Targeting Apoptotic Activity Against Prostate Cancer Stem Cells



Abstract
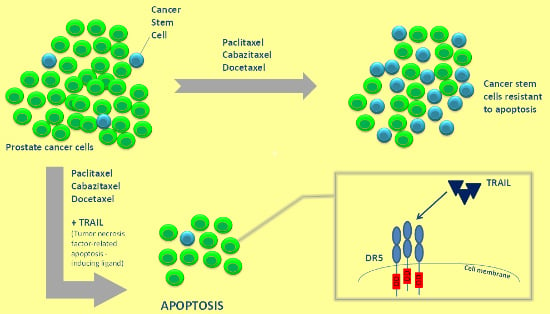
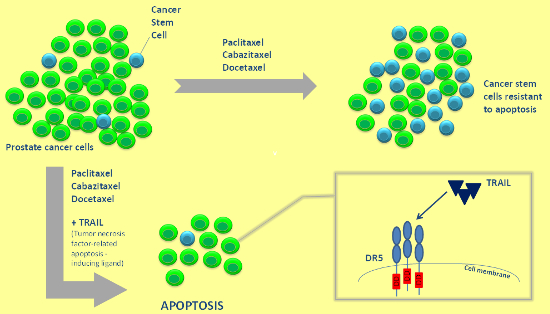{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
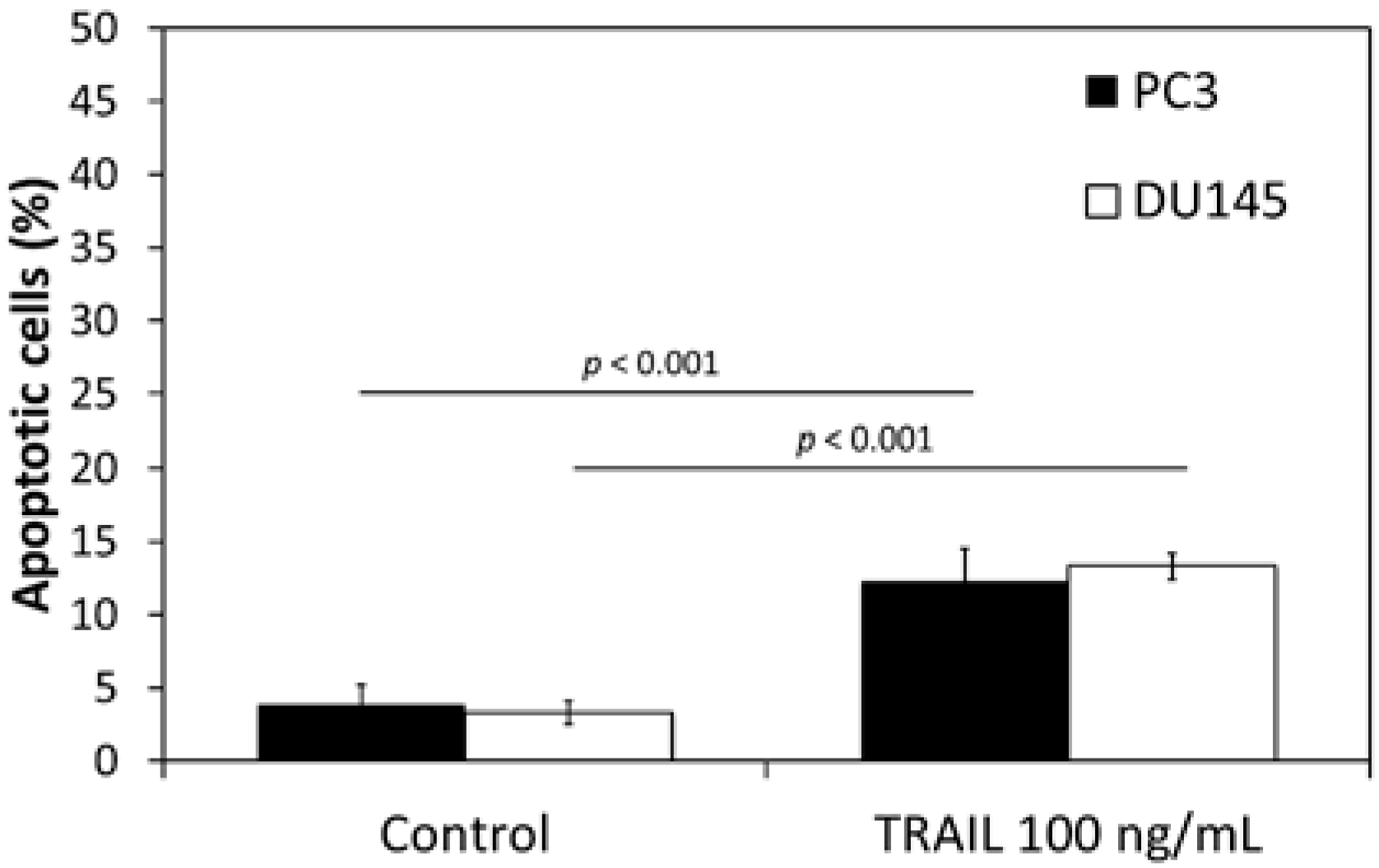
2.1. Apoptotic Activity of TRAIL (Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand) in DU145 and PC3 Prostate Cancer Cell Lines
2.2. Apoptotic Activity of Paclitaxel, Cabazitaxel or Docetaxel in DU145 and PC3 Prostate Cancer Cell Lines
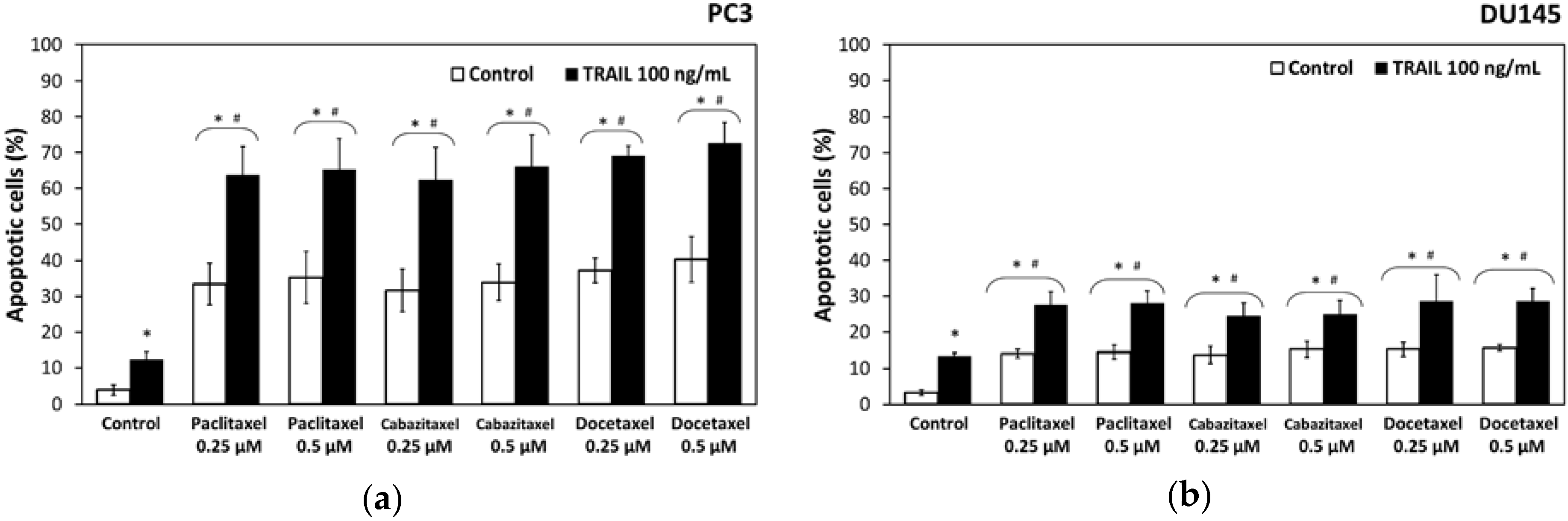
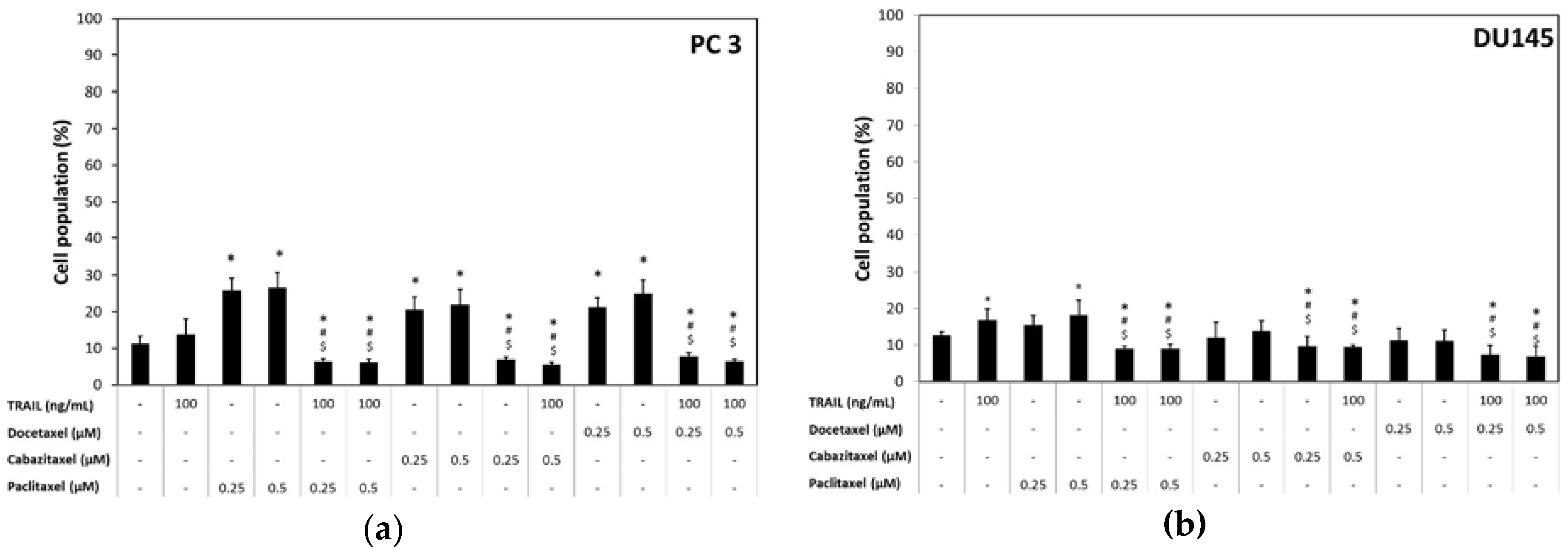
2.3. Apoptotic Activity of TRAIL in Combination with Paclitaxel, Cabazitaxel or Docetaxel in DU145 and PC3 Prostate Cancer Cell Lines
2.4. Necrotic Effect of TRAIL and/or Paclitaxel, Cabazitaxel, Docetaxel in DU145 and PC3 Prostate Cancer Cell Lines
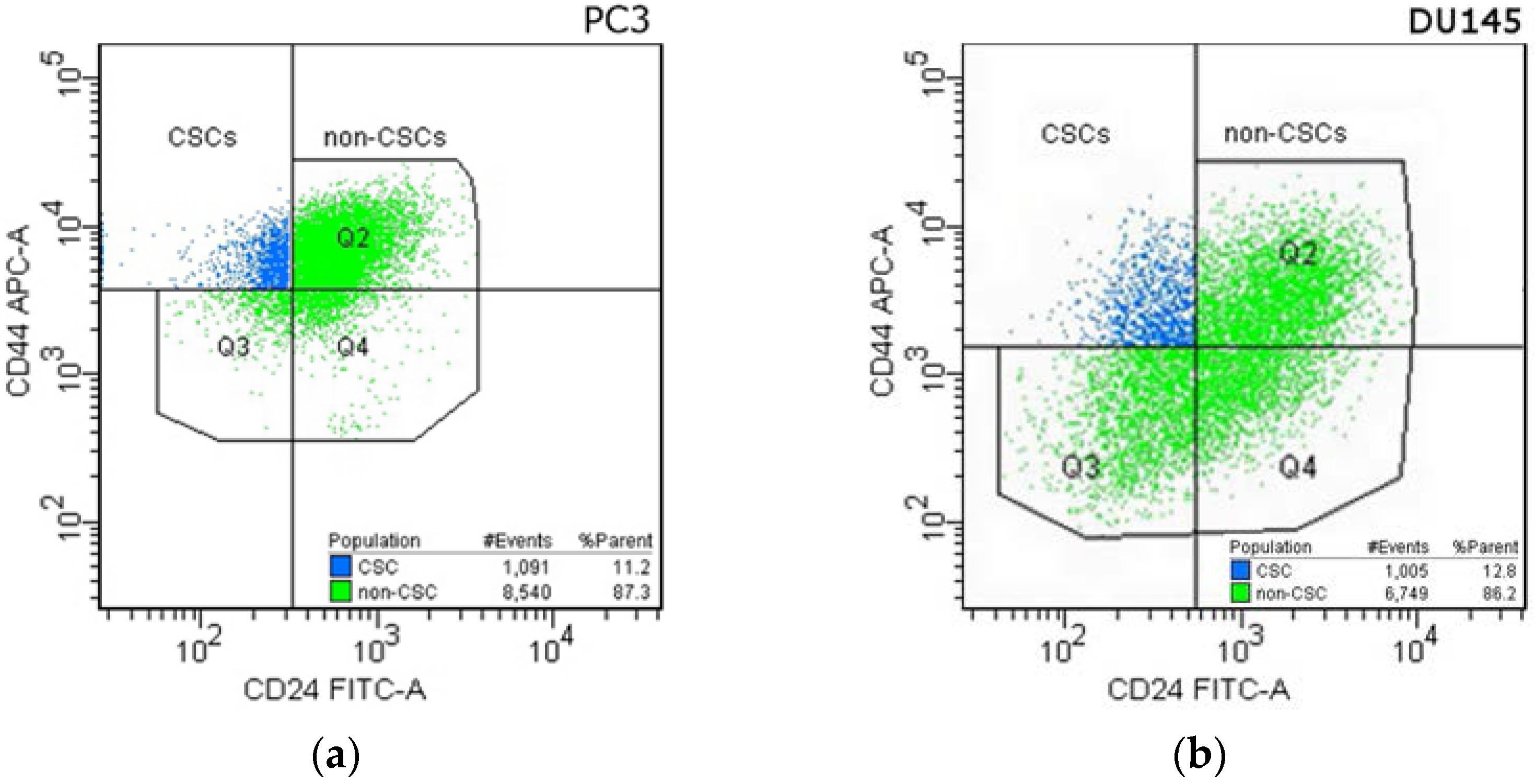
2.5. Cancer Stem Cells in DU145 and PC3 Prostate Cancer Cell Lines
2.6. Apoptotic Activities of TRAIL within the Subpopulations in DU145 and PC3 Prostate Cancer Cell Lines
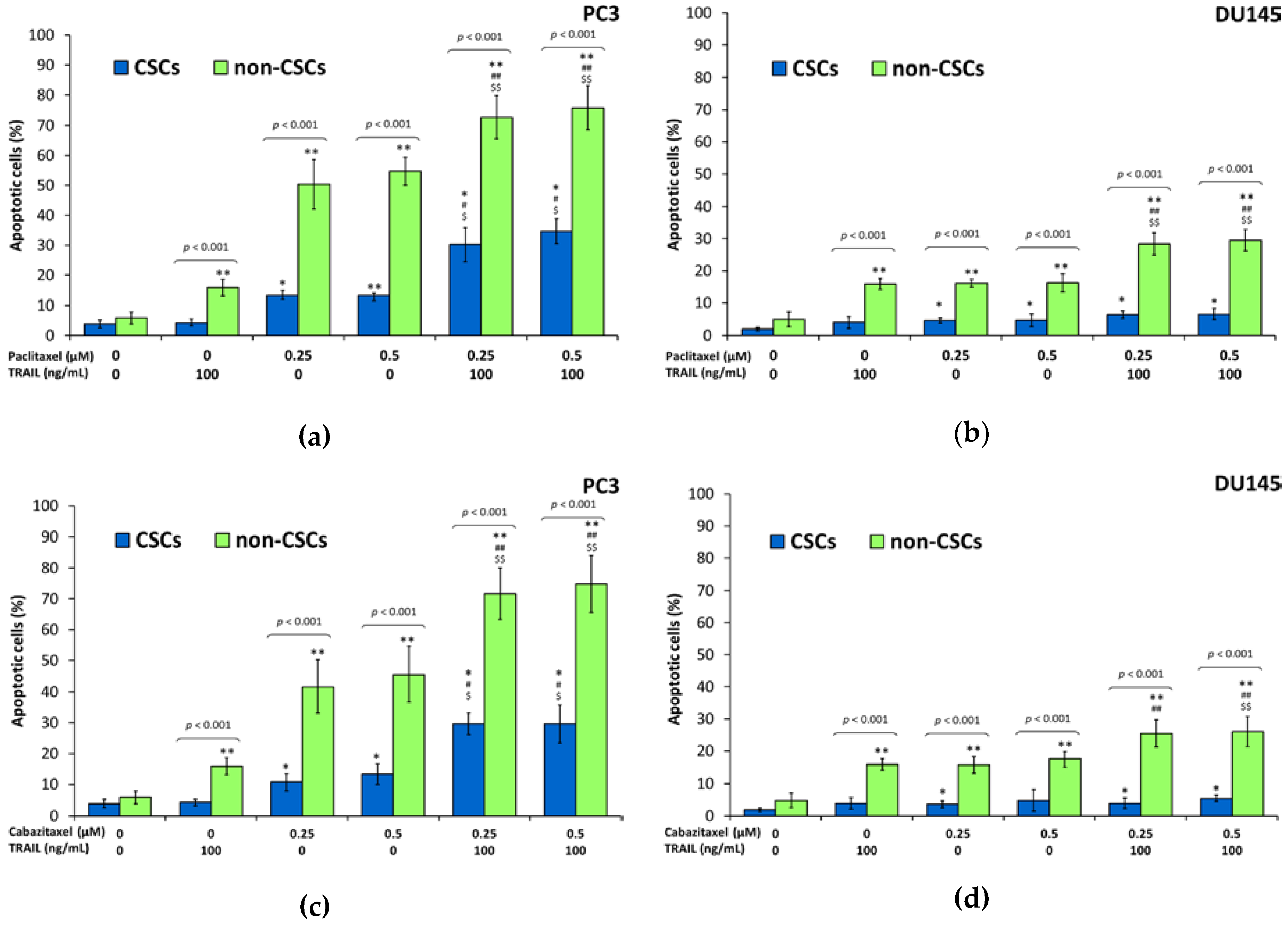
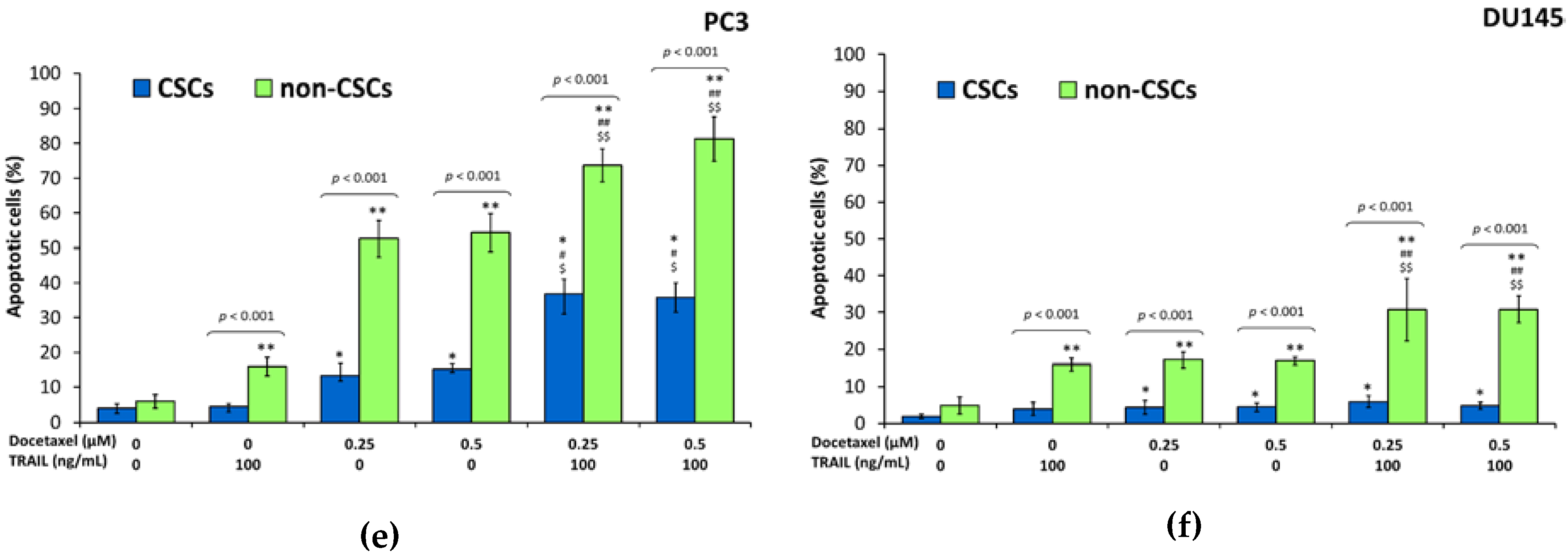
2.7. Apoptotic Activities of Paclitaxel, Cabazitaxel or Docetaxel within the Subpopulations in DU145 and PC3 Prostate Cancer Cell Lines
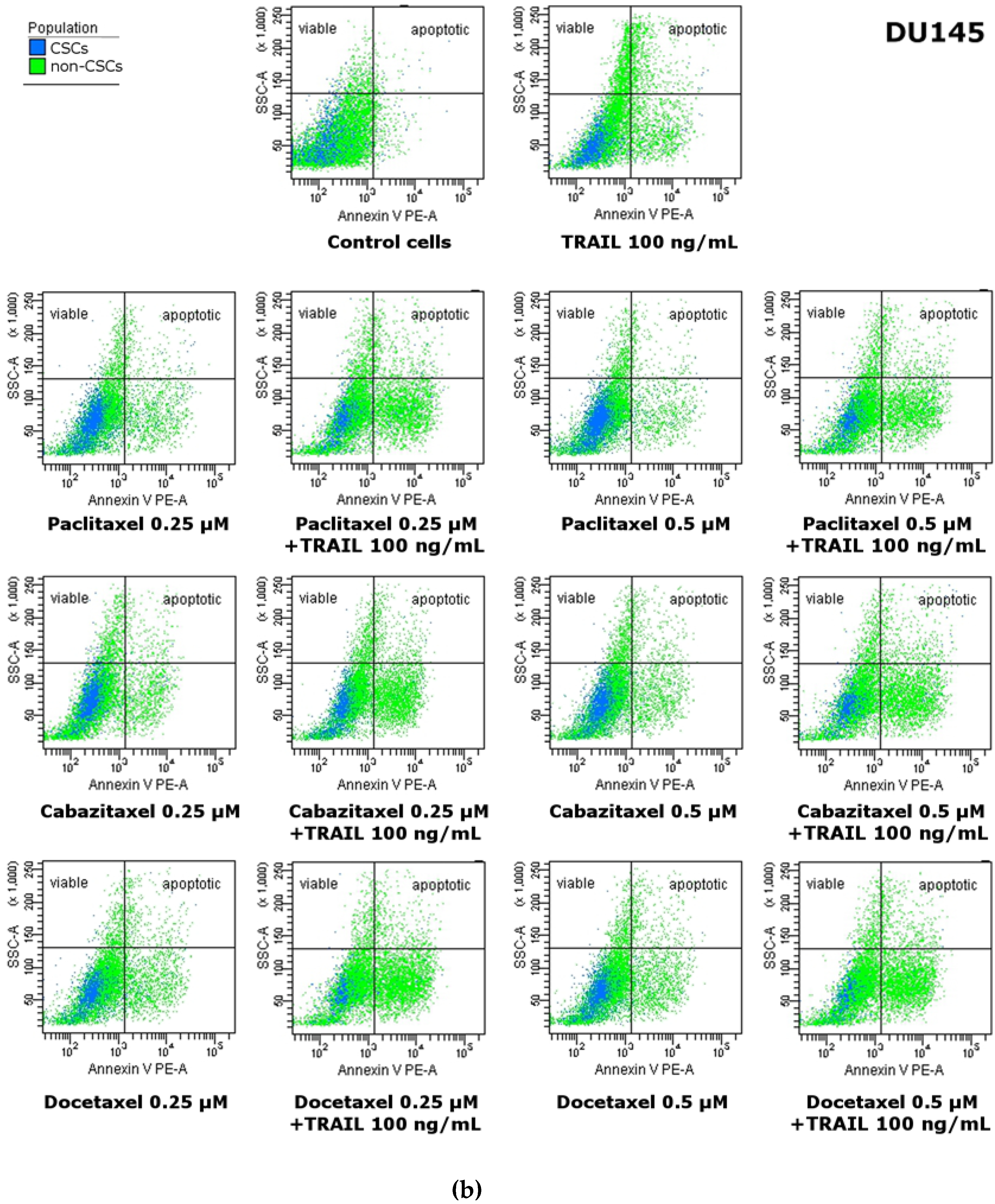
2.8. Apoptotic Activities of TRAIL in combination with Paclitaxel, Cabazitaxel or Docetaxel within the Subpopulations in DU145 and PC3 Prostate Cancer Cell Lines
2.9. CD44+/CD24− Cancer Stem Cell Number in DU145 and PC3 Cell Lines after the Treatment of TRAIL and/or Paclitaxel, Cabazitaxel, Docetaxel
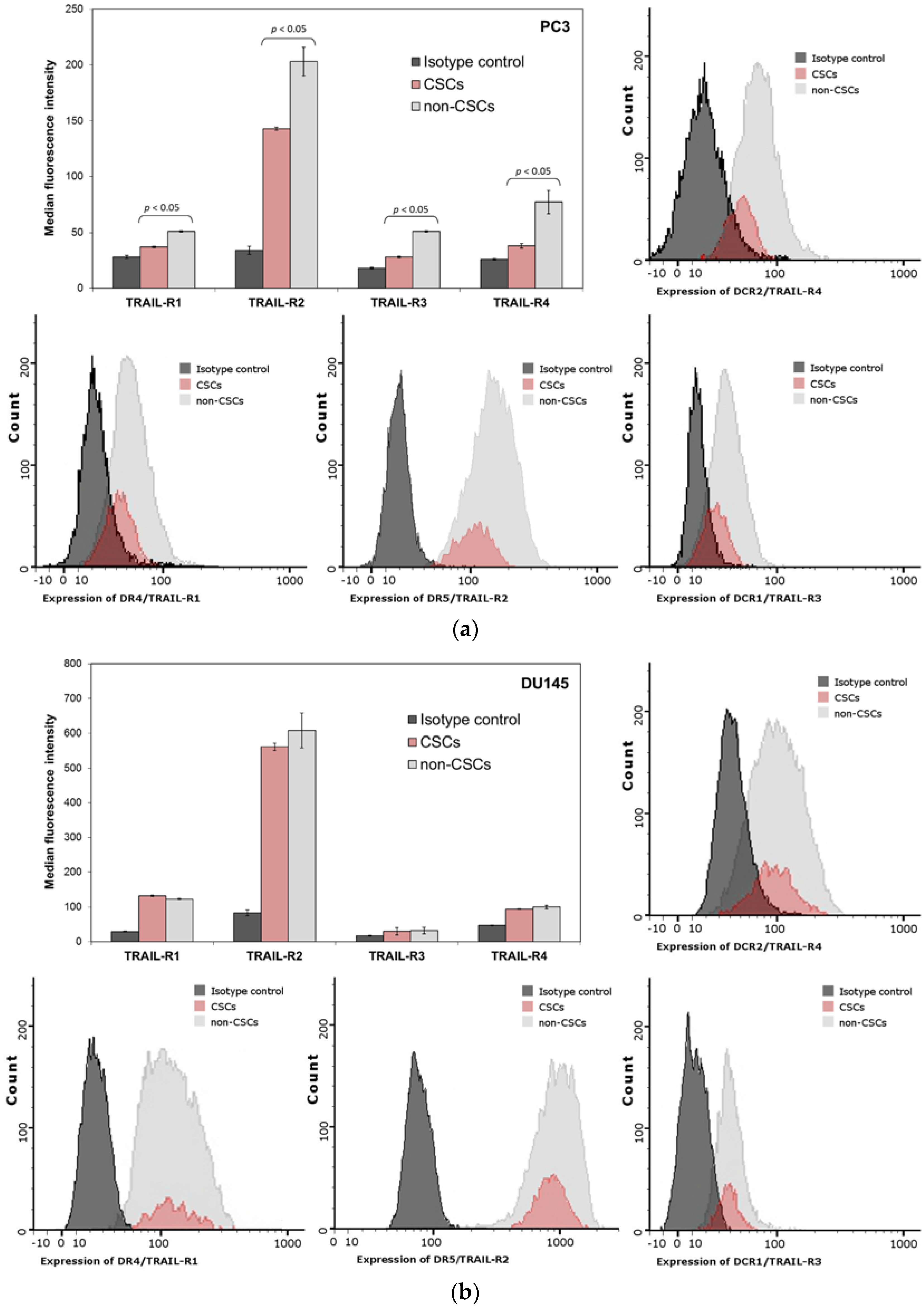
2.10. Expression of TRAIL-Receptors (R1, R2, R3 and R4) on the Surface of PC3 and DU145 Prostate Cancer Cells
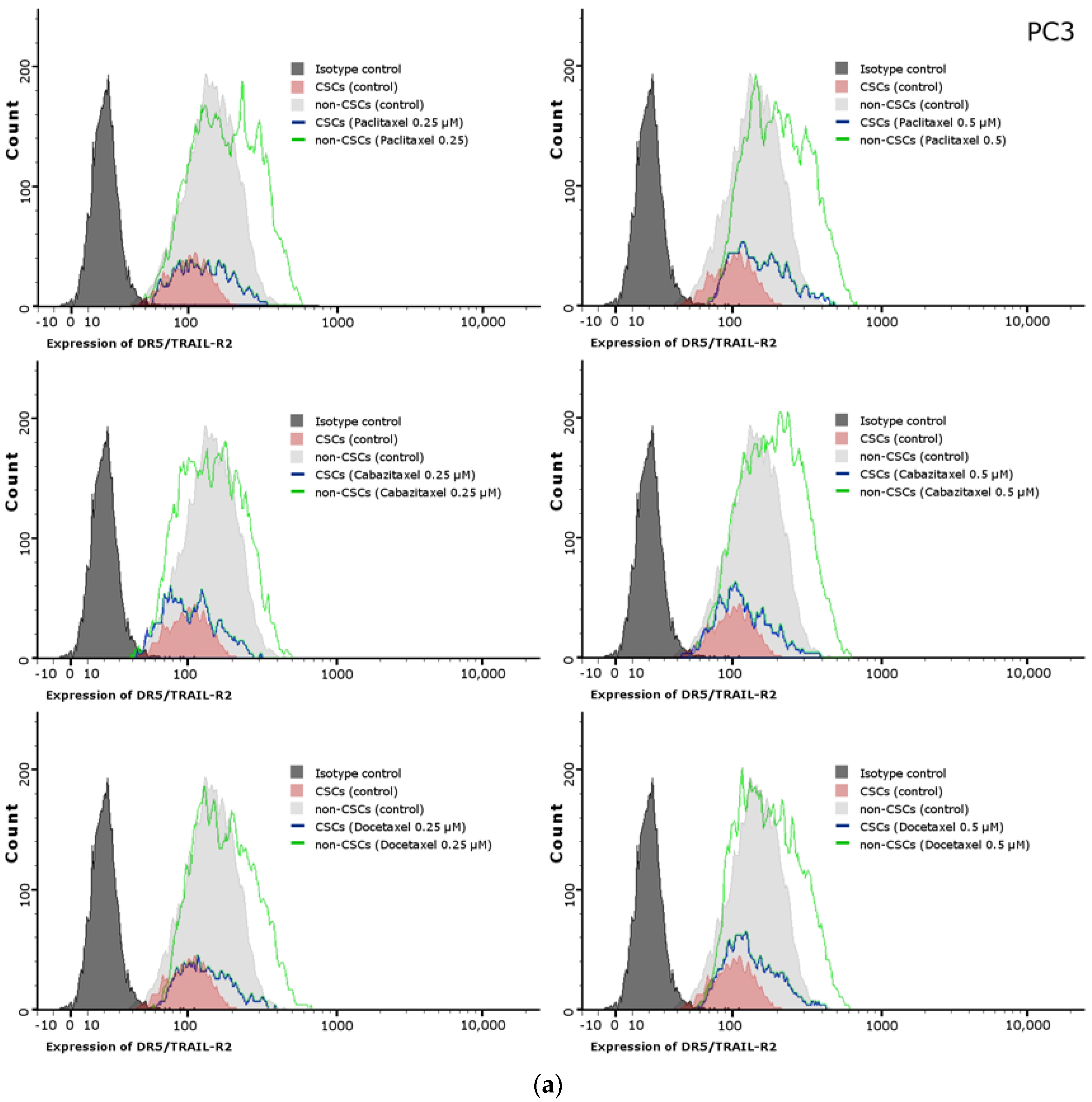
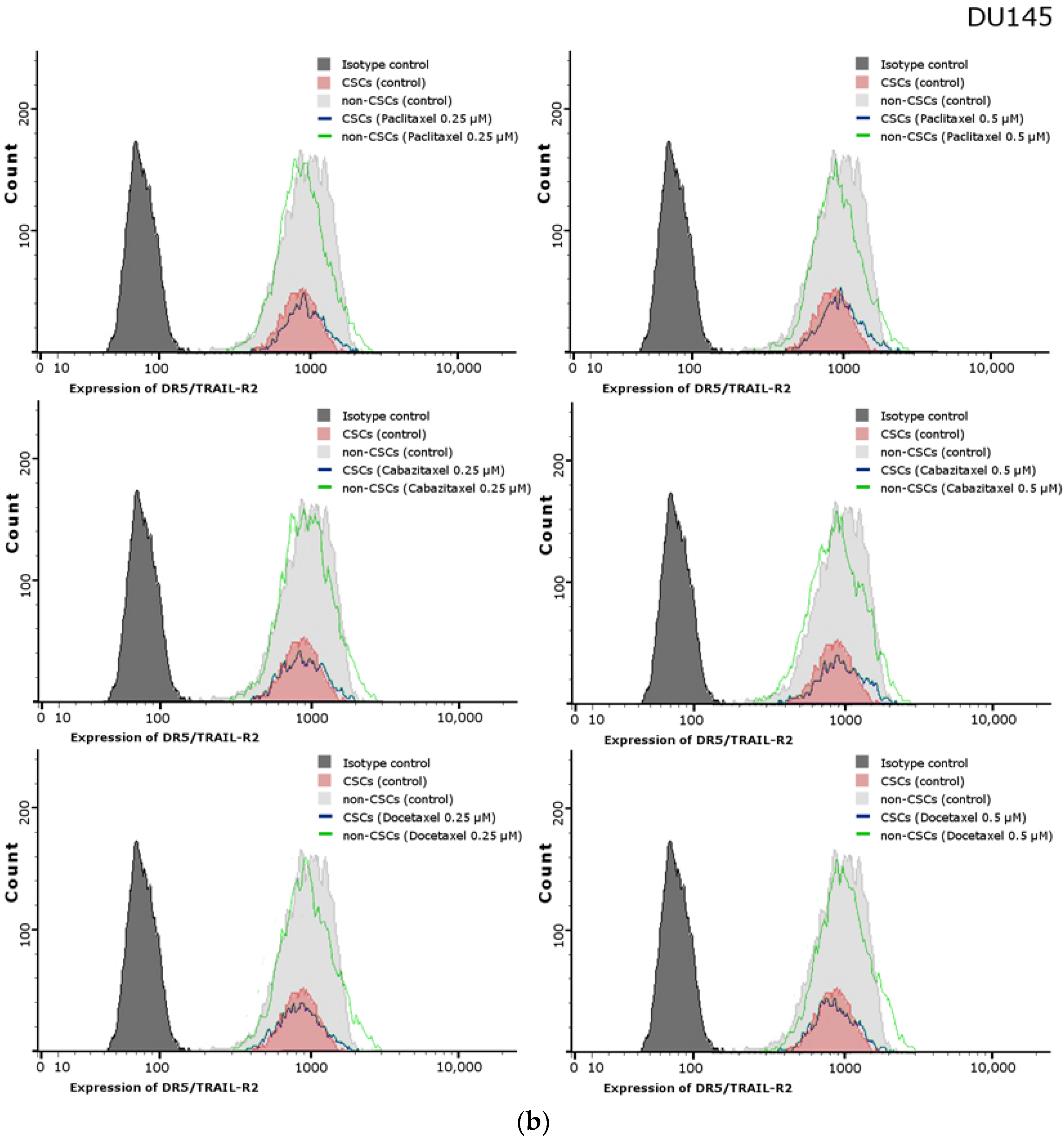
2.11. Effects of Paclitaxel, Cabazitaxel or Docetaxel on TRAIL-Receptors (R1, R2) Expression on the Surface of PC3 and DU145 Prostate Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Reagents
4.3. Detection of Apoptotic and Necrotic Cell Death by Flow Cytometry
4.4. Lactate Dehydrogenase Release Assay
4.5. Detection of CD44+/CD24− Cancer Stem Cells by Flow Cytometry
4.6. Detection of Apoptotic Cell Death by Flow Cytometry in the Subpopulations of Cells in DU145 and PC3 Prostate Cancer Cell Lines
