
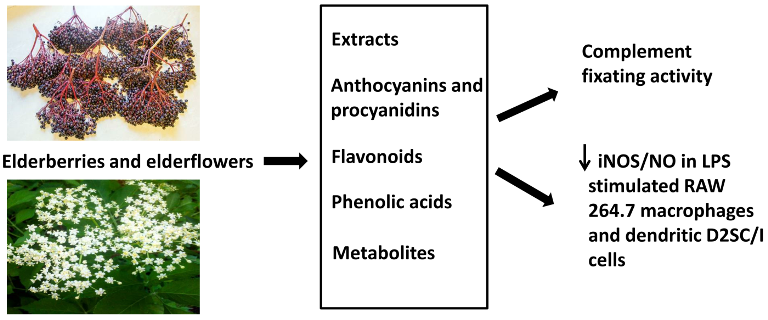
Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells


Abstract
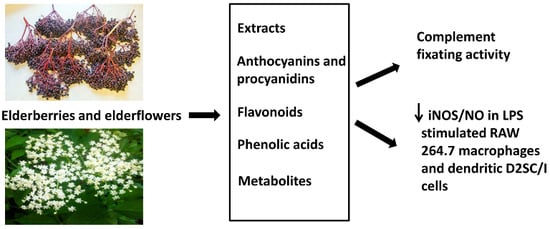
:
1. Introduction
2. Results and Discussion
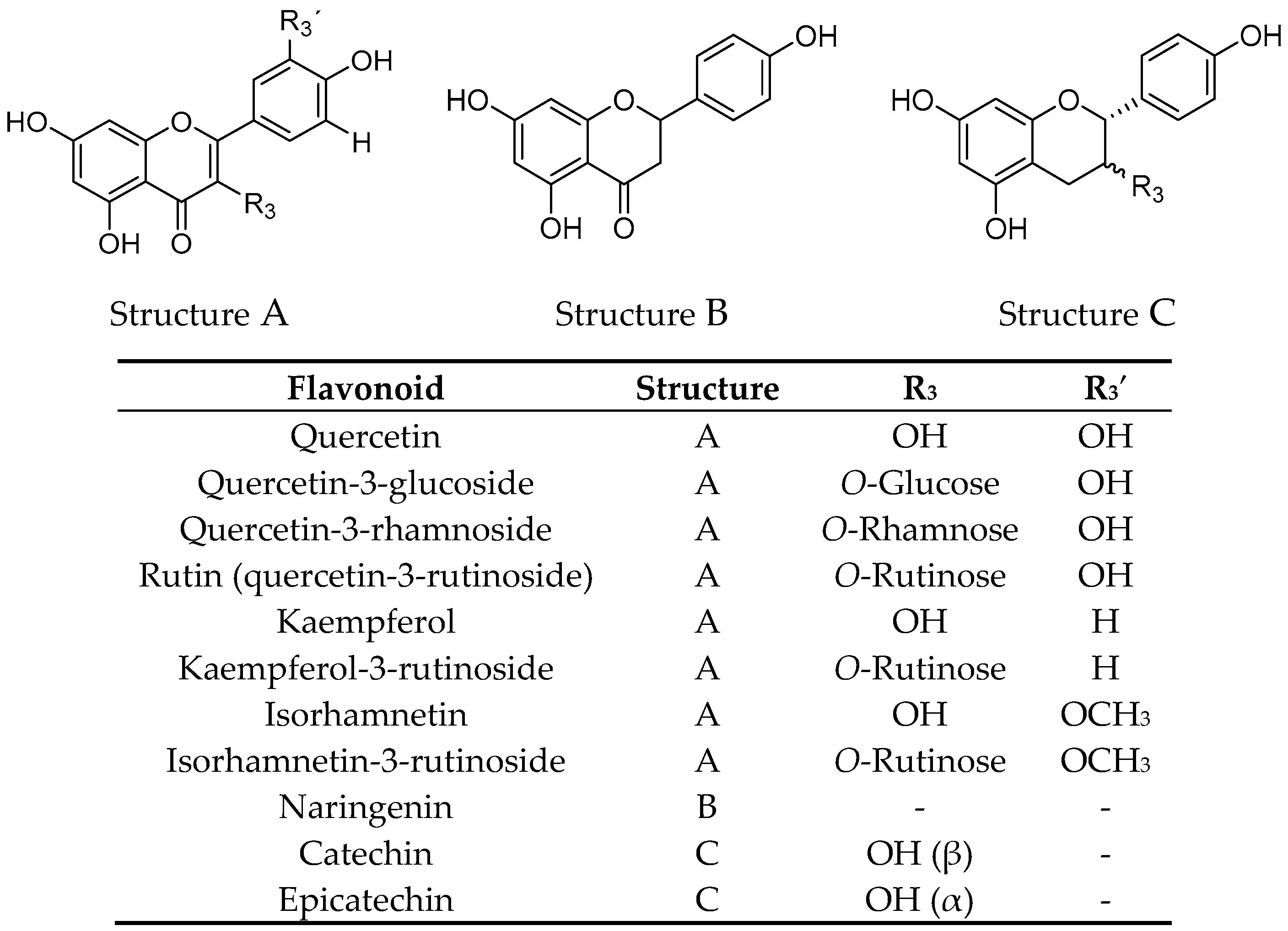
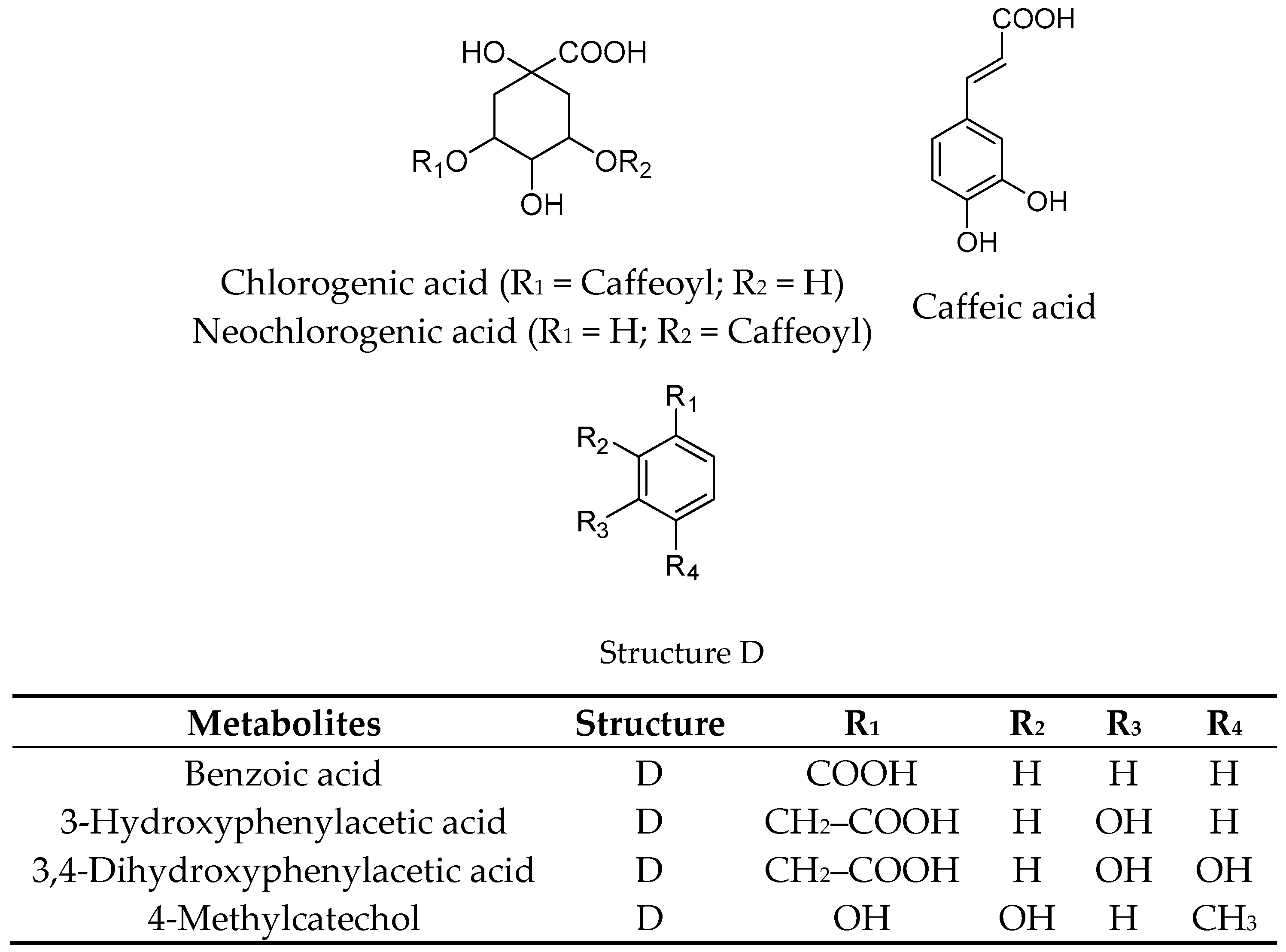
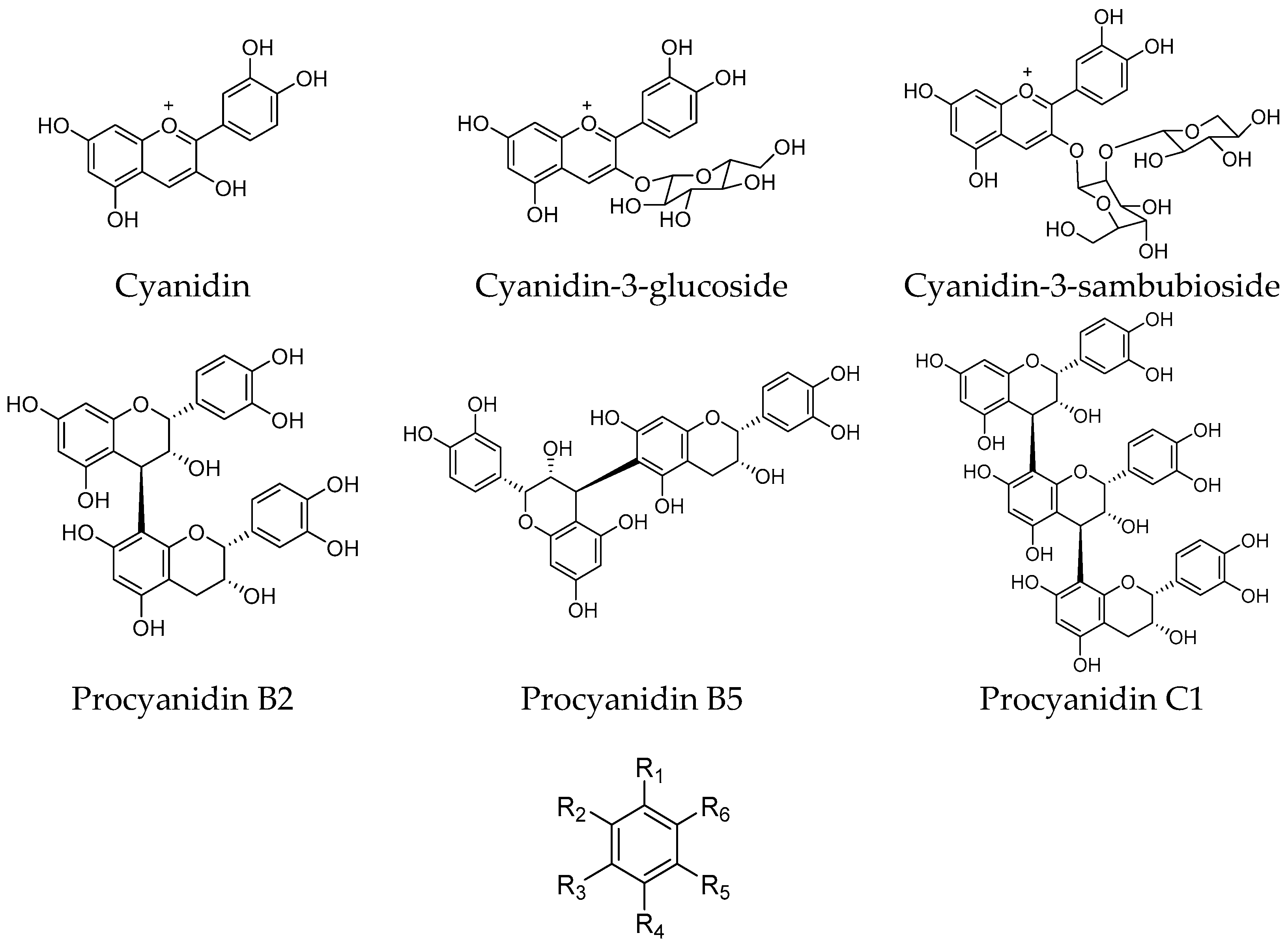
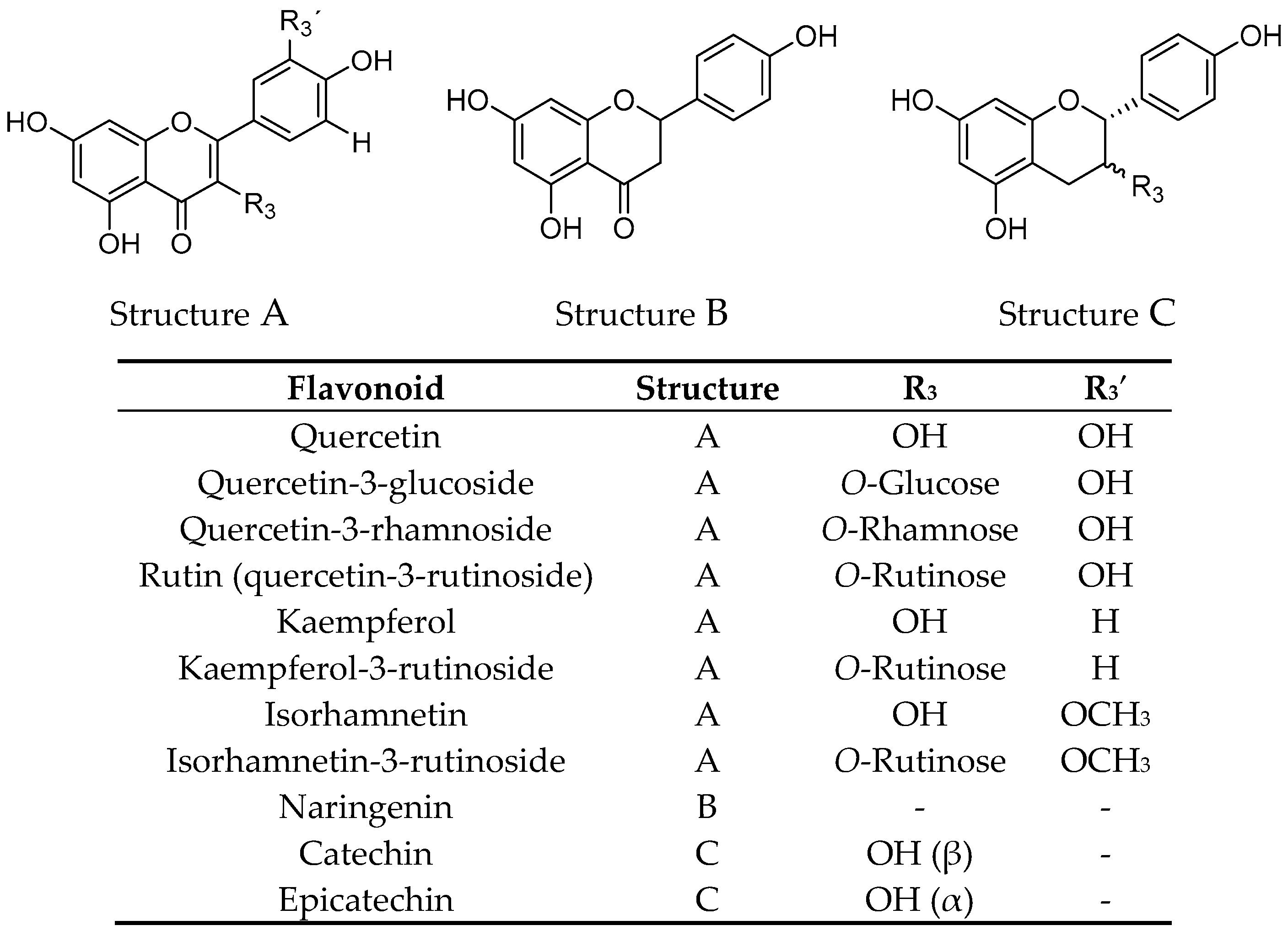
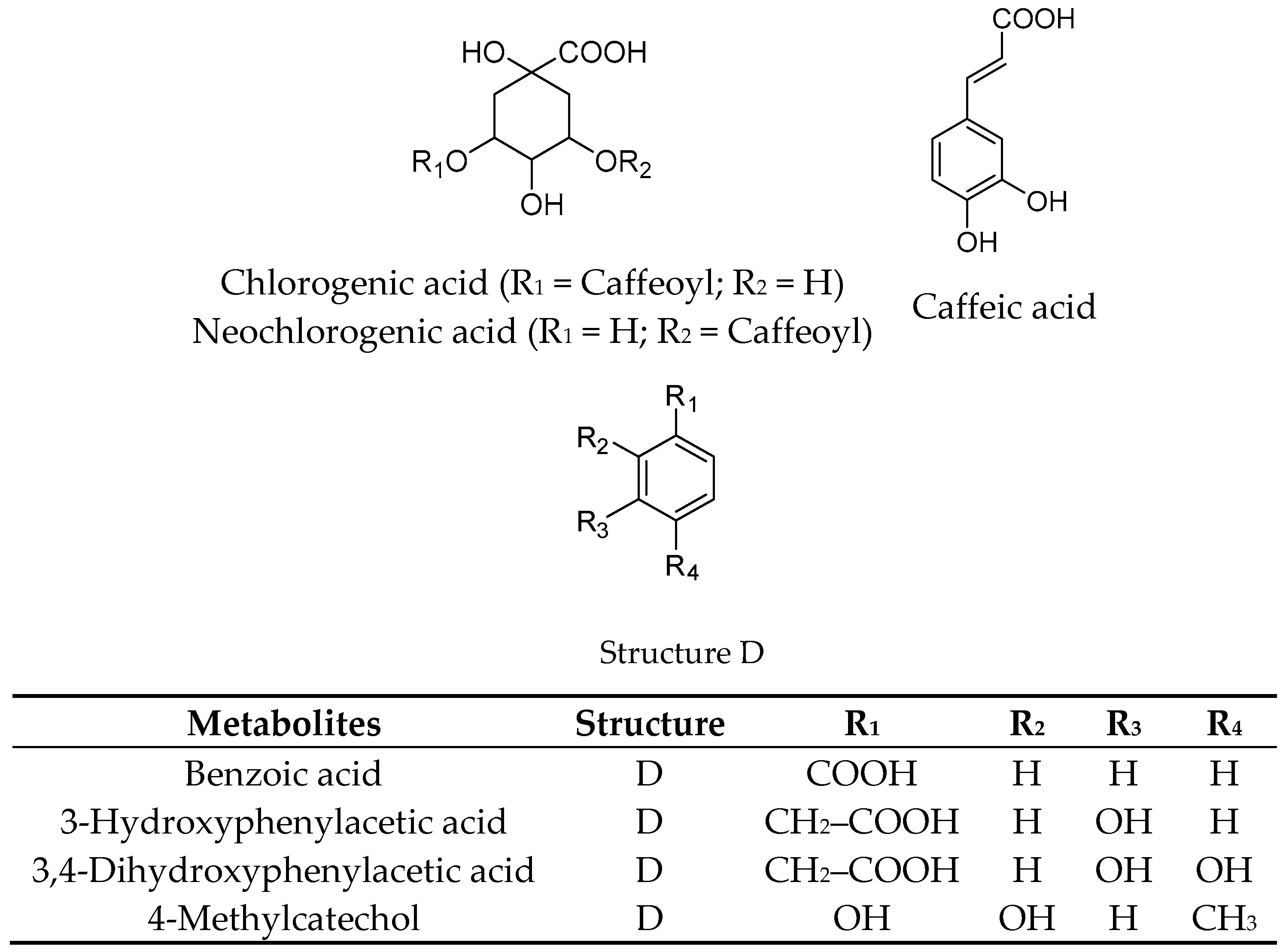
2.1. Extraction, Fractionation, and Chemical Characterization
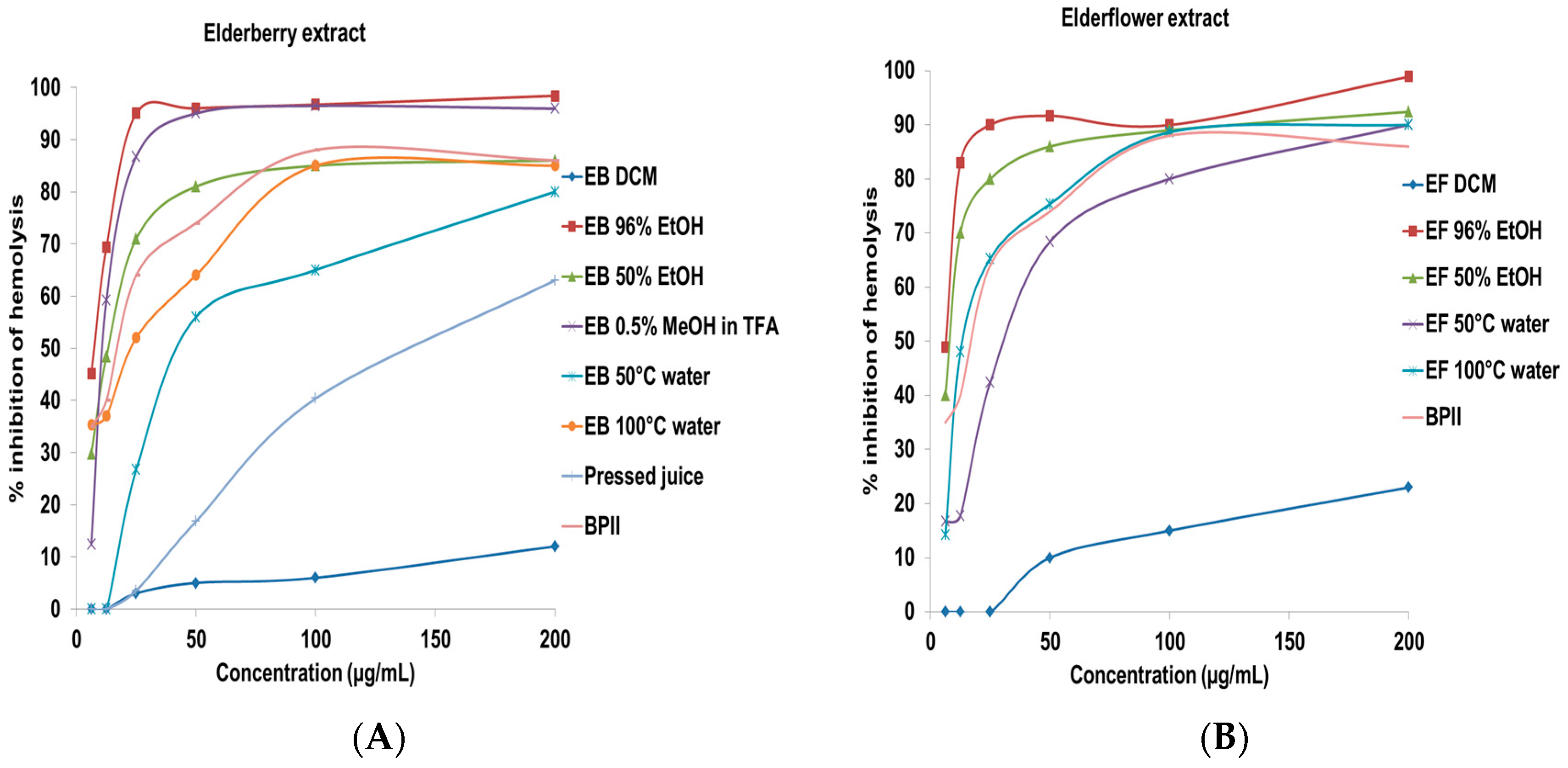
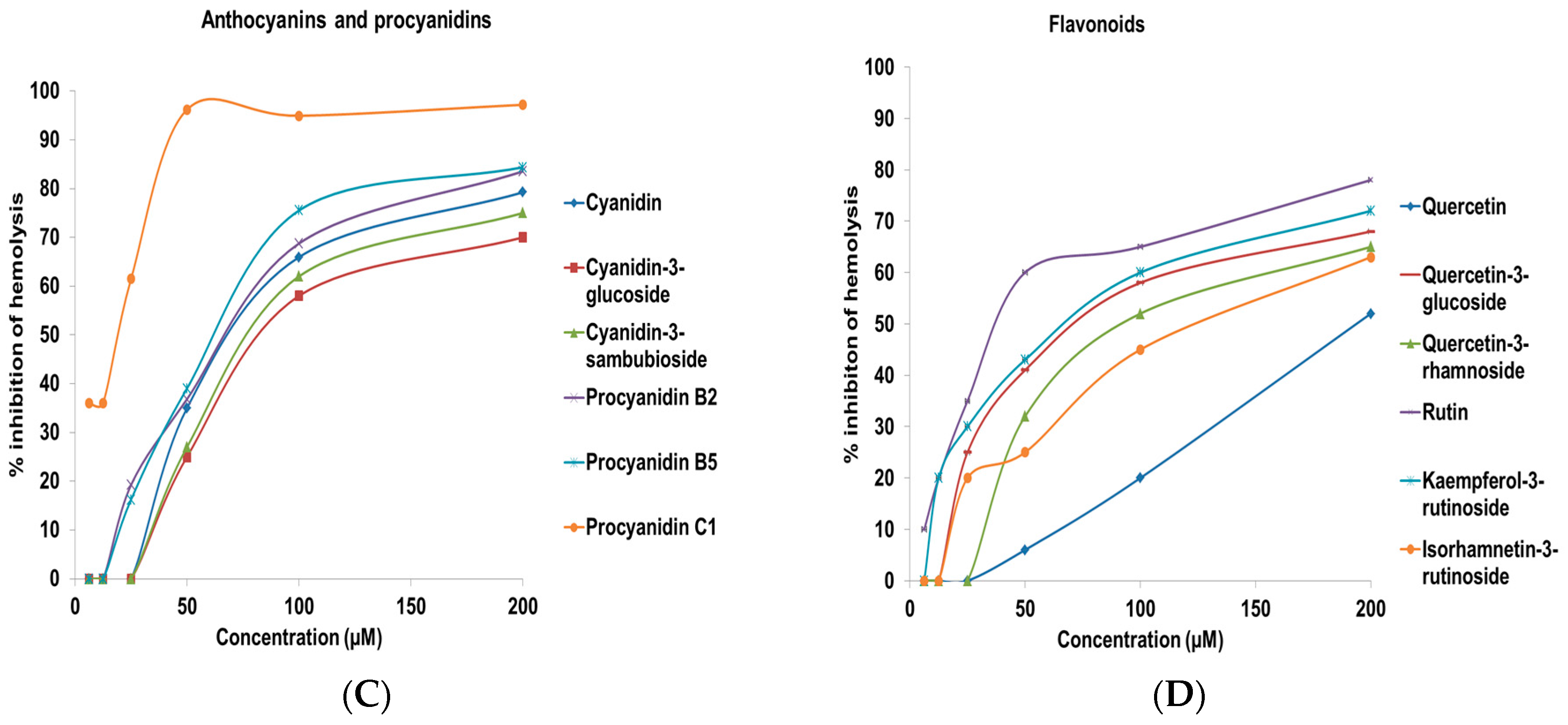
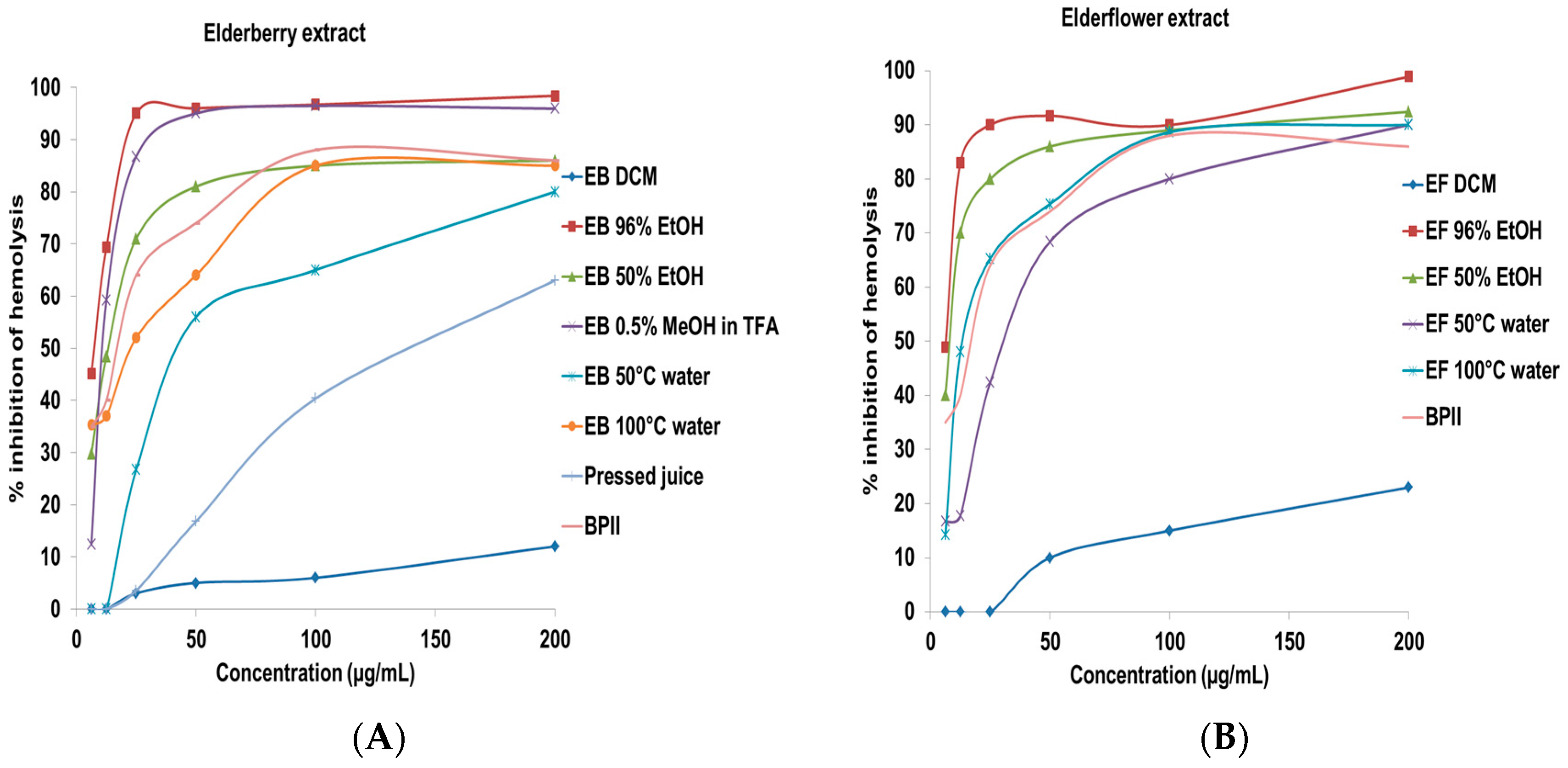
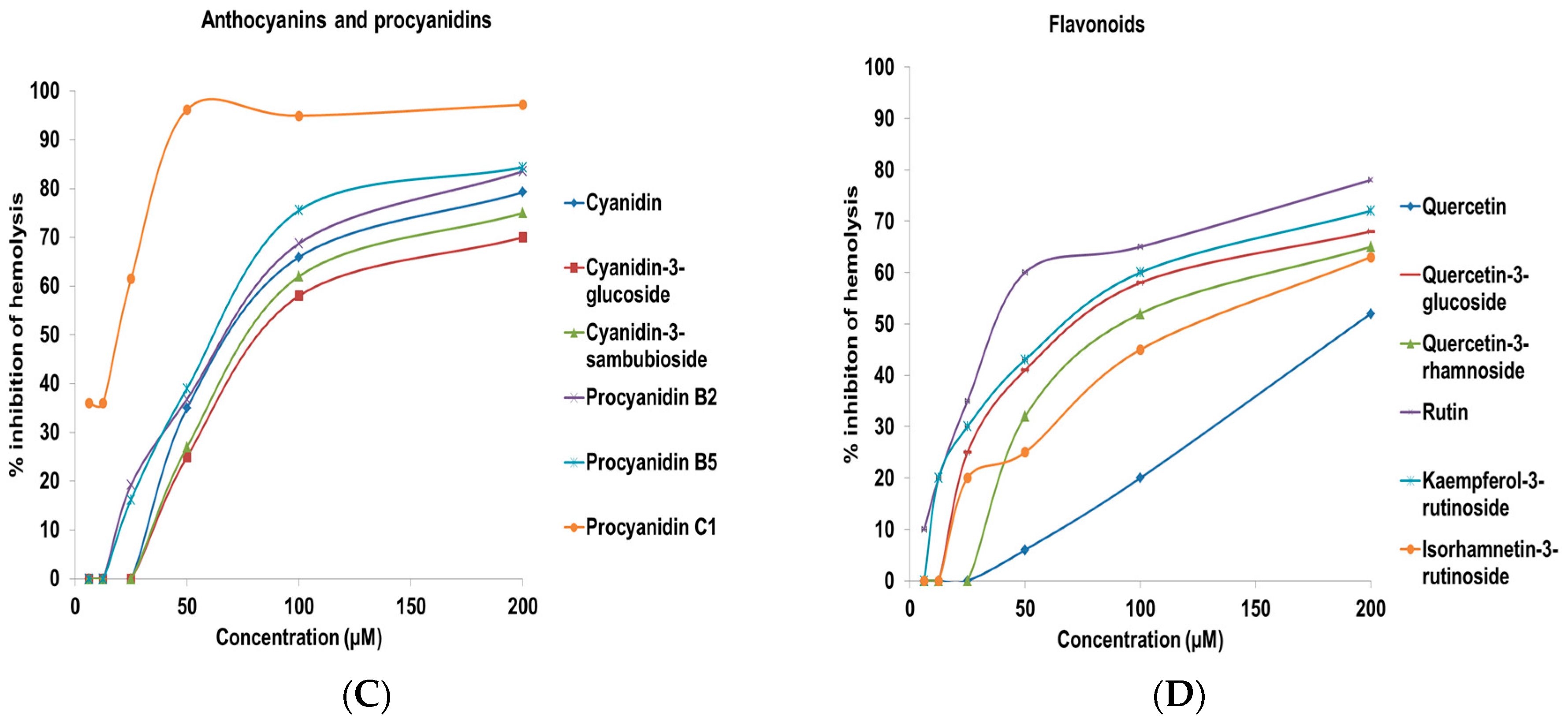
2.2. Complement Fixating Activity
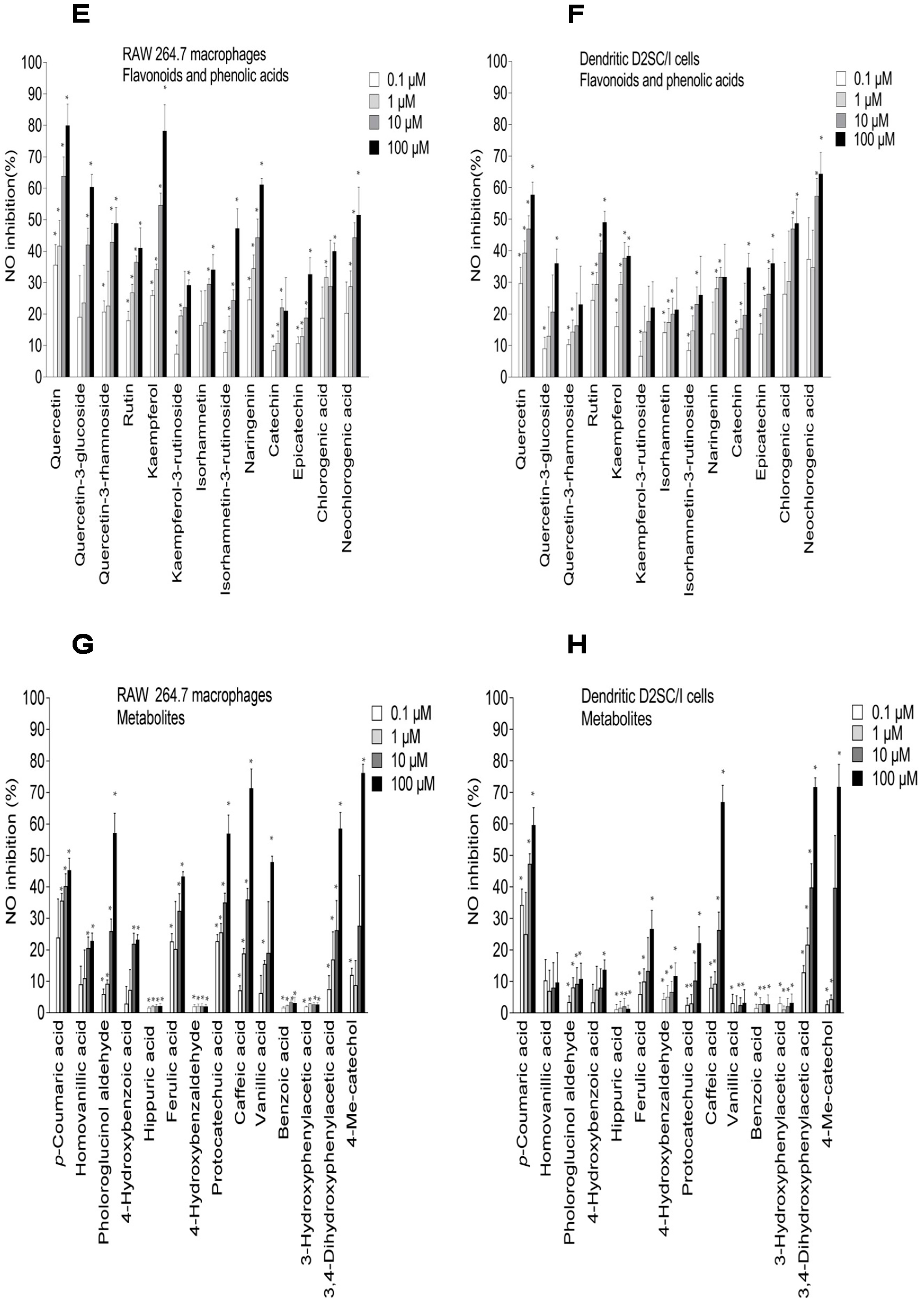
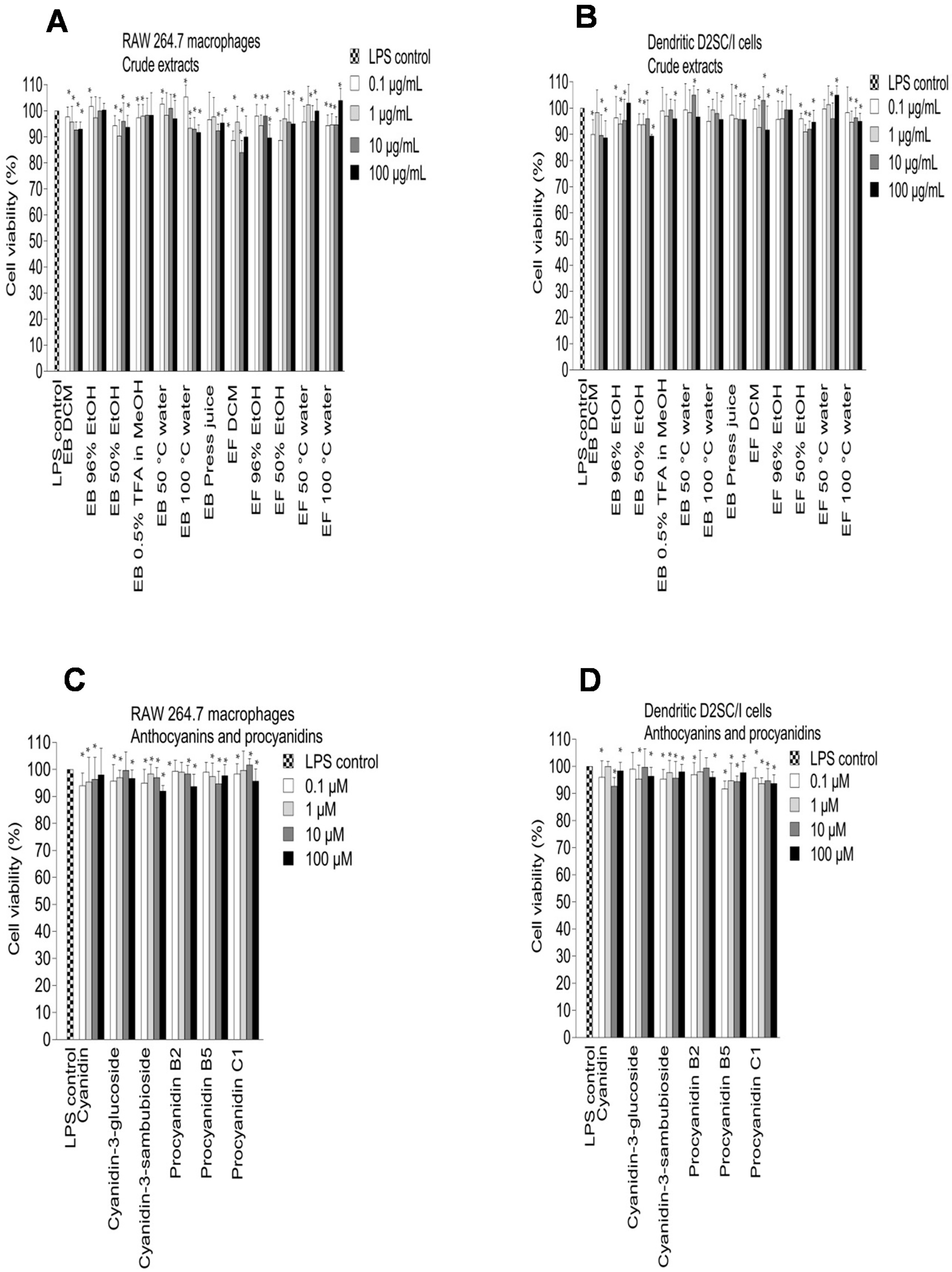
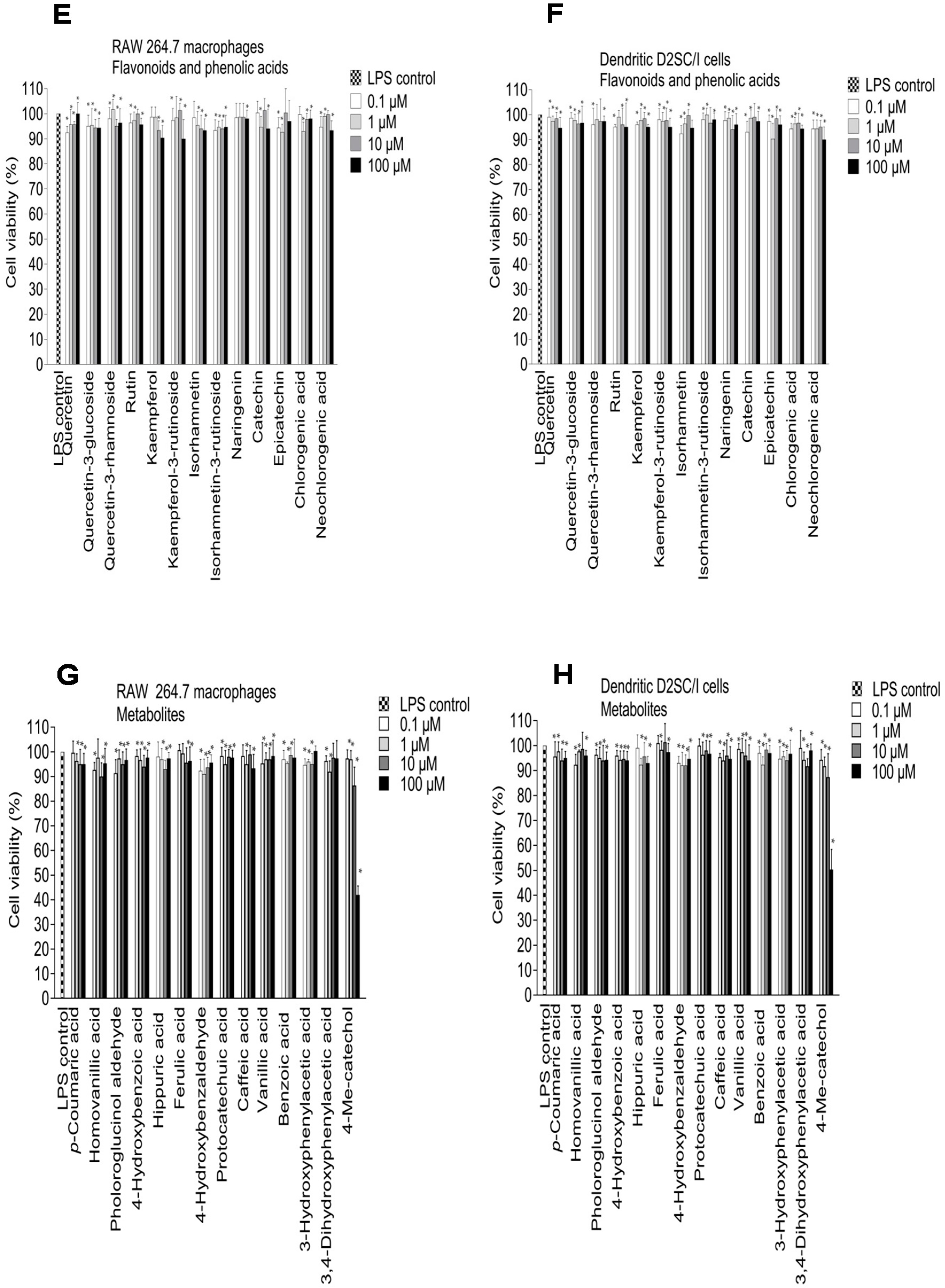
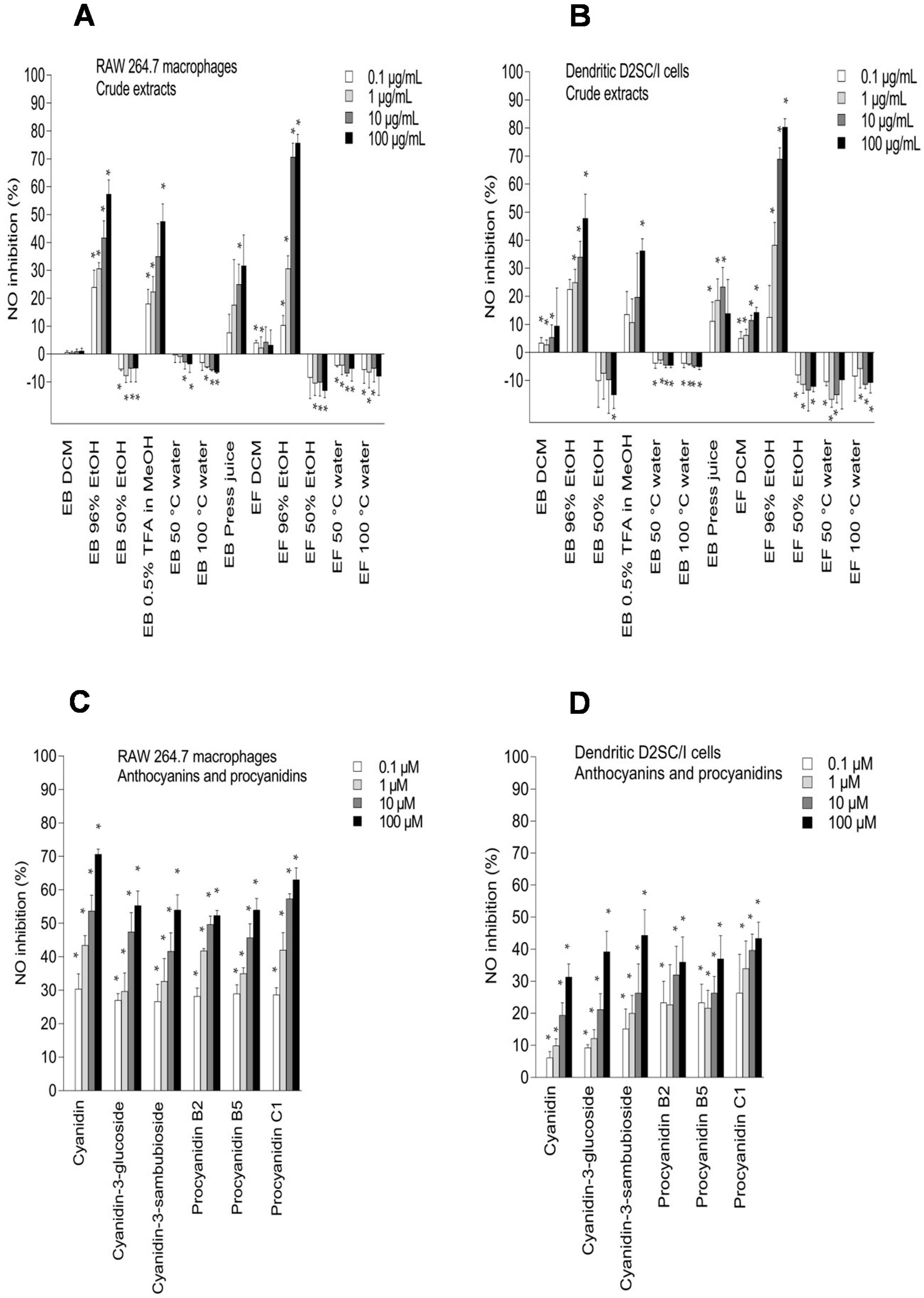
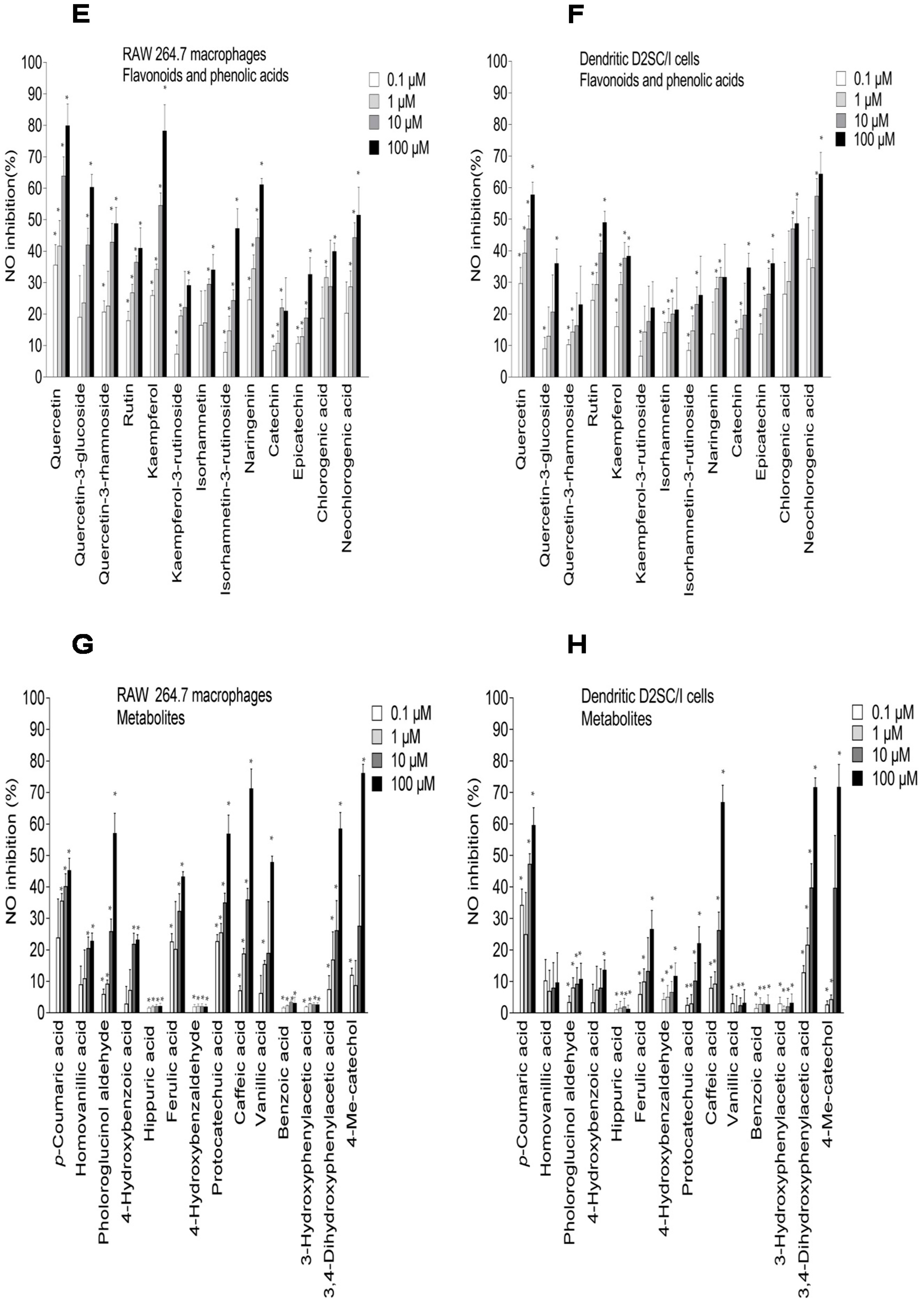
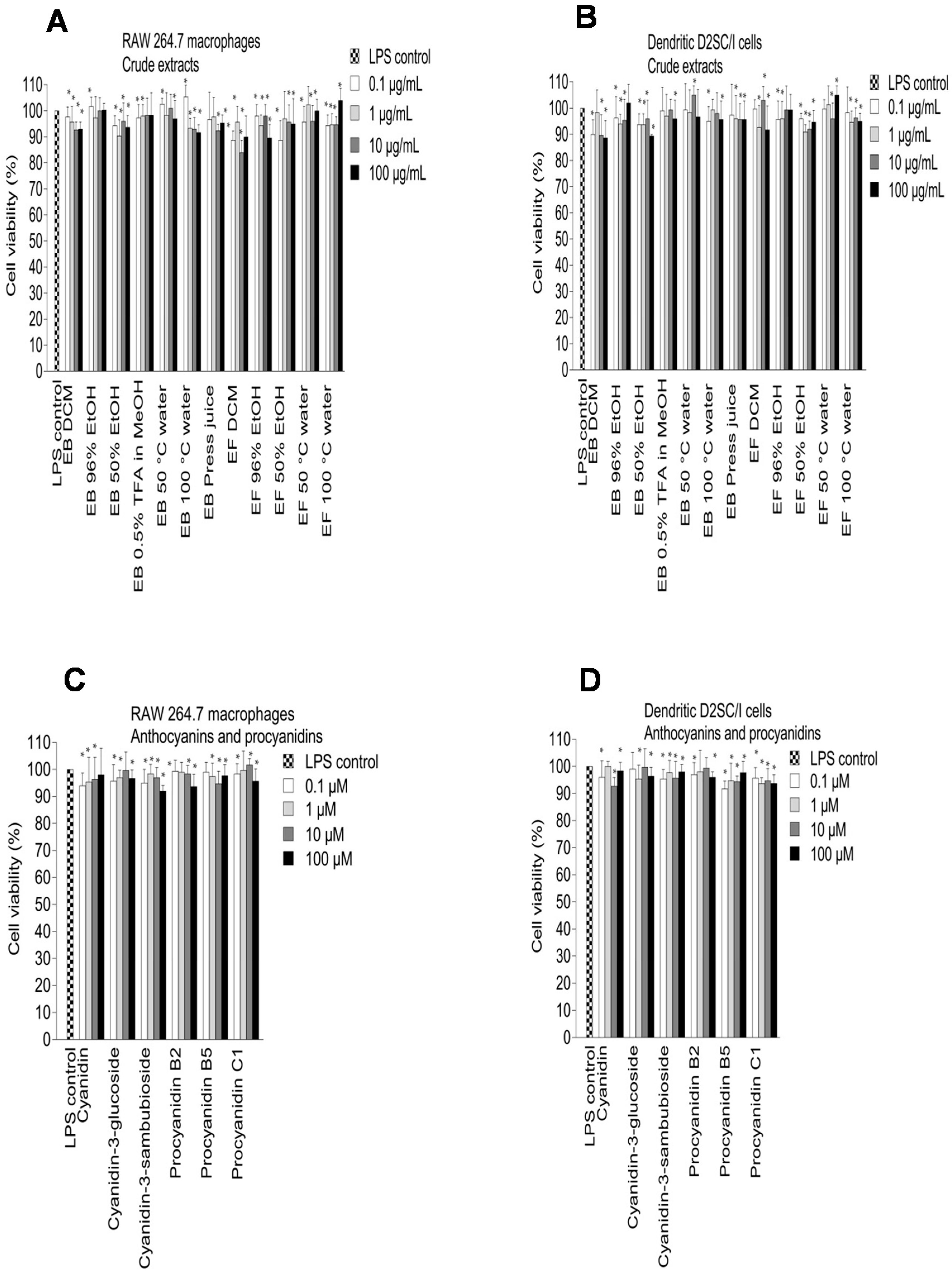
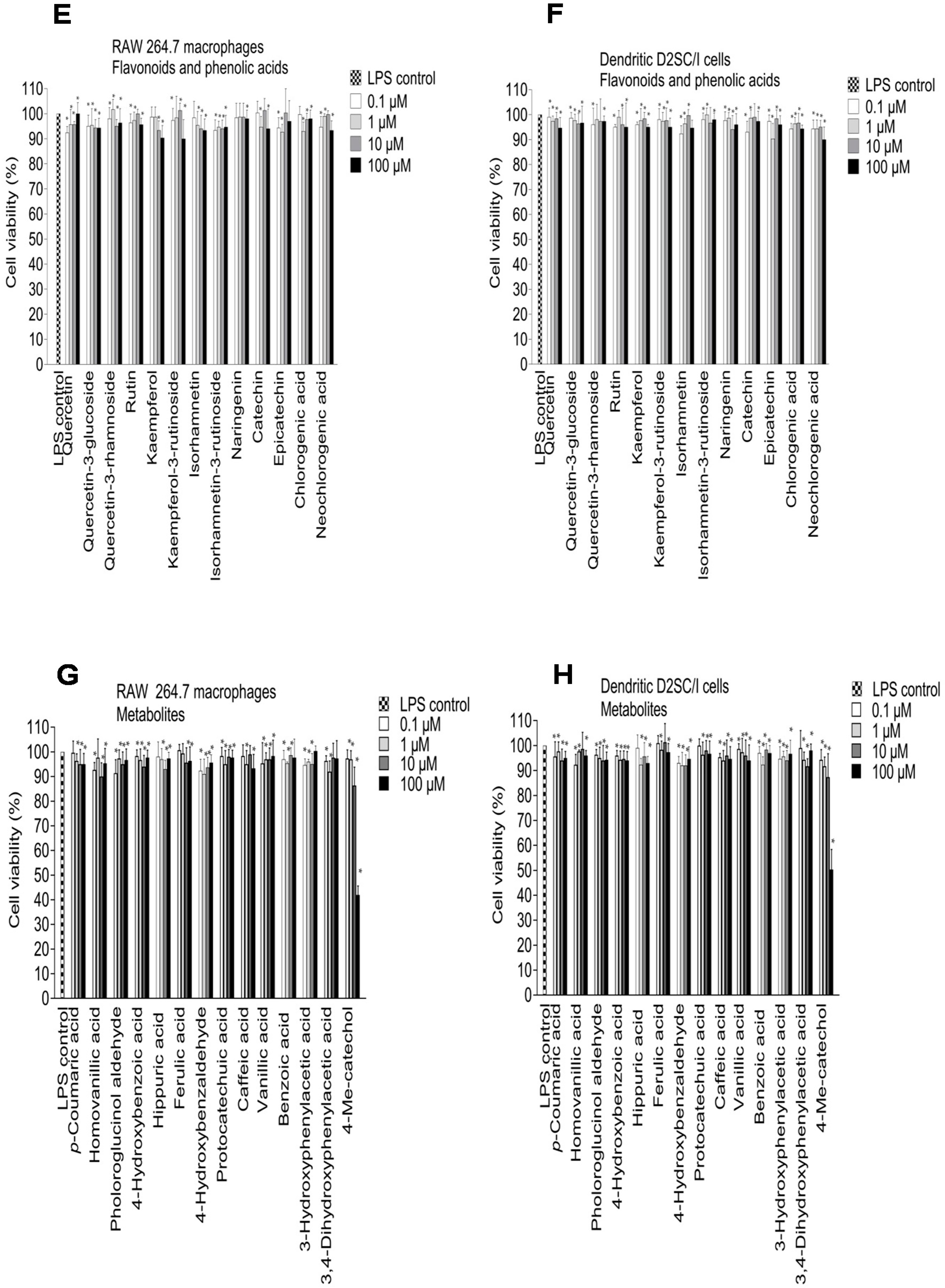
2.3. Inhibition of NO Production in LPS-Stimulated RAW 264.7 Macrophages and Dendritic D2SC/I Cells, and Cell Viability
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals
3.3. Extraction and Isolation of Anthocyanins
3.4. NMR Spectroscopy
3.5. Complement Fixating Assay
3.6. RAW 264.7 and Dendritic D2SC/I Cells
3.7. Nitrite Assay
3.8. Cell Viability Assay
3.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DCM | Dichloromethane |
| DMSO | Dimethylsulfoxide |
| EB | Elderberry |
| EF | Elderflower |
| EtOH IC50 | Ethanol Inhibitory concentration to give 50% effect |
| LPS | Lipopolysaccharide |
| MeOH | Methanol |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NO | Nitric oxide |
