
Vanillin Suppresses Cell Motility by Inhibiting STAT3-Mediated HIF-1α mRNA Expression in Malignant Melanoma Cells

 and
and 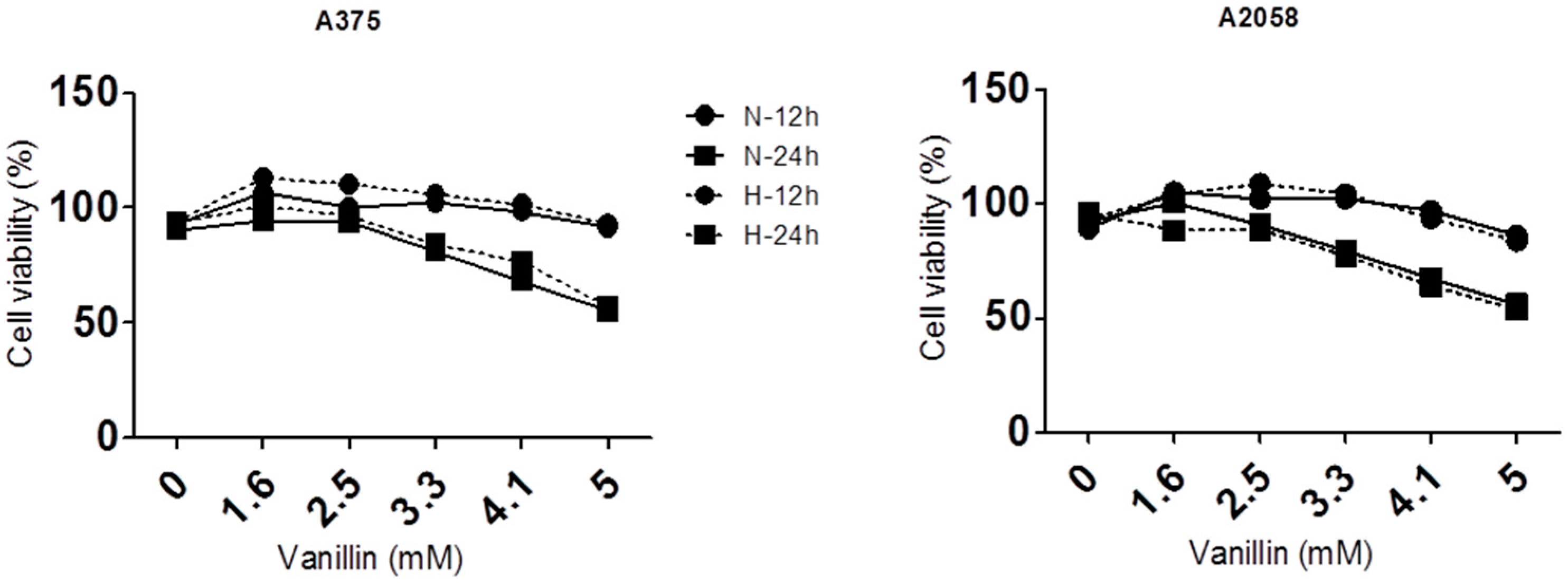{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
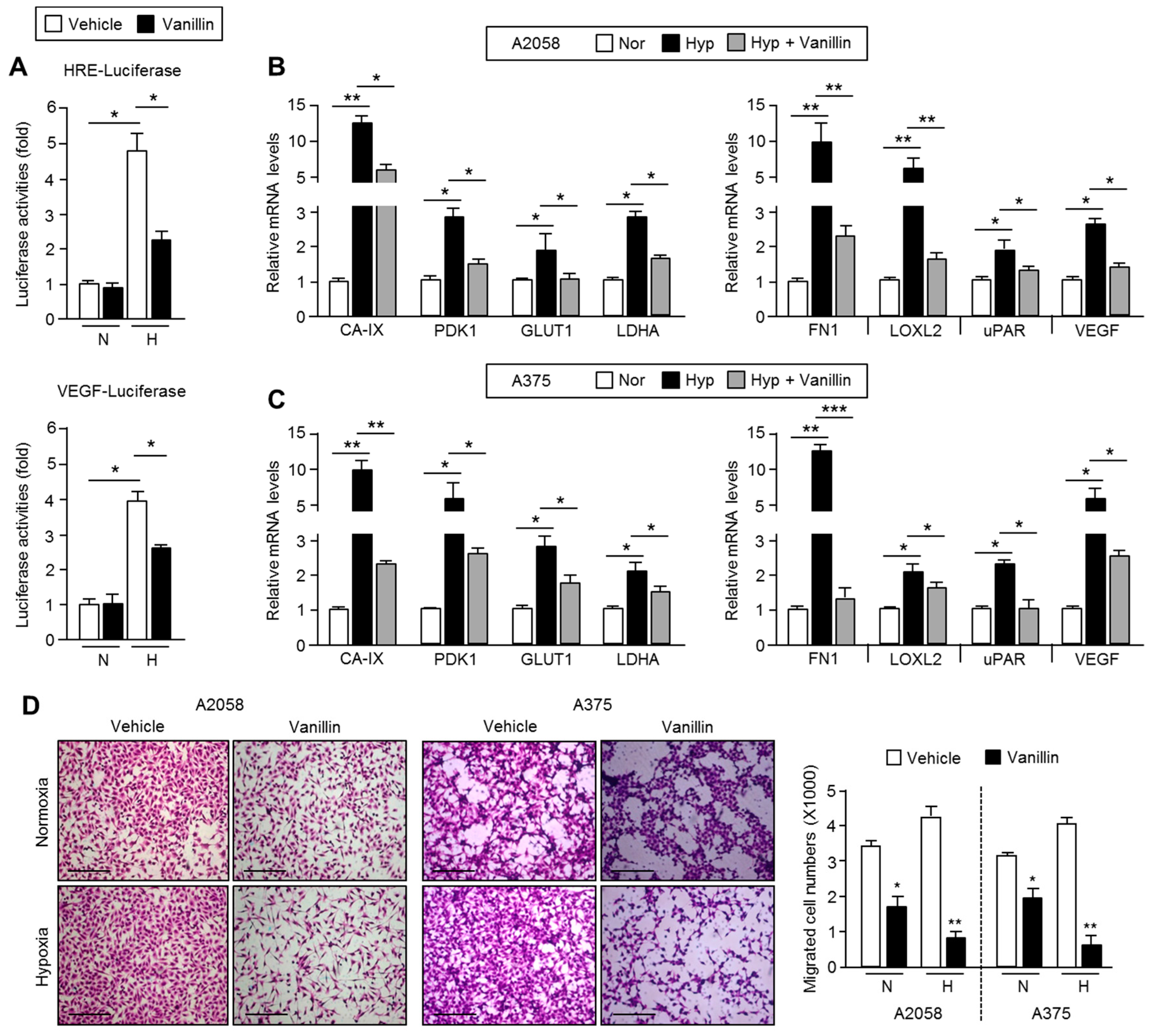
2.1. Anti-Cancer Effects of Vanillin in Human A2058 and A375 Malignant Melanoma Cells
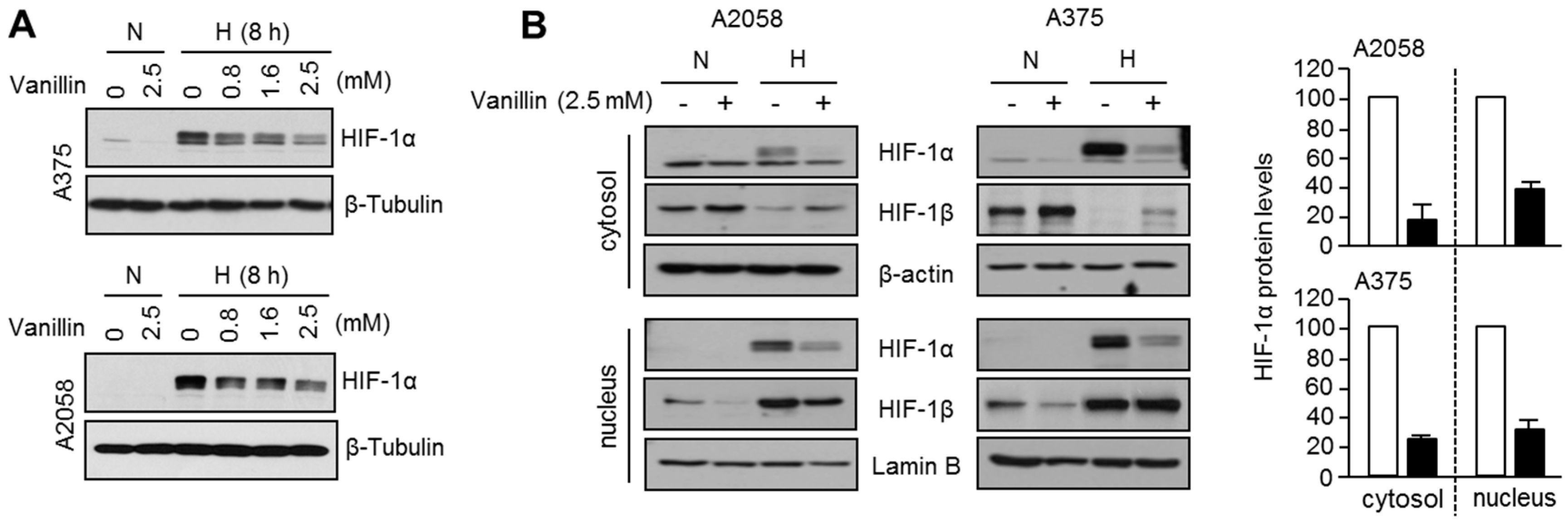
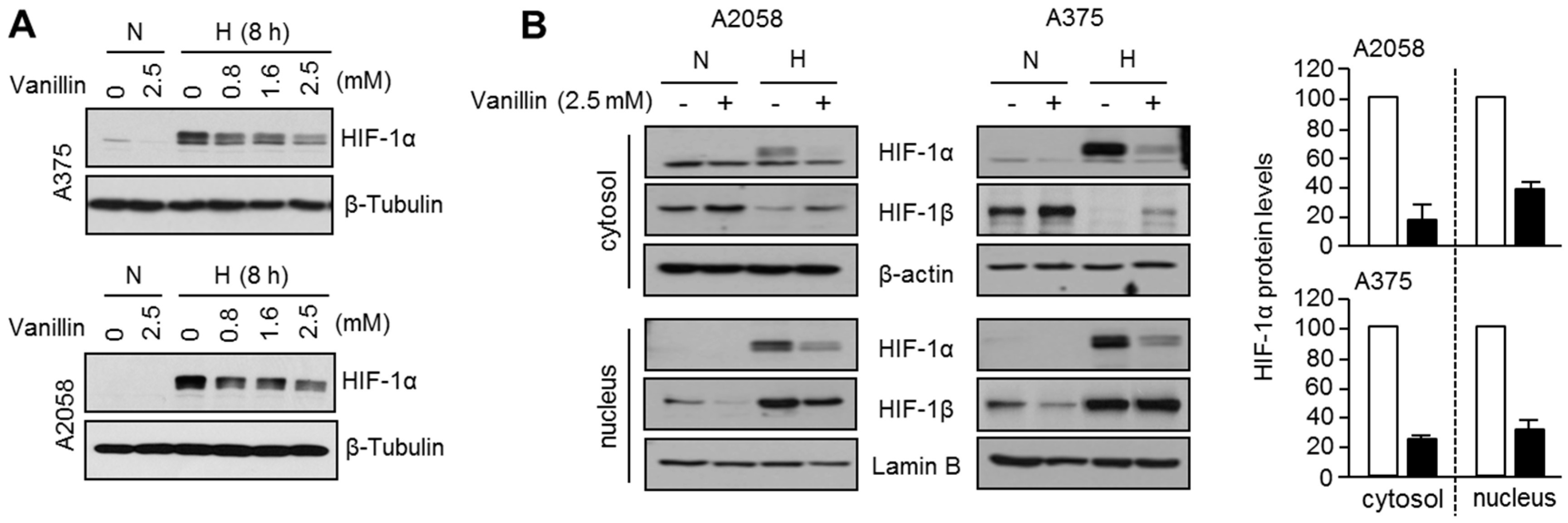
2.2. Vanillin Decreases HIF-1α Protein Levels under Hypoxia in A2058 and A375 Malignant Melanoma Cells
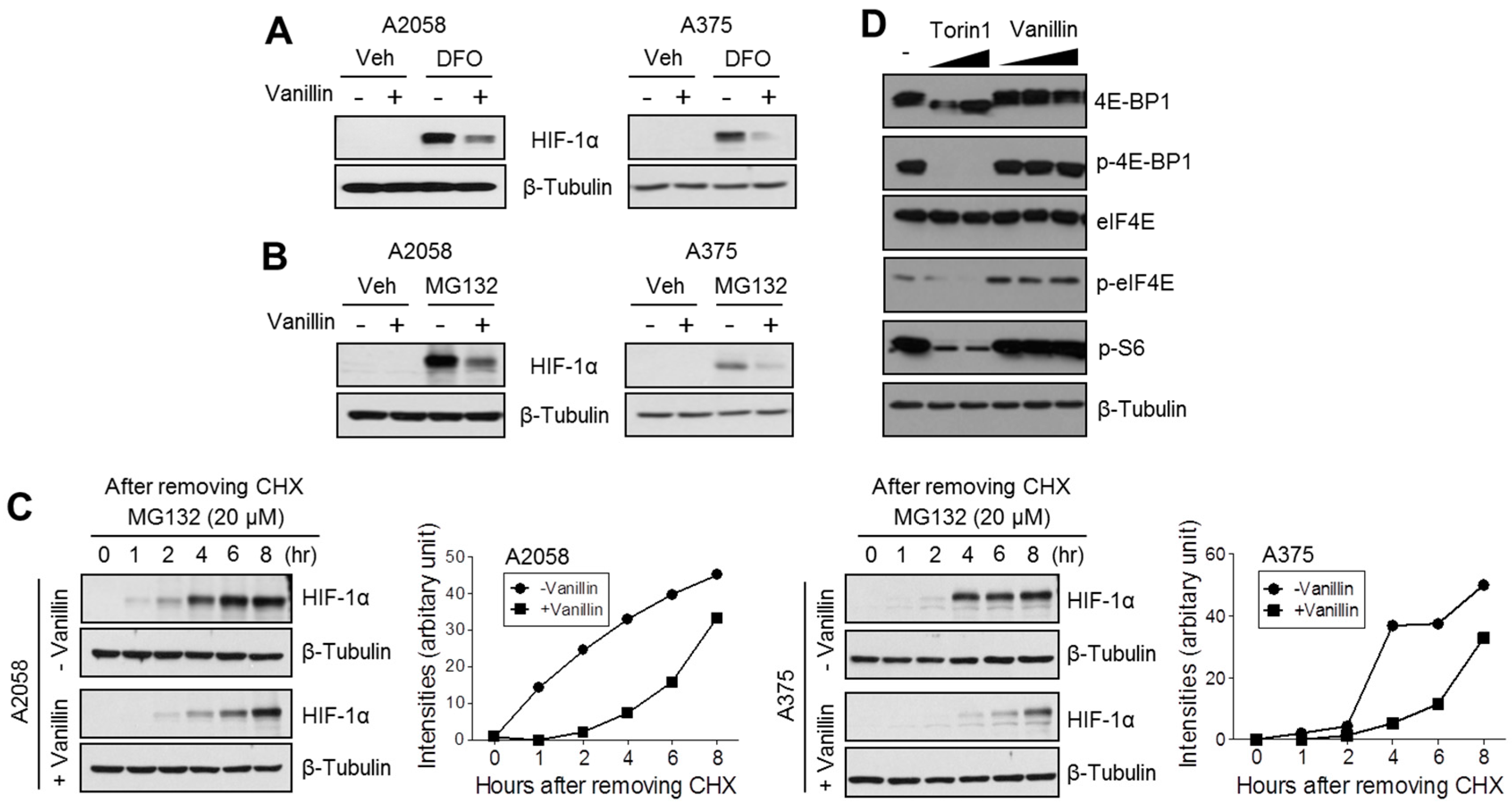
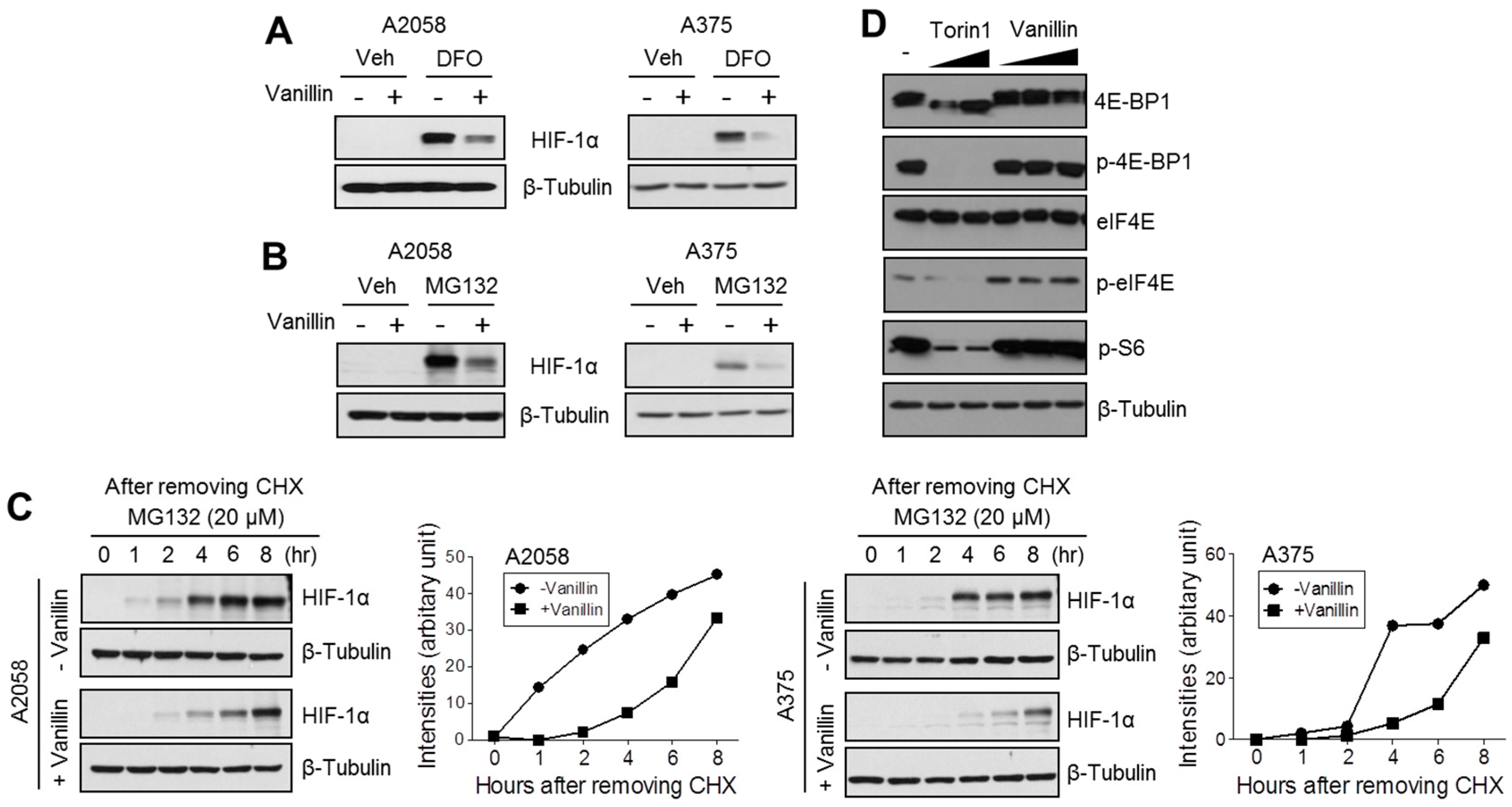
2.3. Vanillin Attenuates HIF-1α Protein Synthesis
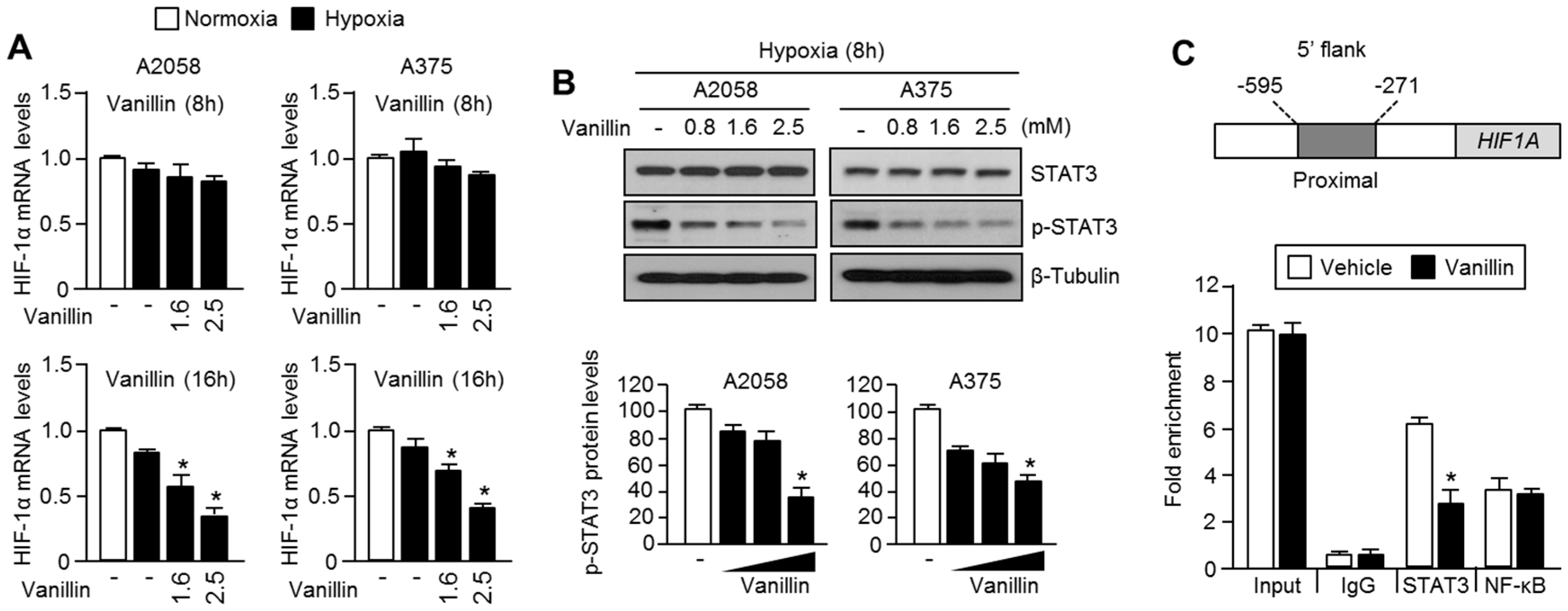
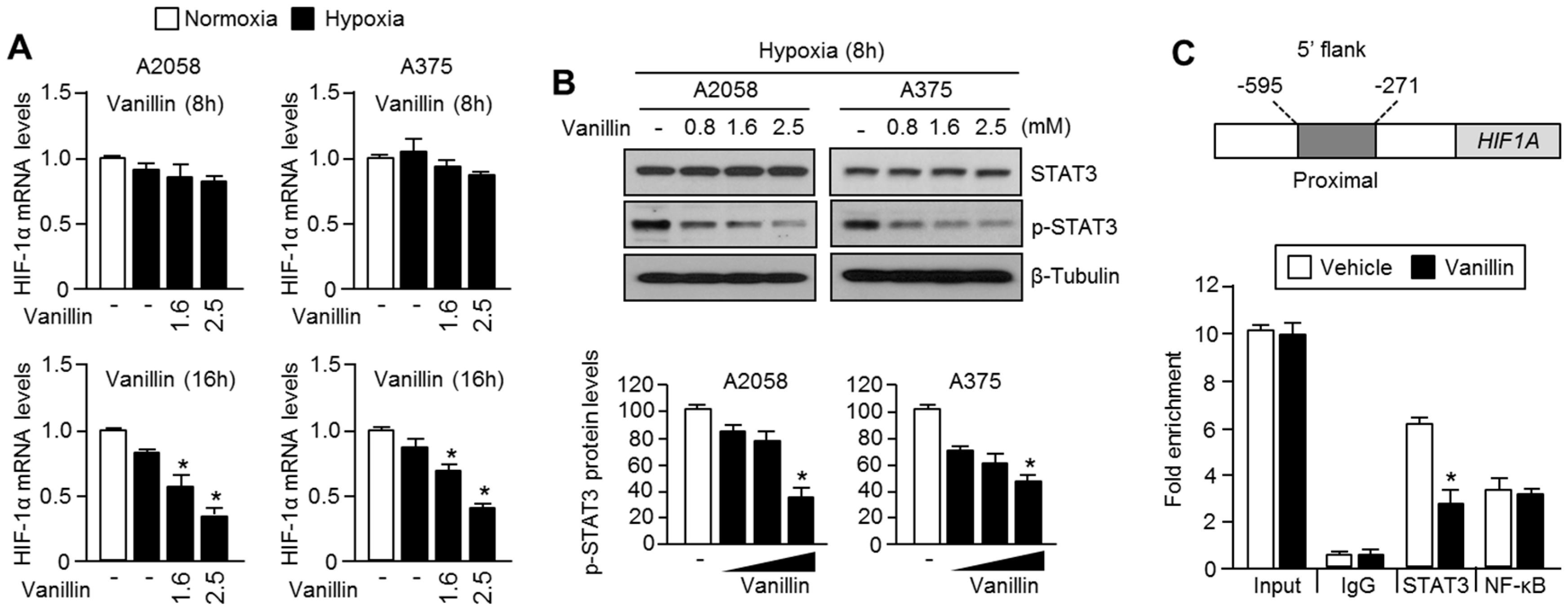
2.4. Vanillin Decreases HIF-1α mRNA Levels by Inhibiting Promoter Occupancy of STAT3 at HIF1A
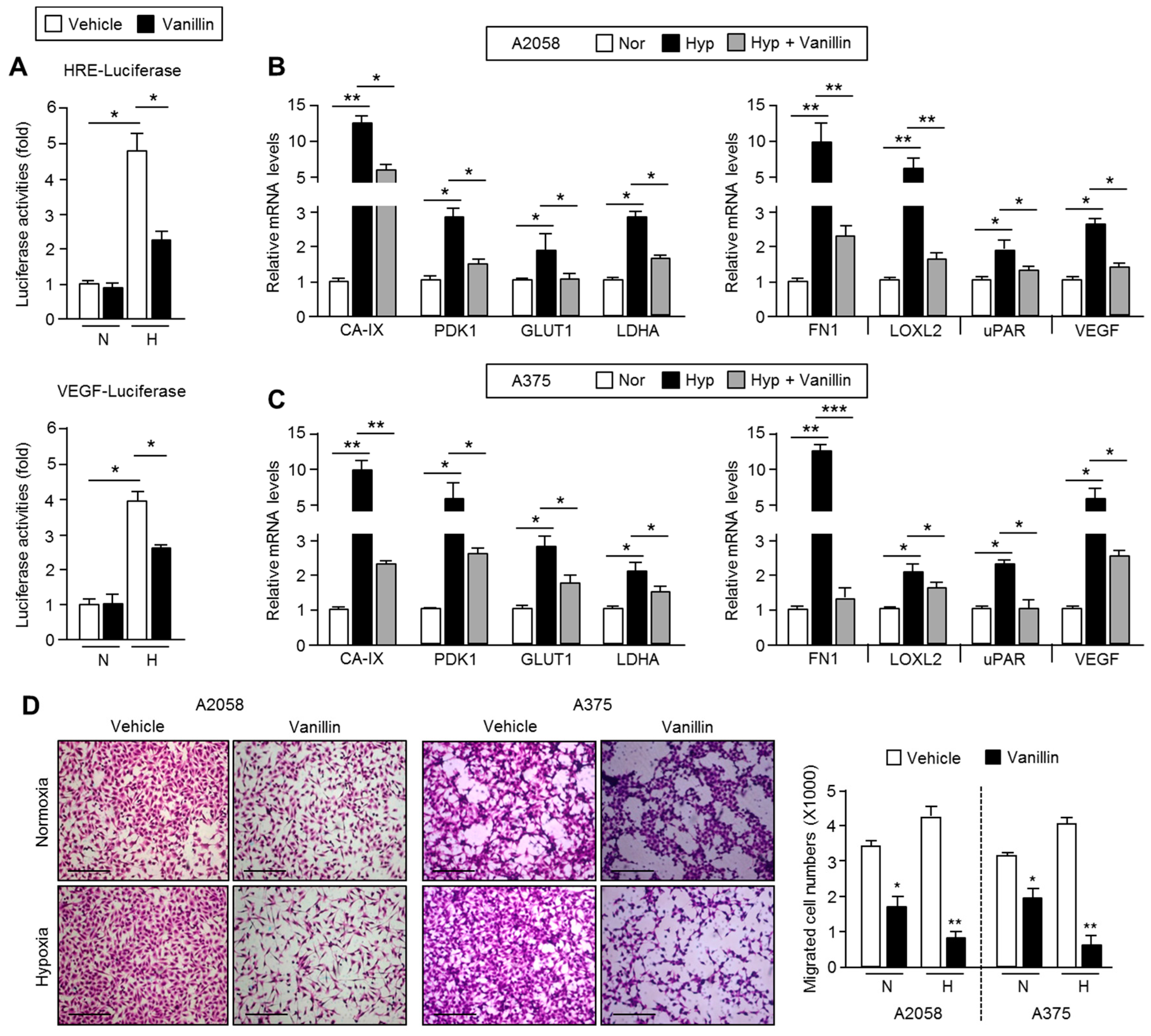
2.5. Vanillin Down-Regulates HIF-1α Target Gene Expression and Causes Suppression of Cell Motility
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Cytosolic and Nuclear Extract Preparation
