Candida albicans Impairments Induced by Peppermint and Clove Oils at Sub-Inhibitory Concentrations


Abstract
:
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Yeast
3.2. Essential Oils
3.3. Colony Morphology Assay
3.4. Biochemical and Enzymatic Profiles
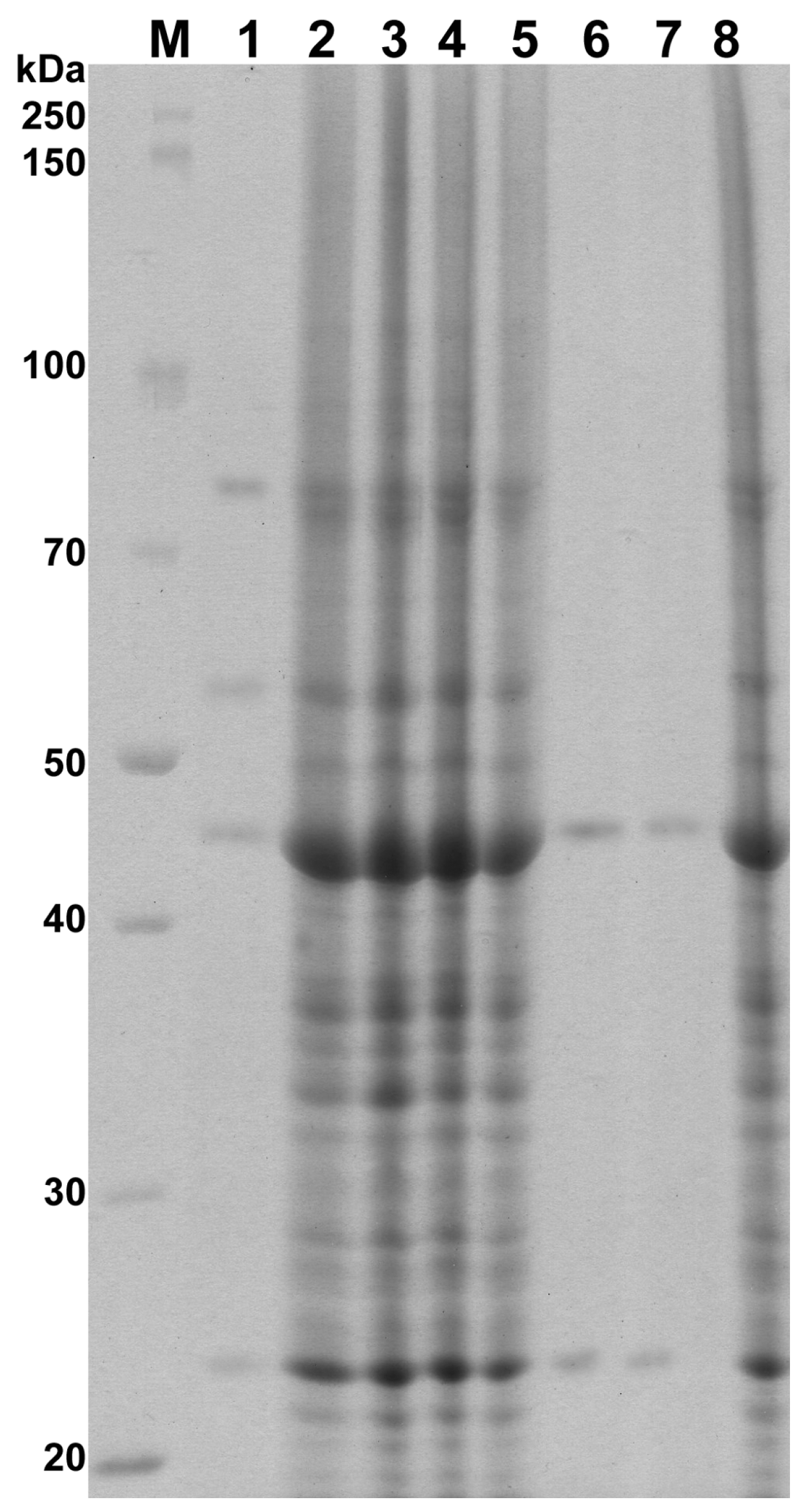
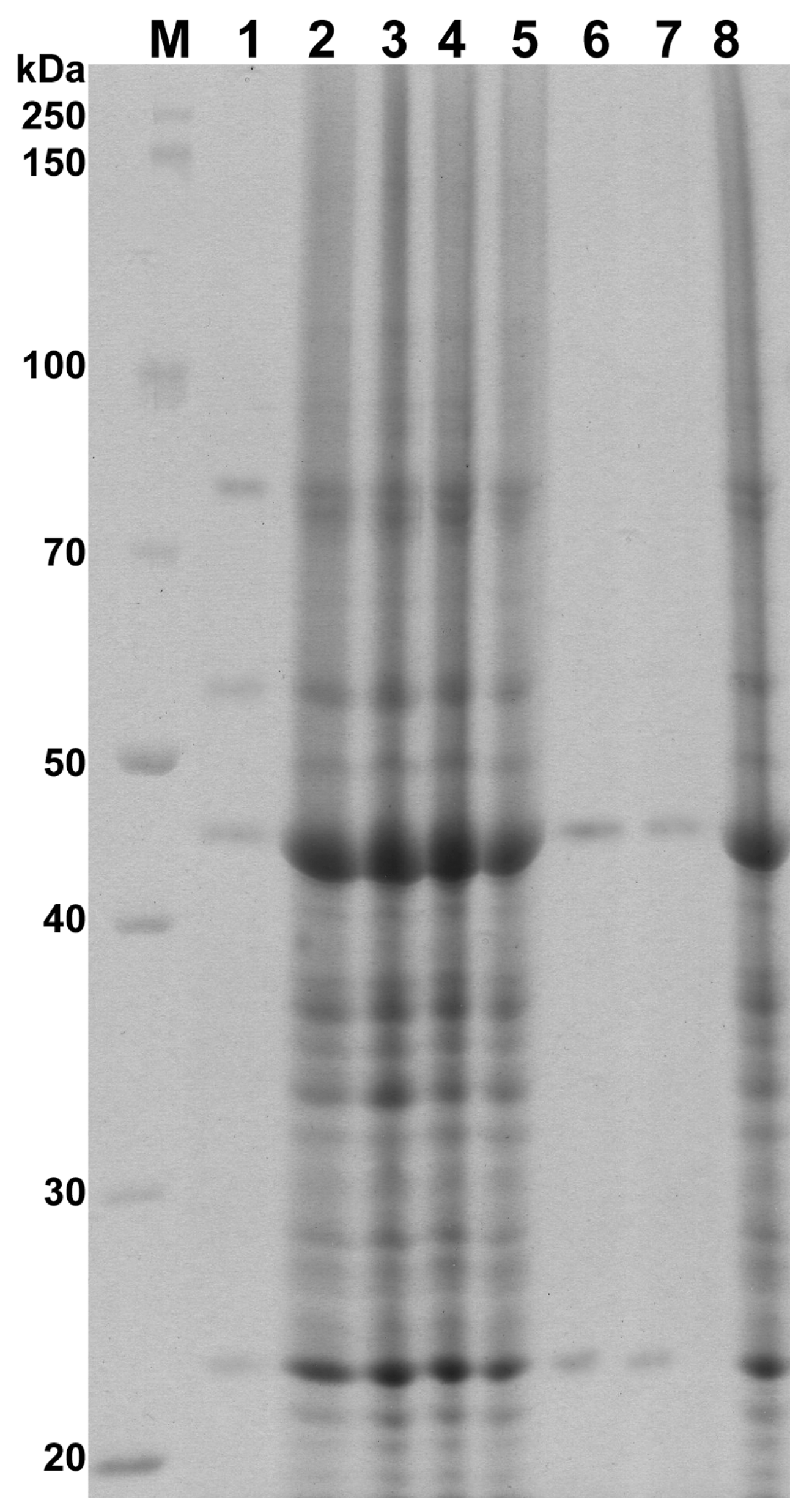
3.5. Protein Preparation and Electrophoresis
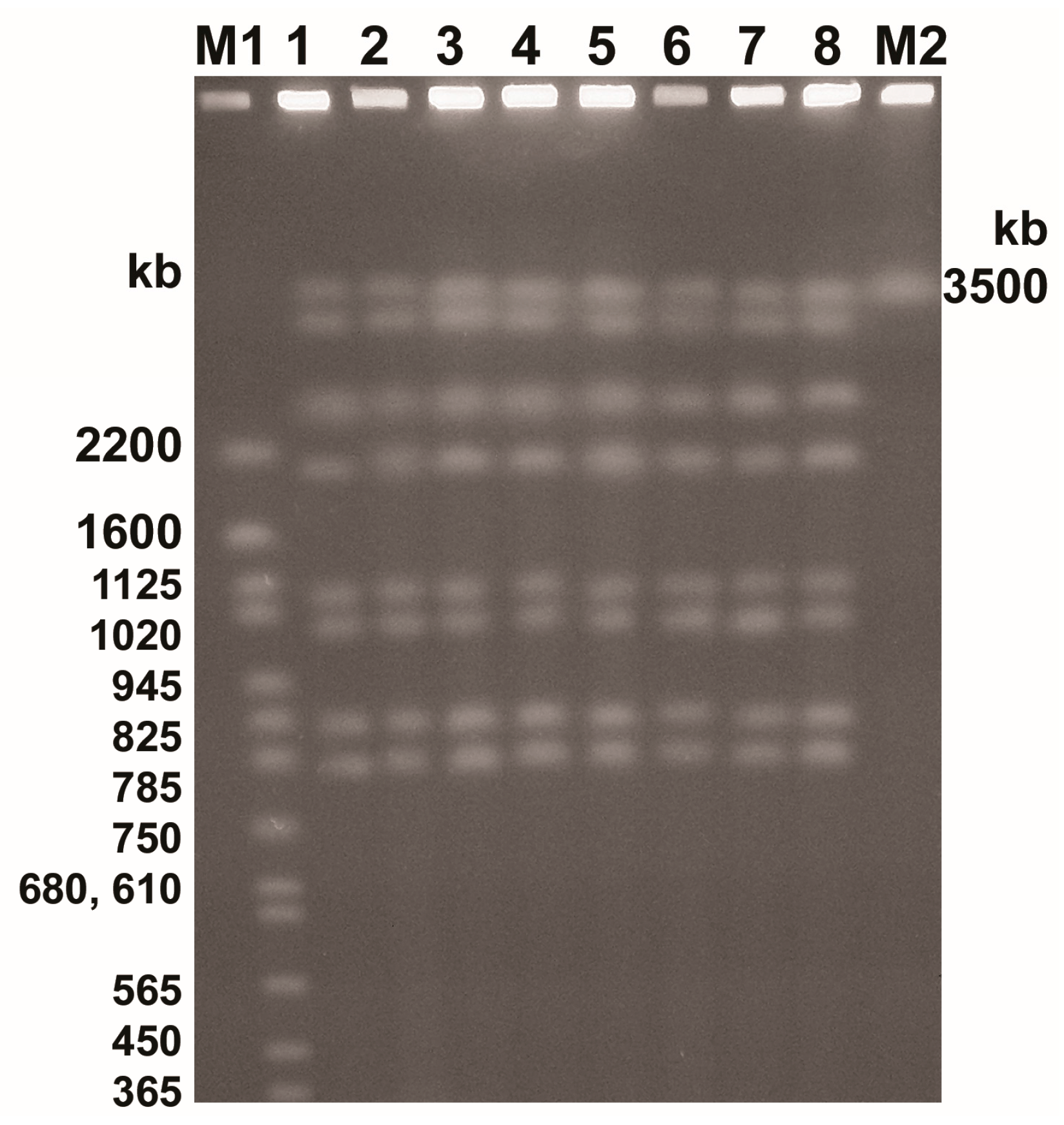
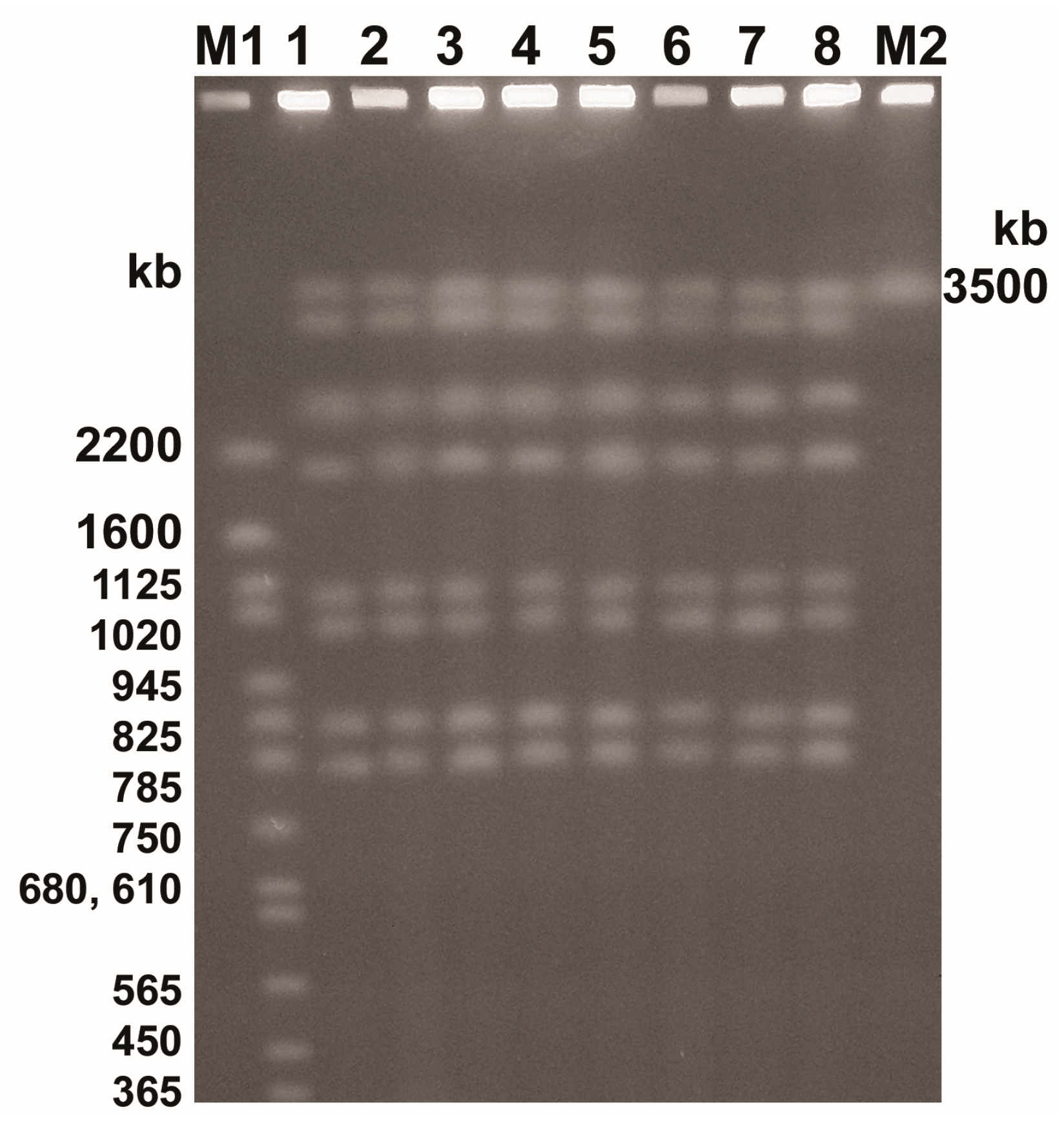
3.6. Chromosomal DNA Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| EO | Essential oil |
| PO | Peppermint oil |
| CLO | Clove oil |
| C. albicans | Candida albicans |
| MIC | Minimal inhibitory concentration |
References
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A.; Global Antifungal Surveillance Group. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [PubMed]
- Nawrot, U.; Pajączkowska, M.; Fleischer, M.; Przondo-Mordarska, H.; Samet, A.; Piasecka-Pazik, D.; Komarnicka, J.; Sulik-Tyszka, B.; Swoboda-Kopeć, E.; Cieślik, J.; et al. Candidaemia in polish hospitals—A multicentre survey. Mycoses 2013, 56, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Cross, E.W.; Park, S.; Perlin, D.S. Cross-resistance of clinical isolates of Candida albicans and Candida glabrata to over-the-counter azoles used in the treatment of vaginitis. Microb. Drug Resist. 2000, 6, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Das, I.; Nightingale, P.; Patel, M.; Jumaa, P. Epidemiology, clinical characteristics, and outcome of candidemia: Experience in a tertiary referral center in the UK. Int. J. Infect. Dis. 2011, 15, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Kahla-Nakbi, A.B.