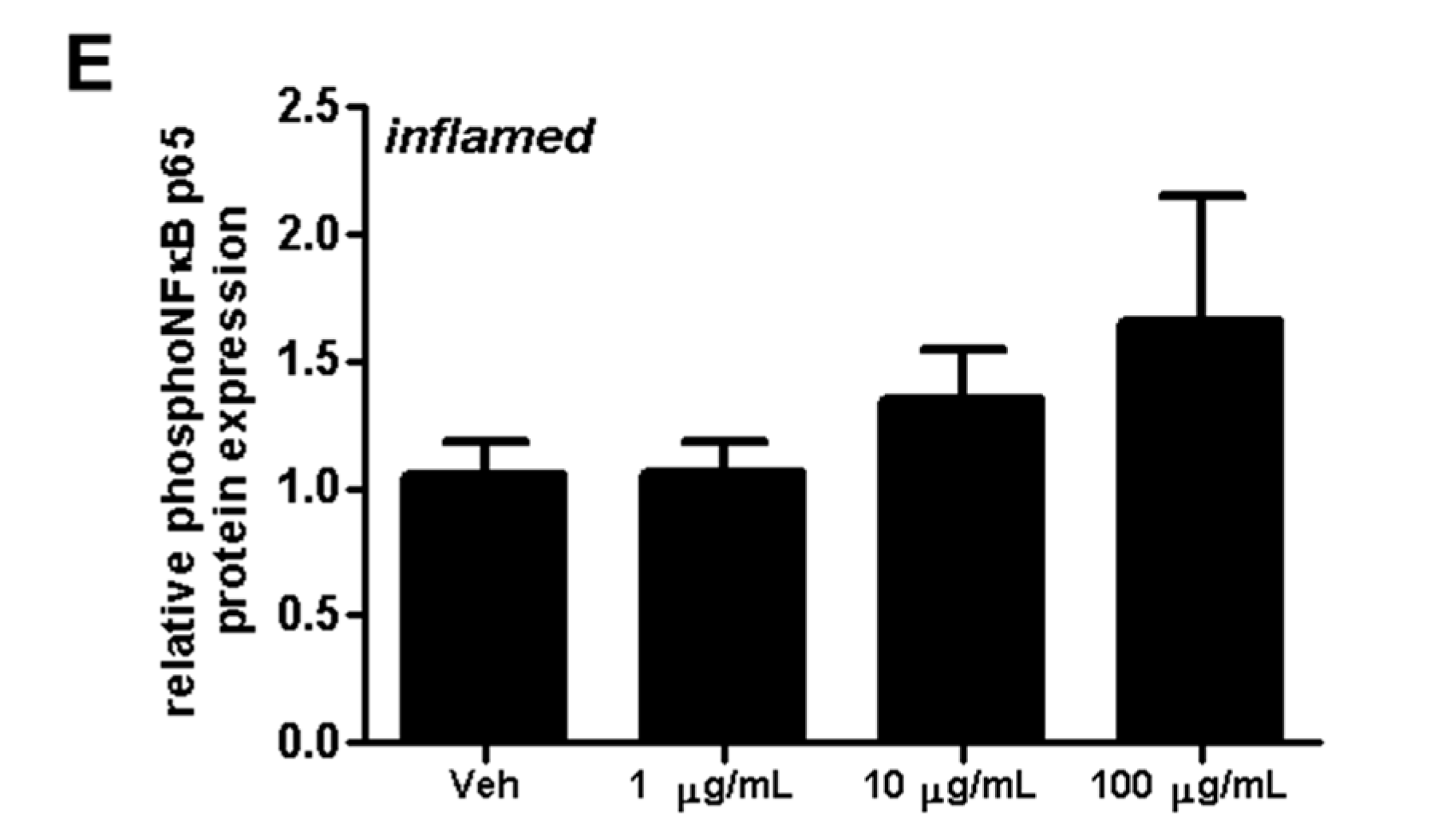
2.4. Fullerenols Impact erk1/2 and NFĸB Activation in MBMEC
3. Discussion
4. Materials and Methods
4.1. Materials
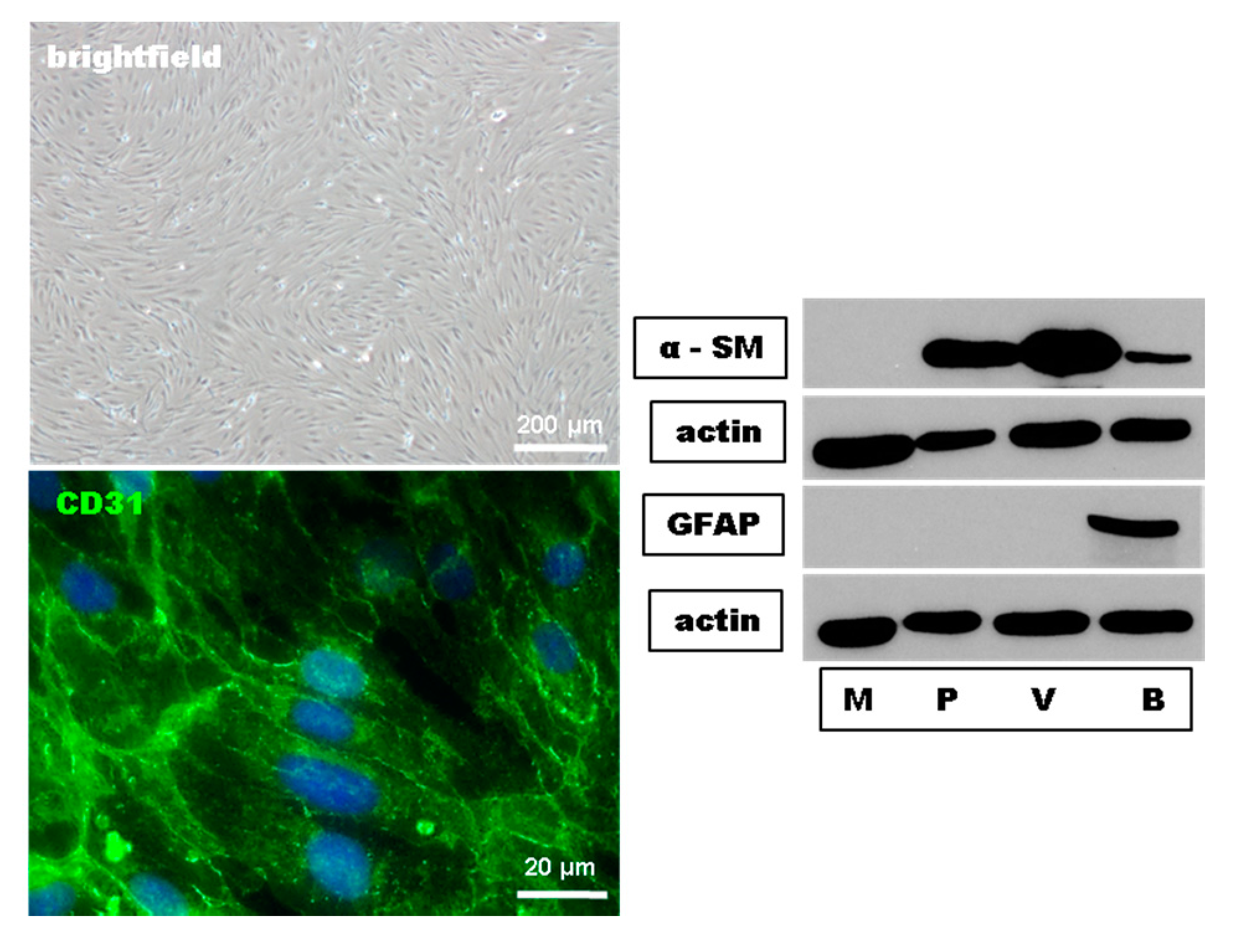
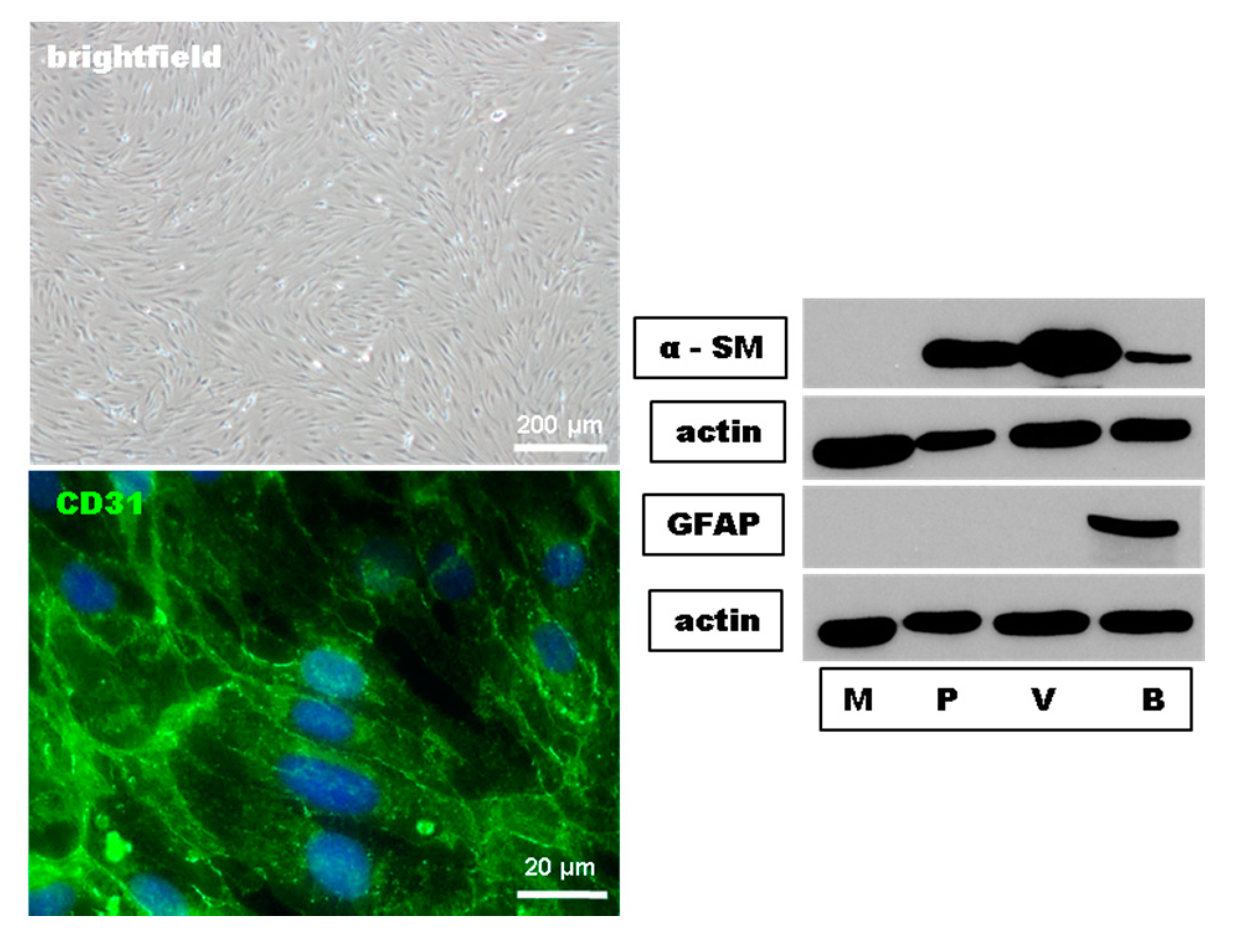
4.2. Preparation and Cultivation of MBMEC
4.3. Immunocytochemistry
4.4. Measurement of GSH
4.5. Measurement of Transendothelial Electrical Resistance (TEER)
4.6. Western Blot Assays
4.7. RT-PCR
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Engelhardt, B.; Ransohoff, R.M. Capture, crawl, cross: The T cell code to breach the blood-brain barriers. Trends Immunol. 2012, 33, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Barten, D.M.; Ruddle, N.H. Vascular cell adhesion molecule-1 modulation by tumor necrosis factor in experimental allergic encephalomyelitis. J. Neuroimmunol. 1994, 51, 123–133. [Google Scholar] [CrossRef]
- Wong, D.; Prameya, R.; Dorovini-Zis, K. In vitro adhesion and migration of T lymphocytes across monolayers of human brain microvessel endothelial cells: Regulation by icam-1, vcam-1, e-selectin and pecam-1. J. Neuropathol. Exp. Neurol. 1999, 58, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Bhattacharya, S.; Tcheranova, D.; Qu, Y.; Regan, R.F.; Leffler, C.W.; Parfenova, H. Ho-2 provides endogenous protection against oxidative stress and apoptosis caused by TNF-α in cerebral vascular endothelial cells. Am. J. Physiol. Cell Physiol. 2006, 291, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, K.D.; Collins, L.E.; Murphy, R.P.; Cummins, P.M. Downregulation of blood-brain barrier phenotype by proinflammatory cytokines involves NADPH oxidase-dependent ROS generation: Consequences for interendothelial adherens and tight junctions. PLoS ONE 2014, 9, e101815. [Google Scholar] [CrossRef] [PubMed]