4.5. Immunoblotting
4.6. Quantitative Real-Time PCR
4.7. In Vitro Migration Assay
4.8. Chromatin Immunoprecipitation
4.9. Luciferase Activity Assay
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HIF-1α | Hypoxia-inducible factor-1α |
| STAT3 | Signal transducer and activator of transcription 3 |
| NF-κB | Nuclear factor-κB |
| FN1 | Fibronectin 1 |
| LOXL2 | Lysyl oxidase-like 2 |
| uPAR | Urokinase plasminogen activator receptor |
| VHL | Von Hippel-Lindau protein |
| MAPK | Mitogen activated protein kinase |
| HRE | Hypoxia-response element |
| MMPs | Matrix metalloproteinases |
| PI3K | Phosphoinositide 3-kinase |
| mTOR | Mammalian target of rapamycin |
| 4E-BP1 | Eukaryotic translation initiation factor 4E-binding protein 1 |
| eIF4E | Eukaryotic translation initiation factor 4E |
References
- Haq, R.; Fisher, D.E. Targeting melanoma by small molecules: Challenges ahead. Pigment Cell. Melanoma Res. 2013, 26, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell. Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Lee, Y.M.; Lee, G.; Choi, Y.J.; Lim, B.O.; Kim, Y.J.; Choi, D.K.; Park, J.W. PRMT5 is essential for the eIF4E-mediated 5′-cap dependent translation. Biochem. Biophys. Res. Commun. 2014, 452, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Dodd, K.M.; Tee, A.R. STAT3 and mTOR: Co-operating to drive HIF and angiogenesis. Oncoscience 2015, 2, 913–914. [Google Scholar] [PubMed]
- Lee, Y.M.; Lim, J.H.; Yoon, H.; Chun, Y.S.; Park, J.W. Antihepatoma activity of chaetocin due to deregulated splicing of hypoxia-inducible factor 1α pre-mRNA in mice and in vitro. Hepatology 2011, 53, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Briggs, J.; Deng, J.; Ma, Y.; Lee, H.; Kortylewski, M.; Kujawski, M.; Kay, H.; Cress, W.D.; Jove, R.; et al. Signal transducer and activator of transcription 3 is required for hypoxia-inducible factor-1α RNA expression in both tumor cells and tumor-associated myeloid cells. Mol. Cancer Res. 2008, 6, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of hypoxia-inducible factor-1α by NF-κB. Biochem. J. 2008, 412, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. 