4.7. Analysis of Death Receptor Expression on the Cancer Cell Surface by Flow Cytometry
4.8. Statistical analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CSC | cancer stem cell |
| TRAIL | tumor necrosis factor-related apoptosis-inducing ligand |
| PE | phycoerythrin |
| TRAIL-R | receptor of tumor necrosis factor-related apoptosis-inducing ligand |
| NOD/SCID | non-obese diabetic/severe combined immunodeficiency |
| TNF | tumor necrosis factor |
| DR | death receptor |
| CRPC | castration-resistant prostate cancer |
| FDA | food and drug administration |
| SD | standard deviation |
| LDH | lactate dehydrogenase |
| 7-AAD | 7-aminoactinomycin D |
| non-CSC | non-cancer stem cell |
| FITC | fluorescein isothiocyanate |
| APC | allophycocyanin |
| DCR | decoy receptor |
| ATP | adenosine triphosphate |
| AIF | apoptosis inducing factor |
| MAPK | mitogen-activated protein kinase |
| DSMZ | Deutsche Sammlung von Mikroorganismen und Zellkulturen |
| RPMI | Roswell Park Memorial Institute medium |
| ATCC | American Type Culture Collection |
| DMSO | dimethyl sulfoxide |
| EDTA | ethylenediaminetetracetic acid |
| PBS | phosphate-buffered saline |
| PerCP | peridinin chlorophyll |
| ANOVA | analysis of variance |
References
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Mokdad, A.H.; Dwyer-Lindgren, L.; Fitzmaurice, C.; Stubbs, R.W.; Bertozzi-Villa, A.; Morozoff, C.; Charara, R.; Allen, C.; Naghavi, M.; Murray, C.J. Trends and patterns of disparities in cancer mortality among US counties, 1980–2014. JAMA 2017, 317, 388–406. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.F.; Dick, J.E.; Dirks, P.B.; Eaves, C.J.; Jamieson, C.H.; Jones, D.L.; Visvader, J.; Weissman, I.L.; Wahl, G.M. Cancer stem cells—perspectives on current status and future directions: AACR workshop on cancer stem cells. Cancer Res. 2006, 66, 9339–9344. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.F.; Fuller, M. Stem cells and cancer: Two faces of eve. Cell. 2006, 124, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.T.; Berry, P.A.; Hyde, C.; Stower, M.J.; Maitland, N.J. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005, 65, 10946–10951. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Yuan, J.; Wills, M.; Kasper, S. Prostate cancer cells with stem cell characteristics reconstitute the original human tumor in vivo. Cancer Res. 2007, 67, 4807–4815. [Google Scholar] [CrossRef] [PubMed]