- Bo, L.; Dawson, T.M.; Wesselingh, S.; Mork, S.; Choi, S.; Kong, P.A.; Hanley, D.; Trapp, B.D. Induction of nitric oxide synthase in demyelinating regions of multiple sclerosis brains. Ann. Neurol. 1994, 36, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Leppert, D.; Waubant, E.; Galardy, R.; Bunnett, N.W.; Hauser, S.L. T cell gelatinases mediate basement membrane transmigration in vitro. J. Immunol. 1995, 154, 4379–4389. [Google Scholar] [PubMed]
- Traore, K.; Sharma, R.; Thimmulappa, R.K.; Watson, W.H.; Biswal, S.; Trush, M.A. Redox-regulation of erk1/2-directed phosphatase by reactive oxygen species: Role in signaling TPA-induced growth arrest in ML-1 cells. J. Cell. Physiol. 2008, 216, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Winyard, P.G.; Blake, D.R. Antioxidants, redox-regulated transcription factors, and inflammation. Adv. Pharmacol. 1997, 38, 403–421. [Google Scholar] [PubMed]
- Karg, E.; Klivenyi, P.; Nemeth, I.; Bencsik, K.; Pinter, S.; Vecsei, L. Nonenzymatic antioxidants of blood in multiple sclerosis. J. Neurol. 1999, 246, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, E.; Isobe, H. Functionalized fullerenes in water. The first 10 years of their chemistry, biology, and nanoscience. Acc. Chem. Res. 2003, 36, 807–815. [Google Scholar] [PubMed]
- Bosi, S.; da Ros, T.; Spalluto, G.; Prato, M. Fullerene derivatives: An attractive tool for biological applications. Eur. J. Med. Chem. 2003, 38, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Dugan, L.L.; Turetsky, D.M.; Du, C.; Lobner, D.; Wheeler, M.; Almli, C.R.; Shen, C.K.; Luh, T.Y.; Choi, D.W.; Lin, T.S. Carboxyfullerenes as neuroprotective agents. Proc. Natl. Acad. Sci. USA 1997, 94, 9434–9439. [Google Scholar] [CrossRef] [PubMed]
- Krusic, P.J.; Wasserman, E.; Keizer, P.N.; Morton, J.R.; Preston, K.F. Radical reactions of C60. Science 1991, 254, 1183–1185. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.S.; Chen, W.J.; Chiang, L.Y. Free radical scavenging activity of fullerenol on the ischemia-reperfusion intestine in dogs. World J. Surg. 2000, 24, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.B.; Pluta, R.M.; Walbridge, S.; Wink, D.A.; Oldfield, E.H.; Boock, R.J. Production of reactive oxygen species after reperfusion in vitro and in vivo: Protective effect of nitric oxide. J. Neurosurg. 2000, 93, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fluri, F.; Grunstein, D.; Cam, E.; Ungethuem, U.; Hatz, F.; Schafer, J.; Samnick, S.; Israel, I.; Kleinschnitz, C.; Orts-Gil, G.; et al. Fullerenols and glucosamine fullerenes reduce infarct volume and cerebral inflammation after ischemic stroke in normotensive and hypertensive rats. Exp. Neurol. 2015, 265, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Basso, A.S.; Frenkel, D.; Quintana, F.J.; Costa-Pinto, F.A.; Petrovic-Stojkovic, S.; Puckett, L.; Monsonego, A.; Bar-Shir, A.; Engel, Y.; Gozin, M.; et al. Reversal of axonal loss and disability in a mouse model of progressive multiple sclerosis. J. Clin. Investig. 2008, 118, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Vani, J.R.; Mohammadi, M.T.; Foroshani, M.S.; Jafari, M. Polyhydroxylated fullerene nanoparticles attenuate brain infarction and oxidative stress in rat model of ischemic stroke. EXCLI J. 2016, 15, 378–390. [Google Scholar] [PubMed]
- Yamawaki, H.; Iwai, N. Cytotoxicity of water-soluble fullerene in vascular endothelial cells. Am. J. Physiol. Cell Physiol. 2006, 290, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Gelderman, M.P.; Simakova, O.; Clogston, J.D.; Patri, A.K.; Siddiqui, S.F.; Vostal, A.C.; Simak, J. Adverse effects of fullerenes on endothelial cells: Fullerenol C60(OH)24 induced tissue factor and ICAM-1 membrane expression and apoptosis in vitro. Int. J. Nanomed. 2008, 3, 59–68. [Google Scholar]
- Schuhmann, M.K.; Bittner, S.; Meuth, S.G.; Kleinschnitz, C.; Fluri, F. Fingolimod (FTY720-P) does not stabilize the blood-brain barrier under inflammatory conditions in an in vitro model. Int. J. Mol. Sci. 2015, 16, 29454–29466. [Google Scholar] [CrossRef] [PubMed]
