
Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections

Abstract
:1. Introduction
2. Lactoferrin: Distribution, Structure and Biological Function as an Antimicrobial
3. Lactoferrin as Anti-Inflammatory in Infectious Diseases
3.1. Gastrointestinal Tract
3.1.1. Gastrointestinal Tract Inflammation: An Overview
3.1.2. Modulatory Effects of Lactoferrin on the Inflammatory Response Associated to Gut Infections
3.1.3. Effect of Lactoferrin on Gut-Associated Sepsis
3.1.4. Gut and Systemic Lipopolysaccharide (LPS)-Related Inflammation
3.2. Respiratory Tract
3.2.1. Respiratory Tract Inflammation and Infections: An Overview
3.2.2. Modulatory Effects of Lactoferrin on the Infection-Associated Inflammatory Response in the Respiratory Tract
3.3. Modulatory Effects of Lactoferrin on Other Infection-Associated Inflammatory Processes Inflammation
Colostrum, Milk and Mastitis by Staphylococci
4. Perspectives
Acknowledgments
Conflicts of Interest
Abbreviations
| apo-Lf | apo-lactoferrin (iron-free lactoferrin) |
| bLf | bovine lactoferrin |
| Lfcin B | bovine lactoferricin B |
| d-GalN | d-galactosamine |
| IFNγ | γ interferon |
| holo-Lf | holo-lactoferrin (iron-loaded lactoferrin) |
| holo-Tf | holo-transferrin (iron-loaded transferrin) |
| hLf | human Lf |
| IBD | inflammatory bowel disease |
| IL | interleukin |
| Lfcin | lactoferricin |
| Lf | lactoferrin |
| LfR | lactoferrin receptor |
| NF-κB | nuclear factor κB |
| ovoTf | ovotransferrin |
| PMN | polymorphonuclear |
| r-hLf | recombinant human lactoferrin |
| RSV | respiratory syncytial virus |
| RT | respiratory tract |
| sIgA | secretory immunoglobulin A |
| Tf | transferrin |
| TNFα | tumor necrosis factor α |
References
- Vorland, L.H. Lactoferrin: A multifunctional glycoprotein. APMIS 1999, 107, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Panella, G.; Leboffe, L.; Antonini, G. Lactoferrin from Milk: Nutraceutical and Pharmacological Properties. Pharmaceuticals 2016, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D. Overview of Lactoferrin as a Natural Immune Modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef] [PubMed]
- Farnaud, S.; Evans, R.W. Lactoferrin—A multifunctional protein with antimicrobial properties. Mol. Immunol. 2003, 40, 395–405. [Google Scholar] [CrossRef]
- Puddu, P.; Valenti, P.; Gessani, S. Immunomodulatory effects of lactoferrin on antigen presenting cells. Biochimie 2009, 91, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Gupta, I.; Sehgal, R.; Kanwar, R.K.; Punj, V.; Kanwar, J.R. Nanocapsules loaded with iron-saturated bovine lactoferrin have antimicrobial therapeutic potential and maintain calcium, zinc and iron metabolism. Nanomedicine 2015, 10, 1289–1314. [Google Scholar] [CrossRef] [PubMed]
- Groves, M.L. The Isolation of a Red Protein from Milk 2. J. Am. Chem. Soc. 1960, 82, 3345–3350. [Google Scholar] [CrossRef]
- Montreuil, J.; Tonnelat, J.; Mullet, S. Preparation and properties of lactosiderophilin (lactotransferrin) of human milk. Biochim. Biophys. Acta 1960, 45, 413–421. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M. A structural framework for understanding the multifunctional character of lactoferrin. Biochimie 2009, 91, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Moguilevsky, N.; Retegui, L.A.; Masson, P.L. Comparison of human lactoferrins from milk and neutrophilic leucocytes. Relative molecular mass, isoelectric point, iron-binding properties and uptake by the liver. Biochem. J. 1985, 229, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Steijns, J.M.; van Hooijdonk, A.C. Occurrence, structure, biochemical properties and technological characteristics of lactoferrin. Br. J. Nutr. 2000, 84, S11–S17. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.L.; Heremans, J.F. Metal-combining properties of human lactoferrin (red milk protein). 1. The involvement of bicarbonate in the reaction. Eur. J. Biochem. 1968, 6, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Testa, U. Proteins of Iron Metabolism; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Brock, J.H. The physiology of lactoferrin. Biochem. Cell Biol. 2002, 80, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.L.; Heremans, J.F. Lactoferrin in milk from different species. Comp. Biochem. Physiol. B 1971, 39, 119–129. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F.; Dive, C.H. An iron-binding protein common to many external secretions. Clin. Chim. Acta 1966, 14, 735–739. [Google Scholar] [CrossRef]
- Legrand, D.; Elass, E.; Pierce, A.; Mazurier, J. Lactoferrin and host defence: An overview of its immuno-modulating and anti-inflammatory properties. Biometals 2004, 17, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, L.; Calvo, M.; Brock, J.H. Biological role of lactoferrin. Arch. Dis. Child. 1992, 67, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Welty, F.K.; Smith, K.L.; Schanbacher, F.L. Lactoferrin concentration during involution of the bovine mammary gland. J. Dairy Sci. 1976, 59, 224–231. [Google Scholar] [CrossRef]
- Hirai, Y.; Kawakata, N.; Satoh, K.; Ikeda, Y.; Hisayasu, S.; Orimo, H.; Yoshino, Y. Concentrations of lactoferrin and iron in human milk at different stages of lactation. J. Nutr. Sci. Vitaminol. 1990, 36, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, L.; Aranda, P.; Pérez, M.D.; Calvo, M. Concentration of lactoferrin and transferrin throughout lactation in cow’s colostrum and milk. Biol. Chem. Hoppe Seyler 1988, 369, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- Lambert, L.A. Molecular evolution of the transferrin family and associated receptors. Biochim. Biophys. Acta 2012, 1820, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Swiss-Prot (2007) Swiss-Prot Database. Available online: http://web.expasy.org/docs/swiss-prot_guideline.html (accessed on 1 January 2017).