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): A short review. Phytother. Res. 2007, 21, 501–506. [Google Scholar]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Kunicka-Styczyńska, A.; Sikora, M.; Kalemba, D. Lavender, tea tree and lemon oils as antimicrobials in washing liquids and soft body balms. Int. J. Cosmet. Sci. 2011, 33, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V.; Nielsen, J.B. A review of the toxicity of Melaleuca alternifolia (tea tree) oil. Food Chem. Toxicol. 2006, 44, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Bennis, S.; Chami, F.; Chami, N.; Bouchikhi, T.; Remmal, A. Surface alteration of Saccharomyces cerevisiae induced by thymol and eugenol. Lett. Appl. Microbiol. 2004, 38, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Tampieri, M.P.; Galuppi, R.; Macchioni, F.; Carelle, M.S.; Falcioni, L.; Cioni, P.L.; Morelli, I. The inhibition of Candida albicans by selected essential oils and their major components. Mycopathologia 2005, 159, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Stringaro, A.; Vavala, E.; Colone, M.; Pepi, F.; Mignogna, G.; Garzoli, S.; Cecchetti, S.; Ragno, R.; Angiolella, L. Effects of Mentha suaveolens essential oil alone or in combination with other drugs in Candida albicans. J. Evid. Based Complement. Altern. Med. 2014, 2014, 125904. [Google Scholar]
- Rajkowska, K.; Kunicka-Styczyńska, A.; Maroszyńska, M.; Dąbrowska, M. The effect of thyme and tea tree oils on morphology and metabolism of Candida albicans. Acta Biochim. Pol. 2014, 61, 305–310. [Google Scholar] [PubMed]