- Klarmann, G.J.; Hurt, E.M.; Mathews, L.A.; Zhang, X.; Duhagon, M.A.; Mistree, T.; Thomas, S.B.; Farrar, W.L. Invasive prostate cancer cells are tumor initiating cells that have a stem cell-like genomic signature. Clin. Exp. Metastasis 2009, 26, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Hurt, E.M.; Kawasaki, B.T.; Klarmann, G.J.; Thomas, S.B.; Farrar, W.L. CD44+ CD24(−) prostate cells are early cancer progenitor/stem cells that provide a model for patients with poor prognosis. Br. J. Cancer 2008, 98, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; Lawson, D.A.; Cheng, D.; Sun, W.; Garraway, I.P.; Witte, O.N. Trop2 identifies a subpopulation of murine and human prostate basal cells with stem cell characteristics. Proc. Natl. Acad. Sci. USA 2008, 105, 20882–20887. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Hindoyan, A.; Wang, S.; Tran, L.M.; Goldstein, A.S.; Lawson, D.; Chen, D.; Li, Y.; Guo, C.; Zhang, B.; et al. Identification of CD166 as a surface marker for enriching prostate stem/progenitor and cancer initiating cells. PLoS ONE 2012, 7, e42564. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.T.; Maitland, N.J. Prostate cancer stem cells. Eur. J. Cancer 2006, 42, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.D.; Robson, C.N.; Lang, S.H.; Neal, D.E.; Maitland, N.J.; Collins, A.T. CD133, a novel marker for human prostatic epithelial stem cells. J. Cell. Sci. 2004, 117, 3539–3545. [Google Scholar] [CrossRef] [PubMed]
- Patrawala, L.; Calhoun, T.; Schneider-Broussard, R.; Li, H.; Bhatia, B.; Tang, S.; Reilly, J.G.; Chandra, D.; Zhou, J.; Claypool, K.; et al. Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene 2006, 25, 1696–1708. [Google Scholar] [CrossRef] [PubMed]
- Rybak, A.P.; He, L.; Kapoor, A.; Cutz, J.C.; Tang, D. Characterization of sphere-propagating cells with stem-like properties from DU145 prostate cancer cells. Biochim. Biophys. Acta 2011, 1813, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, L.; Caporuscio, F.; Verdina, A.; Starace, G.; Crispi, S.; Nicotra, M.R.; Russo, A.; Calogero, R.A.; Morgante, E.; Natali, P.G.; et al. Cell-to-cell signaling influences the fate of prostate cancer stem cells and their potential to generate more aggressive tumors. PLoS ONE 2012, 7, e31467. [Google Scholar] [CrossRef] [PubMed]