2003, 3, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Akagi, K.; Hirose, M.; Hoshiya, T.; Mizoguchi, Y.; Ito, N.; Shirai, T. Modulating effects of ellagic acid, vanillin and quercetin in a rat medium term multi-organ carcinogenesis model. Cancer Lett. 1995, 94, 113–121. [Google Scholar] [CrossRef]
- King, A.A.; Shaughnessy, D.T.; Mure, K.; Leszczynska, J.; Ward, W.O.; Umbach, D.M.; Xu, Z.; Ducharme, D.; Taylor, J.A.; Demarini, D.M.; et al. Antimutagenicity of cinnamaldehyde and vanillin in human cells: Global gene expression and possible role of DNA damage and repair. Mutat. Res. 2007, 616, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, D.T.; Schaaper, R.M.; Umbach, D.M.; DeMarini, D.M. Inhibition of spontaneous mutagenesis by vanillin and cinnamaldehyde in Escherichia coli: Dependence on recombinational repair. Mutat. Res. 2006, 602, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.; Yazan, L.S.; Ismail, N.; Ismail, M. Apoptosis and cell cycle arrest of human colorectal cancer cell line HT-29 induced by vanillin. Cancer Epidemiol. 2009, 33, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Lirdprapamongkol, K.; Sakurai, H.; Suzuki, S.; Koizumi, K.; Prangsaengtong, O.; Viriyaroj, A.; Ruchirawat, S.; Svasti, J.; Saiki, I. Vanillin enhances TRAIL-induced apoptosis in cancer cells through inhibition of NF-κB activation. In Vivo 2010, 24, 501–506. [Google Scholar] [PubMed]
- Marton, A.; Kusz, E.; Kolozsi, C.; Tubak, V.; Zagotto, G.; Buzas, K.; Quintieri, L.; Vizler, C. Vanillin Analogues O-vanillin and 2,4,6-Trihydroxybenzaldehyde Inhibit NF-κB Activation and Suppress Growth of A375 Human Melanoma. Anticancer Res. 2016, 36, 5743–5750. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.A.; Wu, S.L.; Lo, H.Y.; Hsiang, C.Y.; Ho, T.Y. Vanillin inhibits matrix metalloproteinase-9 expression through down-regulation of nuclear factor-κB signaling pathway in human hepatocellular carcinoma cells. Mol. Pharmacol. 