- Steiner, O.; Coisne, C.; Cecchelli, R.; Boscacci, R.; Deutsch, U.; Engelhardt, B.; Lyck, R. Differential roles for endothelial ICAM-1, ICAM-2, and VCAM-1 in shear-resistant T cell arrest, polarization, and directed crawling on blood-brain barrier endothelium. J. Immunol. 2010, 185, 4846–4855. [Google Scholar] [CrossRef] [PubMed]
- Taskin, O.; Buhur, A.; Birincioglu, M.; Burak, F.; Atmaca, R.; Yilmaz, I.; Wheeler, J.M. The effects of duration of CO2 insufflation and irrigation on peritoneal microcirculation assessed by free radical scavengers and total glutathion levels during operative laparoscopy. J. Am. Assoc. Gynecol. Laparosc. 1998, 5, 129–133. [Google Scholar] [CrossRef]
- Xu, J.Y.; Su, Y.Y.; Cheng, J.S.; Li, S.X.; Liu, R.L.; Li, W.X.; Xu, G.T.; Li, Q.N. Protective effects of fullerenol on carbon tetrachloride-induced acute hepatotoxicity and nephrotoxicity in rats. Carbon 2010, 48, 1388–1396. [Google Scholar] [CrossRef]
- White, C.D.; Sacks, D.B. Regulation of MAP kinase signaling by calcium. Methods Mol. Biol. 2010, 661, 151–165. [Google Scholar] [PubMed]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.L.; van der Pol, S.M.; di Dio, F.; van Het Hof, B.; Kooij, G.; de Vries, H.E.; van Horssen, J. Protective effects of monomethyl fumarate at the inflamed blood-brain barrier. Microvasc. Res. 2016, 105, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Read, M.A.; Neish, A.S.; Whitley, M.Z.; Thanos, D.; Maniatis, T. Transcriptional regulation of endothelial cell adhesion molecules: NF-κB and cytokine-inducible enhancers. FASEB J. 1995, 9, 899–909. [Google Scholar] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and nf-kappab signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D.; Gilmore, T.D. Good cop, bad cop: The different faces of NF-κB. Cell Death Differ. 