- Bullen, J.J. The significance of iron in infection. Rev. Infect. Dis. 1981, 3, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Van Snick, J.L.; Masson, P.L.; Heremans, J.F. The involvement of lactoferrin in the hyposideremia of acute inflammation. J. Exp. Med. 1974, 140, 1068–1084. [Google Scholar] [CrossRef] [PubMed]
- Levay, P.F.; Viljoen, M. Lactoferrin: A general review. Haematologica 1995, 80, 252–267. [Google Scholar] [PubMed]
- Jenssen, H.; Hancock, R.E.W. Antimicrobial properties of lactoferrin. Biochimie 2009, 91, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, E.D. The therapeutic potential of lactoferrin. Expert Opin. Investig. Drugs 2003, 12, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.M.; Kokocinski, T. Lactoferrin content of peripheral blood cells. Br. J. Haematol. 1978, 39, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Rado, T.A.; Bollekens, J.; St Laurent, G.; Parker, L.; Benz, E.J. Lactoferrin biosynthesis during granulocytopoiesis. Blood 1984, 64, 1103–1109. [Google Scholar] [PubMed]
- Bennett, R.M.; Kokocinski, T. Lactoferrin turnover in man. Clin. Sci. 1979, 57, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.L.; Heremans, J.F.; Schonne, E. Lactoferrin, an iron-binding protein in neutrophilic leukocytes. J. Exp. Med. 1969, 130, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, T.; Savilahti, E.; Kolho, K.-L.; Nuutinen, H.; Turunen, U.; Färkkilä, M. Crohn’s disease activity assessed by fecal calprotectin and lactoferrin: Correlation with Crohn’s disease activity index and endoscopic findings. Inflamm. Bowel Dis. 2008, 14, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.F.; Baker, H.M.; Dodson, E.J.; Norris, G.E.; Rumball, S.V.; Waters, J.M.; Baker, E.N. Structure of human lactoferrin at 3.2-A resolution. Proc. Natl. Acad. Sci. USA 1987, 84, 1769–1773. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A.; Anderson, B.F.; Groom, C.R.; Haridas, M.; Baker, E.N. Three-dimensional structure of diferric bovine lactoferrin at 2.8 A resolution. J. Mol. Biol. 1997, 274, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Coddeville, B.; Strecker, G.; Wieruszeski, J.M.; Vliegenthart, J.F.; van Halbeek, H.; Peter-Katalinić, J.; Egge, H.; Spik, G. Heterogeneity of bovine lactotransferrin glycans. Characterization of α-d-Galp-(1→3)-β-d-Gal- and α-NeuAc-(2→6)-β-d-GalpNAc-(1→4)-β-d-GlcNAc-substituted N-linked glycans. Carbohydr. Res. 1992, 236, 145–164. [Google Scholar] [CrossRef]
- Mirza, S.; Benjamin, W.H.; Coan, P.A.; Hwang, S.-A.; Winslett, A.-K.; Yother, J.; Hollingshead, S.K.; Fujihashi, K.; Briles, D.E. The effects of differences in pspA alleles and capsular types on the resistance of Streptococcus pneumoniae to killing by apolactoferrin. Microb. Pathog. 2016, 99, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.R.; Russell, J.E.; Champion, W.J.; Brewer, M.; Gauthier, J.J. Bactericidal activity of human lactoferrin: Differentiation from the stasis of iron deprivation. Infect. Immun. 1982, 35, 792–799. [Google Scholar] [PubMed]
- Salamah, A.A.; al-Obaidi, A.S. In vivo and in vitro effects of lactoferrin on Yersinia pseudotuberculosis. New Microbiol. 1995, 18, 267–274. [Google Scholar] [PubMed]
- Yamauchi, K.; Tomita, M.; Giehl, T.J.; Ellison, R.T. Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment. Infect. Immun. 1993, 61, 719–728. [Google Scholar] [PubMed]
- Leon-Sicairos, N.; Reyes-López, M.; Canizalez-Román, A.; Bermúdez-Cruz, R.M.; Serrano-Luna, J.; Arroyo, R.; de la Garza, M. Human hololactoferrin: Endocytosis and use as an iron source by the parasite Entamoeba histolytica. Microbiology 2005, 151, 3859–3871. [Google Scholar] [CrossRef] [PubMed]
- Ortíz-Estrada, G.; Luna-Castro, S.; Piña-Vázquez, C.; Samaniego-Barrón, L.; León-Sicairos, N.; Serrano-Luna, J.; de la Garza, M. Iron-saturated lactoferrin and pathogenic protozoa: Could this protein be an iron source for their parasitic style of life? Future Microbiol. 2012, 7, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Mickelsen, P.A.; Blackman, E.; Sparling, P.F. Ability of Neisseria gonorrhoeae, Neisseria meningitidis, and commensal Neisseria species to obtain iron from lactoferrin. Infect. Immun. 1982, 35, 915–920. [Google Scholar] [PubMed]
- Majka, G.; Więcek, G.; Śróttek, M.; Śpiewak, K.; Brindell, M.; Koziel, J.; Marcinkiewicz, J.; Strus, M. The impact of lactoferrin with different levels of metal saturation on the intestinal epithelial barrier function and mucosal inflammation. Biometals 2016, 29, 1019–1033. [Google Scholar] [CrossRef] [PubMed]
- Cavestro, G.M.; Ingegnoli, A.V.; Aragona, G.; Iori, V.; Mantovani, N.; Altavilla, N.; Dal Bò, N.; Pilotto, A.; Bertelè, A.; Franzè, A.; et al. Lactoferrin: Mechanism of action, clinical significance and therapeutic relevance. Acta Biomed. 2002, 73, 71–73. [Google Scholar] [PubMed]
- Pierce, A.; Legrand, D.; Mazurier, J. La lactoferrine: Une protéine multifonctionnelle. Méd. Sci. 2009, 25, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Iyer, S. Lactoferrin: Molecular Structure and Biological Function. Annu. Rev. Nutr. 1995, 15, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Leitch, E.C.; Willcox, M.D.P. Elucidation of the antistaphylococcal action of lactoferrin and lysozyme. J. Med. Microbiol. 1999, 48, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Van Hooijdonk, A.C.; Kussendrager, K.D.; Steijns, J.M. In vivo antimicrobial and antiviral activity of components in bovine milk and colostrum involved in non-specific defence. Br. J. Nutr. 2000, 84, S127–S134. [Google Scholar] [CrossRef] [PubMed]
- Appelmelk, B.J.; Geerts, M.; Thijs, B.G.; de Boer, H.A.; MAcLAREN, D.M.; de Graaff, J.; Nuijens, J.H.; Pharming Europe, G.B. Lactoferrin Is a Lipid A-Binding Protein. Infect. Immun. 1994, 62, 2628–2632. [Google Scholar] [PubMed]
