An Appropriate Modulation of Lymphoproliferative Response and Cytokine Release as Possible Contributors to Longevity


Abstract
:
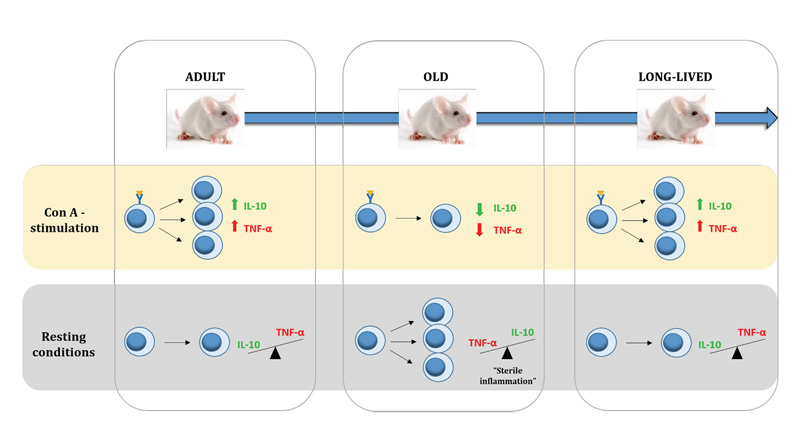{kind=link}
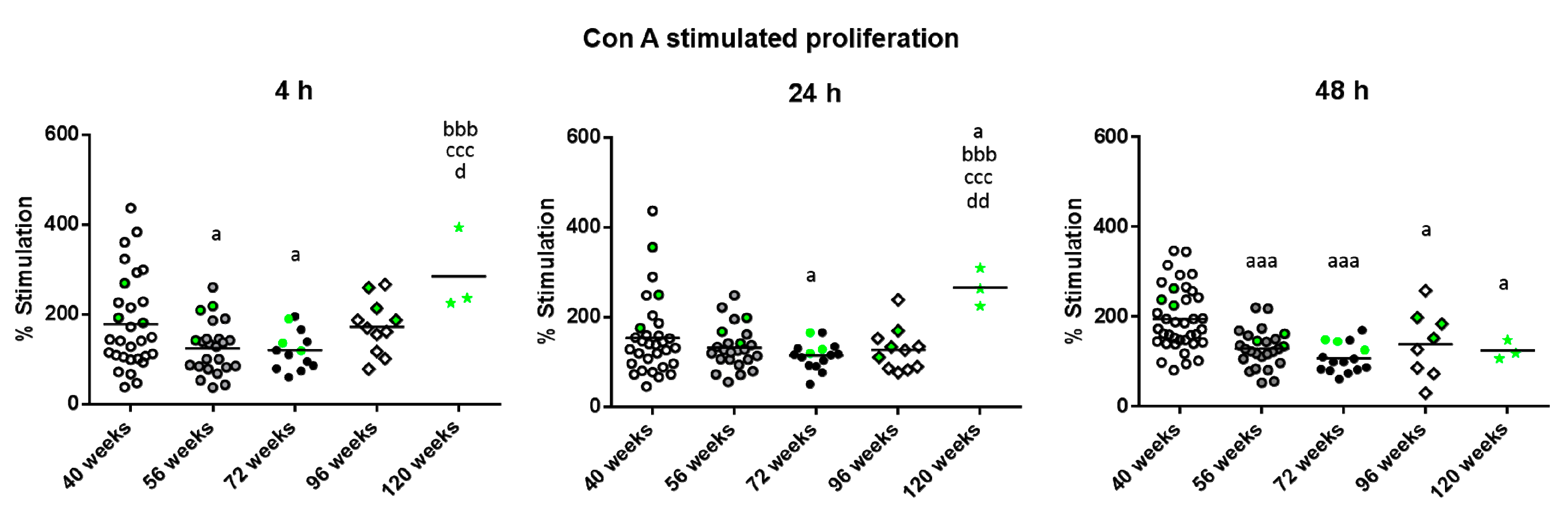{kind=link}
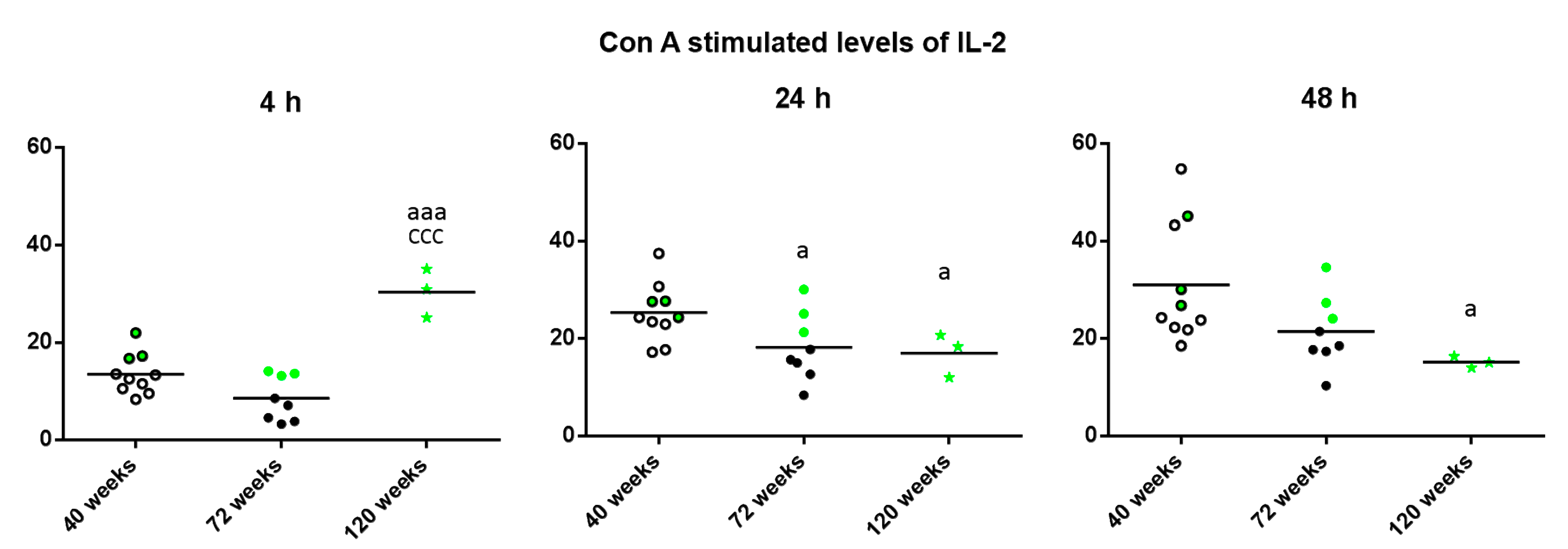{kind=link}
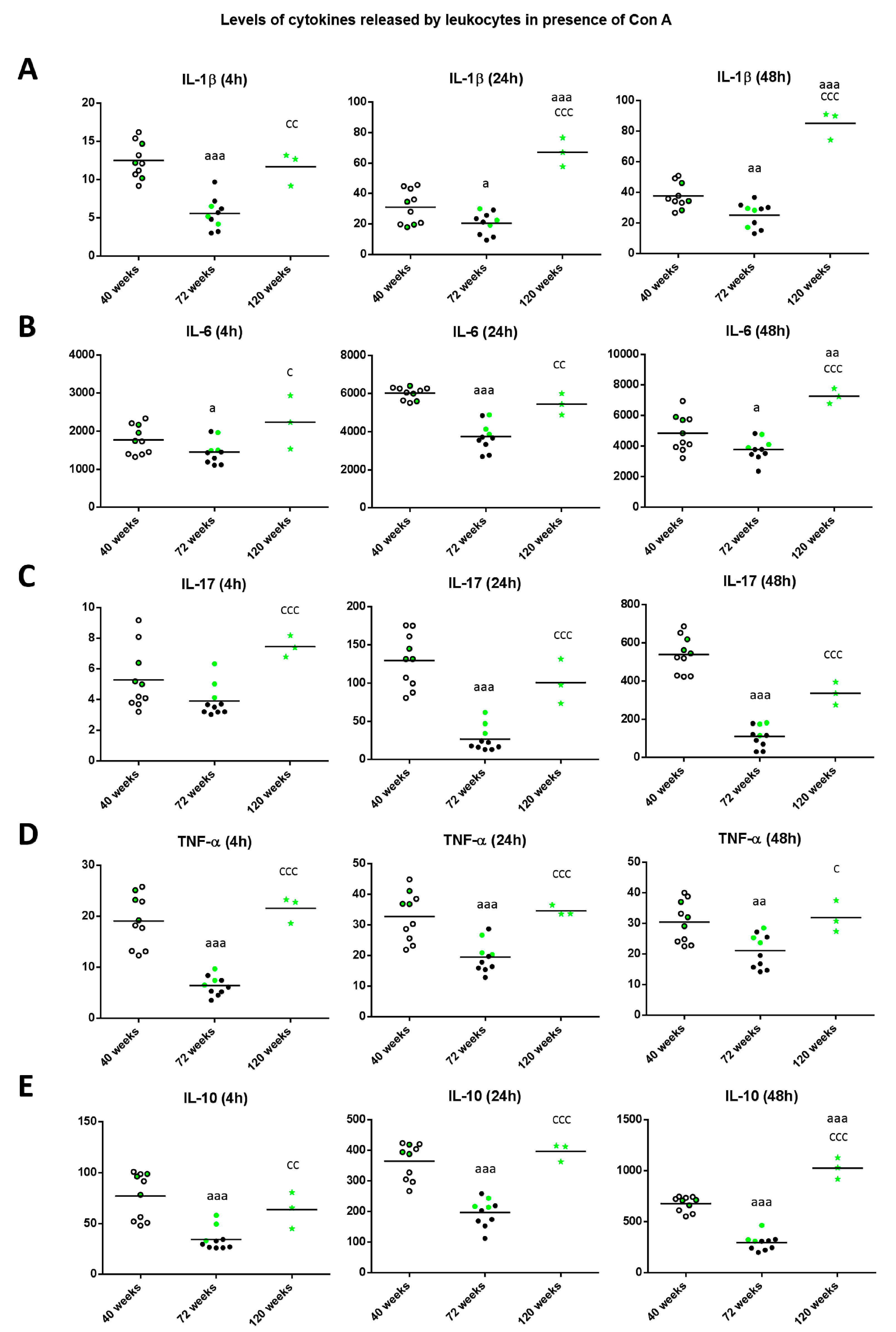{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
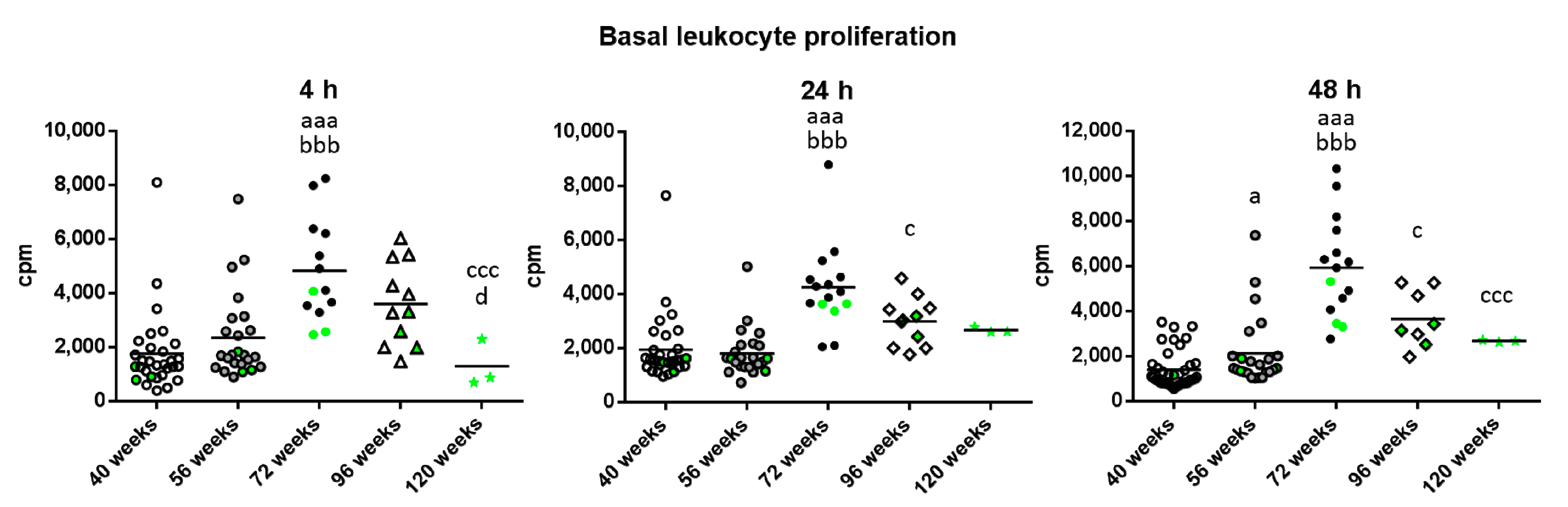
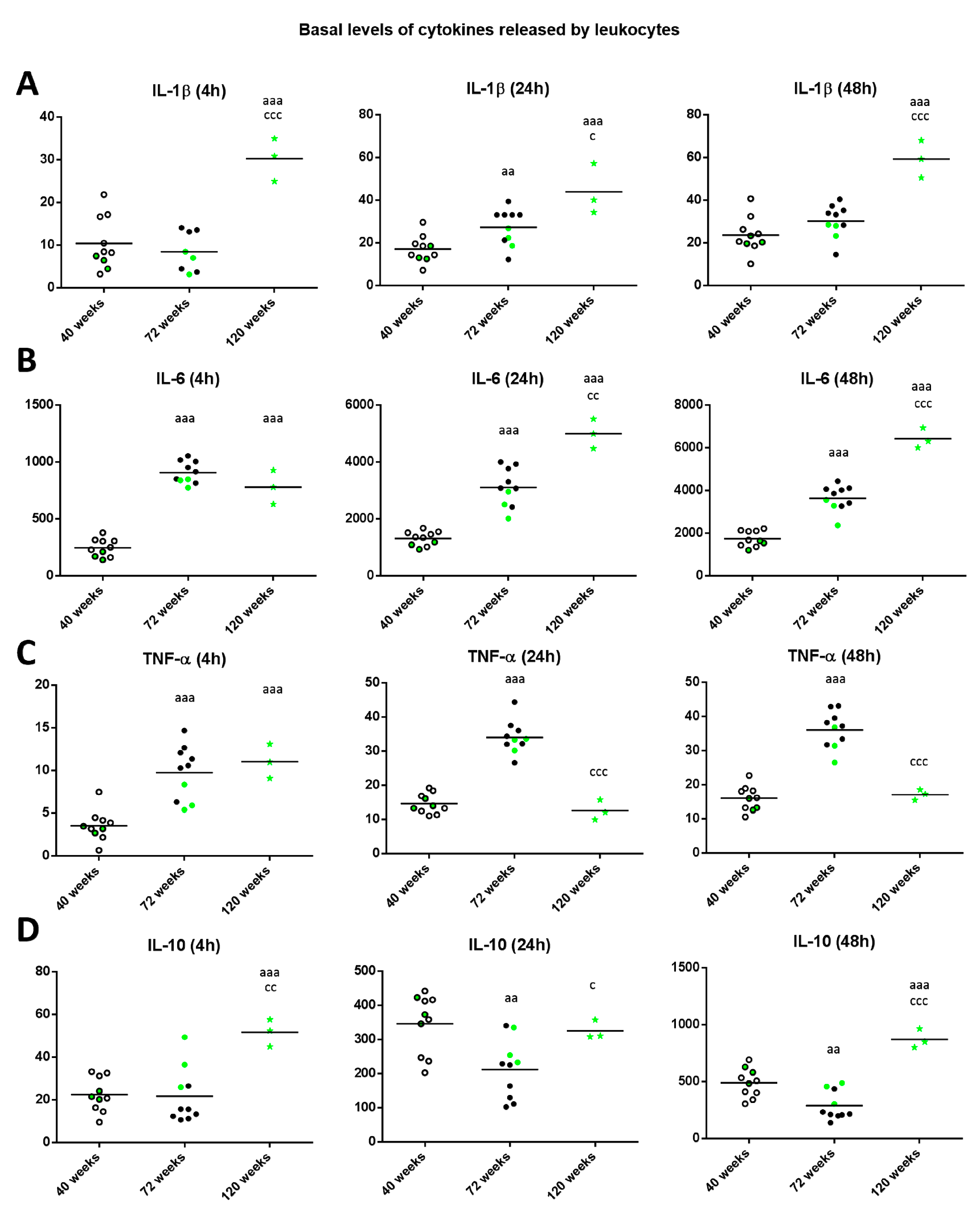
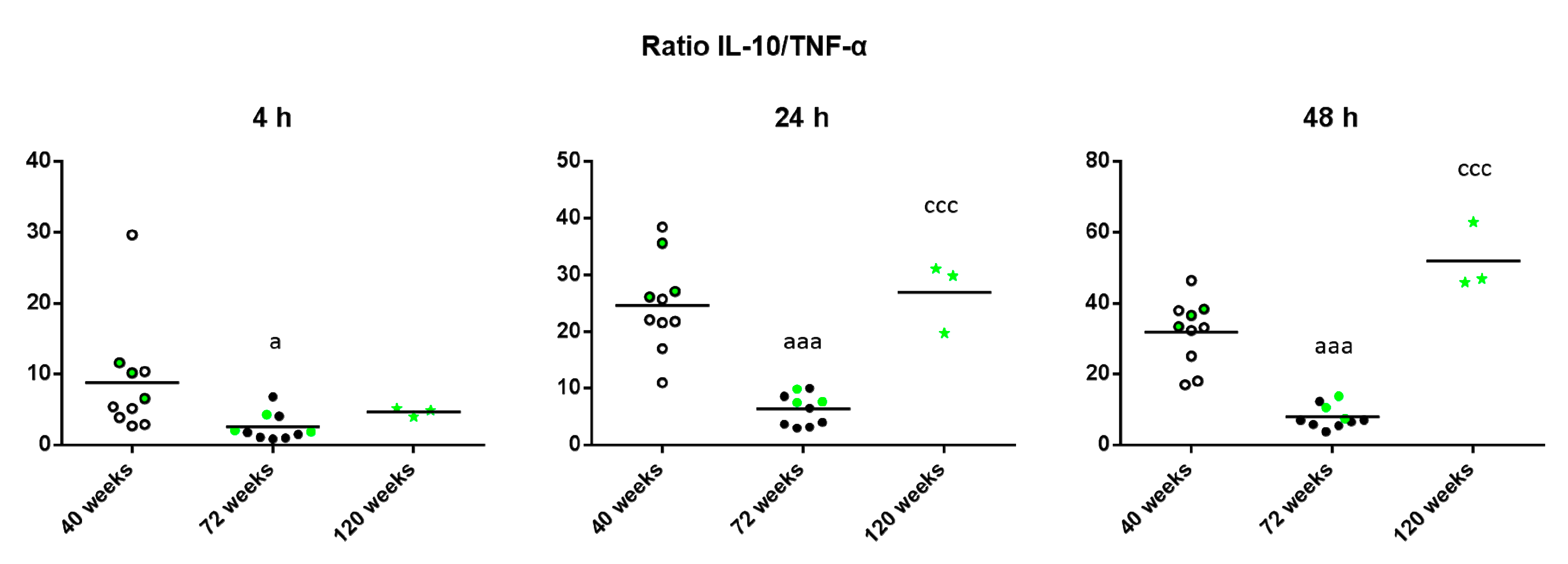
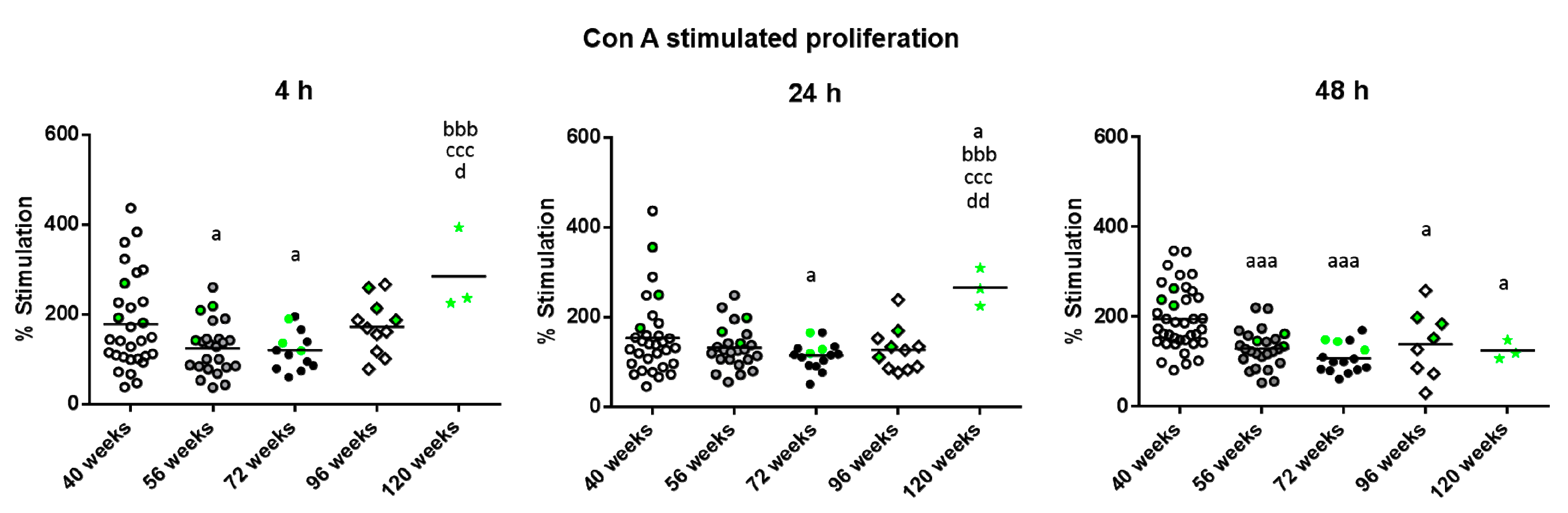
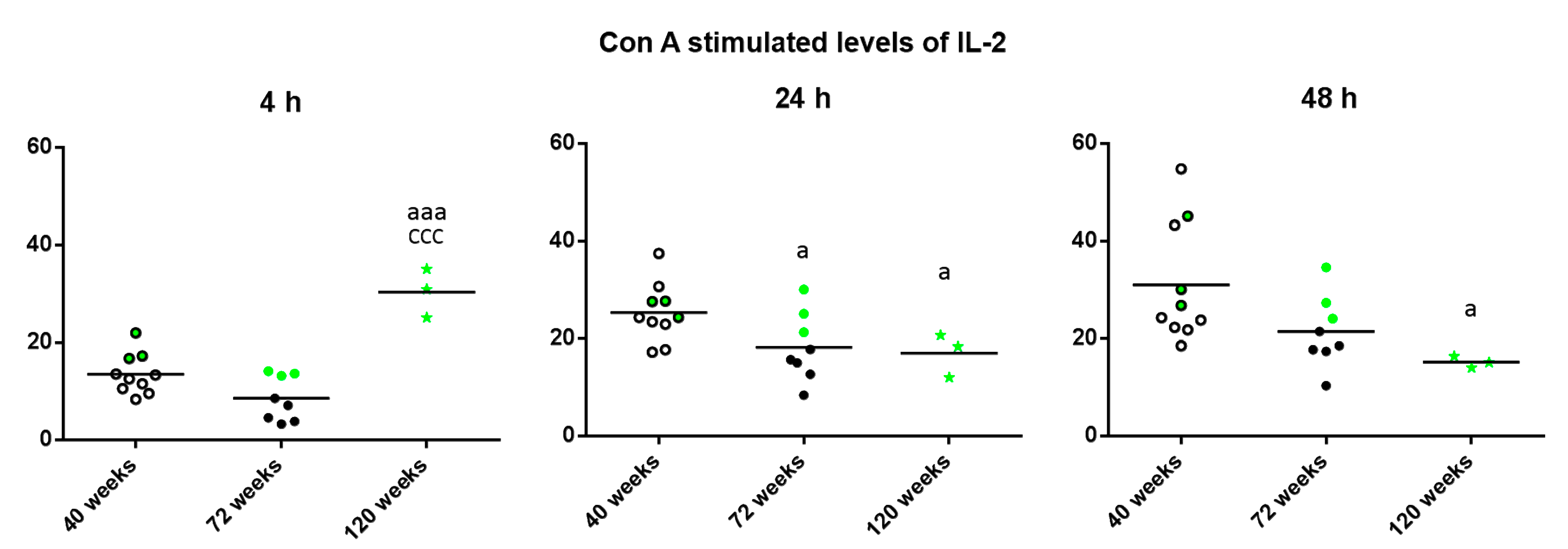
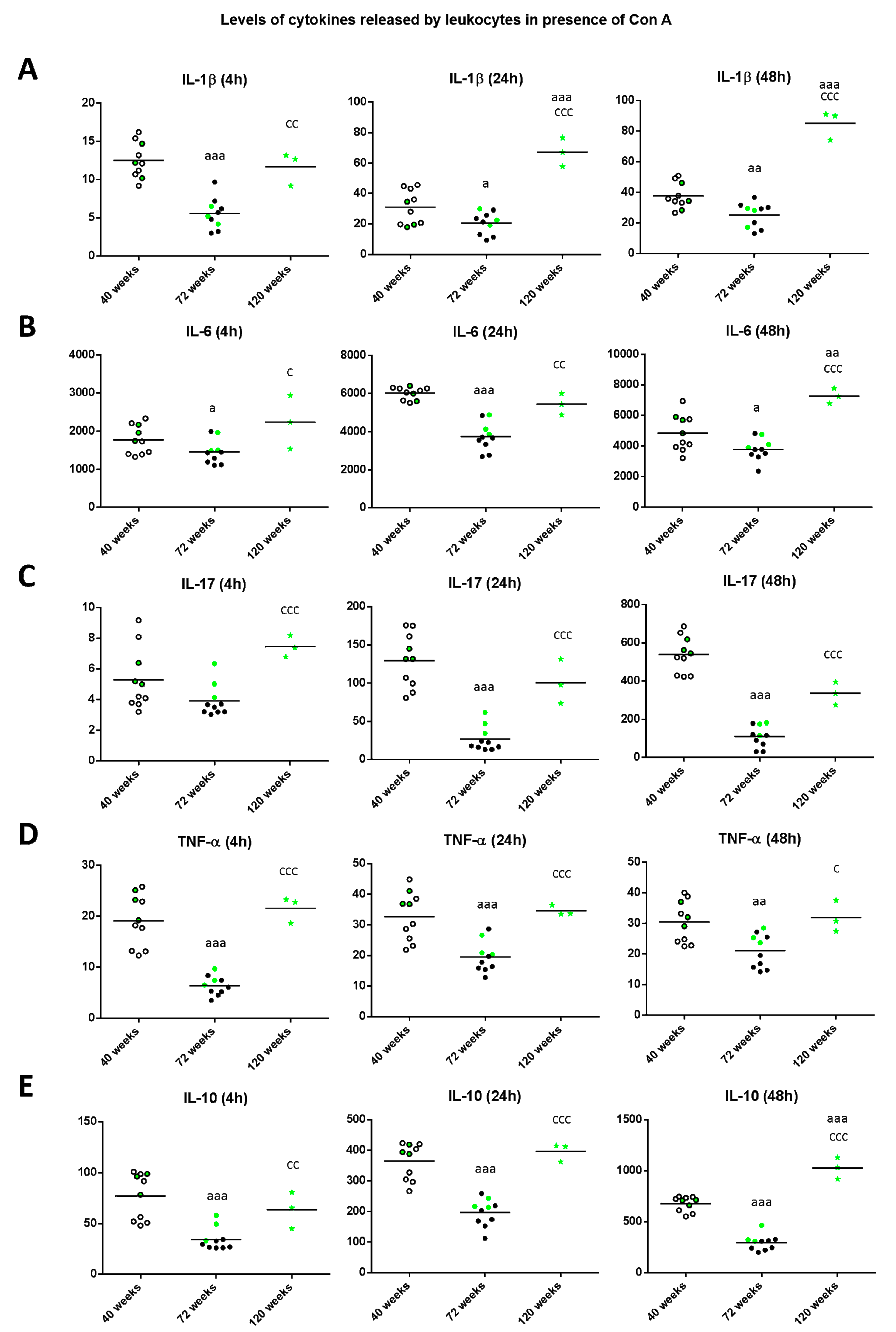
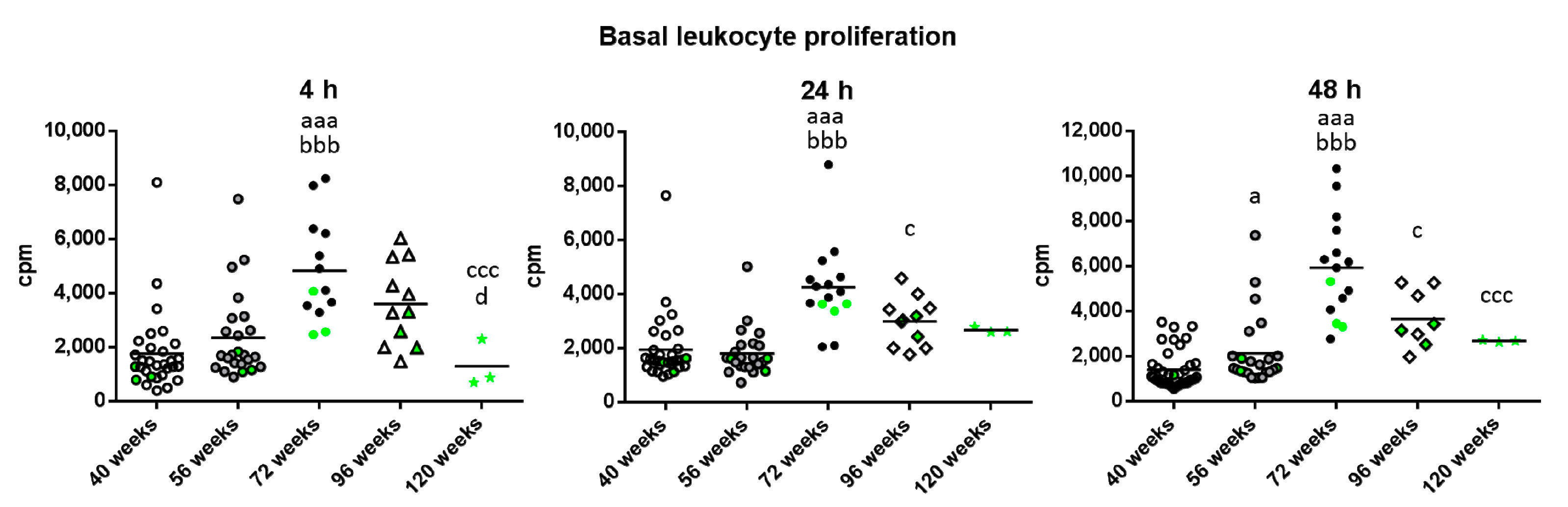
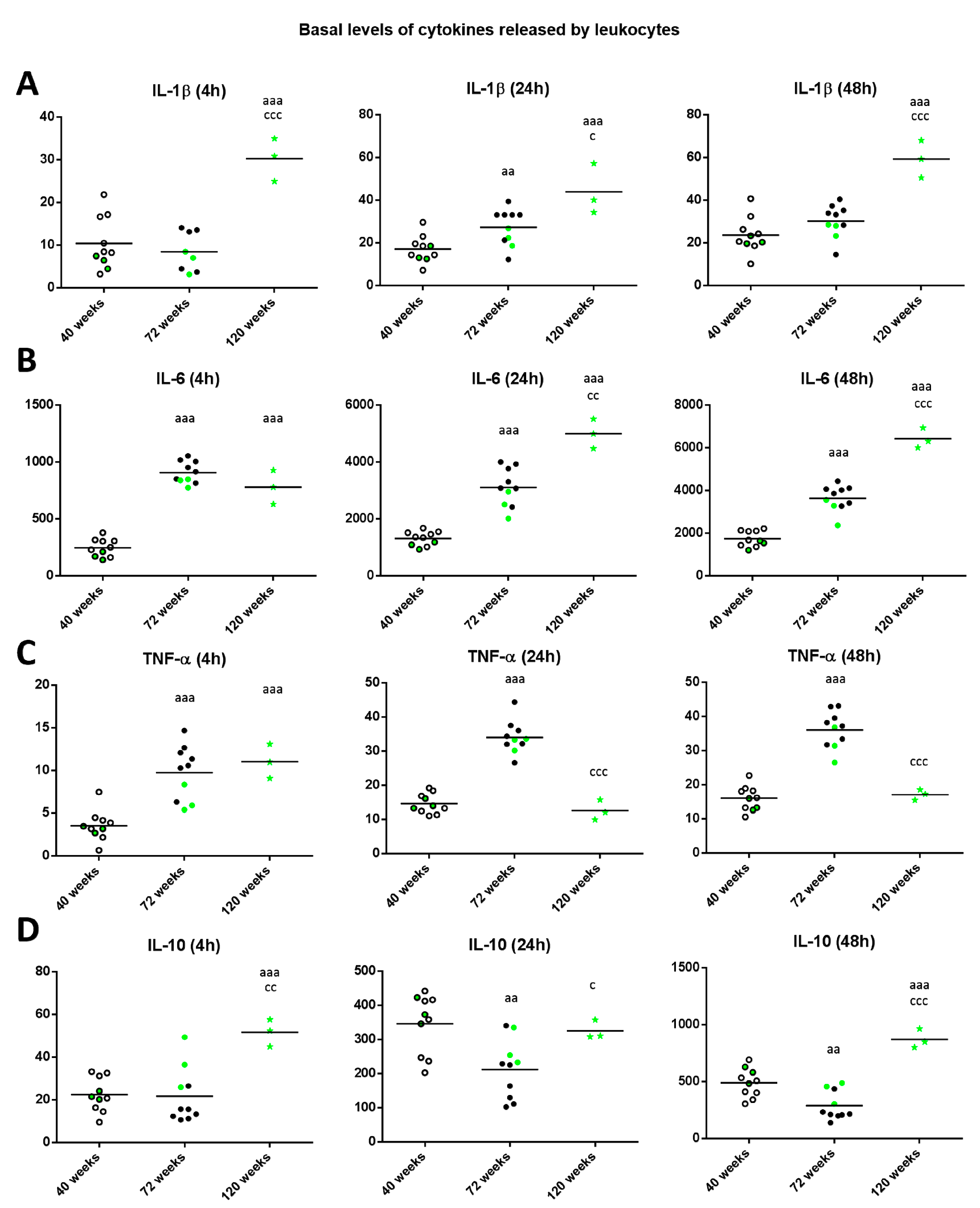
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Collection of Peritoneal Leukocytes
4.3. Lymphoproliferation
4.4. Cytokine Measurement
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AICD | Activation-induced cell death |
| Con A | Concanavalin A |
| DAMPs | Damage associated molecular patterns |
| LPS | Lipopolysaccharide |
References
- Fulop, T.; Larbi, A.; Kotb, R.; de Angelis, F.; Pawelec, G. Aging, immunity, and cancer. Discov. Med. 2011, 11, 537–550. [Google Scholar] [PubMed]
- Dewan, S.K.; Zheng, S.B.; Xia, S.J.; Bill, K. Senescent remodeling of the immune system and its contribution to the predisposition of the elderly to infections. Chin. Med. J. 2012, 125, 3325–3331. [Google Scholar] [PubMed]
- De la Fuente, M.; Miquel, J. An update of the oxidation-inflammation theory of aging: The involvement of the immune system in oxi-inflamm-aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E.; De la Fuente, M. The role of oxidative and inflammatory stress and persistent viral infections in immunosenescence. Mech. Ageing Dev. 2016, 158, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Wayne, S.J.; Rhyne, R.L.; Garry, P.J.; Goodwin, J.S. Cell-mediated immunity as a predictor of morbidity and mortality in subjects over 60. J. Gerontol. 1990, 114, 80–88. [Google Scholar] [CrossRef]