- van Leenders, G.J.; Sookhlall, R.; Teubel, W.J.; de Ridder, C.M.; Reneman, S.; Sacchetti, A.; Vissers, K.J.; van Weerden, W.; Jenster, G. Activation of c-MET induces a stem-like phenotype in human prostate cancer. PLoS ONE 2011, 6, e26753. [Google Scholar] [CrossRef] [PubMed]
- Kangwan, N.; Park, J.M.; Kim, E.H.; Hahm, K.B. Chemoquiescence for ideal cancer treatment and prevention: Where are we now? J. Cancer Prev. 2014, 19, 89–96. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, M. TRAIL-induced signalling and apoptosis. Toxicol Lett. 2003, 139, 89–97. [Google Scholar] [CrossRef]
- Oldenhuis, C.N.; Stegehuis, J.H.; Walenkamp, A.M.; de Jong, S.; de Vries, E.G. Targeting TRAIL death receptors. Curr. Opin. Pharmacol. 2008, 8, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.; Allen, J.E.; Prabhu, V.V.; Talekar, M.K.; Finnberg, N.K.; El-Deiry, W.S. Targeting TRAIL in the treatment of cancer: New developments. Expert Opin. Ther. Targets 2015, 19, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Ni, J.; Wei, Y.F.; Yu, G.; Gentz, R.; Dixit, V.M. An antagonist decoy receptor and a death domain-containing receptor for TRAIL. Science 1997, 277, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Oldenhuis, C.N.; Szegezdi, E.; Giles, F.J.; de Vries, E.G.; de Jong, S.; Nawrocki, S.T. Targeting TRAIL towards the clinic. Curr. Drug Targets 2011, 12, 2079–2090. [Google Scholar] [CrossRef] [PubMed]
- Signore, M.; Ricci-Vitiani, L.; De Maria, R. Targeting apoptosis pathways in cancer stem cells. Cancer Lett. 2013, 332, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Seidenfeld, J.; Samson, D.J.; Hasselblad, V.; Aronson, N.; Albertsen, P.C.; Bennett, C.L.; Wilt, T.J. Single-therapy androgen suppression in men with advanced prostate cancer: A systematic review and meta-analysis. Ann. Intern. Med. 2000, 132, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Karantanos, T.; Corn, P.G.; Thompson, T.C. Prostate cancer progression after androgen deprivation therapy: Mechanisms of castrate resistance and novel therapeutic approaches. Oncogene 2013, 32, 5501–5511. [Google Scholar] [CrossRef] [PubMed]
- de Bono, J.S.; Logothetis, C.J.; Molina, A.; Fizazi, K.; North, S.; Chu, L.; Chi, K.N.; Jones, R.J.; Goodman, O.B.; Saad, F.; et al. Abiraterone and increased survival in metastatic prostate cancer. N. Engl. J. Med. 2011, 364, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Fizazi, K.; Saad, F.; Taplin, M.E.; Sternberg, C.N.; Miller, K.; de Wit, R.; Mulders, P.; Chi, K.N.; Shore, N.D.; et al. Increased survival with enzalutamide in prostate cancer after chemotherapy. N. Engl. J. Med. 2012, 367, 1187–1197. [Google Scholar] [PubMed]