2009, 75, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Lirdprapamongkol, K.; Kramb, J.P.; Suthiphongchai, T.; Surarit, R.; Srisomsap, C.; Dannhardt, G.; Svasti, J. Vanillin suppresses metastatic potential of human cancer cells through PI3K inhibition and decreases angiogenesis in vivo. J. Agric. Food Chem. 2009, 57, 3055–3063. [Google Scholar] [CrossRef] [PubMed]
- Lirdprapamongkol, K.; Sakurai, H.; Kawasaki, N.; Choo, M.K.; Saitoh, Y.; Aozuka, Y.; Singhirunnusorn, P.; Ruchirawat, S.; Svasti, J.; Saiki, I. Vanillin suppresses in vitro invasion and in vivo metastasis of mouse breast cancer cells. Eur. J. Pharm. Sci. 2005, 25, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Nagle, D.G.; Zhou, Y.D. Natural product-based inhibitors of hypoxia-inducible factor-1 (HIF-1). Curr. Drug Targets 2006, 7, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Fang, J.; Xia, C.; Shi, X.; Jiang, B.H. Trans-3,4,5′-Trihydroxystibene inhibits hypoxia-inducible factor 1α and vascular endothelial growth factor expression in human ovarian cancer cells. Clin. Cancer Res. 2004, 10, 5253–5263. [Google Scholar] [CrossRef] [PubMed]