- Ferguson, F.G.; Wikby, A.; Maxson, P.; Olsson, J.; Johansson, B. Immune parameters in a longitudinal study of a very old population of Swedish people: A comparison between survivors and nonsurvivors. J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, B378–B382. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Franceschi, C.; Monti, D.; Ginaldi, L. Inflammation markers predicting frailty and mortality in the elderly. Exp. Mol. Pathol. 2006, 80, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Guayerbas, N.; Puerto, M.; Víctor, V.M.; Miquel, J.; De la Fuente, M. Leukocyte function and life span in a murine model of premature immunosenescence. Exp. Gerontol. 2002, 37, 249–256. [Google Scholar] [CrossRef]
- Guayerbas, N.; De la Fuente, M. An impairment of phagocytic function is linked to a shorter life span in two strains of prematurely aging mice. Dev. Comp. Immunol. 2003, 27, 339–350. [Google Scholar] [CrossRef]
- Franceschi, C.; Monti, D.; Sansoni, P.; Cossarizza, A. The immunology of exceptional individuals: The lesson of centenarians. Immunol. Today 1995, 16, 12–16. [Google Scholar] [CrossRef]
- Puerto, M.; Guayerbas, G.; Álvarez, P.; De la Fuente, M. Modulation of neuropeptide Y and norepinephrine on several leucocyte functions in adult, old and very old mice. J. Neuroimmunol. 2005, 165, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Fernández, P.; Puerto, M.; Maté, I.; Ribera, J.M.; De la Fuente, M. The neutrophils of centenarians show function levels similar to those of adults. J. Am. Geriatr. Soc. 2008, 56, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Caamaño, J.; Lord, J.M.; De la Fuente, M. Preserved immune functions and controlled leukocyte oxidative stress in naturally long-lived mice: Possible role of nuclear factor-κB. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 941–950. [Google Scholar] [CrossRef] [PubMed]