| NMR | Nuclear magnetic resonance |
| TMS | Tetramethylsilane |
| TFA | Trifluoroacetic acid |
References
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.M.; Vupputuri, S.; Myers, L.; Whelton, P.K. Fruit and vegetable intake and risk of cardiovascular disease in US adults: The first national health and nutrition examination survey epidemiologic follow-up study. Am. J. Clin. Nutr. 2002, 76, 93–99. [Google Scholar] [PubMed]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007. [Google Scholar] [CrossRef]
- Gandini, S.; Merzenich, H.; Robertson, C.; Boyle, P. Meta-analysis of studies on breast cancer risk and diet: The role of fruit and vegetable consumption and the intake of associated micronutrients. Eur. J. Cancer 2000, 36, 636–646. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Baek, S.J. Molecular targets of dietary polyphenols with anti-inflammatory properties. Yonsei Med. J. 2005, 46, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.D.; Almeida, M.C.; Lopes, N.P.; De Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of the natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, H.; Kiderlen, A.F. Antileishmanial activity and immune modulatory effects of tannins and related compounds on Leishmania parasitised RAW 264.7 cells. Phytochemistry 2005, 66, 2056–2071. [Google Scholar] [CrossRef] [PubMed]
- Markiewski, M.M.; Lambris, J.D. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Sarma, J.V.; Ward, P.A. The complement system. Cell Tissue Res. 2011, 343, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Juan, S.-H.; Shen, S.-C.; Hsu, F.-L.; Chen, Y.-C. Inhibition of lipopolysaccharide-induced nitric oxide production by flavonoids in RAW 264.7 macrophages involves heme oxygenase-1. Biochem. Pharmacol. 2003, 66, 1821–1832. [Google Scholar] [CrossRef]