- Elass-Rochard, E.; Legrand, D.; Salmon, V.; Roseanu, A.; Trif, M.; Tobias, P.S.; Mazurier, J.; Spik, G. Lactoferrin inhibits the endotoxin interaction with CD14 by competition with the lipopolysaccharide-binding protein. Infect. Immun. 1998, 66, 486–491. [Google Scholar] [PubMed]
- Samaniego-Barron, L.; Luna-Castro, S.; Piña-Vázquez, C.; Suárez-Güemes, F. Two outer membrane proteins are bovine lactoferrin-binding proteins in Mannheimia haemolytica A1. Vet. Res. 2016, 47. [Google Scholar] [CrossRef] [PubMed]
- Almaas, H.; Holm, H.; Langsrud, T.; Flengsrud, R.; Vegarud, G.E. In vitro studies of the digestion of caprine whey proteins by human gastric and duodenal juice and the effects on selected microorganisms. Br. J. Nutr. 2006, 96, 562–569. [Google Scholar] [PubMed]
- el Agamy, E.I.; Ruppanner, R.; Ismail, A.; Champagne, C.P.; Assaf, R. Antibacterial and antiviral activity of camel milk protective proteins. J. Dairy Res. 1992, 59, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wan, J.; Roginski, H.; Lee, A.; Shiell, B.; Michalski, W.P.; Coventry, M.J. Comparison of the effects of acylation and amidation on the antimicrobial and antiviral properties of lactoferrin. Lett. Appl. Microbiol. 2007, 44, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Oda, H.; Yamauchi, K.; Abe, F. Lactoferrin for prevention of common viral infections. J. Infect. Chemother. 2014, 20, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.C.; Swart, P.J.; de Béthune, M.P.; Pauwels, R.; de Clercq, E.; The, T.H.; Meijer, D.K. Antiviral effects of plasma and milk proteins: Lactoferrin shows potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J. Infect. Dis. 1995, 172, 380–388. [Google Scholar] [CrossRef] [PubMed]
- EL-Fakharany, E.M.; Sánchez, L.; Al-Mehdar, H.A.; Redwan, E.M. Effectiveness of human, camel, bovine and sheep lactoferrin on the hepatitis C virus cellular infectivity: Comparison study. Virol. J. 2013, 10, 199. [Google Scholar] [CrossRef] [PubMed]
- Ordaz-Pichardo, C.; Leon-Sicairos, N.; Canizales-Román, A.; Cornejo-Cortés, M.; Ortiz-Estrada, G.; de la Garza, M. Lactoferrin: A protein of the innate immune system capable of killing parasitic protozoa. In Parasites: Ecology, Diseases and Management; Erzinger, G.S., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2013; pp. 177–213. [Google Scholar]
- Tomita, M.; Wakabayashi, H.; Shin, K.; Yamauchi, K.; Yaeshima, T.; Iwatsuki, K. Twenty-five years of research on bovine lactoferrin applications. Biochimie 2009, 91, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Leon-Sicairos, N.; Reyes-López, M.; Ordaz-Pichardo, C.; de la Garza, M. Microbicidal action of lactoferrin and lactoferricin and their synergistic effect with metronidazole in Entamoeba histolytica. Biochem. Cell Biol. 2006, 84, 327–336. [Google Scholar] [PubMed]
- Ordaz-Pichardo, C.; Leon-Sicairos, N.; Hernández-Ramírez, V.; Talamás-Rohana, P.; de la Garza, M. Effect of bovine lactoferrin in a therapeutic hamster model of hepatic amoebiasis. Biochem. Cell Biol. 2012, 90, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Coote, P.J. Bactericidal synergy of lysostaphin in combination with antimicrobial peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-W.; Campbell, L.T.; Wilkins, M.R.; Pang, C.N.I.; Chen, S.; Carter, D.A. Synergy and antagonism between iron chelators and antifungal drugs in Cryptococcus. Int. J. Antimicrob. Agents 2016, 48, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Vaerman, J.P. Effector mechanisms of IgA. Ann. Biol. Clin. 1984, 42, 61–70. [Google Scholar]
- León-Sicairos, N.; López-Soto, F.; Reyes-López, M.; Godínez-Vargas, D.; Ordaz-Pichardo, C.; de la Garza, M. Amoebicidal activity of milk, apo-lactoferrin, slgA and lysozyme. Clin. Med. Res. 2006, 4, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.S.; Watts, J.L. Enhancement of the Activity of Novobiocin Against Escherichia coli by Lactoferrin. J. Dairy Sci. 1999, 82, 494–499. [Google Scholar] [CrossRef]
- Luna-Castro, S.; Aguilar-Romero, F.; Samaniego-Barrón, L.; Godínez-Vargas, D.; de la Garza, M. Effect of bovine apo-lactoferrin on the growth and virulence of Actinobacillus pleuropneumoniae. BioMetals 2014, 27, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Zarember, K.A.; Cruz, A.R.; Huang, C.-Y.; Gallin, J.I. Antifungal Activities of Natural and Synthetic Iron Chelators Alone and in Combination with Azole and Polyene Antibiotics against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2009, 53, 2654–2656. [Google Scholar] [CrossRef] [PubMed]
- Troost, F.J.; Steijns, J.; Saris, W.H.; Brummer, R.J. Gastric digestion of bovine lactoferrin in vivo in adults. J. Nutr. 2001, 131, 2101–2104. [Google Scholar] [PubMed]
- Suzuki, Y.A.; Lönnerdal, B. Characterization of mammalian receptors for lactoferrin. Biochem. Cell Biol. 2002, 80, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Human Milk: Bioactive Proteins/Peptides and Functional Properties. In Nestle Nutrition Institute Workshop Series; Nestec Ltd., Vevey/S. Karger AG: Basel, Switzerland, 2016; Volume 86, pp. 97–107. [Google Scholar]
- Oo, T.Z.; Cole, N.; Garthwaite, L.; Willcox, M.D.P.; Zhu, H. Evaluation of synergistic activity of bovine lactoferricin with antibiotics in corneal infection. J. Antimicrob. Chemother. 2010, 65, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-W.; Ho, S.-P.; Shyu, C.-L.; Mao, F.C. Effects of bovine lactoferrin hydrolysate on the in vitro antimicrobial susceptibility of Escherichia coli strains isolated from baby pigs. Am. J. Vet. Res. 2004, 65, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gomez, S.; Japelj, B.; Jerala, R.; Moriyon, I.; Fernandez Alonso, M.; Leiva, J.; Blondelle, S.E.; Andra, J.; Brandenburg, K.; Lohner, K.; et al. Structural Features Governing the Activity of Lactoferricin-Derived Peptides That Act in Synergy with Antibiotics against Pseudomonas aeruginosa In Vitro and In Vivo. Antimicrob. Agents Chemother. 2011, 55, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Bolscher, J.G.M.; Adão, R.; Nazmi, K.; van den Keybus, P.A.M.; van’t Hof, W.; Nieuw Amerongen, A.V.; Bastos, M.; Veerman, E.C.I. Bactericidal activity of LFchimera is stronger and less sensitive to ionic strength than its constituent lactoferricin and lactoferrampin peptides. Biochimie 2009, 91, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Flores-Villaseñor, H.; Canizalez-Román, A.; Reyes-Lopez, M.; Nazmi, K.; de la Garza, M.; Zazueta-Beltrán, J.; León-Sicairos, N.; Bolscher, J.G.M. Bactericidal effect of bovine lactoferrin, LFcin, LFampin and LFchimera on antibiotic-resistant Staphylococcus aureus and Escherichia coli. Biometals 2010, 23, 569–578. [Google Scholar] [CrossRef] [PubMed]