- De Toda, I.M.; Maté, I.; Vida, C.; Cruces, J.; De la Fuente, M. Immune function parameters as markers of biological age and predictors of longevity. Aging 2016, 8, 3110–3119. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, R.; Andrew, D. Thymic involution in aging. J. Clin. Immunol. 2000, 20, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Shanley, D.P.; Aw, D.; Manley, N.R.; Palmer, D.B. An evolutionary perspective on the mechanisms of immunosenescence. Trends Immunol. 2009, 30, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Solana, R.; Villanueva, J.L.; Pena, J.; De la Fuente, M. Cell mediated immunity in aging. Comp. Biochem. Physiol. 1991, 99, 1–4. [Google Scholar] [CrossRef]
- Pawelec, G.; Adibzadeh, M.; Solana, R.; Beckman, I. The T-cell in the aging individual. Mech. Aging Dev. 1997, 93, 35–45. [Google Scholar] [CrossRef]
- Hallgren, H.M.; Bergh, N.; Rodysill, K.J.; O’Leary, J.J. Lymphocyte proliferative response to PHA and anti-CD3/Ti monoclonal antibodies T cell surface marker expression, and serum IL-2 receptor levels as biomarkers of age and health. Mech. Aging Dev. 1998, 43, 175–185. [Google Scholar] [CrossRef]
- Wikby, A.; Ferguson, F.; Forsey, R.; Thompson, J.; Strindhall, J.; Lofgren, S.; Nilsson, B.O.; Ernerudh, J.; Pawelec, G.; Johansson, B. An immune risk phenotype, cognitive impairment, and survival in very late life: Impact of allostatic load in Swedish octogenarian and nonagenarian humans. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 556–565. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, O.; Pawelec, G.; Peralbo, E.; Wikby, A.; Mariani, E.; Mocchegiani, E.; Tarazona, R.; Solana, R. Immunological biomarkers of ageing in man: Changes in both innate and adaptive immunity are associated with health and longevity. Biogerontology. 2006, 7, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Murasko, D.M.; Weiner, P.; Kaye, D. Decline in mitogen induced proliferation of lymphocytes with increasing age. Clin. Exp. Immunol. 1987, 70, 440–448. [Google Scholar] [PubMed]