- Wang, J.; Mazza, G. Inhibitory effects of anthocyanins and other phenolic compounds on nitric oxide production in LPS/IFN-γ-activated RAW 264.7 macrophages. J. Agric. Food Chem. 2002, 50, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Sidor, A.; Gramza-Michałowska, A. Advanced research on the antioxidant and health benefit of elderberry (Sambucus nigra) in food—A review. J. Funct. Foods 2014, 18B, 941–958. [Google Scholar] [CrossRef]
- Mascolo, N.; Capasso, F.; Menghini, A.; Fasulo, M.P. Biological screening of Italian medicinal plants for anti-inflammatory activity. Phytother. Res. 1987, 1, 28–31. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Ferrars, R.; Czank, C.; Zhang, Q.; Botting, N.; Kroon, P.; Cassidy, A.; Kay, C. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [PubMed]
- Ferrars, R.M.; Cassidy, A.; Curtis, P.; Kay, C.D. Phenolic metabolites of anthocyanins following a dietary intervention study in post-menopausal women. Mol. Nutr. Food Res. 2014, 58, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Napolitano, J.G.; Lankin, D.C.; Chen, S.N.; Pauli, G.F. Complete 1H NMR spectral analysis of ten chemical markers of Ginkgo biloba. Magn. Reson. Chem. 2012, 50, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Dürüst, N.; Özden, S.; Umur, E.; Dürüst, Y.; Kucukislamoglu, M. The isolation of carboxylic acids from the flowers of Delphinium formosum. Turk. J. Chem. 2001, 25, 93–97. [Google Scholar]