- Rajkowska, K.; Kunicka-Styczyńska, A.; Maroszyńska, M. Selected essential oils as antifungal agents against antibiotic-resistant Candida spp.: In vitro study on clinical and food-borne isolates. Microb. Drug Resist. 2017, 23, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Granek, J.A.; Magwene, P.M. Environmental and genetic determinants of colony morphology in yeast. PLoS Genet. 2010, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gyurko, C.; Lendenmann, U.; Troxler, R.F.; Oppenheim, F.G. Candida albicans mutants deficient in respiration are resistant to the small cationic salivary antimicrobial peptide histatin 5. Antimicrob. Agents Chemother. 2000, 44, 348–354. [Google Scholar] [CrossRef]
- Uribe, S.; Ramirez, J.; Pena, A. Effects of β-pinene on yeast membrane functions. J. Bacteriol. 1985, 161, 1195–1200. [Google Scholar] [PubMed]
- Rajkowska, K.; Nowak, A.; Kunicka-Styczyńska, A.; Siadura, A. Biological effects of various chemically characterized essential oils: Investigation of the mode of action against Candida albicans and HeLa cells. RSC Adv. 2016, 6, 97199–97207. [Google Scholar] [CrossRef]
- Ernst, J.F. Transcription factors in Candida albicans—Environmental control of morphogenesis. Microbiology 2000, 146, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Forche, A.; Berman, J. Genomic plasticity of the human fungal pathogen Candida albicans. Eukaryot. Cell 2010, 9, 991–1008. [Google Scholar] [CrossRef] [PubMed]