- Osada, M.; Imaoka, S.; Funae, Y. Apigenin suppresses the expression of VEGF, an important factor for angiogenesis, in endothelial cells via degradation of HIF-1α protein. FEBS Lett. 2004, 575, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Jiang, B.H.; Semenza, G.L. Effect of protein kinase and phosphatase inhibitors on expression of hypoxia-inducible factor 1. Biochem. Biophys. Res. Commun. 1995, 216, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Welsh, S.J.; Williams, R.R.; Birmingham, A.; Newman, D.J.; Kirkpatrick, D.L.; Powis, G. The thioredoxin redox inhibitors 1-methylpropyl 2-imidazolyl disulfide and pleurotin inhibit hypoxia-induced factor 1α and vascular endothelial growth factor formation. Mol. Cancer Ther. 2003, 2, 235–243. [Google Scholar] [PubMed]
- Gupta, S.C.; Kim, J.H.; Prasad, S.; Aggarwal, B.B. Regulation of survival, proliferation, invasion, angiogenesis, and metastasis of tumor cells through modulation of inflammatory pathways by nutraceuticals. Cancer Metastasis Rev. 2010, 29, 405–434. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, E.-J.; Lee, Y.-M.; Oh, T.-I.; Kim, B.M.; Lim, B.-O.; Lim, J.