- López-Soto, F.; León-Sicairos, N.; Nazmi, K.; Bolscher, J.G.; de la Garza, M. Microbicidal effect of the lactoferrin peptides lactoferricin17-30, lactoferrampin265-284, and lactoferrin chimera on the parasite Entamoeba histolytica. Biometals 2010, 23, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Omata, Y.; Satake, M.; Maeda, R.; Saito, A.; Shimazaki, K.; Yamauchi, K.; Uzuka, Y.; Tanabe, S.; Sarashina, T.; Mikami, T. Reduction of the infectivity of Toxoplasma gondii and Eimeria stiedai sporozoites by treatment with bovine lactoferricin. J. Vet. Med. Sci. 2001, 63, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Díaz, H.; Canizalez-Roman, A.; Nepomuceno-Mejia, T.; Gallardo-Vera, F.; Hornelas-Orozco, Y.; Nazmi, K.; Bolscher, J.G.M.; Carrero, C.; Leon-Sicairos, C.; Leon-Sicairos, N. Parasiticidal effect of synthetic bovine Lactoferrin peptides on the enteric parasite Giardia intestinalis. Biochem. Cell Biol. 2016, 95, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Xiong, W.; Hu, Q.; Zuo, P.; Shao, B.; Lan, F.; Lu, X.; Xu, Y.; Xiong, S. Lactoferrin-derived peptides and Lactoferricin chimera inhibit virulence factor production and biofilm formation in Pseudomonas aeruginosa. J. Appl. Microbiol. 2010, 109, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.M.L.; Schryvers, A.B. Perspectives on interactions between lactoferrin and bacteria. Biochem. Cell Biol. 2006, 84, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Stowell, K.M.; Rado, T.A.; Funk, W.D.; Tweedie, J.W. Expression of cloned human lactoferrin in baby-hamster kidney cells. Biochem. J. 1991, 349–355. [Google Scholar] [CrossRef]
- Xiaonan, P.; Xiao, H.; Xuan, W.; Xiwen, C.; Jia, L.; Defu, C. Research progress in physicochemical characteristics of lactoferrin and its recombinant expression systems. Yi Chuan 2015, 37, 873–884. [Google Scholar] [PubMed]
- Suzuki, Y.A.; Kelleher, S.L.; Yalda, D.; Wu, L.; Huang, J.; Huang, N.; Lönnerdal, B. Expression, characterization, and biologic activity of recombinant human lactoferrin in rice. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Recombinant human milk proteins. Nestlé Nutr. Work. Ser. Paediatr. Program. 2006, 58, 207–215. [Google Scholar]
- Yemets, A.I.; Tanasienko, I.V.; Krasylenko, Y.A.; Blume, Y.B. Plant-based biopharming of recombinant human lactoferrin. Cell Biol. Int. 2014, 38, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, W.; Zhao, J.; Wang, J.; Dai, Y.; Zhao, Y.; Meng, Q.; Li, N. Supplementation transgenic cow’s milk containing recombinant human lactoferrin enhances systematic and intestinal immune responses in piglets. Mol. Biol. Rep. 2014, 41, 2119–2128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, J.; Wang, J.; Li, N. Nutritional composition analysis of meat from human lactoferrin transgenic bulls. Anim. Biotechnol. 2013, 24, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Chen, Y.; Wang, H.Y.; Wang, R.-F. Mechanisms and pathways of innate immune activation and regulation in health and cancer. Hum. Vaccin. Immunother. 2014, 10, 3270–3285. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Lawrence, T.; Nizet, V. Innate immunity gone awry: Linking microbial infections to chronic inflammation and cancer. Cell 2006, 124, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.M.; Neish, A.S. Recognition of bacterial pathogens and mucosal immunity. Cell. Microbiol. 2011, 13, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.-D. Converging roles of caspases in inflammasome activation, cell death and innate immunity. Nat. Rev. Immunol. 2016, 16, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Elass, E.; Carpentier, M.; Mazurier, J. Interactions of lactoferrin with cells involved in immune functionThis paper is one of a selection of papers published in this Special Issue, entitled 7th International Conference on Lactoferrin: Structure, Function, and Applications, and has undergone TH. Biochem. Cell Biol. 2006, 84, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Jiang, R.; Du, X. Bovine lactoferrin can be taken up by the human intestinal lactoferrin receptor and exert bioactivities. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.A.; Wong, H.; Ashida, K.-Y.; Schryvers, A.B.; Lönnerdal, B. The N1 domain of human lactoferrin is required for internalization by Caco-2 cells and targeting to the nucleus. Biochemistry 2008, 47, 10915–10920. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.A.; Lopez, V.; Lönnerdal, B. Mammalian lactoferrin receptors: Structure and function. Cell. Mol. Life Sci. 2005, 62, 2560–2575. [Google Scholar] [CrossRef] [PubMed]
- Gómez, H.F.; Ochoa, T.J.; Herrera-Insua, I.; Carlin, L.G.; Cleary, T.G. Lactoferrin protects rabbits from Shigella flexneri-induced inflammatory enteritis. Infect. Immun. 2002, 70, 7050–7053. [Google Scholar] [CrossRef] [PubMed]
