
Protective Effect of Fragaria ananassa Crude Extract on Cadmium-Induced Lipid Peroxidation, Antioxidant Enzymes Suppression, and Apoptosis in Rat Testes

 ,
,
Abstract
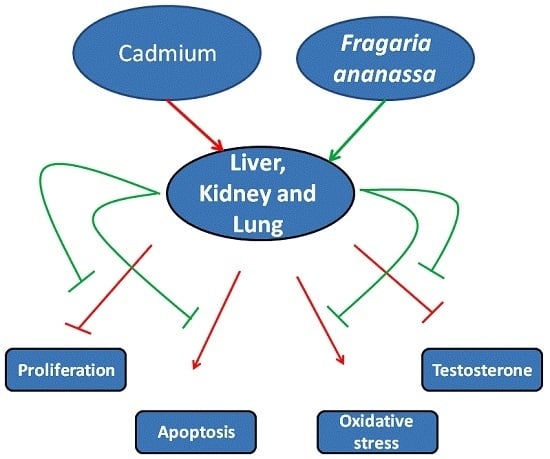
:
1. Introduction
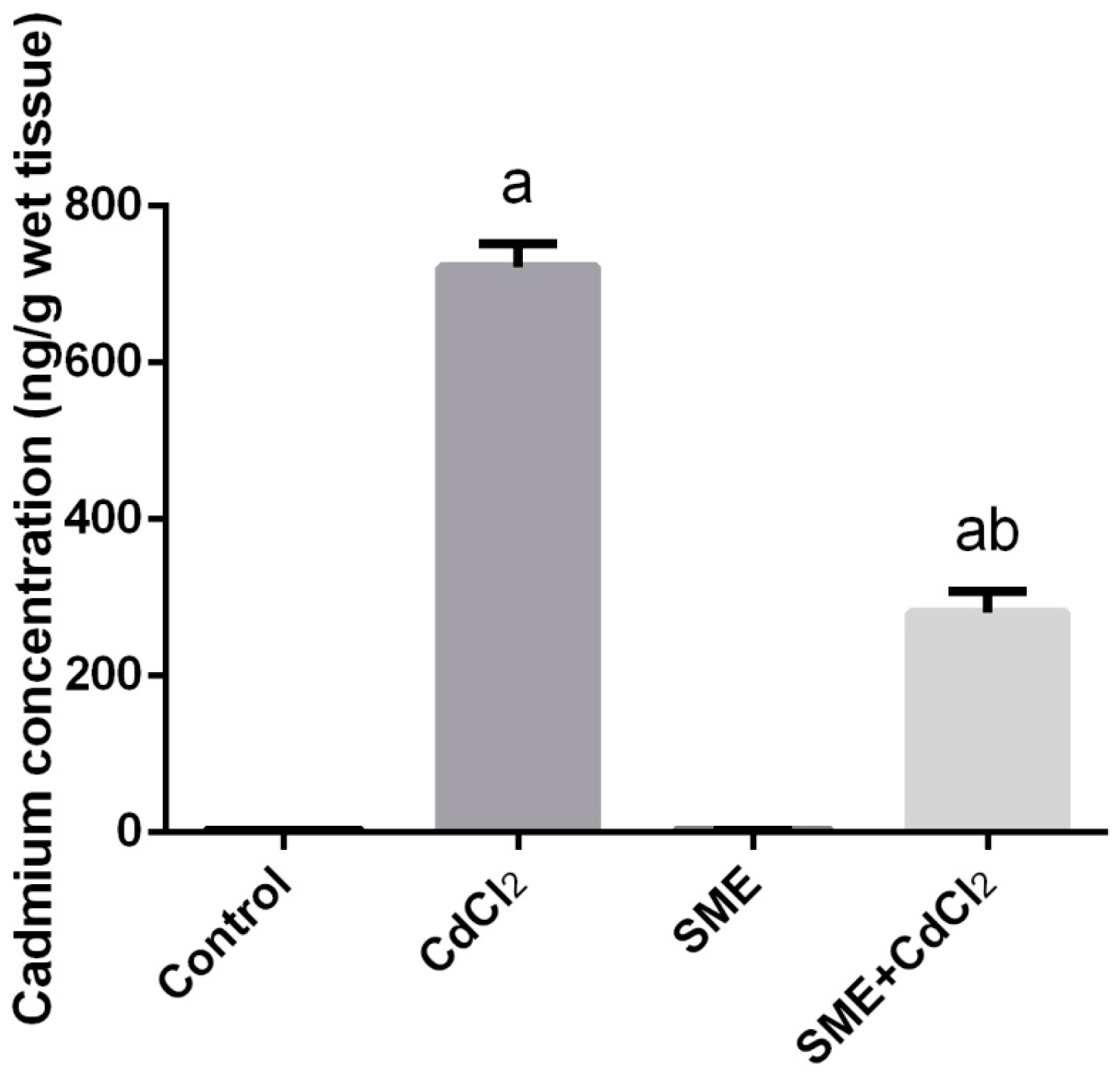
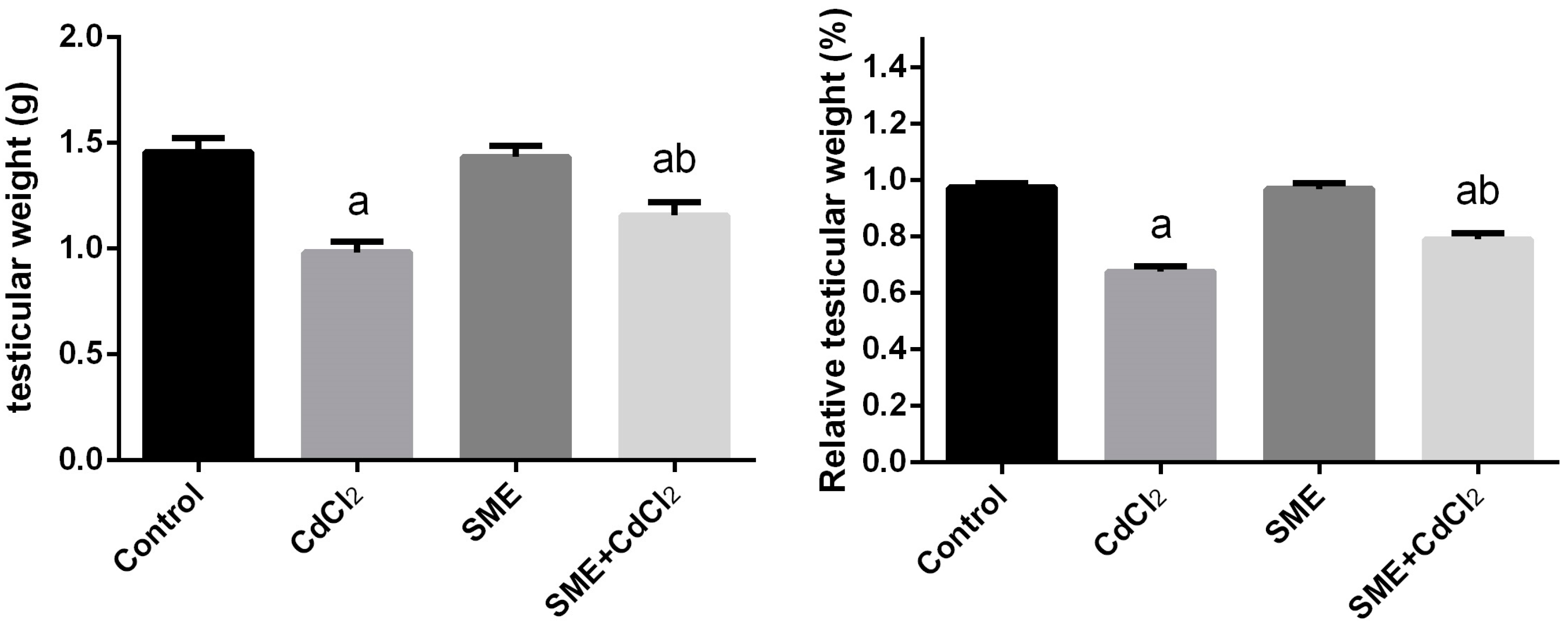
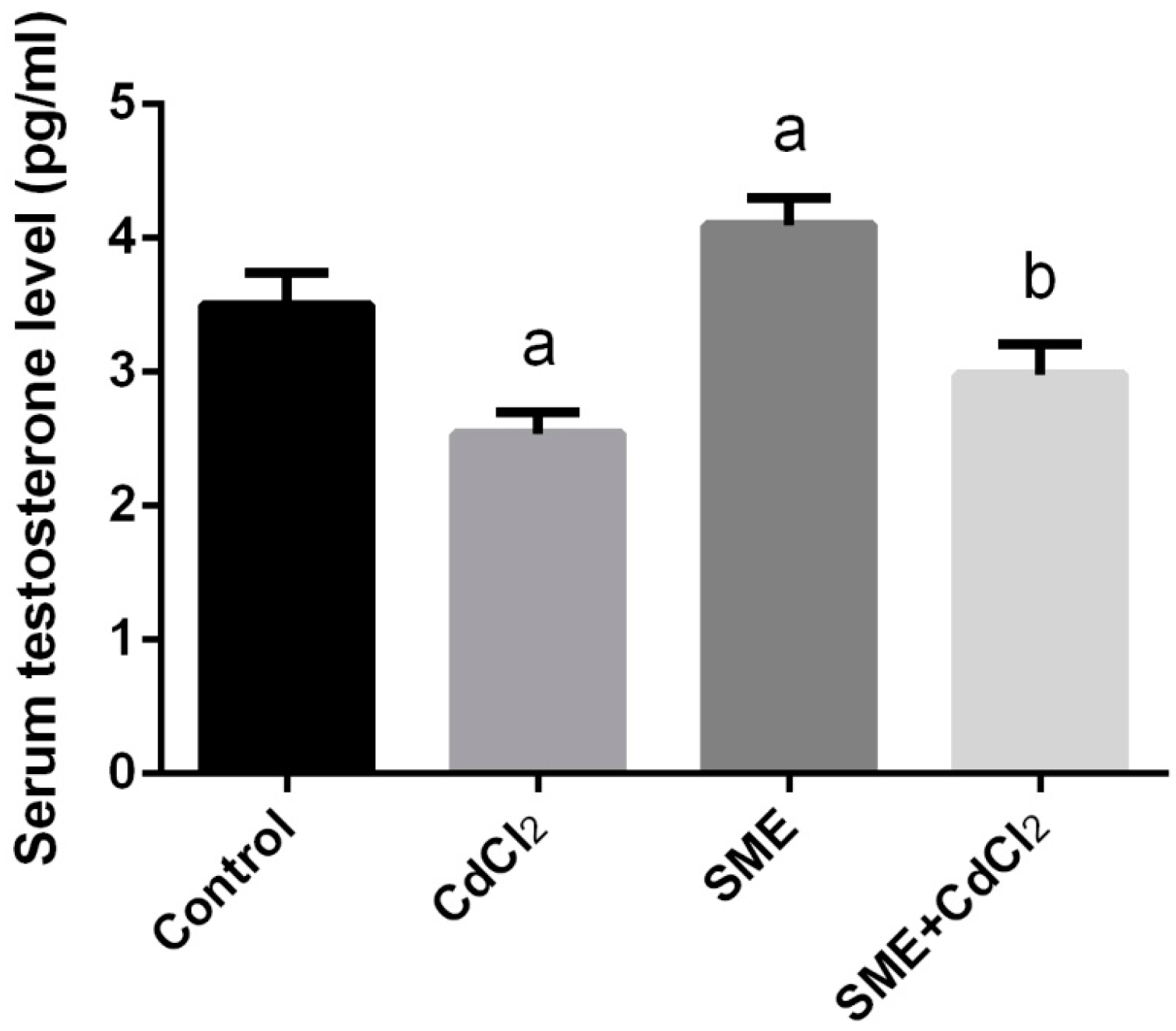
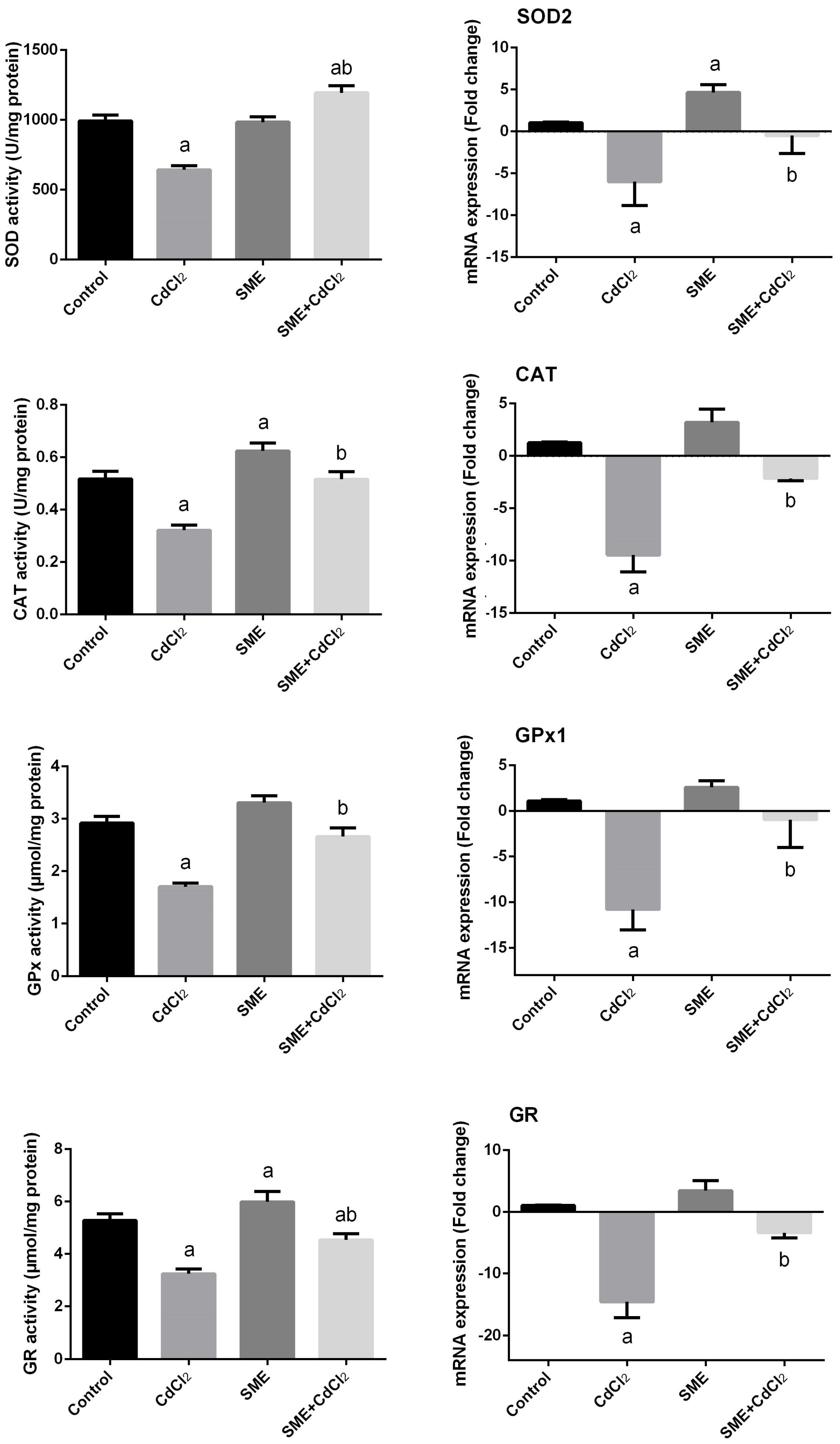
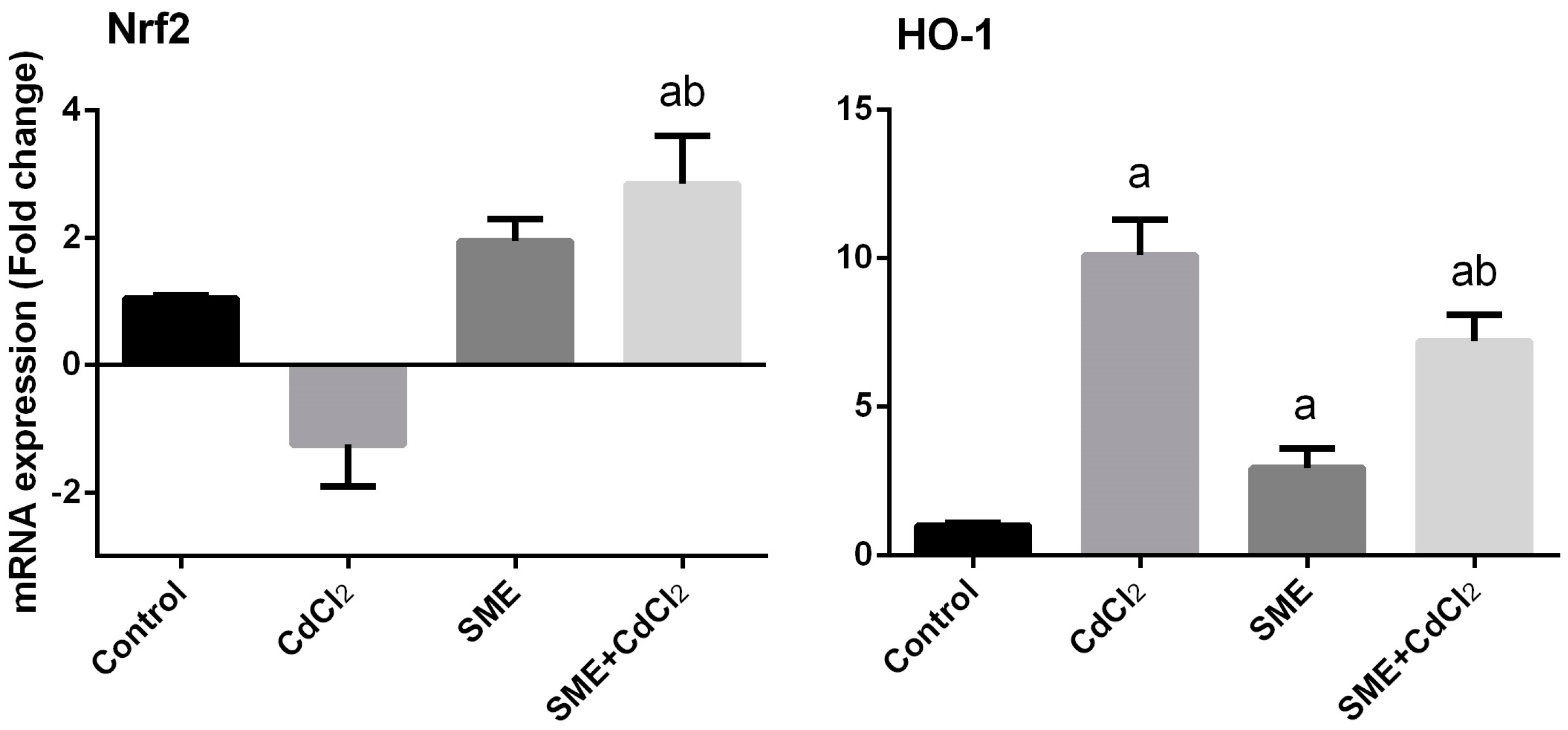
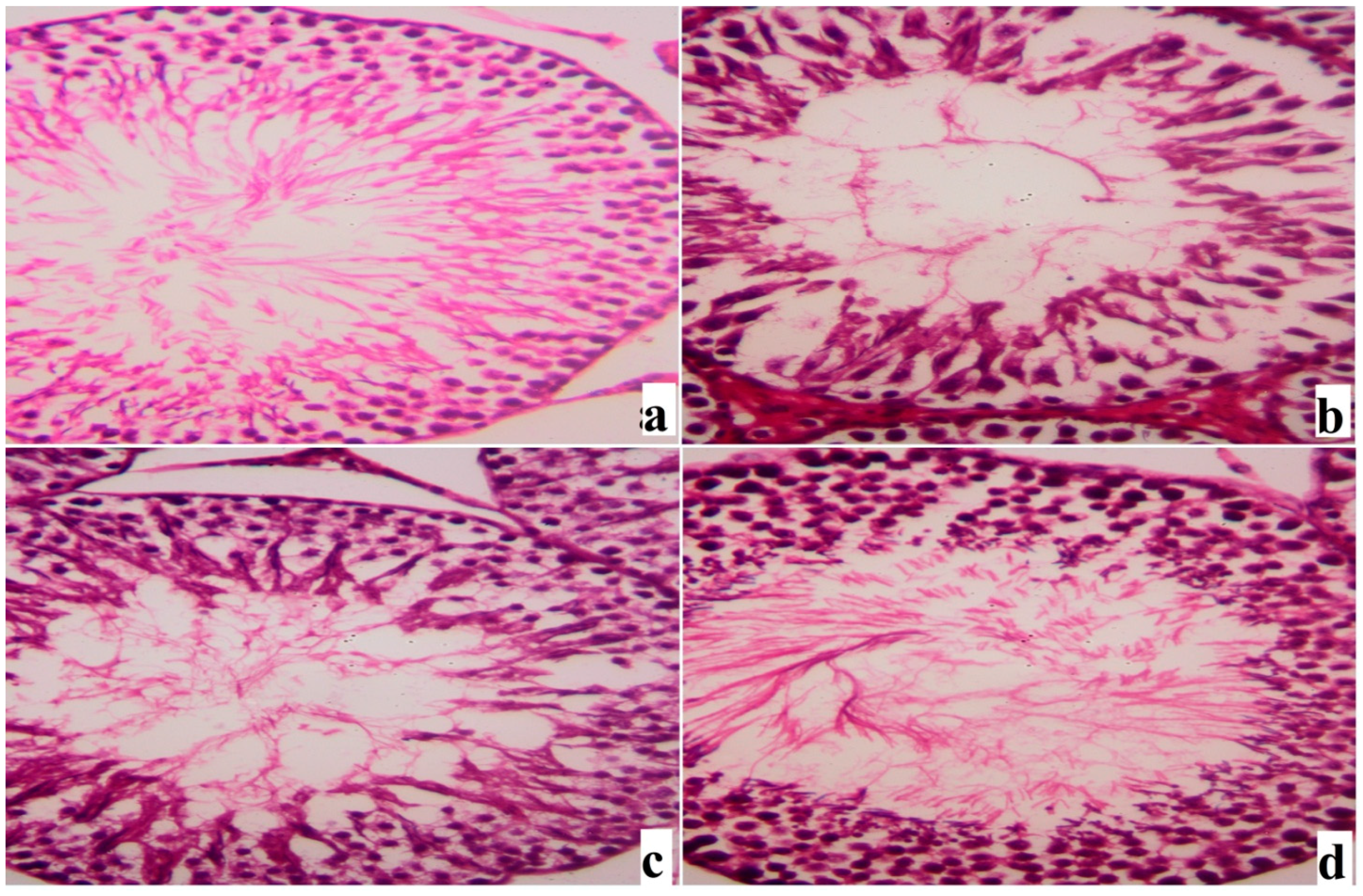
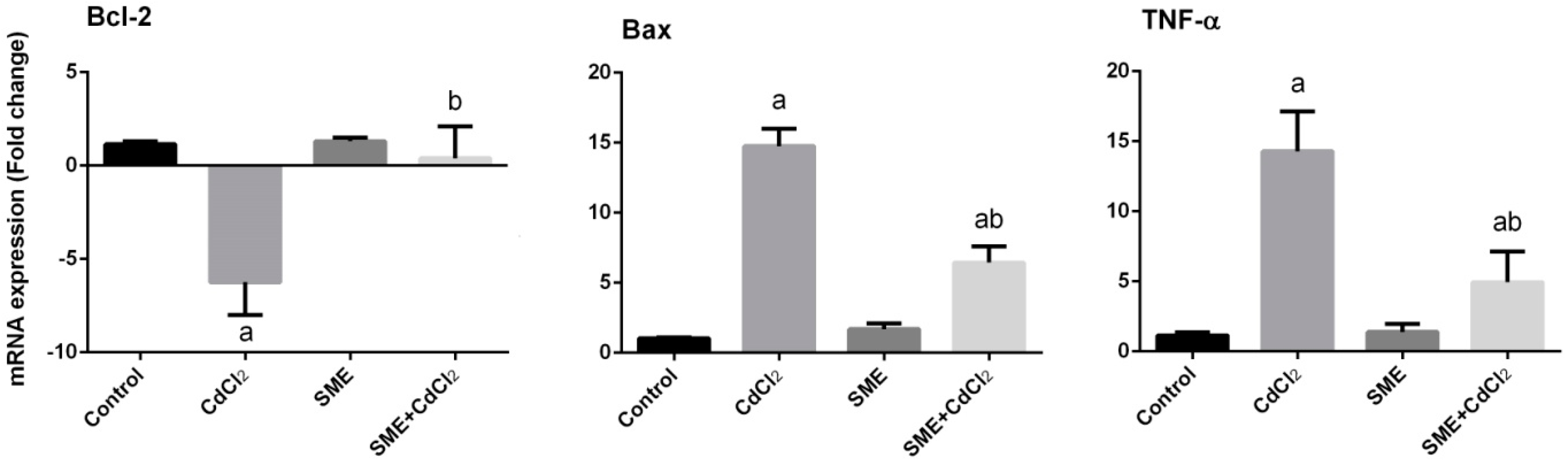
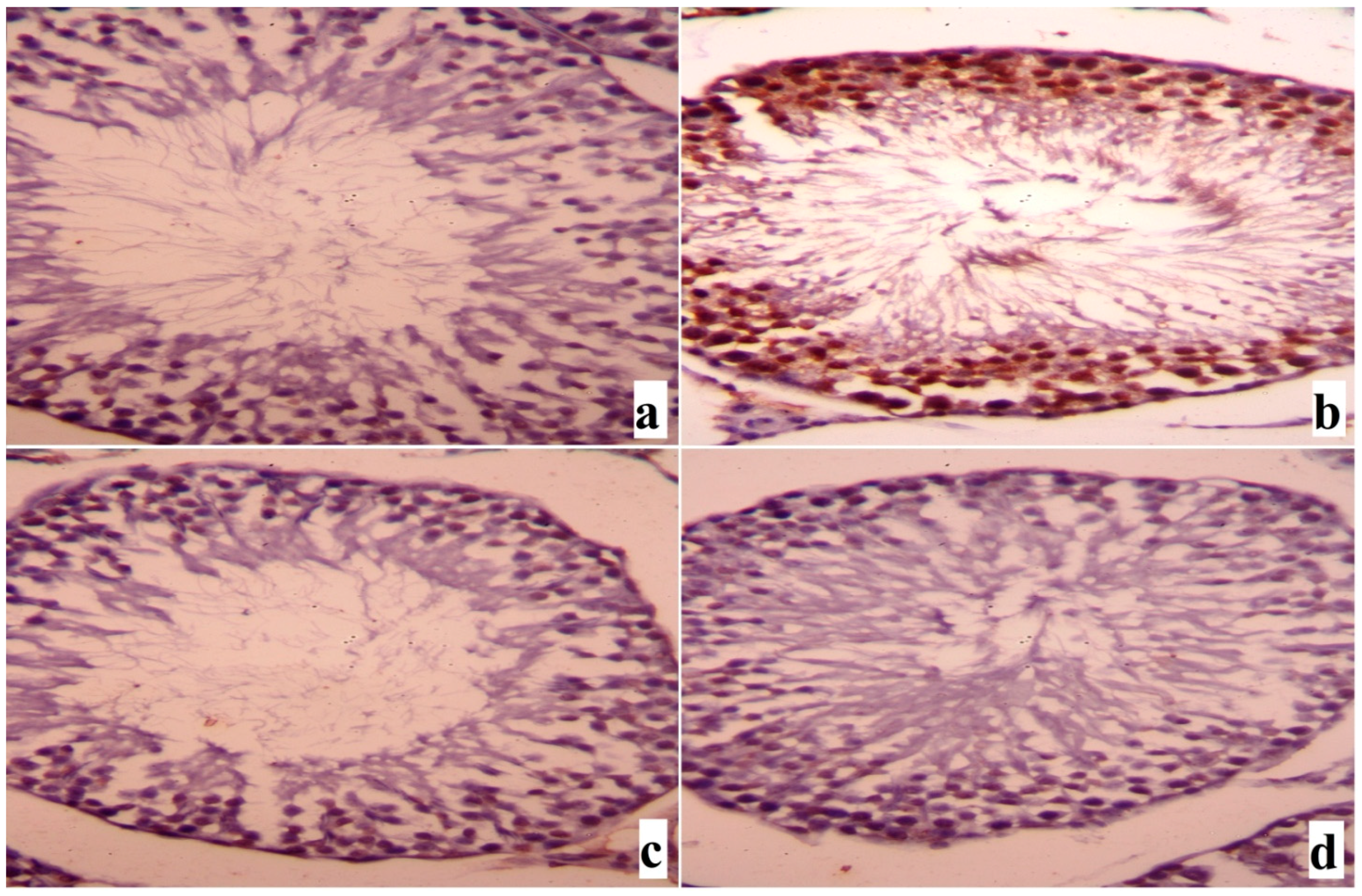
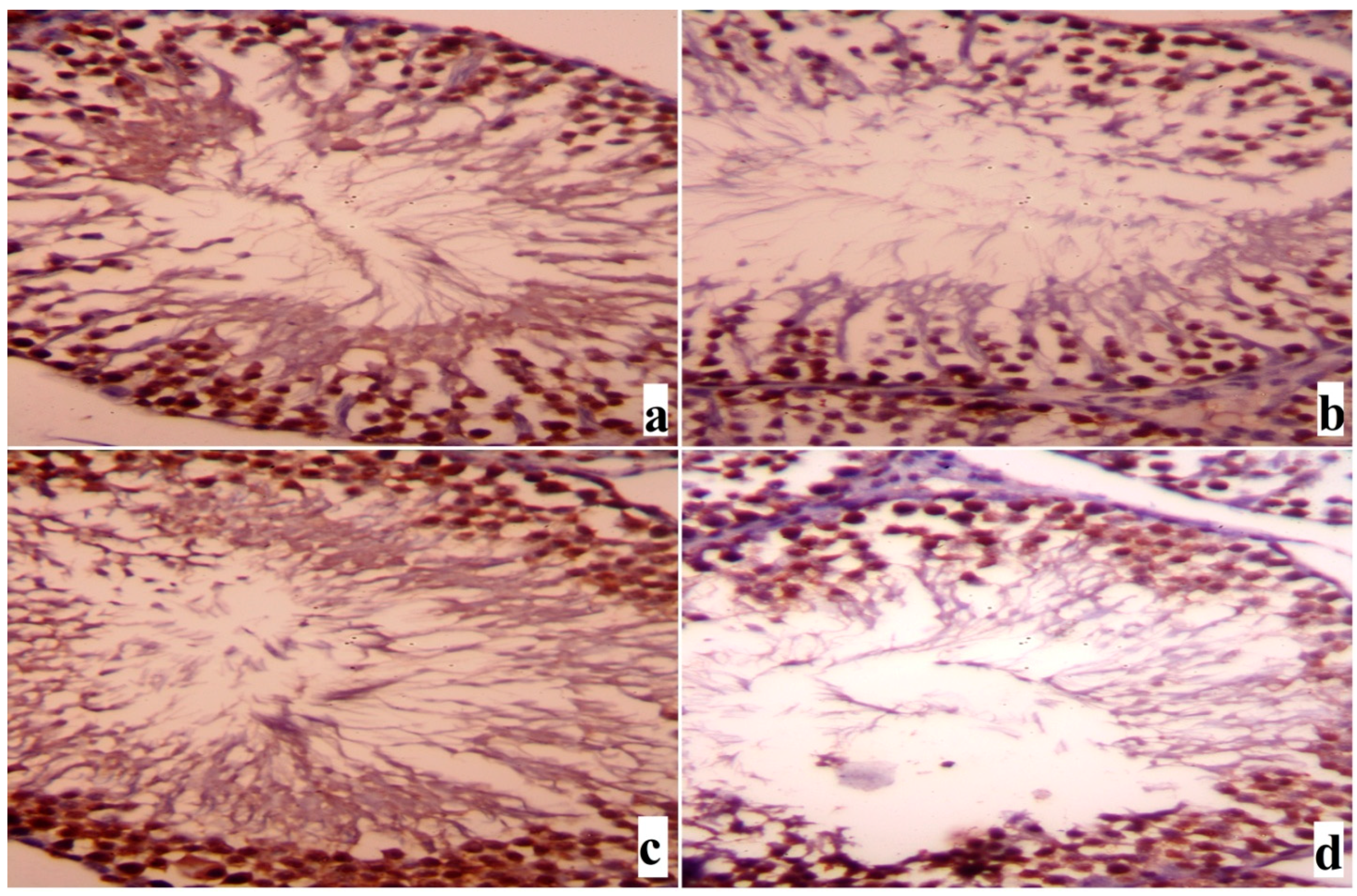
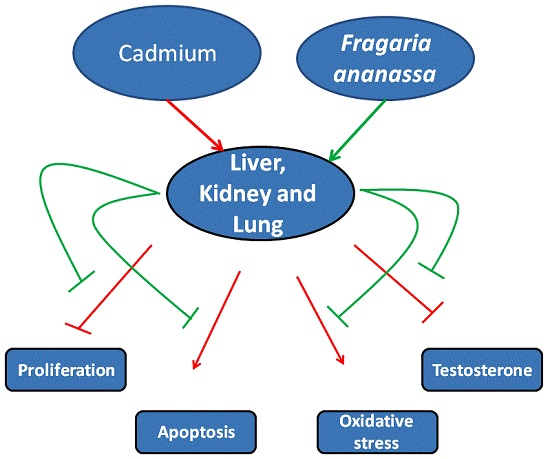
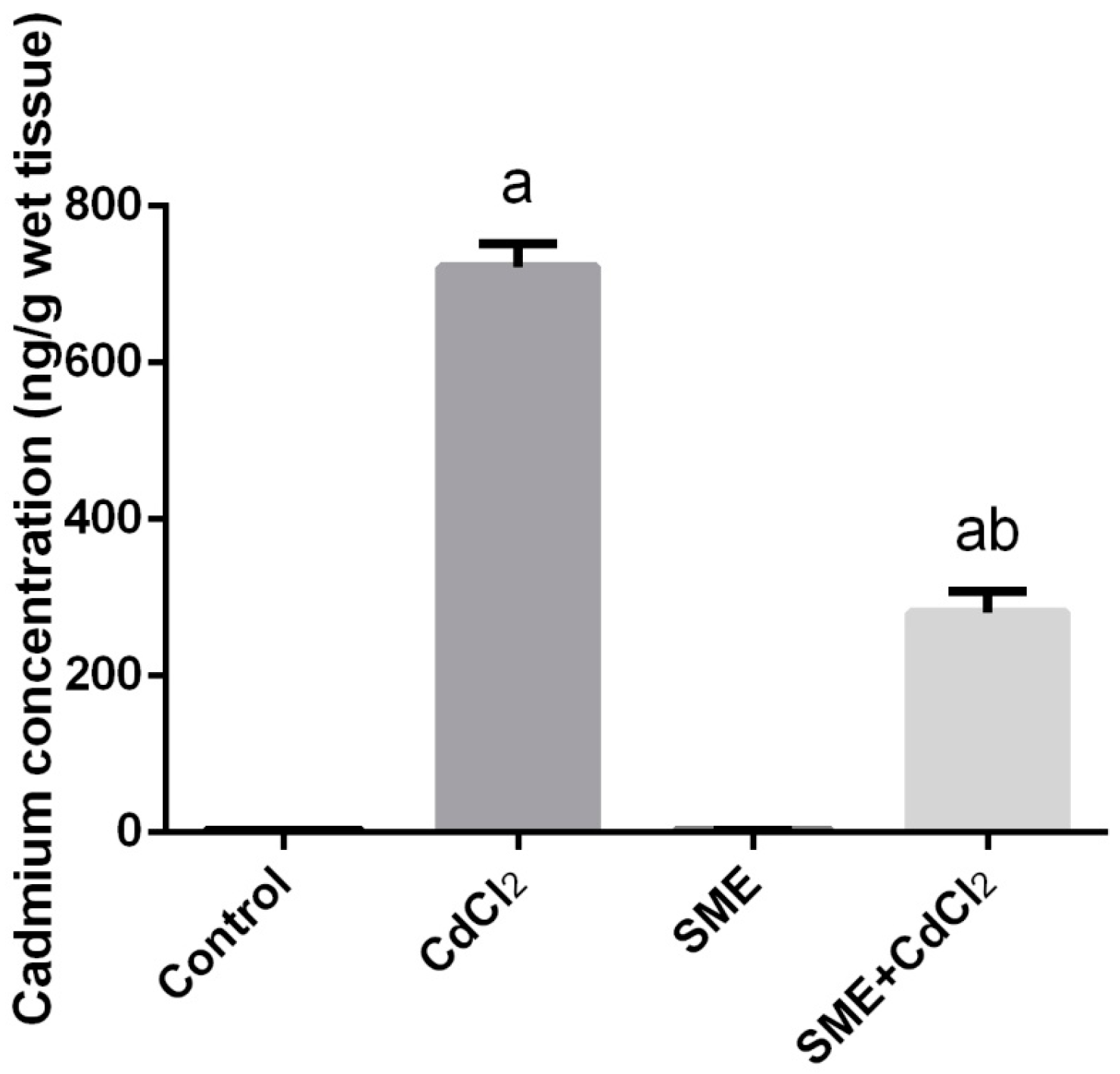
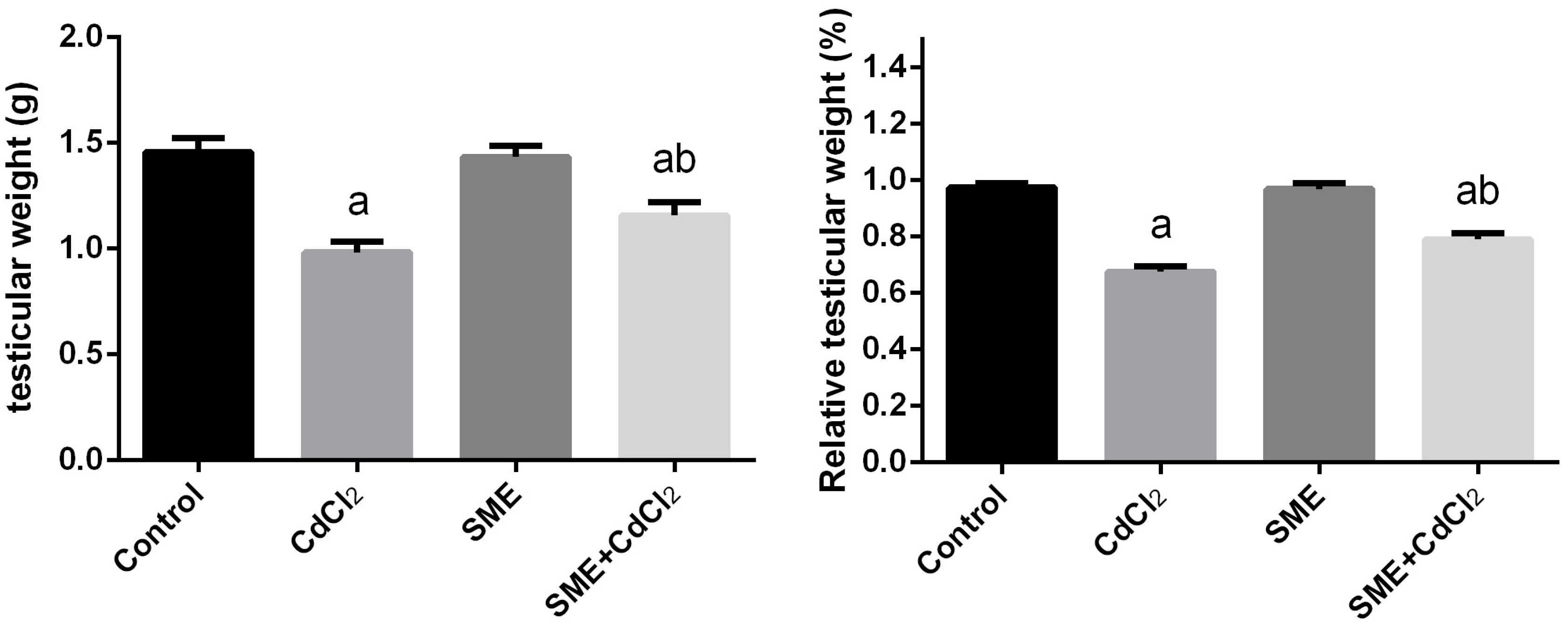
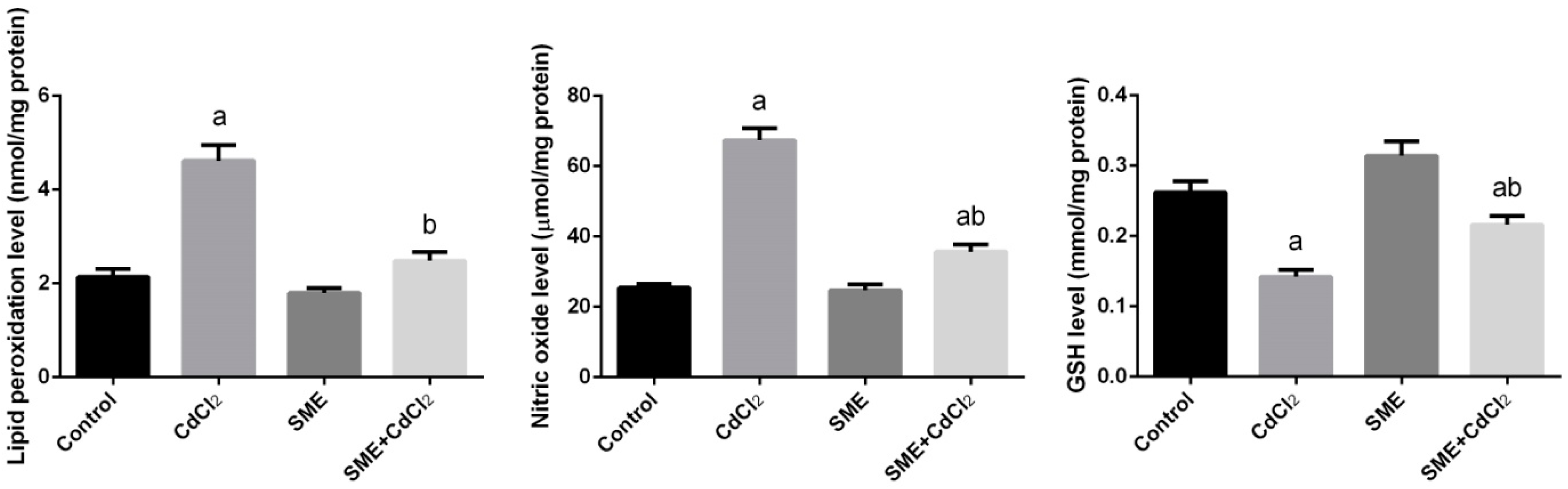
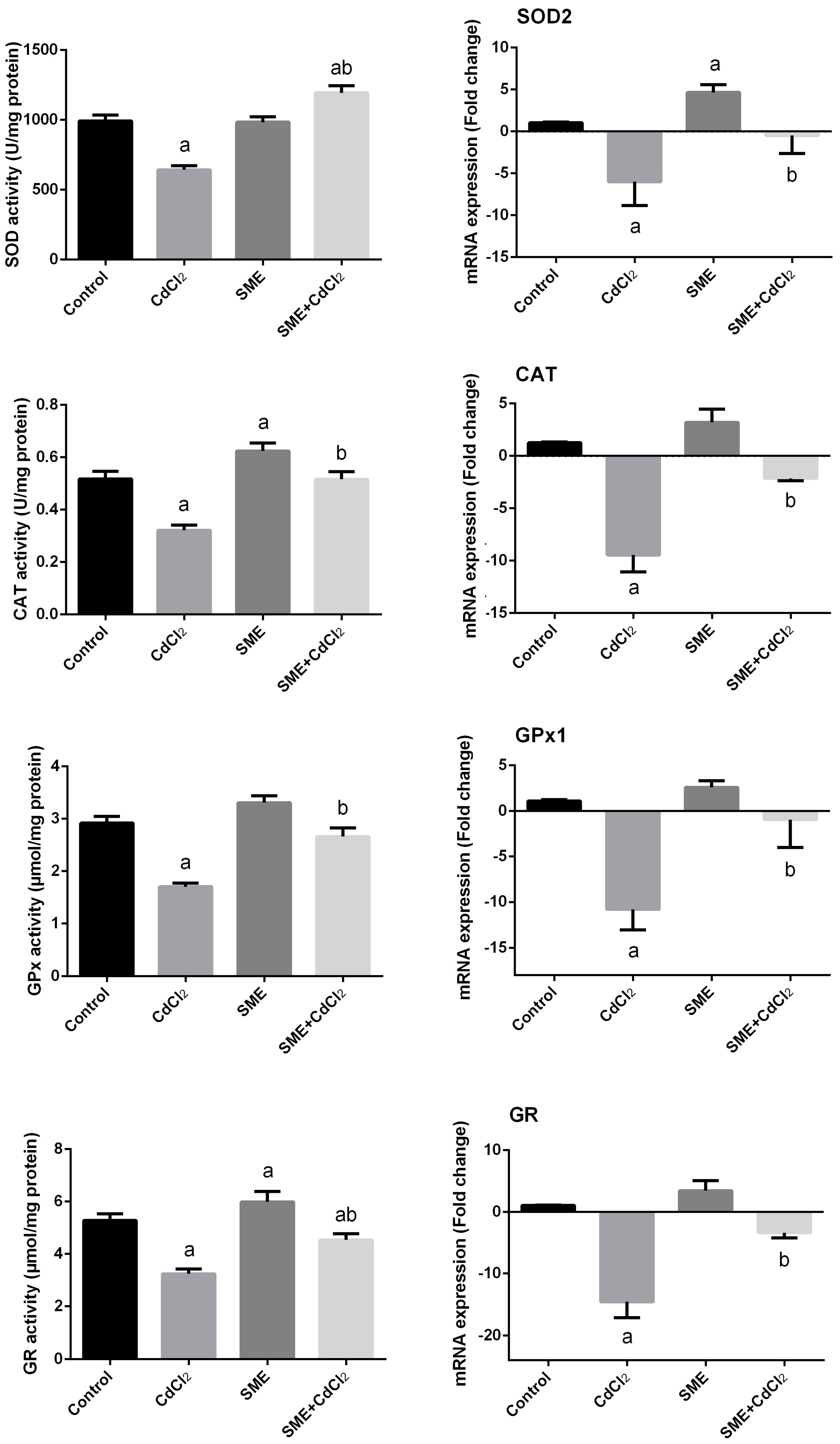
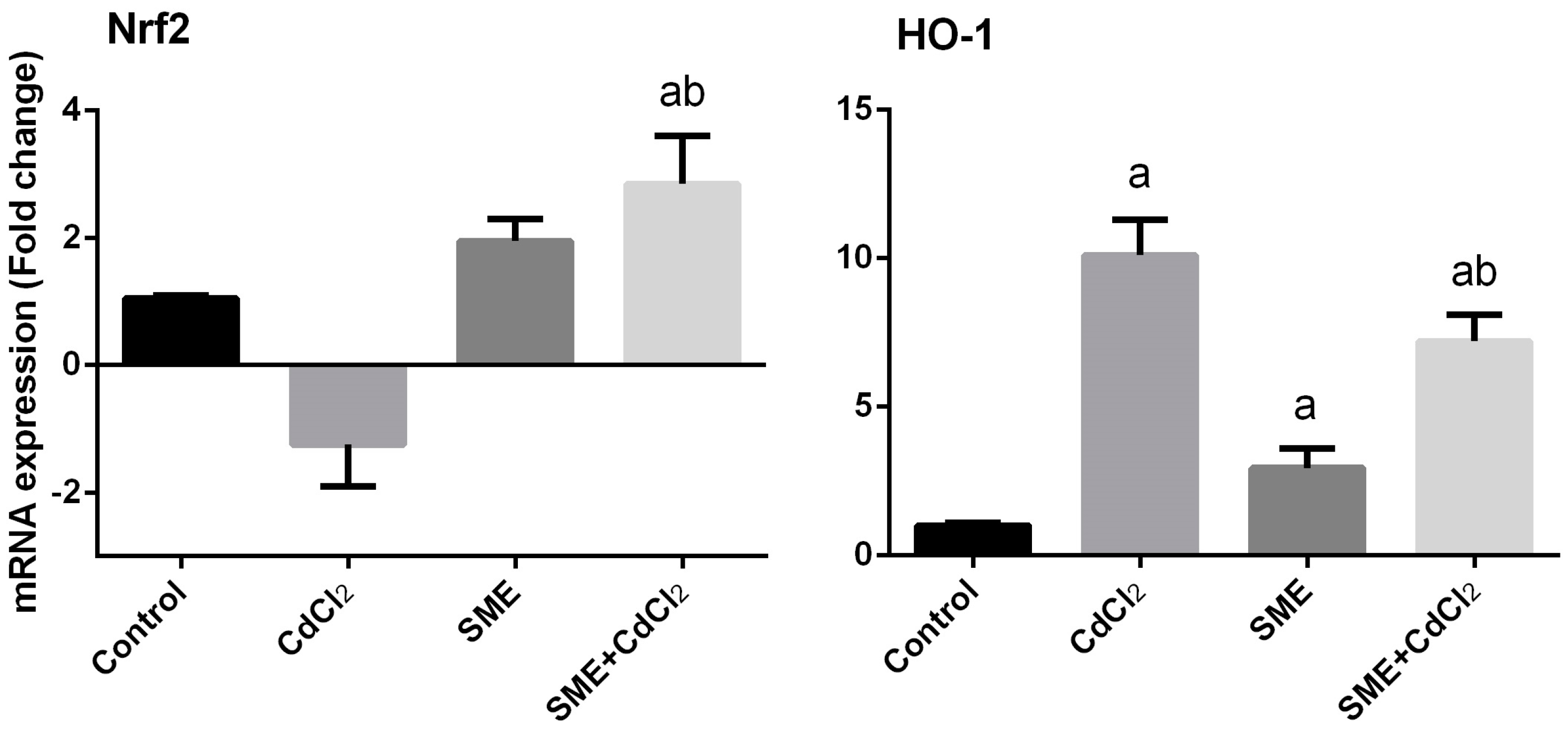
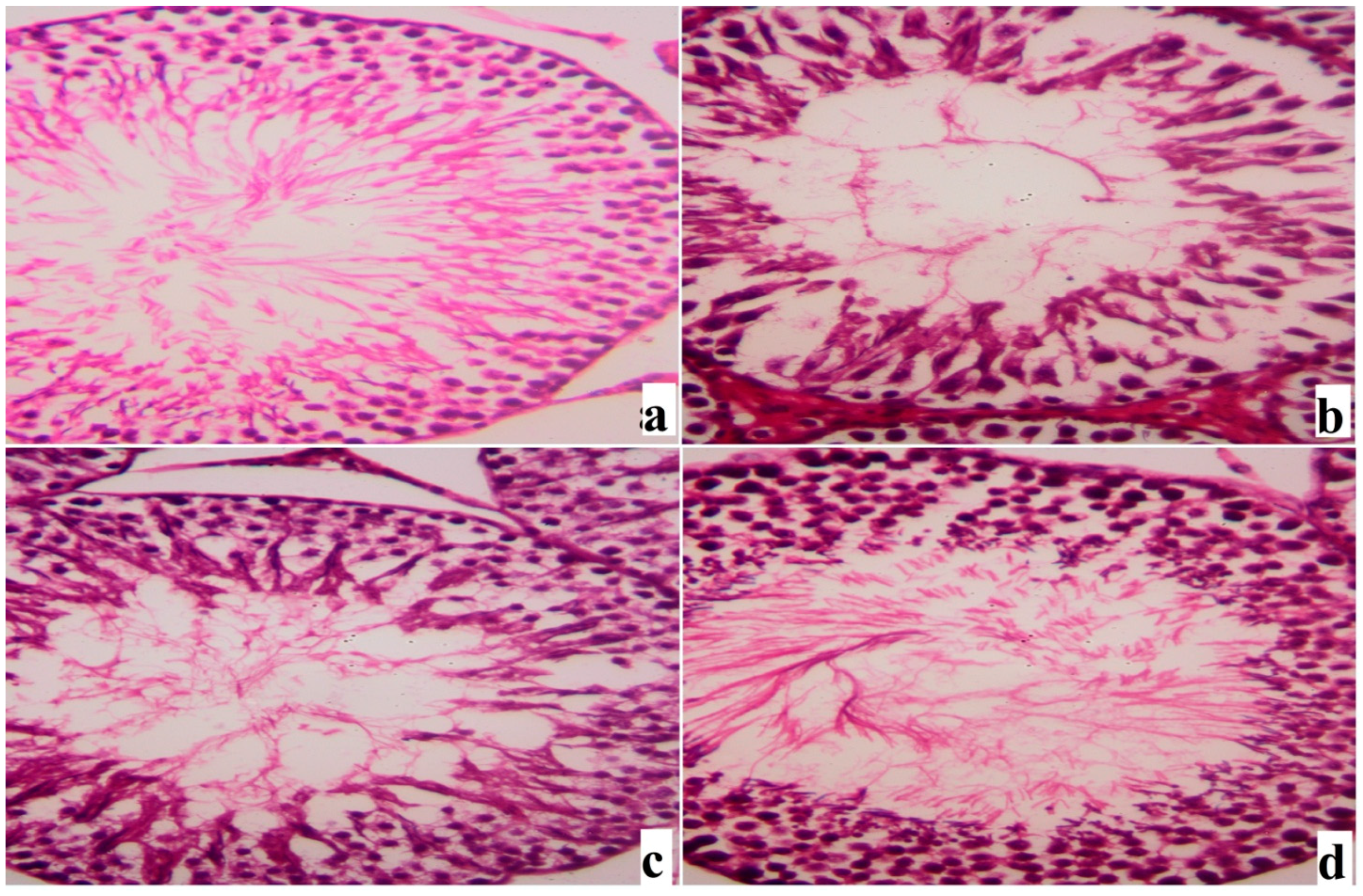
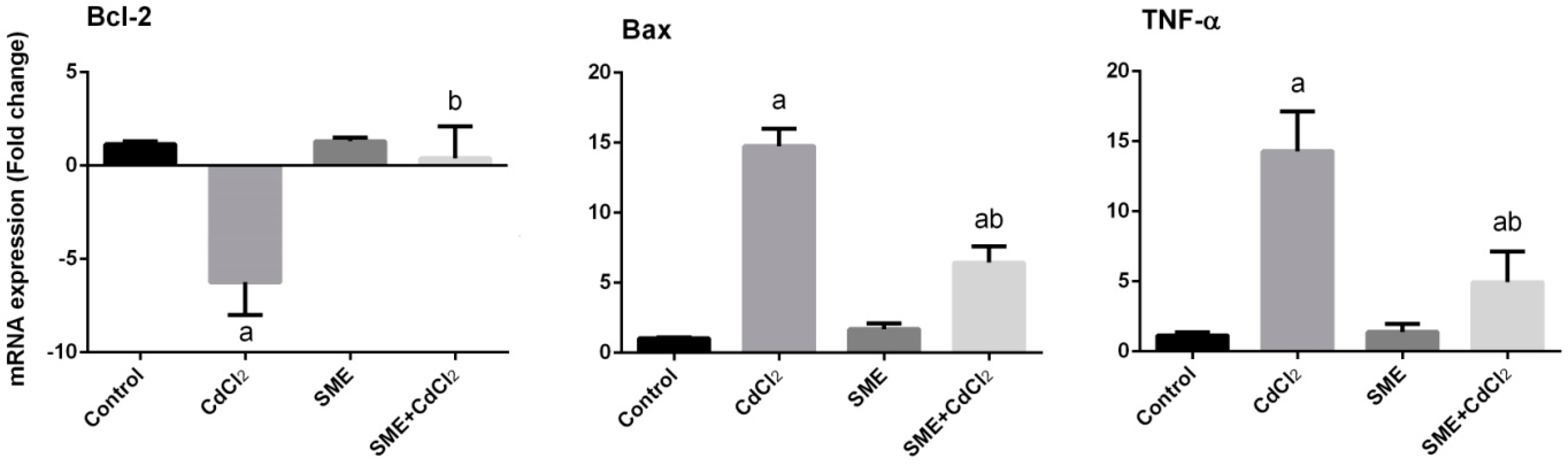
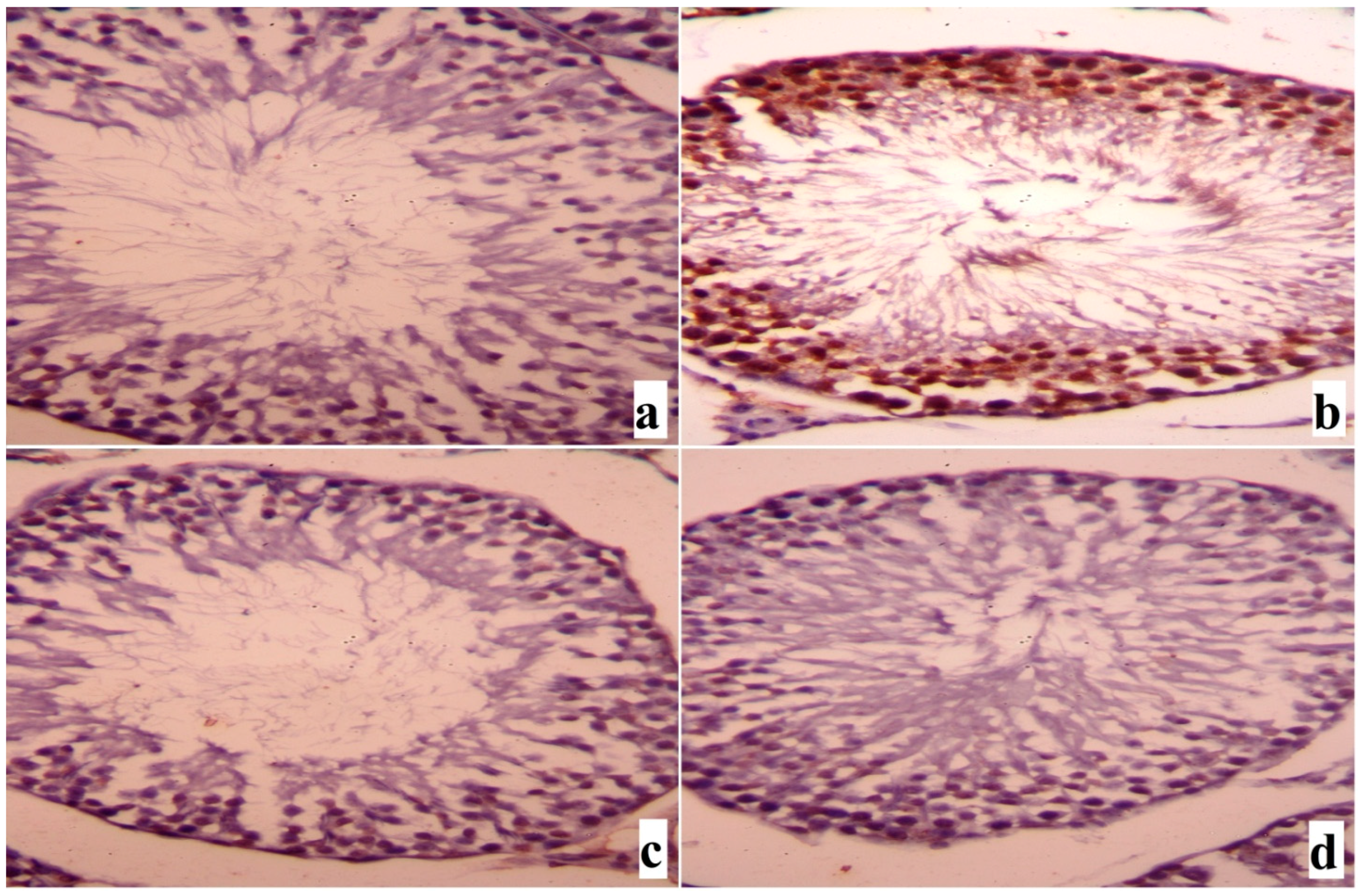
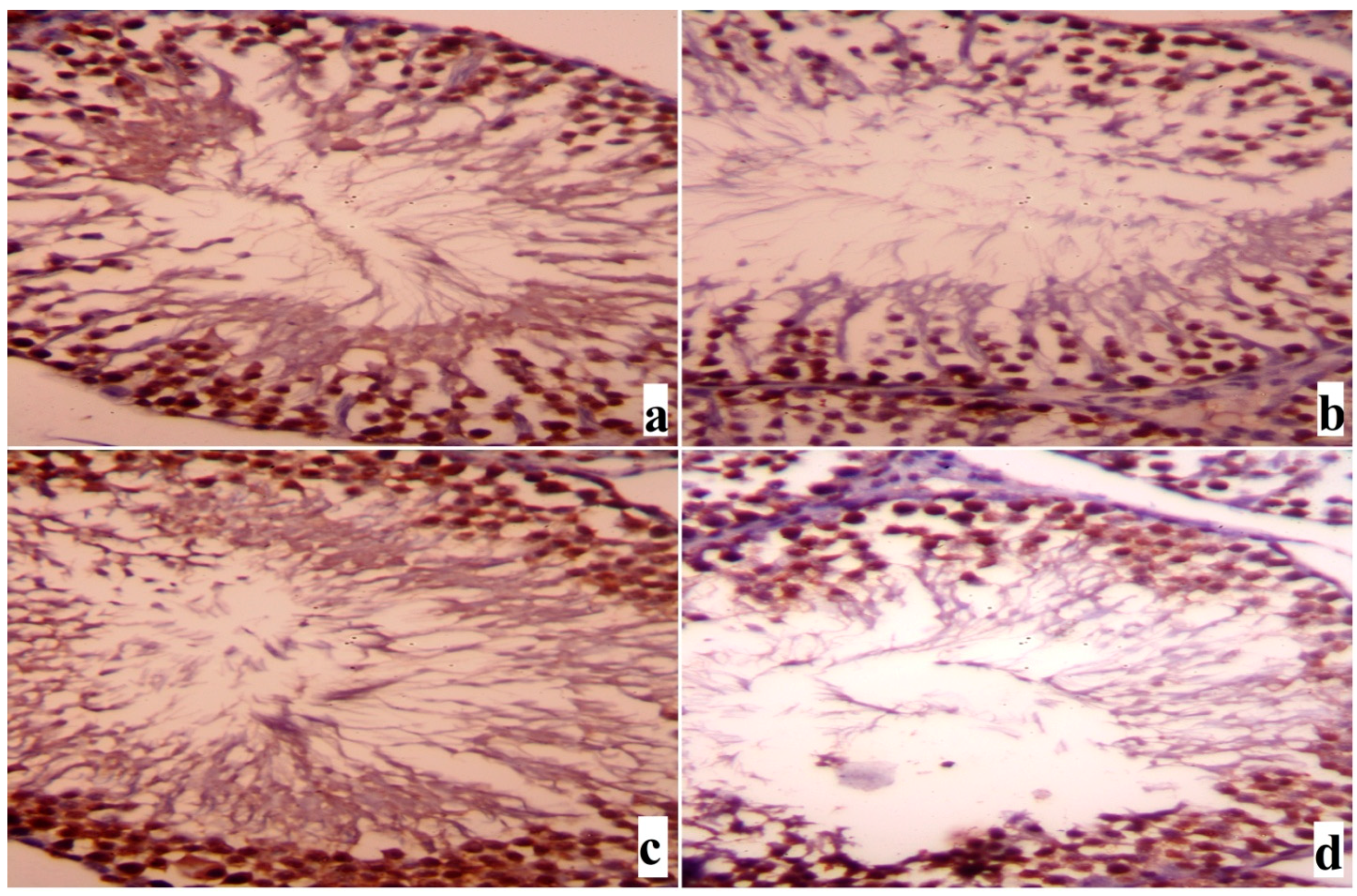
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Preparation of Strawberry Crude Extract
4.3. Animals and Experimental Design
4.4. Determination of Cadmium Concentration in Testes
4.5. Testicular Weight Measurements
4.6. Biochemical Analysis
4.6.1. Serum Testosterone
4.6.2. Lipid Peroxidation (LPO)
4.6.3. Nitric Oxide (NO) Level
4.6.4. Reduced Glutathione (GSH)
4.6.5. Catalase (CAT) Activity