- Mikulic-Petkovsek, M.; Samoticha, J.; Eler, K.; Stampar, F.; Veberic, R. Traditional elderflower beverages: A rich source of phenolic compounds with high antioxidant activity. J. Agric. Food Chem. 2015, 63, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Ø.M.; Aksnes, D.W.; Nerdal, W.; Johansen, O.P. Structure elucidation of cyanidin-3-sambubioside and assignments of the 1H and 13C NMR resonances through two-dimensional shift-correlated NMR techniques. Phytochem. Anal. 1991, 2, 175–183. [Google Scholar] [CrossRef]
- Johansen, O.-P.; Andersen, Ø.M.; Nerdal, W.; Aksnes, D.W. Cyanidin 3-[6-(p-coumaroyl)-2-(xylosyl)-glucoside]-5-glucoside and other anthocyanins from fruits of Sambucus canadensis. Phytochemistry 1991, 30, 4137–4141. [Google Scholar] [CrossRef]
- Alban, S.; Classen, B.; Brunner, G.; Blaschek, W. Differentiation between the complement modulating effects of an arabinogalactan-protein from Echinacea purpurea and heparin. Planta Med. 2002, 68, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Grønhaug, T.E.; Kiyohara, H.; Sveaass, A.; Diallo, D.; Yamada, H.; Paulsen, B.S. β-d-(1→4)-galactan-containing side chains in RG-I regions of pectic polysaccharides from Biophytum petersianum Klotzsch. contribute to expression of immunomodulating activity against intestinal Peyer’s patch cells and macrophages. Phytochemistry 2011, 72, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Inngjerdingen, K.T.; Coulibaly, A.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. A complement fixing polysaccharide from Biophytum petersianum Klotzsch, a medicinal plant from Mali, West Africa. Biomacromolecules 2006, 7, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.T.T.; Ahmed, A.; Zou, Y.