-H. Vanillin Suppresses Cell Motility by Inhibiting STAT3-Mediated HIF-1α mRNA Expression in Malignant Melanoma Cells. Int. J. Mol. Sci. 2017, 18, 532. https://doi.org/10.3390/ijms18030532
Park E-J, Lee Y-M, Oh T-I, Kim BM, Lim B-O, Lim J-H. Vanillin Suppresses Cell Motility by Inhibiting STAT3-Mediated HIF-1α mRNA Expression in Malignant Melanoma Cells. International Journal of Molecular Sciences. 2017; 18(3):532. https://doi.org/10.3390/ijms18030532
Chicago/Turabian StylePark, Eun-Ji, Yoon-Mi Lee, Taek-In Oh, Byeong Mo Kim, Beong-Ou Lim, and Ji-Hong Lim. 2017. "Vanillin Suppresses Cell Motility by Inhibiting STAT3-Mediated HIF-1α mRNA Expression in Malignant Melanoma Cells" International Journal of Molecular Sciences 18, no. 3: 532. https://doi.org/10.3390/ijms18030532
APA StylePark, E.-J., Lee, Y.-M., Oh, T.-I., Kim, B. M., Lim, B.-O., & Lim, J.-H. (2017). Vanillin Suppresses Cell Motility by Inhibiting STAT3-Mediated HIF-1α mRNA Expression in Malignant Melanoma Cells. International Journal of Molecular Sciences, 18(3), 532. https://doi.org/10.3390/ijms18030532