4.6.6. Superoxide Dismutase (SOD) Activity
4.6.7. Glutathione Peroxidase (GPx) Activity
4.6.8. Glutathione Reductase (GR) Activity
4.7. Real Time-PCR
4.8. Histological Procedures
4.9. Immunohistochemistry Analysis
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nordberg, M.; Winblad, B.; Basun, H. Cadmium concentration in blood in an elderly urban population. Biometals 2000, 13, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Elkhadragy, M.F.; Abdel Moneim, A.E. Protective effect of fragaria ananassa methanolic extract on cadmium chloride (CDCL2)-induced hepatotoxicity in rats. Toxicol. Mech. Methods 2017, 27, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef]
- Sajjad, S.; Malik, H.; Farooq, U.; Rashid, F.; Nasim, H.; Tariq, S.; Rehman, S. Cadmium chloride toxicity revisited: Effect on certain andrological, endocrinological and biochemical parameters of adult male rabbits. Physiol. Res. 2014, 63, 505–512. [Google Scholar] [PubMed]
- Thompson, J.; Bannigan, J. Cadmium: Toxic effects on the reproductive system and the embryo. Reprod. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Ognjanovic, B.I.; Markovic, S.D.; Ethordevic, N.Z.; Trbojevic, I.S.; Stajn, A.S.; Saicic, Z.S. Cadmium-induced lipid peroxidation and changes in antioxidant defense system in the rat testes: Protective role of coenzyme Q 10 and vitamin E. Reprod. Toxicol. 2010, 29, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.F.; Arrieta, M.C.; Villafañe, M.N.; Klyver, S.M.R.; Odstrcil, I.M.A.; González, M.E. Early signs of toxicity in testes and sperm of rats exposed to low cadmium doses. Toxicol. Ind. Health 2017. [Google Scholar] [CrossRef]