-F.; Aslaksen, T.; Wangensteen, H.; Barsett, H. Structure–activity relationship of immunomodulating pectins from elderberries. Carbohydr. Polym. 2015, 125, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Shahat, A.A.; Hammouda, F.; Ismail, S.I.; Azzam, S.A.; De Bruyne, T.; Lasure, A.; van Poel, B.; Pieters, L.; Vlietinck, A.J. Anti-complementary activity of Crataegus sinaica. Planta Med. 1996, 62, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.T.; Bräunlich, M.; Austarheim, I.; Wangensteen, H.; Malterud, K.E.; Slimestad, R.; Barsett, H. Immunomodulating activity of Aronia melanocarpa polyphenols. Int. J. Mol. Sci. 2014, 15, 11626–11636. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.T.T.; Zou, Y.-F.; Aslaksen, T.H.; Wangensteen, H.; Barsett, H. Structural characterization of bioactive pectic polysaccharides from elderflowers (Sambuci flos). Carbohydr. Polym. 2016, 135, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.T.T.; Zou, Y.-F.; Wangensteen, H.; Barsett, H. RG-I regions from elderflower pectins substituted on GalA are strong immunomodulators. Int. J. Biol. Macromol. 2016, 92, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Wen, Q.; Cao, J.; Yin, C.; Chen, D.; Cheng, Z. Flavonol glycosides and other phenolic compounds from Viola tianshanica and their anti-complement activities. Pharm. Biol. 2015, 54, 1140–1147. [Google Scholar] [PubMed]
- Min, B.-S.; Lee, S.-Y.; Kim, J.-H.; Lee, J.-K.; Kim, T.-J.; Kim, D.-H.; Kim, Y.-H.; Joung, H.; Lee, H.-K.; Nakamura, N. Anti-complement activity of constituents from the stem-bark of Juglans mandshurica. Biol. Pharm. Bull. 2003, 26, 1042–1044. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.Y.; Oh, S.R.; Park, S.-H.; Lee, I.S.; Ahn, K.S.; Lee, J.J.; Lee, H.-K. Anti-complement activity of tiliroside from the flower buds of Magnolia fargesii. Biol. Pharm. Bull. 1998, 21, 1077–1078. [Google Scholar] [CrossRef] [PubMed]