- Sansoni, P.; Brianti, V.; Fagnoni, F.; Snelli, G.; Marcato, A.; Passeri, G.; Monti, D.; Cossarizza, A.; Franceschi, C. NK cell activity and T-lymphocyte proliferation in healthy centenariansa. Ann. N. Y. Acad. Sci. 1992, 663, 505–507. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Lord, J.M.; De la Fuente, M. Preserved ex-vivo inflammatory status and cytokine responses in naturally long-lived mice. Age. 2010, 32, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M. Centenarians as a model for healthy aging. Biochem. Soc. Trans. 2003, 31, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Salvioli, S.; Capri, S.; Valensin, S.; Tieri, P.; Monti, D.; Ottaviani, E.; Franceschi, C. Inflamm-aging, cytokines and aging: State of the art, new hypothesis on the role of mitochondria and new perspectives from systems biology. Curr. Pharm. Des. 2006, 12, 3161–3171. [Google Scholar] [CrossRef] [PubMed]
- Mooradian, A.D.; Reed, R.L.; Osterweil, D.; Scuderi, P. Detectable serum levels of tumor necrosis factor α may predict early mortality in elderly institutionalized patients. J. Am. Geriatr. Soc. 1991, 39, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.B.; Ferrucci, L.; Tracy, R.P.; Corti, M.C.; Wacholder, S.; Ettinger, W.H., Jr.; Heimovitz, H.; Cohen, H.J.; Wallace, R. Associations of elevated interleukin-6 and C-reactive protein levels with mortality in the elderly. Am. J. Med. 1999, 106, 506–512. [Google Scholar] [CrossRef]
- Ferrucci, L.; Harris, T.B.; Guralnik, J.M.; Tracy, R.P.; Corti, M.C.; Cohen, H.J.; Penninx, B.; Pahor, M.; Wallace, R.; Havlik, R.J. Serum IL-6 level and the development of disability in older persons. J. Am. Geriatr. Soc. 1999, 47, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, W.; Rockwell, J.; Engleman, C.; Gewirtz, A.; Katz, J.; Sambhara, S. Cutting edge: Impaired Toll-like receptor expression and function in aging. J. Immunol. 2002, 169, 4697–4701. [Google Scholar] [CrossRef] [PubMed]