- Ponnusamy, M.; Pari, L. Protective role of diallyl tetrasulfide on cadmium-induced testicular damage in adult rats: A biochemical and histological study. Toxicol. Ind. Health 2011, 27, 407–416. [Google Scholar] [CrossRef] [PubMed]
- De Souza Predes, F.; Diamante, M.A.; Dolder, H. Testis response to low doses of cadmium in wistar rats. Int. J. Exp. Pathol. 2010, 91, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tam, P.P.; Liu, W.K. Gonadal development and fertility of mice treated prenatally with cadmium during the early organogenesis stages. Teratology 1985, 32, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Luevano, J.; Damodaran, C. A review of molecular events of cadmium-induced carcinogenesis. J. Environ. Pathol. Toxicol. Oncol. 2014, 33, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Waisberg, M.; Joseph, P.; Hale, B.; Beyersmann, D. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology 2003, 192, 95–117. [Google Scholar] [CrossRef]
- Joseph, P. Mechanisms of cadmium carcinogenesis. Toxicol. Appl. Pharmacol. 2009, 238, 272–279. [Google Scholar] [CrossRef] [PubMed]
- El-Refaiy, A.I.; Eissa, F.I. Histopathology and cytotoxicity as biomarkers in treated rats with cadmium and some therapeutic agents. Saudi J. Biol. Sci. 2013, 20, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. Hplc-msn identification and quantification of flavonol glycosides in 28 wild and cultivated berry species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, J.; Dibarrart, A.J.; Fuentes-Fuentes, M.C.; Saez-Orellana, F.; Quinones, K.; Guzman, L.; Perez, C.; Becerra, J.; Aguayo, L.G. Synaptic failure and adenosine triphosphate imbalance induced by amyloid-beta aggregates are prevented by blueberry-enriched polyphenols extract. J. Neurosci. Res. 2011, 89, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Malin, D.H.; Lee, D.R.; Goyarzu, P.; Chang, Y.H.; Ennis, L.J.; Beckett, E.; Shukitt-Hale, B.; Joseph, J.A. Short-term blueberry-enriched diet prevents and reverses object recognition memory loss in aging rats. Nutrition 2011, 27, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.; Moyano, R.; Molina Lopez, A.M.; Blanco, C.; Flores-Acuna, R.; Garcia-Flores, J.R.; Espada, M.; Monterde, J.G. Preneoplastic and neoplastic changes in the leydig cells population in mice exposed to low doses of cadmium. Toxicol. Ind. Health 2010, 26, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Siu, E.R.; Mruk, D.D.; Porto, C.S.; Cheng, C.Y. Cadmium-induced testicular injury. Toxicol. Appl. Pharmacol. 2009, 238, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Iserhienrhien, B.O.; Badejo, T.A. Rutin- and selenium-attenuated cadmium-induced testicular pathophysiology in rats. Hum. Exp. Toxicol. 