- Schweizer, M.T.; Antonarakis, E.S. Abiraterone and other novel androgen-directed strategies for the treatment of prostate cancer: A new era of hormonal therapies is born. Ther. Adv. Urol. 2012, 4, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Cozzi, P.; Hao, J.; Duan, W.; Graham, P.; Kearsley, J.; Li, Y. Cancer stem cells in prostate cancer chemoresistance. Curr. Cancer Drug Targets 2014, 14, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Zhou, J.; Lin, C.J.; Hernandez, E.; Fazli, L.; Gleave, M.; Hsieh, J.T. Targeting cancer stem cells in castration-resistant prostate cancer. Clin. Cancer Res. 2016, 22, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, N.; Kawasaki, B.T.; Hurt, E.M.; Farrar, W.L. Stem cells in prostate cancer: Resolving the castrate-resistant conundrum and implications for hormonal therapy. Cancer Biol. Ther. 2006, 5, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Liu, X.; Laffin, B.; Chen, X.; Choy, G.; Jeter, C.R.; Calhoun-Davis, T.; Li, H.; Palapattu, G.S.; Pang, S.; et al. The PSA(-/lo) prostate cancer cell population harbors self-renewing long-term tumor-propagating cells that resist castration. Cell Stem Cell 2012, 10, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Vander Griend, D.J.; Karthaus, W.L.; Dalrymple, S.; Meeker, A.; DeMarzo, A.M.; Isaacs, J.T. The role of CD133 in normal human prostate stem cells and malignant cancer-initiating cells. Cancer Res. 2008, 68, 9703–9711. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Peng, J.; Zhang, Y.; Cho, W.C.; Jin, K. The implications of cancer stem cells for cancer therapy. Int. J. Mol. Sci. 2012, 13, 16636–16657. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.; Toivanen, R.; Risbridger, G.P. Stem cells in prostate cancer: Treating the root of the problem. Endocr. Relat. Cancer 2010, 17, R273–R285. [Google Scholar] [CrossRef] [PubMed]
- Signoretti, S.; Loda, M. Prostate stem cells: From development to cancer. Semin. Cancer Biol. 2007, 17, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Maitland, N.J.; Bryce, S.D.; Stower, M.J.; Collins, A.T. Prostate cancer stem cells: A target for new therapies. Ernst. Schering Found. Symp. Proc. 2006, 155–179. [Google Scholar]