- Klempp-Selb, B.; Rimek, D.; Kappe, R. Karyotyping of Candida albicans and Candida glabrata from patients with Candida sepsis. Mycoses 2000, 43, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Zhiri, A.; Idaomar, M. Cytotoxicity and gene induction by some essential oils in the yeast Saccharomyces cerevisiae. Mutat. Res. 2005, 585, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fleck, A.B.; Schöbel, F.; Brock, M. Nutrient acquisition by pathogenic fungi: Nutrient availability, pathway regulation, and differences in substrate utilization. Int. J. Med. Microbiol. 2011, 301, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, W.L.; López-Ribot, J.L.; Casanova, M.; Gozalbo, D.; Martínez, J.P. Cell wall and secreted proteins of Candida albicans: Identification, function, and expression. Microbiol. Mol. Biol. Rev. 1998, 62, 130–180. [Google Scholar] [PubMed]
- Abaci, O. Investigation of extracellular phospholipase and proteinase activities of Candida species isolated from individuals denture wearers and genotypic distribution of Candida albicans strains. Curr. Microbiol. 2011, 62, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Monod, M.; Borg-von, Z.M. Secreted proteinases and other virulence mechanisms of Candida albicans. Chem. Immunol. 2002, 81, 114–128. [Google Scholar] [PubMed]
- Shahid, M.; Malik, A.; Rizvi, M.W.; Singhai, M. Protein profile of a fluconazole-resistant Candida albicans isolated from HIV-1 infected patient: Evaluation of protein extraction methods and development of a simple procedure. Glob. J. Biochem. Biotechnol. 2006, 1, 1–6. [Google Scholar]