2006, 13, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, Y.; Sun, B.; Li, H.; Dong, J.; Zhang, L.; Wang, L.; Wang, P.; Zhao, Y.; Chen, C. Polyhydroxylated metallofullerenols stimulate IL-1β secretion of macrophage through TLRs/MyD88/ NF-κB pathway and NLRP3 inflammasome activation. Small 2014, 10, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Aoshima, H.; Saitoh, Y.; Miwa, N. Fullerene-polyvinylpyrrolidone clathrate localizes in the cytoplasm to prevent ultraviolet-A ray-induced DNA-fragmentation and activation of the transcriptional factor NF-κB. J. Cell. Biochem. 2010, 111, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Seo, J.H.; Wong, K.H.; Terasaki, Y.; Park, J.; Bong, K.; Arai, K.; Lo, E.H.; Irimia, D. Three-dimensional blood-brain barrier model for in vitro studies of neurovascular pathology. Sci. Rep. 2015, 5, 15222. [Google Scholar] [CrossRef] [PubMed]
- Bittner, S.; Ruck, T.; Schuhmann, M.K.; Herrmann, A.M.; Moha ou Maati, H.; Bobak, N.; Gobel, K.; Langhauser, F.; Stegner, D.; Ehling, P.; et al. Endothelial TWIK-related potassium channel-1 (TREK1) regulates immune-cell trafficking into the CNS. Nat. Med. 2013, 19, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuhmann, M.K.; Fluri, F. Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells. Int. J. Mol. Sci. 2017, 18, 1783. https://doi.org/10.3390/ijms18081783
Schuhmann MK, Fluri F. Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells. International Journal of Molecular Sciences. 2017; 18(8):1783. https://doi.org/10.3390/ijms18081783
Chicago/Turabian StyleSchuhmann, Michael K., and Felix Fluri. 2017. "Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells" International Journal of Molecular Sciences 18, no. 8: 1783. https://doi.org/10.3390/ijms18081783
APA StyleSchuhmann, M. K., & Fluri, F. (2017). Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells. International Journal of Molecular Sciences, 18(8), 1783. https://doi.org/10.3390/ijms18081783