- Botchkina, G.I.; Zuniga, E.S.; Rowehl, R.H.; Park, R.; Bhalla, R.; Bialkowska, A.B.; Johnson, F.; Golub, L.M.; Zhang, Y.; Ojima, I.; et al. Prostate cancer stem cell-targeted efficacy of a new-generation taxoid, SBT-1214 and novel polyenolic zinc-binding curcuminoid, CMC2.24. PLoS ONE 2013, 8, e69884. [Google Scholar] [CrossRef] [PubMed]
- Maitland, N.J.; Collins, A.T. Prostate cancer stem cells: A new target for therapy. J. Clin. Oncol. 2008, 26, 2862–2870. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, J.Y.; Wu, G.S. Tumor necrosis factor-related apoptosis-inducing ligand is required for tumor necrosis factor α-mediated sensitization of human breast cancer cells to chemotherapy. Cancer Res. 2006, 66, 10092–10099. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, J.Y.; Tainsky, M.A.; Wu, G.S. Evidence that tumor necrosis factor-related apoptosis-inducing ligand induction by 5-aza-2’-deoxycytidine sensitizes human breast cancer cells to adriamycin. Cancer Res. 2007, 67, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Daigeler, A.; Chromik, A.M.; Geisler, A.; Bulut, D.; Hilgert, C.; Krieg, A.; Klein-Hitpass, L.; Lehnhardt, M.; Uhl, W.; Mittelkötter, U. Synergistic apoptotic effects of taurolidine and TRAIL on squamous carcinoma cells of the esophagus. Int. J. Oncol. 2008, 32, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; El-Deiry, W.S. Requirement of p53 targets in chemosensitization of colonic carcinoma to death ligand therapy. Proc. Natl. Acad. Sci. USA 2003, 100, 15095–15100. [Google Scholar] [CrossRef] [PubMed]
- Baritaki, S.; Huerta-Yepez, S.; Sakai, T.; Spandidos, D.A.; Bonavida, B. Chemotherapeutic drugs sensitize cancer cells to TRAIL-mediated apoptosis: Up-regulation of DR5 and inhibition of yin yang 1. Mol. Cancer Ther. 2007, 6, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.X.; Jin, X.H.; Zeng, Y.; El Hamed, A.M.; Kakehi, Y. Low concentrations of doxorubicin sensitizes human solid cancer cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-receptor (R) 2-mediated apoptosis by inducing TRAIL-R2 expression. Cancer Sci. 2007, 98, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Hunter, T.B.; Manimala, N.J.; Luddy, K.A.; Catlin, T.; Antonia, S.J. Paclitaxel and TRAIL synergize to kill paclitaxel-resistant small cell lung cancer cells through a caspase-independent mechanism mediated through AIF. Anticancer Res. 2011, 31, 3193–3204. [Google Scholar] [PubMed]