2013, 32, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Erboga, M.; Kanter, M.; Aktas, C.; Bozdemir Donmez, Y.; Fidanol Erboga, Z.; Aktas, E.; Gurel, A. Anti-apoptotic and anti-oxidant effects of caffeic acid phenethyl ester on cadmium-induced testicular toxicity in rats. Biol. Trace Elem. Res. 2016, 171, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Eybl, V.; Kotyzova, D.; Bludovska, M. The effect of curcumin on cadmium-induced oxidative damage and trace elements level in the liver of rats and mice. Toxicol. Lett. 2004, 151, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.S.; Alyhya, N.A.; Elkhadragy, M.F.; Al-Olayan, E.M.; Alagami, R.A.; Hassan, Z.K.; Baker, S.B.; Abdel Moneim, A.E. The protective properties of the strawberry (Fragaria ananassa) against carbon tetrachloride-induced hepatotoxicity in rats mediated by anti-apoptotic and upregulation of antioxidant genes expression effects. Front. Physiol. 2016, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, H.; Liu, X.; Liu, Z. N-acetylcysteine protects against cadmium-induced oxidative stress in rat hepatocytes. J. Vet. Sci. 2014, 15, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Shaikh, Z.A. Cadmium-induced apoptosis in rat kidney epithelial cells involves decrease in nuclear factor-kappa b activity. Toxicol. Sci. 2006, 91, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Abdel Moneim, A.E. Indigofera oblongifolia prevents lead acetate-induced hepatotoxicity, oxidative stress, fibrosis and apoptosis in rats. PLoS ONE 2016, 11, e0158965. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar] [PubMed]
- Choi, A.M.; Alam, J. Heme oxygenase-1: Function, regulation, and implication of a novel stress-inducible protein in oxidant-induced lung injury. Am. J. Respir. Cell Mol. Biol. 1996, 15, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Al-Azemi, M.; Omu, F.E.; Kehinde, E.O.; Anim, J.T.; Oriowo, M.A.; Omu, A.E. Lithium protects against toxic effects of cadmium in the rat testes. J. Assist. Reprod. Genet. 2010, 27, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Shivji, K.K.; Kenny, M.K.; Wood, R.D. Proliferating cell nuclear antigen is required for DNA excision repair. Cell 1992, 69, 367–374. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Dkhil, M.A.; Al-Quraishy, S.; Diab, M.M.; Othman, M.S.; Aref, A.M.; Abdel Moneim, A.E. The potential protective role of physalis peruviana l. Fruit in cadmium-induced hepatotoxicity and nephrotoxicity. Food Chem. Toxicol. 2014, 74, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P.; Street, J.C.; Shupe, J.L.; Bourcier, D.R. Accumulation and depletion of cadmium and lead in tissues and milk of lactating cows fed small amounts of these metals. J. Dairy Sci. 1982, 65, 972–979. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Sastry, K.V.; Moudgal, R.P.; Mohan, J.; Tyagi, J.S.; Rao, G.S. Spectrophotometric determination of serum nitrite and nitrate by copper-cadmium alloy. Anal. Biochem. 2002, 306, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. 1976. Biochem. Biophys. Res. Commun. 2012, 425, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.G.; Puebla, M.; Acevedo, A.; Tapia, P.J.; Gutierrez, E.; Zepeda, A.; Calaf, G.; Juantok, C.; Reyes, J.G. Oxidative stress in rat testis and epididymis under intermittent hypobaric hypoxia: Protective role of ascorbate supplementation. J. Androl. 2010, 31, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Accession Number | Sense (5′–3′) | Antisense (5′–3′) |
|---|---|---|---|
| GAPDH | NM_017008.4 | GCATCTTCTTGTGCAGTGCC | GATGGTGATGGGTTTCCCGT |
| SOD2 | NM_001270850.1 | AGCTGCACCACAGCAAGCAC | TCCACCACCCTTAGGGCTCA |
| CAT | NM_012520.2 | TCCGGGATCTTTTTAACGCCATTG | TCGAGCACGGTAGGGACAGTTCAC |
| GPX1 | NM_017006.2 | CGGTTTCCCGTGCAATCAGT | ACACCGGGGACCAAATGATG |
| GR | NM_053906.2 | TGCACTTCCCGGTAGGAAAC | GATCGCAACTGGGGTGAGAA |
| NFE2L2 | NM_031789.2 | GGTTGCCCACATTCCCAAAC | GGCTGGGAATATCCAGGGC |
| HMOX1 | NM_012580.2 | GCGAAACAAGCAGAACCCA | GCTCAGGATGAGTACCTCCCA |
| BCL2 | NM_016993.1 | CTGGTGGACAACATCGCTCTG | GGTCTGCTGACCTCACTTGTG |
| BAX | NM_017059.2 | GGCGAATTGGCGATGAACTG | ATGGTTCTGATCAGCTCGGG |