- Berber, I.; Ekin, S. Discrimination and numerical analysis of human pathogenic Candida albicans strains based on SDS-PAGE protein profiles. Afr. J. Biotechnol. 2008, 7, 425–429. [Google Scholar]
- Boriollo, M.F.G.; Rosa, E.A.R.; Rosa, R.T.; Höfling, J.F. Criteria for Candida albicans numerical analysis based on electrophoretic protein patterns. Rev. Argent. Microbiol. 2000, 32, 123–128. [Google Scholar] [PubMed]
- Owen, R.J.; Jackman, P.J.H. The similarities between Pseudomonas paucimobilis and allied bacteria derived from analysis of deoxyribonucleic acids and electrophoretic protein patterns. J. Gen. Microbiol. 1982, 128, 2945–2954. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Hood, J.R.; Wilkinson, J.M.; Cavanagh, H.M.A. Evaluation of common antibacterial screening methods utilized in essential oil research. J. Essent. Oil Res. 2003, 15, 428–433. [Google Scholar] [CrossRef]
- Duran, N.; Öztürk, F.; Açık, L.; Aslantaş, Ö.; Aslan, G. Numerical analysis of Candida species from uriner system infections based on SDS-PAGE and detection of antifungal resistance. Eur. J. Gen. Med. 2007, 4, 100–106. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.C.; Cantor, C. Separation of yeast chromosome-sized DNAs by pulsed field gradient gel electrophoresis. Cell 1984, 37, 67–75. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil | Oil Concentration (% v/v) | The Number of Colonies with Different Morphology/the Total Number of Colonies on the Plate 1 | Representative Colony with Different Morphology | Isolate Acronym |
|---|---|---|---|---|
| Peppermint oil | 0.25 | 1/1 |  | PO/0.25 |
| 0.015 | 11/19 |  | PO/0.015 | |
| 0.0075 | 10/26 |  | PO/0.0075/1 | |