- Mosquito, S.; Ochoa, T.J.; Cok, J.; Cleary, T.G. Effect of bovine lactoferrin in Salmonella ser. Typhimurium infection in mice. Biometals 2010, 23, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Berlutti, F.; Schippa, S.; Morea, C.; Sarli, S.; Perfetto, B.; Donnarumma, G.; Valenti, P. Lactoferrin downregulates pro-inflammatory cytokines upexpressed in intestinal epithelial cells infected with invasive or noninvasive Escherichia coli strains. Biochem. Cell Biol. 2006, 84, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Bertuccini, L.; Costanzo, M.; Iosi, F.; Tinari, A.; Terruzzi, F.; Stronati, L.; Aloi, M.; Cucchiara, S.; Superti, F. Lactoferrin prevents invasion and inflammatory response following E. coli strain LF82 infection in experimental model of Crohn’s disease. Dig. Liver Dis. 2014, 46, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; di Patti, M.C.B.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; Puddu, P.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. Biometals 2014, 27, 843–856. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Martínez-Pardo, L.; Sánchez-Hernández, B.; de la Garza, M.; Carrero, J.C. Oral lactoferrin treatment resolves amoebic intracecal infection in C3H/HeJ mice. Biochem. Cell Biol. 2012, 90, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hirmo, S.; Willén, R.; Wadström, T. Inhibition of Helicobacter pylori infection by bovine milk glycoconjugates in a BAlb/cA mouse model. J. Med. Microbiol. 2001, 50, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.-H.; Chiu, C.-H.; Huang, Y.-C.; Lin, T.-Y. Effects of lactoferrin-containing formula in the prevention of enterovirus and rotavirus infection and impact on serum cytokine levels: A randomized trial. Chang Gung Med. J. 2011, 34, 395–402. [Google Scholar] [PubMed]
- Flores-Villasenor, H.; Canizalez-Roman, A.; Velazquez-Roman, J.; Nazmi, K.; Bolscher, J.G.; Leon-Sicairos, N. Protective effects of lactoferrin chimera and bovine lactoferrin in a mouse model of enterohaemorrhagic Escherichia coli O157:H7 infection. Biochem. Biol. 2012, 411, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Artym, J.; Chodaczek, G.; Kocieba, M.; Kruzel, M.L. Protective effects of lactoferrin in Escherichia coli-induced bacteremia in mice: Relationship to reduced serum TNF α level and increased turnover of neutrophils. Inflamm. Res. 2004, 53, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Artym, J.; Zimecki, M.; Kruzel, M.L. Enhanced clearance of Escherichia coli and Staphylococcus aureus in mice treated with cyclophosphamide and lactoferrin. Int. Immunopharmacol. 2004, 4, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Kruzel, M.L.; Harari, Y.; Mailman, D.; Actor, J.K.; Zimecki, M. Differential effects of prophylactic, concurrent and therapeutic lactoferrin treatment on LPS-induced inflammatory responses in mice. Clin. Exp. Immunol. 2002, 130, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.H.; Mann, D.M.; Tsai, C.M. Neutralization of endotoxin in vitro and in vivo by a human lactoferrin-derived peptide. Infect. Immun. 1999, 67, 1353–1358. [Google Scholar] [PubMed]
- Hwang, S.-A.; Kruzel, M.L.; Actor, J.K. Lactoferrin augments BCG vaccine efficacy to generate T helper response and subsequent protection against challenge with virulent Mycobacterium tuberculosis. Int. Immunopharmacol. 2005, 5, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-A.; Wilk, K.M.; Budnicka, M.; Olsen, M.; Bangale, Y.A.; Hunter, R.L.; Kruzel, M.L.; Actor, J.K. Lactoferrin enhanced efficacy of the BCG vaccine to generate host protective responses against challenge with virulent Mycobacterium tuberculosis. Vaccine 2007, 25, 6730–6743. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-A.; Kruzel, M.L.; Actor, J.K. CHO expressed recombinant human lactoferrin as an adjuvant for BCG. Int. J. Immunopathol. Pharmacol. 2015, 28, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Welsh, K.J.; Hwang, S.-A.; Boyd, S.; Kruzel, M.L.; Hunter, R.L.; Actor, J.K. Influence of oral lactoferrin on Mycobacterium tuberculosis induced immunopathology. Tuberculosis 2011, 91, S105–S113. [Google Scholar] [CrossRef] [PubMed]
- Valenti, P.; Catizone, A.; Pantanella, F.; Frioni, A.; Natalizi, T.; Tendini, M.; Berlutti, F. Lactoferrin decreases inflammatory response by cystic fibrosis bronchial cells invaded with Burkholderia cenocepacia iron-modulated biofilm. Int. J. Immunopathol. Pharmacol. 2011, 24, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, D.; Chen, H.; Pan, X.; Kong, Q.; Pang, Q. Lactoferrin protects against lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2012, 12, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Gualdi, L.; Mertz, S.; Gomez, A.M.; Ramilo, O.; Wittke, A.; Mejias, A. Lack of effect of bovine lactoferrin in respiratory syncytial virus replication and clinical disease severity in the mouse model. Antivir. Res. 2013, 99, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Welliver, R.C.; Hintz, K.M.; Shalwitz, R.M. Lactoferrin reverses respiratory abnormalities in respiratory syncytial virus (RSV) infection of mice. Pediatric Res. 1999, 45, 178A. [Google Scholar] [CrossRef]
- Shin, K.; Wakabayashi, H.; Yamauchi, K.; Teraguchi, S.; Tamura, Y.; Kurokawa, M.; Shiraki, K. Effects of orally administered bovine lactoferrin and lactoperoxidase on influenza virus infection in mice. J. Med. Microbiol. 2005, 54, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Komine, Y.; Komine, K.; Asai, K.; Kuroishi, T.; Kozutsumi, T.; Itagaki, M.; Ohta, M.; Kumagai, K. Effects of bovine lactoferrin by the intramammary infusion in cows with Staphylococcal mastitis during the early non-lactating period. J. Vet. Med. Sci. 2002, 64, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Komine, Y.; Komine, K.-I.; Kai, K.; Itagaki, M.; Kuroishi, T.; Aso, H.; Obara, Y.; Kumagai, K. Effect of combination therapy with lactoferrin and antibiotics against staphylococcal mastitis on drying cows. J. Vet. Med. Sci. 2006, 68, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Komine, K.; Komine, Y.; Kuroishi, T.; Kobayashi, J.; Obara, Y.; Kumagai, K. Small Molecule Lactoferrin with an Inflammatory Effect But No Apparent Antibacterial Activity in Mastitic Mammary Gland Secretion. J. Vet. Med. Sci. 2005, 67, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Komine, Y.; Kuroishi, T.; Kobayashi, J.; Aso, H.; Obara, Y.; Kumagai, K.; Sugawara, S.; Komine, K. Inflammatory effect of cleaved bovine lactoferrin by elastase on staphylococcal mastitis. J. Vet. Med. Sci. 2006, 68, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Takakura, N.; Wakabayashi, H.; Ishibashi, H.; Yamauchi, K.; Teraguchi, S.; Tamura, Y.; Yamaguchi, H.; Abe, S. Effect of orally administered bovine lactoferrin on the immune response in the oral candidiasis murine model. J. Med. Microbiol. 