- Fossiez, F.; Djossou, O.; Chomarat, P.; Flores-Romo, L.; Ait-Yahia, S.; Maat, C.; Pin, J.-J.; Garrone, P.; Garcia, E.; Saeland, S. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J. Exp. Med. 1996, 183, 2593–2603. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, F.M.; Duma, D.; Assreuy, J.; Buzzi, F.C.; Niero, R.; Campos, M.M.; Calixto, J.B. Caffeic acid derivatives: In vitro and in vivo anti-inflammatory properties. Free Radic. Res. 2004, 38, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Min, S.-W.; Ryu, S.-N.; Kim, D.-H. Anti-inflammatory effects of black rice, cyanidin-3-O-β-d-glycoside, and its metabolites, cyanidin and protocatechuic acid. Int. Immunopharmacol. 2010, 10, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [PubMed]
- Ho, G.T.T.; Kase, E.T.; Wangensteen, H.; Barsett, H. Effect of phenolic compounds from elderflowers on glucose- and fatty acid uptake in human myotubes and HepG2-cells. Molecules 2017, 22, 90. [Google Scholar] [CrossRef] [PubMed]
- Bräunlich, M.; Slimestad, R.; Wangensteen, H.; Brede, C.; Malterud, K.E.; Barsett, H. Extracts, anthocyanins and procyanidins from Aronia melanocarpa as radical scavengers and enzyme inhibitors. Nutrients 2013, 5, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, T.; Gilje, A.; Samuelsen, A.; Høgåsen, K.; Paulsen, B. Interaction between human complement and a pectin type polysaccharide fraction, PMII, from the leaves of Plantago major L. Scand. J. Immunol. 