- Li, L.; Wen, X.Z.; Bu, Z.D.; Cheng, X.J.; Xing, X.F.; Wang, X.H.; Zhang, L.H.; Guo, T.; Du, H.; Hu, Y.; et al. Paclitaxel enhances tumoricidal potential of TRAIL via inhibition of MAPK in resistant gastric cancer cells. Oncol. Rep. 2016, 35, 3009–3017. [Google Scholar] [CrossRef] [PubMed]
- Nimmanapalli, R.; Perkins, C.L.; Orlando, M.; O’Bryan, E.; Nguyen, D.; Bhalla, K.N. Pretreatment with paclitaxel enhances apo-2 ligand/tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis of prostate cancer cells by inducing death receptors 4 and 5 protein levels. Cancer Res. 2001, 61, 759–763. [Google Scholar] [PubMed]
- Yoo, J.; Park, S.S.; Lee, Y.J. Pretreatment of docetaxel enhances TRAIL-mediated apoptosis in prostate cancer cells. J. Cell. Biochem. 2008, 104, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Hu, H.; Tan, H.; Chow, L.W.; Yip, A.Y.; Loo, W.T. Relationship of CD44+CD24−/low breast cancer stem cells and axillary lymph node metastasis. J. Transl. Med. 2012, 10, S6. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C.; Kishimoto, H.; Fuchs, R.K.; Mehrotra, S.; Bhat-Nakshatri, P.; Turner, C.H.; Goulet, R.; Badve, S.; Nakshatri, H. CD44+/CD24− breast cancer cells exhibit enhanced invasive properties: An early step necessary for metastasis. Breast Cancer Res. 2006, 8, R59. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xu, F.; Du, X.; Lai, D.; Zhao, Y.; Huang, Q.; Jiang, L.; Huang, W.; Cheng, W.; Liu, Z. Establishment and characterization of multi-drug resistant, prostate carcinoma-initiating stem-like cells from human prostate cancer cell lines 22RV1. Mol. Cell. Biochem. 2010, 340, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiao, M.; Li, L.; Wu, D.; Wu, K.; Li, X.; Zhu, G.; Dang, Q.; Wang, X.; Hsieh, J.T.; et al. Tumorspheres derived from prostate cancer cells possess chemoresistant and cancer stem cell properties. J. Cancer Res. Clin. Oncol. 2012, 138, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Iczkowski, K.A. Cell adhesion molecule CD44: Its functional roles in prostate cancer. Am. J. Transl. Res. 2010, 3, 1–7. [Google Scholar] [PubMed]