2004, 53, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Park, J.-H.; Seok, S.-H.; Baek, M.-W.; Kim, D.-J.; Lee, B.-H.; Kang, P.-D.; Kim, Y.-S.; Park, J.-H. Potential antimicrobial effects of human lactoferrin against oral infection with Listeria monocytogenes in mice. J. Med. Microbiol. 2005, 54, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Håversen, L.A.; Engberg, I.; Baltzer, L.; Dolphin, G.; Hanson, L.A.; Mattsby-Baltzer, I. Human lactoferrin and peptides derived from a surface-exposed helical region reduce experimental Escherichia coli urinary tract infection in mice. Infect. Immun. 2000, 68, 5816–5823. [Google Scholar] [CrossRef] [PubMed]
- Guillén, C.; McInnes, I.B.; Vaughan, D.M.; Kommajosyula, S.; van Berkel, P.H.C.; Leung, B.P.; Aguila, A.; Brock, J.H. Enhanced Th1 response to Staphylococcus aureus infection in human lactoferrin-transgenic mice. J. Immunol. 2002, 168, 3950–3957. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.; Pallone, F.; Monteleone, G.; Fantini, M.C. Intestinal inflammation and colorectal cancer: A double-edged sword? World J. Gastroenterol. 2011, 17, 3092–3100. [Google Scholar] [PubMed]
- Petschow, B.W.; Talbott, R.D.; Batema, R.P. Ability of lactoferrin to promote the growth of Bifidobacterium spp. in vitro is independent of receptor binding capacity and iron saturation level. J. Med. Microbiol. 1999, 48, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Oda, H.; Wakabayashi, H.; Yamauchi, K.; Abe, F. Lactoferrin and bifidobacteria. Biometals 2014, 27, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Dial, E.J.; Lichtenberger, L.M. Effect of lactoferrin on Helicobacter felis induced gastritis. Biochem. Cell Biol. 2002, 80, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H.Q.; Campbell, M.A.F.; Couper, R.T.L.; Tran, C.D.; Lawrence, A.; Butler, R.N. Lactoferrin and desferrioxamine are ineffective in the treatment of Helicobacter pylori infection and may enhance H. pylori growth and gastric inflammation in mice. Lett. Appl. Microbiol. 2009, 48, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Guttner, Y.; Windsor, H.M.; Viiala, C.H.; Marshall, B.J. Human recombinant lactoferrin is ineffective in the treatment of human Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2003, 17, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Wu, Q.; Cheng, G.; Liu, X.; Liu, S.; Luo, J.; Zhang, A.; Bian, L.; Chen, J.; Lv, J.; et al. Recombinant human lactoferrin enhances the efficacy of triple therapy in mice infected with Helicobacter pylori. Int. J. Mol. Med. 2015, 36, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B. Multi-faceted functions of secretory IgA at mucosal surfaces. Front. Immunol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Drago-Serrano, M.E.; Rivera-Aguilar, V.; Reséndiz-Albor, A.A.; Campos-Rodríguez, R. Lactoferrin increases both resistance to Salmonella typhimurium infection and the production of antibodies in mice. Immunol. Lett. 2010, 134, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Negroni, A.; Costanzo, M.; Vitali, R.; Superti, F.; Bertuccini, L.; Tinari, A.; Minelli, F.; di Nardo, G.; Nuti, F.; Pierdomenico, M.; et al. Characterization of adherent-invasive Escherichia coli isolated from pediatric patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Carrero, J.C.; Contreras-Rojas, A.; Sánchez-Hernández, B.; Petrosyan, P.; Bobes, R.J.; Ortiz-Ortiz, L.; Laclette, J.P. Protection against murine intestinal amoebiasis induced by oral immunization with the 29 kDa antigen of Entamoeba histolytica and cholera toxin. Exp. Parasitol. 2010, 126, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Johansen, F.-E.; Brandtzaeg, P. Transcriptional regulation of the mucosal IgA system. Trends Immunol. 2004, 25, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Schulte, W.; Bernhagen, J.; Bucala, R. Cytokines in sepsis: Potent immunoregulators and potential therapeutic targets—An updated view. Mediat. Inflamm. 2013, 2013, 165974. [Google Scholar] [CrossRef] [PubMed]
- Weng, T.-Y.; Chen, L.-C.; Shyu, H.-W.; Chen, S.-H.; Wang, J.-R.; Yu, C.-K.; Lei, H.-Y.; Yeh, T.-M. Lactoferrin inhibits enterovirus 71 infection by binding to VP1 protein and host cells. Antivir. Res. 2005, 67, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kvistgaard, A.S.; Pallesen, L.T.; Arias, C.F.; López, S.; Petersen, T.E.; Heegaard, C.W.; Rasmussen, J.T. Inhibitory effects of human and bovine milk constituents on rotavirus infections. J. Dairy Sci. 2004, 87, 4088–4096. [Google Scholar] [CrossRef]
- Pérez-Cano, F.J.; Marín-Gallén, S.; Castell, M.; Rodríguez-Palmero, M.; Rivero, M.; Castellote, C.; Franch, A. Supplementing suckling rats with whey protein concentrate modulates the immune response and ameliorates rat rotavirus-induced diarrhea. J. Nutr. 2008, 138, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Deutschman, C.S.; Tracey, K.J. Sepsis: Current dogma and new perspectives. Immunity 2014, 40, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Keestra, A.M.; Tsolis, R.M.; Bäumler, A.J. The blessings and curses of intestinal inflammation. Cell Host Microbe 2010, 8, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Edde, L.; Hipolito, R.B.; Hwang, F.F.; Headon, D.R.; Shalwitz, R.A.; Sherman, M.P. Lactoferrin protects neonatal rats from gut-related systemic infection. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1140–G1150. [Google Scholar] [PubMed]
- Zagulski, T.; Lipiński, P.; Zagulska, A.; Broniek, S.; Jarzabek, Z. Lactoferrin can protect mice against a lethal dose of Escherichia coli in experimental infection in vivo. Br. J. Exp. Pathol. 1989, 70, 697–704. [Google Scholar] [PubMed]