2000, 52, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Roche Cell Proliferation Kit I (MTT). Available online: https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Roche/Bulletin/1/11465007001bul.pdf (accessed on 19 June 2016).

| Chemical Compound | R1 | R2 | R3 | R4 | R5 | R6 |
|---|---|---|---|---|---|---|
| p-Coumaric acid | CH=CH–COOH | H | H | OH | H | H |
| Homovanillic acid | CH2–COOH | H | H | OH | OCH3 | H |
| Phloroglucinol aldehyde | CHO | OH | H | OH | H | OH |
| 4-Hydroxybenzoic acid | COOH | H | H | OH | H | H |
| Hippuric acid | CO–NH–CH2–COOH | H | H | H | H | H |
| Ferulic acid | CH=CH–COOH | H | OCH3 | OH | H | H |
| 4-Hydroxybenzaldehyde | CHO | H | H | OH | H | H |
| Protocatechuic acid | COOH | H | H | OH | OH | H |
| Caffeic acid | CH=CH–COOH | H | H | OH | OH | H |
| Vanillic acid | COOH | H | H | OH | OCH3 | H |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Compounds | IC50 (µg/mL) a |
|---|---|
| Crude extracts from elderberry | |
| EB DCM | >200 |
| EB 96% EtOH | 7.8 ± 2.3 |
| EB 50% EtOH | 13.4 ± 2.9 |
| EB 0.5% TFA in MeOH | 12.3 ± 1.9 |
| EB 50 °C water | 44.9 ± 5.3 |
| EB 100 °C water | 23.3 ± 3.5 |
| Pressed juice | 142.4 ± 13.1 |
| Crude extracts from elderflower | |
| EF DCM | >200 |
| EF 96% EtOH | 6.5 ± 1.5 |
| EF 50% EtOH | 8.9 ± 2.2 |
| EF 50 °C water | 32.3 ± 5.2 |
| EF 100 °C water | 14.4 ± 3.1 |
| BPII (positive control) | 17.7 ± 1.5 |
| Test Compounds | IC50 (µM) a |
|---|---|
| Flavonoids | |
| Cyanidin | 74.2 ± 3.4 |
| Cyanidin-3-glucoside | 87.1 ± 5.6 |
| Cyanidin-3-sambubioside | 82.8 ± 3.9 |
| Procyanidin B2 | 70.6 ± 4.5 |
| Procyanidin B5 | 65.0 ± 3.1 |