- Boehmer, E.D.; Meehan, M.J.; Cutro, B.T.; Kovacs, E.J. Aging negatively skews macrophage TLR2- and TLR4-mediated proinflammatory responses without affecting the IL-2-stimulated pathway. Mech. Ageing Dev. 2005, 126, 1305–1313. [Google Scholar]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Monti, D.; Cossarizza, A.; Fagnoni, F.; Passeri, G.; Sansoni, P. Aging, longevity and cancer: Studies in Down´s syndrome and in centenarians. Ann. N. Y. Acad. Sci. 1991, 621, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Lectin receptors as lymphocyte surface markers. Adv. Immunol. 1983, 34, 213–298. [Google Scholar] [PubMed]
- Hirokawa, K. Immunity and aging. In Principles and Practice of Geriatric Medicine; Pathy, M.S.J., Sinclair, A.J., Eds.; John Wiley and Sons Ltd.: London, UK, 1998; Volume 3, pp. 35–47. [Google Scholar]
- Pawelec, G.; Effros, C.; Caruso, C.; Remarque, E.; Barnett, Y.; Solana, R. T cells and aging. Front. Biosci. 1999, 4, 216–269. [Google Scholar] [CrossRef]
- Salam, N.; Rane, S.; Das, R.; Faulkner, M.; Gund, R.; Kandpal, U.; Lewis, V.; Mattoo, H.; Prabhu, S.; Ranganathan, V.; et al. T cell ageing: Effects of age on development, survival & function. Indian J. Med. Res. 2013, 138, 595–608. [Google Scholar] [PubMed]
- Pahlavani, M.A.; Harris, M.D. Effect of in vitro generation of oxygen free radicals on T cell function in young and old rats. Free Radic. Biol. Med. 1998, 25, 903–913. [Google Scholar] [CrossRef]
- Gougerot-Pocidalo, M.A.; Fay, M.; Roche, Y.; Chollet-Martin, S. Mechanisms by which oxidative injury inhibits the proliferative response of human lymphocytes to PHA. Effect of the thiol compound 2-mercaptoethanol. Immunology 1988, 64, 281–288. [Google Scholar] [PubMed]
- Schieber, M.; Chande, N.S. ROS Function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; González, E.M.; De la Fuente, M. Increase of oxidation and inflammation in nervous and immune systems with aging and anxiety. Curr. Pharm. Des. 2014, 20, 4656–4678. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; de Toda, I.M.; Cruces, J.; Garrido, A.; Gonzalez-Sanchez, M.; De la Fuente, M. Role of macrophages in age-related oxidative stress and lipofuscin accumulation in mice. Redox Biol. 2017, 12, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Effros, R.B.; Pawelec, G. Replicative senescence of T cells: Does the hayflick limit lead to immune exhaustion? Immunol. Today 1997, 18, 450–454. [Google Scholar] [CrossRef]