- Zagulski, T.; Lipinski, P.; Zagulska, A.; Jarzabek, Z. Antibacterial system generated by lactoferrin in mice in vivo is primarily a killing system. Int. J. Exp. Pathol. 1998, 79, 117–123. [Google Scholar] [PubMed]
- Akin, I.M.; Atasay, B.; Dogu, F.; Okulu, E.; Arsan, S.; Karatas, H.D.; Ikinciogullari, A.; Turmen, T. Oral lactoferrin to prevent nosocomial sepsis and necrotizing enterocolitis of premature neonates and effect on T-regulatory cells. Am. J. Perinatol. 2014, 31, 1111–1120. [Google Scholar] [PubMed]
- Drago-Serrano, M.E.; de la Garza-Amaya, M.; Luna, J.S.; Campos-Rodríguez, R. Lactoferrin-lipopolysaccharide (LPS) binding as key to antibacterial and antiendotoxic effects. Int. Immunopharmacol. 2012, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Latorre, D.; Puddu, P.; Valenti, P.; Gessani, S. Reciprocal interactions between lactoferrin and bacterial endotoxins and their role in the regulation of the immune response. Toxins 2010, 2, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Talukder, M.J.R.; Harada, E. Bovine lactoferrin protects lipopolysaccharide-induced diarrhea modulating nitric oxide and prostaglandin E2 in mice. Can. J. Physiol. Pharmacol. 2007, 85, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Kuhara, T.; Tanaka, A.; Yamauchi, K.; Iwatsuki, K. Bovine lactoferrin ingestion protects against inflammation via IL-11 induction in the small intestine of mice with hepatitis. Br. J. Nutr. 2014, 111, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- Nebermann, L.; Döhler, J.R.; Perlick, L. Treatment of enterogenic endotoxinemia with lactoferrin in rats. Langenbeck Arch. Surg. 2001, 386, 146–149. [Google Scholar] [CrossRef]
- Kruzel, M.L.; Harari, Y.; Chen, C.Y.; Castro, G.A. Lactoferrin protects gut mucosal integrity during endotoxemia induced by lipopolysaccharide in mice. Inflammation 2000, 24, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Hu, W.; Song, D.; Li, Z.; Du, H.; Lu, Z.; Wang, Y. Porcine lactoferrin-derived peptide LFP-20 protects intestinal barrier by maintaining tight junction complex and modulating inflammatory response. Biochem. Pharmacol. 2016, 104, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Song, D.; Wang, T.; Xia, X.; Hu, W.; Han, F.; Wang, Y. LFP-20, a porcine lactoferrin peptide, ameliorates LPS-induced inflammation via the MyD88/NF-κB and MyD88/MAPK signaling pathways. Dev. Comp. Immunol. 2015, 52, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Hirotani, Y.; Ikeda, K.; Kato, R.; Myotoku, M.; Umeda, T.; Ijiri, Y.; Tanaka, K. Protective Effects of Lactoferrin against Intestinal Mucosal Damage Induced by Lipopolysaccharide in Human Intestinal Caco-2 Cells. Yakugaku Zasshi 2008, 128, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, E.A.; Duffy, L.C.; Schanbacher, F.L.; Qiao, H.; Dryja, D.; Leavens, A.; Rossman, J.; Rich, G.; Dirienzo, D.; Ogra, P.L. In vivo effects of bifidobacteria and lactoferrin on gut endotoxin concentration and mucosal immunity in BALB/c mice. Dig. Dis. Sci. 2004, 49, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Farmer, J.L.; Hilty, M.; Kim, Y.B. The protective effects of lactoferrin feeding against endotoxin lethal shock in germfree piglets. Infect. Immun. 1998, 66, 1421–1426. [Google Scholar] [PubMed]
- Guerville, M.; Boudry, G. Gastrointestinal and hepatic mechanisms limiting entry and dissemination of lipopolysaccharide into the systemic circulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G1–G15. [Google Scholar] [CrossRef] [PubMed]
- Glant, T.T.; Bárdos, T.; Vermes, C.; Chandrasekaran, R.; Valdéz, J.C.; Otto, J.M.; Gerard, D.; Velins, S.; Lovász, G.; Zhang, J.; et al. Variations in susceptibility to proteoglycan-induced arthritis and spondylitis among C3H substrains of mice: Evidence of genetically acquired resistance to autoimmune disease. Arthritis Rheum. 2001, 44, 682–692. [Google Scholar] [CrossRef]
- Zimecki, M.; Chodaczek, G.; Kocieba, M.; Kruzel, M.L. Lethality in LPS-induced endotoxemia in C3H/HeCr mice is associated with prevalence of proinflammatory cytokines: Lack of protective action of lactoferrin. FEMS Immunol. Med. Microbiol. 2004, 42, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Kuhara, T.; Yamauchi, K.; Iwatsuki, K. Bovine lactoferrin induces interleukin-11 production in a hepatitis mouse model and human intestinal myofibroblasts. Eur. J. Nutr. 2012, 51, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Matsuura, M.; Kobayashi, K.; Sasaki, H.; Yajima, T.; Kuwata, T. Lactoferrin protects against development of hepatitis caused by sensitization of Kupffer cells by lipopolysaccharide. Clin. Diagn. Lab. Immunol. 2001, 8, 1234–1239. [Google Scholar] [CrossRef] [PubMed]
- Cutone, A.; Frioni, A.; Berlutti, F.; Valenti, P.; Musci, G.; Bonaccorsi di Patti, M.C. Lactoferrin prevents LPS-induced decrease of the iron exporter ferroportin in human monocytes/macrophages. Biometals 2014, 27, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Puddu, P.; Latorre, D.; Carollo, M.; Catizone, A.; Ricci, G.; Valenti, P.; Gessani, S. Bovine lactoferrin counteracts Toll-like receptor mediated activation signals in antigen presenting cells. PLoS ONE 2011, 6, e22504. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.J.; Han, S.B.; Kang, J.S.; Yoon, Y.D.; Park, S.-K.; Kim, H.M.; Yang, K.-H.; Joe, C.O. Lactoferrin works as a new LPS-binding protein in inflammatory activation of macrophages. Int. Immunopharmacol. 2004, 4, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Curran, C.S.; Demick, K.P.; Mansfield, J.M. Lactoferrin activates macrophages via TLR4-dependent and -independent signaling pathways. Cell. Immunol. 2006, 242, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Håversen, L.; Ohlsson, B.G.; Hahn-Zoric, M.; Hanson, L.Å.; Mattsby-Baltzer, I. Lactoferrin down-regulates the LPS-induced cytokine production in monocytic cells via NF-κB. Cell. Immunol. 2002, 220, 83–95. [Google Scholar] [CrossRef]