| Procyanidin C1 | 19.4 ± 2.1 |
| Quercetin | 193.8 ± 5.6 |
| Quercetin-3-glucoside | 76.5 ± 2.6 |
| Quercetin-3-rhamnoside | 95.1 ± 3.1 |
| Rutin | 40.0 ± 2.6 |
| Kaempferol | >200 |
| Kaempferol-3-rutinoside | 70.6 ± 3.8 |
| Isorhamnetin | >200 |
| Isorhamnetin-3-rutinoside | 127.8 ± 4.8 |
| Naringenin | >200 |
| Catechin | >200 |
| Epicatechin | >200 |
| Metabolites | |
| p-Coumaric acid | >200 |
| Homovanillic acid | >200 |
| Phloroglucinol aldehyde | >200 |
| 4-Hydroxybenzoic acid | >200 |
| Hippuric acid | >200 |
| Ferulic acid | >200 |
| 4-Hydroxybenzaldehyde | >200 |
| Protocatechuic acid | >200 |
| Caffeic acid | >200 |
| Vanillic acid | >200 |
| Benzoic acid | >200 |
| 3-Hydroxybenzoic acid | >200 |
| 3,4-Dihydroxyphenylacetic acid | >200 |
| 4-Methylcatechol | >200 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, G.T.T.; Wangensteen, H.; Barsett, H. Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells. Int. J. Mol. Sci. 2017, 18, 584. https://doi.org/10.3390/ijms18030584
Ho GTT, Wangensteen H, Barsett H. Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells. International Journal of Molecular Sciences. 2017; 18(3):584. https://doi.org/10.3390/ijms18030584
Chicago/Turabian StyleHo, Giang Thanh Thi, Helle Wangensteen, and Hilde Barsett. 2017. "Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells" International Journal of Molecular Sciences 18, no. 3: 584. https://doi.org/10.3390/ijms18030584
APA StyleHo, G. T. T., Wangensteen, H., & Barsett, H. (2017). Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells. International Journal of Molecular Sciences, 18(3), 584. https://doi.org/10.3390/ijms18030584