- Fülöp, T.; Larbi, A.; Pawelec, G. Human T cell aging and the impact of persistent viral infections. Front. Immunol. 2013, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.H. Activation-induced death of mature T cells in the regulation of immune responses. Curr. Opin. Immunol. 1995, 7, 382–388. [Google Scholar] [CrossRef]
- Kabelitz, D.; Janssen, O. Antigen-induced death of T-lymphocytes. Front. Biosci. 1997, 2, 61–77. [Google Scholar] [CrossRef]
- Marrack, P.; Scott-Browne, J.; MacLeod, M.K. Terminating the immune response. Immunol. Rev. 2010, 236, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, R.; Yamada, A.; Kudo, Y.; Hayashi, Y.; Ishimaru, N. Mechanism of activation-induced cell death of T cells and regulation of FASL expression. Crit. Rev. Immunol. 2014, 34, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Sprent, J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat. Rev. Immunol. 2012, 12, 180–190. [Google Scholar] [CrossRef] [PubMed]
- LaRock, C.N.; Todd, J.; LaRock, D.L.; Olson, J.; O’Donoghue, A.J.; Robertson, A.A.B.; Cooper, M.A.; Hoffman, H.M.; Nizet, V. IL-1β is an innate immune sensor of microbial proteolysis. Sci. Immunol. 2016, 1, eaah3539. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. CSH. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M. Interleukin 17 is a chief orchestrator of immunity. Nat. Immunol. 2017, 18, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Simioni, PU.; Costa, E.H.; Tamashiro, W.M. Aging reduces the primary humoral response and the in vitro cytokine production in mice. Braz. J. Med. Biol. Res. 2007, 40, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Brüünsgaard, H.; Pedersen, B.K. Age-related inflammatory cytokines and disease. Immunol. Allergy Clin. N. Am. 2003, 23, 15–39. [Google Scholar] [CrossRef]