| 13/26 |  | PO/0.0075/2 | ||
| Clove oil | 0.25 | 4/5 |  | CLO/0.25 |
| 0.125 | 1/1 |  | CLO/0.125 | |
| 0.0075 | 9/11 |  | CLO/0.0075 | |
| Without essential oil | — | 0/28 |  | Control |
| Isolate | Whole Cell Proteins (kDa) | Dice Index of Similarity (%) |
|---|---|---|
| Control | 110, 95, 89, 80, 75, 65, 56, 49, 44, 40, 39, 38, 36, 35, 34, 33, 32, 31, 30, 28, 27, 26, 25, 24, 23, 21, 20, 19 | — |
| PO/0.25 | 80, 56, 44, 23 | 25.0 |
| PO/0.015 | 110, 95, 89, 80, 75, 65, 56, 49, 44, 40, 39, 38, 36, 35, 34, 33, 32, 31, 30, 28, 27, 26, 25, 24, 23, 21, 20, 19 | 100.0 |
| PO/0.0075/1 | 110, 95, 89, 80, 75, 65, 56, 49, 44, 40, 39, 38, 36, 35, 34, 33, 32, 31, 30, 28, 27, 26, 25, 24, 23, 21, 20, 19 | 100.0 |
| PO/0.0075/2 | 110, 95, 89, 80, 75, 65, 56, 49, 44, 40, 39, 38, 36, 35, 34, 33, 32, 31, 30, 28, 27, 26, 25, 24, 23, 21, 20, 19 | 100.0 |
| CLO/0.25 | 44, 23 | 13.3 |
| CLO/0.125 | 44, 23 | 13.3 |
| CLO/0.0075 | 110, 95, 89, 80, 75, 65, 56, 49, 44, 40, 39, 38, 36, 35, 34, 33, 32, 31, 30, 28, 27, 26, 25, 24, 23, 21, 20, 19 | 100.0 |
| Compound | RI | Peppermint Oil | Clove oil |
|---|---|---|---|
| Content (%) | |||
| α-Pinene | 934 | 0.6 | – |
| Sabinene | 968 | 0.2 | – |
| β-Pinene | 974 | 1.0 | – |
| p-Cymene | 1016 | 0.3 | – |
| 1,8-Cineole | 1020 | 6.6 | – |
| Limonene | 1025 | 2.4 | – |
| trans-Sabinene hydrate | 1060 | 0.2 | – |
| 2-Methylbutyl 2-methylbutanoate | 1094 | 0.1 | – |
| Menthone | 1138 | 23.1 | – |
| Isomenthone | 1145 | 3.8 | – |
| Menthofuran | 1154 | 2.2 | – |
| Neomenthol | 1155 | 3.3 | – |
| Menthol | 1163 | 43.9 | – |
| Neoisomenthol | 1173 | 1.0 | – |