- Sastry, K.S.; Al-Muftah, M.A.; Li, P.; Al-Kowari, M.K.; Wang, E.; Ismail Chouchane, A.; Kizhakayil, D.; Kulik, G.; Marincola, F.M.; Haoudi, A.; et al. Targeting proapoptotic protein BAD inhibits survival and self-renewal of cancer stem cells. Cell Death Differ. 2014, 21, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Mastelić, A.; Čikeš Čulić, V.; Režić Mužinić, N.; Vuica-Ross, M.; Barker, D.; Leung, E.Y.; Reynisson, J.; Markotić, A. Glycophenotype of breast and prostate cancer stem cells treated with thieno[2,3-b]pyridine anticancer compound. Drug Des. Devel. Ther. 2017, 11, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, A.; Kumar, S.; Arfuso, F.; Newsholme, P.; Dharmarajan, A. Secreted frizzled-related protein 4 (sFRP4) chemo-sensitizes cancer stem cells derived from human breast, prostate, and ovary tumor cell lines. Sci. Rep. 2017, 7, 2256. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Henning, S.M.; Heber, D.; Vadgama, J.V. Sensitization to docetaxel in prostate cancer cells by green tea and quercetin. J. Nutr. Biochem. 2015, 26, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Berndtsson, M.; Konishi, Y.; Bonni, A.; Hägg, M.; Shoshan, M.; Linder, S.; Havelka, A.M. Phosphorylation of BAD at ser-128 during mitosis and paclitaxel-induced apoptosis. FEBS Lett. 2005, 579, 3090–3094. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.Y.; Jiang, C.C.; Croft, A.; Tay, K.H.; Thorne, R.F.; Yang, F.; Liu, H.; Hersey, P.; Zhang, X.D. Contrasting effects of nutlin-3 on TRAIL- and docetaxel-induced apoptosis due to upregulation of TRAIL-R2 and Mcl-1 in human melanoma cells. Mol. Cancer Ther. 2010, 9, 3363–3374. [Google Scholar] [CrossRef] [PubMed]











© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaworska, D.; Szliszka, E. Targeting Apoptotic Activity Against Prostate Cancer Stem Cells. Int. J. Mol. Sci. 2017, 18, 1648. https://doi.org/10.3390/ijms18081648
Jaworska D, Szliszka E. Targeting Apoptotic Activity Against Prostate Cancer Stem Cells. International Journal of Molecular Sciences. 2017; 18(8):1648. https://doi.org/10.3390/ijms18081648
Chicago/Turabian StyleJaworska, Dagmara, and Ewelina Szliszka. 2017. "Targeting Apoptotic Activity Against Prostate Cancer Stem Cells" International Journal of Molecular Sciences 18, no. 8: 1648. https://doi.org/10.3390/ijms18081648
APA StyleJaworska, D., & Szliszka, E. (2017). Targeting Apoptotic Activity Against Prostate Cancer Stem Cells. International Journal of Molecular Sciences, 18(8), 1648. https://doi.org/10.3390/ijms18081648