- Oberley, L.W.; Oberley, T.D.; Buettner, G.R. Cell division in normal and transformed cells: The possible role of superoxide and hydrogen peroxide. Med. Hypotheses 1981, 7, 21–42. [Google Scholar] [CrossRef]
- Kim, B.Y.; Han, M.J.; Chung, A.S. Effects of reactive oxygen species on proliferation of Chinese hamster lung fibroblast (V79) cells. Free Radic. Biol. Med. 2001, 30, 686–698. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Salvioli, S.; Monti, D.; Lanzarini, C.; Conte, M.; Pirazzini, C.; Giulia Bacalini, M.; Garagnani, P.; Giuliani, C.; Fontanesi, E.; Ostan, R.; et al. Immune system, cell senescence, aging and longevity–inflamm-aging reappraised. Curr. Pharm. Des. 2013, 19, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Minciullo, P.L.; Catalano, A.; Mandraffino, G.; Casciaro, M.; Crucitti, A.; Maltese, G.; Morabito, N.; Lasco, A.; Gangemi, S.; Basile, G. Inflammaging and anti-inflammaging: The role of cytokines in extreme longevity. Arch. Immunol. Ther. Exp. 2016, 64, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijević, M.; Aleksić, I.; Vujić, V.; Stanojević, S.; Pilipović, I.; von Hörsten, S.; Leposavić, G. Peritoneal exudate cells from long-lived rats exhibit increased IL-10/IL-1β expression ratio and preserved NO/urea ratio following LPS-stimulation in vitro. Age 2014, 36, 9696. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.I.; Banning, U.; Kim, Y.M.; Verheyen, J.; Hannen, M.; Bönig, H.; Körholz, D. Interleukin 10 inhibits TNF-α production in human monocytes independently of interleukin 12 and interleukin 1β. Immunol. Investig. 1999, 28, 165–175. [Google Scholar] [CrossRef]
- Dagvadorj, J.; Naiki, Y.; Tumurkhuu, G.; Hassan, F.; Islam, S.; Koide, N.; Mori, I.; Yoshida, T.; Yokochi, T. Interleukin-10 inhibits tumor necrosis factor-α production in lipopolysaccharide-stimulated RAW 264.7 cells through reduced MyD88 expression. Innate Immun. 