| TNFA | XM_008772775.2 | AGAACTCAGCGAGGACACCAA | GCTTGGTGGTTTGCTACGAC |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmallah, M.I.Y.; Elkhadragy, M.F.; Al-Olayan, E.M.; Abdel Moneim, A.E. Protective Effect of Fragaria ananassa Crude Extract on Cadmium-Induced Lipid Peroxidation, Antioxidant Enzymes Suppression, and Apoptosis in Rat Testes. Int. J. Mol. Sci. 2017, 18, 957. https://doi.org/10.3390/ijms18050957
Elmallah MIY, Elkhadragy MF, Al-Olayan EM, Abdel Moneim AE. Protective Effect of Fragaria ananassa Crude Extract on Cadmium-Induced Lipid Peroxidation, Antioxidant Enzymes Suppression, and Apoptosis in Rat Testes. International Journal of Molecular Sciences. 2017; 18(5):957. https://doi.org/10.3390/ijms18050957
Chicago/Turabian StyleElmallah, Mohammed I. Y., Manal F. Elkhadragy, Ebtesam M. Al-Olayan, and Ahmed E. Abdel Moneim. 2017. "Protective Effect of Fragaria ananassa Crude Extract on Cadmium-Induced Lipid Peroxidation, Antioxidant Enzymes Suppression, and Apoptosis in Rat Testes" International Journal of Molecular Sciences 18, no. 5: 957. https://doi.org/10.3390/ijms18050957
APA StyleElmallah, M. I. Y., Elkhadragy, M. F., Al-Olayan, E. M., & Abdel Moneim, A. E. (2017). Protective Effect of Fragaria ananassa Crude Extract on Cadmium-Induced Lipid Peroxidation, Antioxidant Enzymes Suppression, and Apoptosis in Rat Testes. International Journal of Molecular Sciences, 18(5), 957. https://doi.org/10.3390/ijms18050957