| α-Terpineol | 1178 | 0.6 | – |
| Isomenthol | 1179 | 0.2 | – |
| Pulegone | 1218 | 1.2 | – |
| Piperitone | 1237 | 0.4 | – |
| Menthyl acetate | 1279 | 4.9 | – |
| Eugenol | 1342 | – | 85.2 |
| β-Burbonene | 1381 | 0.1 | – |
| Methyleugenol | 1386 | – | 0.2 |
| (E)-β-Caryophyllene | 1421 | 1.6 | 9.9 |
| α-Humulene | 1453 | 0.1 | 1.9 |
| Germacrene D | 1474 | 0.3 | – |
| δ-Cadinene | 1513 | – | 0.4 |
| (E)-β-Caryophyllene oxide | 1573 | 0.2 | 0.4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajkowska, K.; Otlewska, A.; Kunicka-Styczyńska, A.; Krajewska, A. Candida albicans Impairments Induced by Peppermint and Clove Oils at Sub-Inhibitory Concentrations. Int. J. Mol. Sci. 2017, 18, 1307. https://doi.org/10.3390/ijms18061307
Rajkowska K, Otlewska A, Kunicka-Styczyńska A, Krajewska A. Candida albicans Impairments Induced by Peppermint and Clove Oils at Sub-Inhibitory Concentrations. International Journal of Molecular Sciences. 2017; 18(6):1307. https://doi.org/10.3390/ijms18061307
Chicago/Turabian StyleRajkowska, Katarzyna, Anna Otlewska, Alina Kunicka-Styczyńska, and Agnieszka Krajewska. 2017. "Candida albicans Impairments Induced by Peppermint and Clove Oils at Sub-Inhibitory Concentrations" International Journal of Molecular Sciences 18, no. 6: 1307. https://doi.org/10.3390/ijms18061307
APA StyleRajkowska, K., Otlewska, A., Kunicka-Styczyńska, A., & Krajewska, A. (2017). Candida albicans Impairments Induced by Peppermint and Clove Oils at Sub-Inhibitory Concentrations. International Journal of Molecular Sciences, 18(6), 1307. https://doi.org/10.3390/ijms18061307