2008, 14, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Van Den Biggelaar, A.H.; De Craen, A.J.; Gussekloo, J.; Huizinga, T.W.; Heijmans, B.T.; Frolich, M.; Kirkwood, T.B.; Westendorp, R.G. Inflammation underlying cardiovascular mortality is a late consequence of evolutionary programming. FASEB J. 2004, 18, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Krabbe, K.S.; Pedersen, M.; Bruunsgaard, H. Inflammatory mediators in the elderly. Exp. Gerontol. 2004, 39, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.T.; Scichilone, N.; Paganelli, R.; Minciullo, P.L.; Patella, V.; Bonini, M.; Passalacqua, G.; Lombardi, C.; Simioni, L.; Ridolo, E.; et al. Allergic diseases in the elderly: Biological characteristics and main immunological and non-immunological mechanisms. Clin. Mol. Allergy 2017, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, C.; Alvarez, P.; Puerto, M.; Gausserès, N.; Jiménez, L.; De la Fuente, M. Dietary supplementation with antioxidants improves functions and decreases oxidative stress of leukocytes from prematurely aging mice. Nutrition 2006, 22, 767–777. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez de Toda, I.; Vida, C.; De la Fuente, M. An Appropriate Modulation of Lymphoproliferative Response and Cytokine Release as Possible Contributors to Longevity. Int. J. Mol. Sci. 2017, 18, 1598. https://doi.org/10.3390/ijms18071598
Martínez de Toda I, Vida C, De la Fuente M. An Appropriate Modulation of Lymphoproliferative Response and Cytokine Release as Possible Contributors to Longevity. International Journal of Molecular Sciences. 2017; 18(7):1598. https://doi.org/10.3390/ijms18071598
Chicago/Turabian StyleMartínez de Toda, Irene, Carmen Vida, and Mónica De la Fuente. 2017. "An Appropriate Modulation of Lymphoproliferative Response and Cytokine Release as Possible Contributors to Longevity" International Journal of Molecular Sciences 18, no. 7: 1598. https://doi.org/10.3390/ijms18071598
APA StyleMartínez de Toda, I., Vida, C., & De la Fuente, M. (2017). An Appropriate Modulation of Lymphoproliferative Response and Cytokine Release as Possible Contributors to Longevity. International Journal of Molecular Sciences, 18(7), 1598. https://doi.org/10.3390/ijms18071598