- Whitsett, J.A.; Alenghat, T. Respiratory epithelial cells orchestrate pulmonary innate immunity. Nat. Immunol. 2014, 16, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.; Mateos, S. Infecciones respiratorias. In Temas de Bacteriología y Virología Médica; Oficina del libro FEFMUR (Fundacion de Ediciones de la Facultad de Medicina de la Universidad de la República): Montevideo, Uruguay, 2006; pp. 137–161. [Google Scholar]
- Tavares, L.P.; Teixeira, M.M.; Garcia, C.C. The inflammatory response triggered by Influenza virus: A two edged sword. Inflamm. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Acıoğlu, E.; Yigit, O.; Alkan, Z.; Azizli, E.; Gelişgen, R.; Uzun, H. The effects of corticosteroid on tissue lactoferrin in patients with nasal polyposis. Am. J. Rhinol. Allergy 2012, 26, e28–e31. [Google Scholar] [CrossRef] [PubMed]
- Maher, D.; Raviglione, M. Global Epidemiology of Tuberculosis. Clin. Chest Med. 2005, 26, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Behr, M.A. BCG—Different strains, different vaccines? Lancet Infect. Dis. 2002, 2, 86–92. [Google Scholar] [CrossRef]
- Hwang, S.-A.; Kruzel, M.L.; Actor, J.K. Influence of bovine lactoferrin on expression of presentation molecules on BCG-infected bone marrow derived macrophages. Biochimie 2009, 91, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-A.; Wilk, K.; Kruzel, M.L.; Actor, J.K. A novel recombinant human lactoferrin augments the BCG vaccine and protects alveolar integrity upon infection with Mycobacterium tuberculosis in mice. Vaccine 2009, 27, 3026–3034. [Google Scholar] [CrossRef] [PubMed]
- Rogan, M.P.; Taggart, C.C.; Greene, C.M.; Murphy, P.G.; O’Neill, S.J.; McElvaney, N.G. Loss of microbicidal activity and increased formation of biofilm due to decreased lactoferrin activity in patients with cystic fibrosis. J. Infect. Dis. 2004, 190, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Britigan, B.E.; Hayek, M.B.; Doebbeling, B.N.; Fick, R.B. Transferrin and lactoferrin undergo proteolytic cleavage in the Pseudomonas aeruginosa-infected lungs of patients with cystic fibrosis. Infect. Immun. 1993, 61, 5049–5055. [Google Scholar] [PubMed]
- Portelli, J.; Gordon, A.; May, J.T. Effect of compounds with antibacterial activities in human milk on respiratory syncytial virus and cytomegalovirus in vitro. J. Med. Microbiol. 1998, 47, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nagai, K.; Tsutsumi, H.; Kuroki, Y. Lactoferrin and surfactant protein A exhibit distinct binding specificity to F protein and differently modulate respiratory syncytial virus infection. Eur. J. Immunol. 2003, 33, 2894–2902. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, G.V.; Trabattoni, D.; Morelli, M.; Borgonovo, S.; Schneider, L.; Clerici, M. Immune modulation by lactoferrin and curcumin in children with recurrent respiratory infections. J. Biol. Regul. Homeost. Agents 2009, 23, 119–123. [Google Scholar] [PubMed]
- Chen, K.; Chai, L.; Li, H.; Zhang, Y.; Xie, H.-M.; Shang, J.; Tian, W.; Yang, P.; Jiang, A.C. Effect of bovine lactoferrin from iron-fortified formulas on diarrhea and respiratory tract infections of weaned infants in a randomized controlled trial. Nutrition 2016, 32, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Psaltis, A.J.; Wormald, P.-J.; Ha, K.R.; Tan, L.W. Reduced levels of lactoferrin in biofilm-associated chronic rhinosinusitis. Laryngoscope 2008, 118, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Komine, K.-I.; Kuroishi, T.; Ozawa, A.; Komine, Y.; Minami, T.; Shimauchi, H.; Sugawara, S. Cleaved inflammatory lactoferrin peptides in parotid saliva of periodontitis patients. Mol. Immunol. 2007, 44, 1498–1508. [Google Scholar] [CrossRef] [PubMed]
- Ueta, E.; Tanida, T.; Osaki, T. A novel bovine lactoferrin peptide, FKCRRWQWRM, suppresses Candida cell growth and activates neutrophils. J. Pept. Res. 2001, 57, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Longhi, C.; Conte, M.P.; Ranaldi, S.; Penta, M.; Valenti, P.; Tinari, A.; Superti, F.; Seganti, L. Apoptotic death of Listeria monocytogenes-infected human macrophages induced by lactoferricin B, a bovine lactoferrin-derived peptide. Int. J. Immunopathol. Pharmacol. 2005, 18, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Byrd, T.F.; Horwitz, M.A. Lactoferrin inhibits or promotes Legionella pneumophila intracellular multiplication in nonactivated and interferon γ-activated human monocytes depending upon its degree of iron saturation. Iron-lactoferrin and nonphysiologic iron chelates reverse mon. J. Clin. Investig. 1991, 88, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Nocerino, N.; Fulgione, A.; Iannaccone, M.; Tomasetta, L.; Ianniello, F.; Martora, F.; Lelli, M.; Roveri, N.; Capuano, F.; Capparelli, R. Biological activity of lactoferrin-functionalized biomimetic hydroxyapatite nanocrystals. Int. J. Nanomed. 2014, 9, 1175–1184. [Google Scholar]
- Chatterton, D.E.W.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int. J. Biochem. Cell Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef] [PubMed]
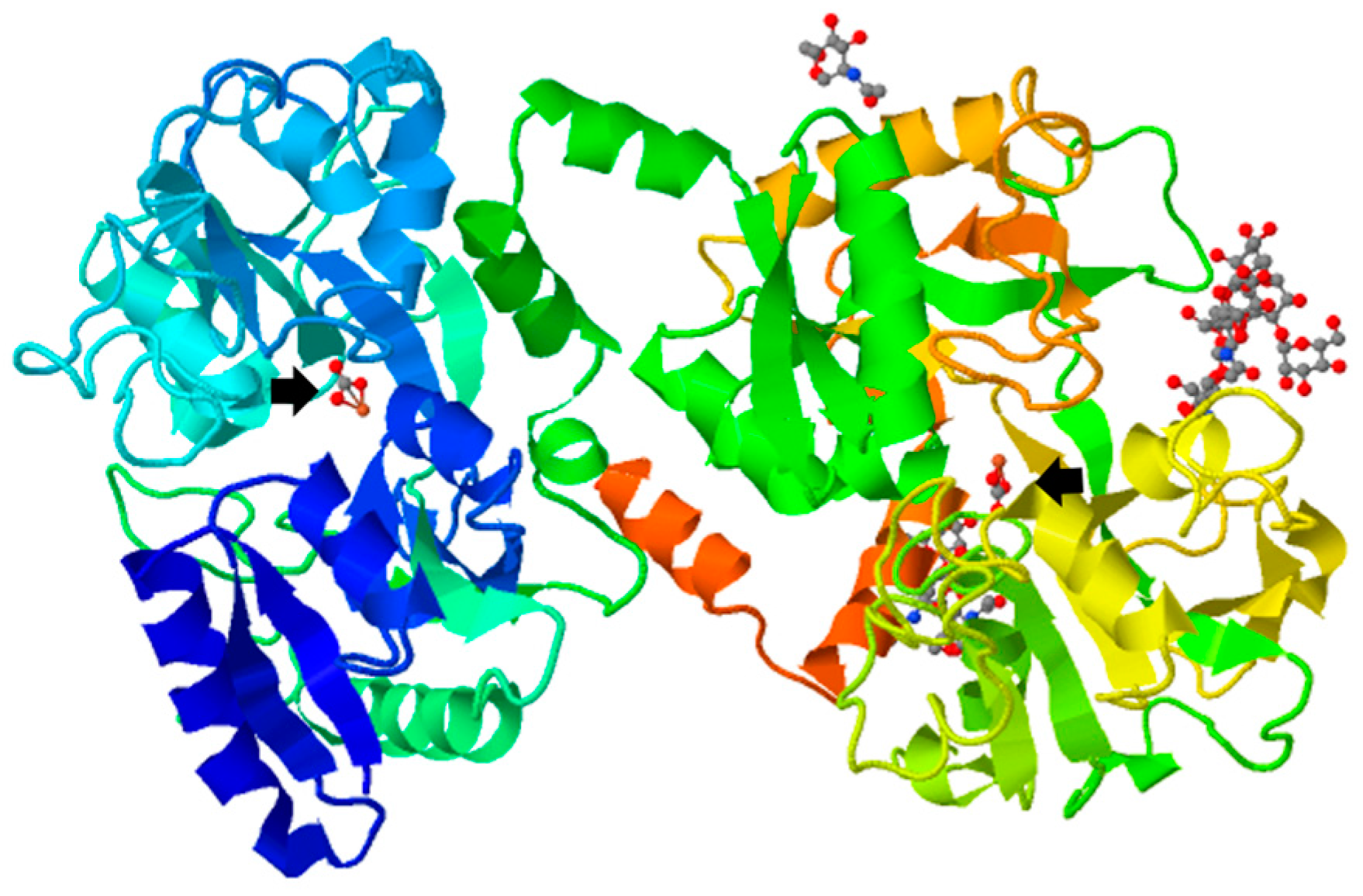

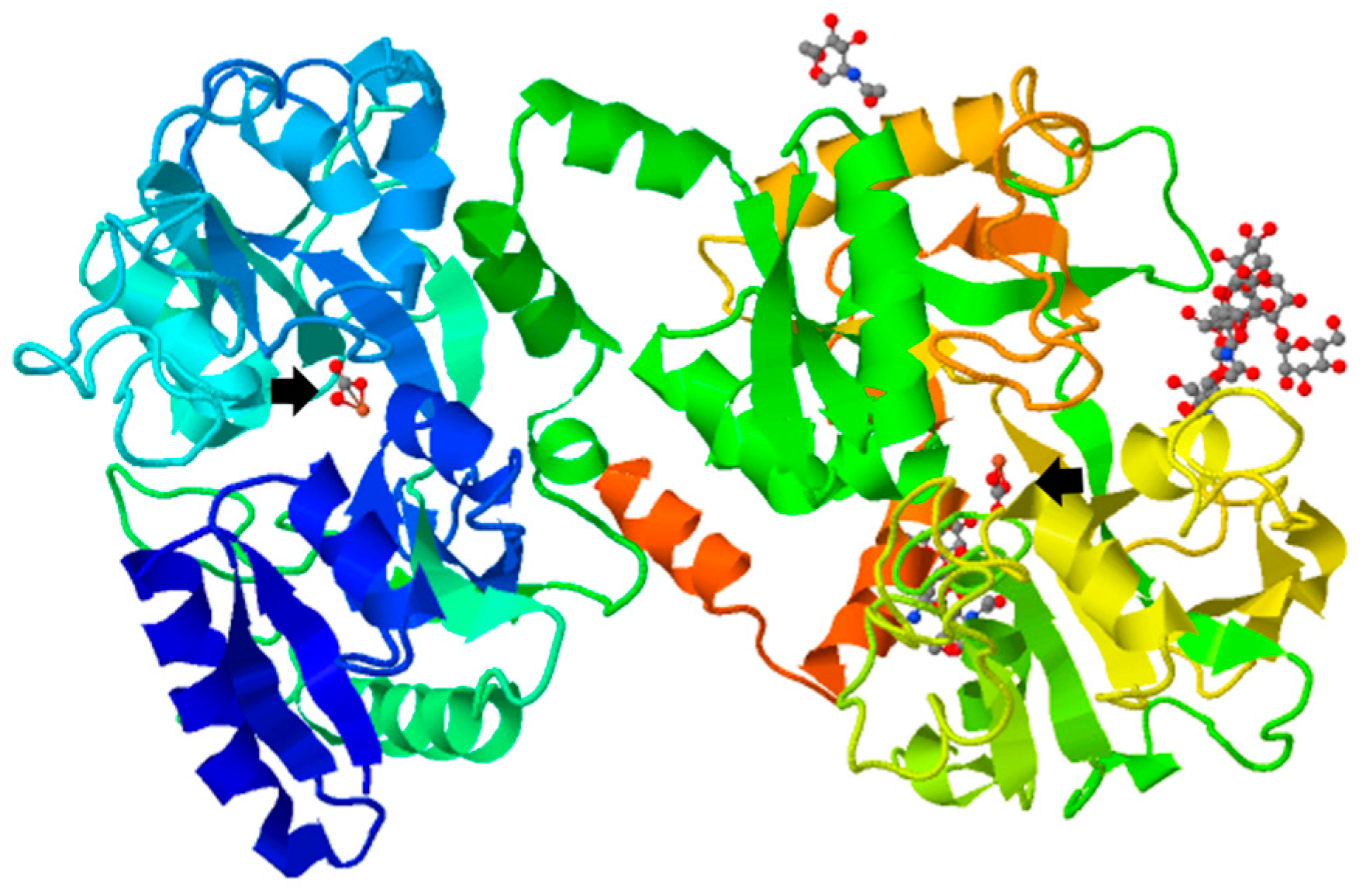{kind=link}
| Model | Finding | Reference |
|---|---|---|
| Gastrointestinal infections | ||
| Bovine Lf (bLf) treatment of rabbits infected with Shigella flexneri (S. flexneri) | ↓Gut inflammation (microscopic observation), ↓submucosal edema, ↓infiltration of leukocytes | [98] |
| bLf treatment of BALB/c mice infected with Salmonella enterica (S. enterica) serovar Typhimurium | ↓Gut inflammation, ↓necrosis | [99] |
| bLf treatment of Caco-2 cell-line cultures infected with Escherichia coli (E. coli) HB101 (non-invasive), or recombinant E. coli HB101 pRI203 (invasive) | ↓IL-8 mRNA expression in Caco-2 cells infected with E. coli HB101; ↓IL-6, ↓IL-8, ↓Tumor Necrosis Factor (TNF)α levels in Caco-2 cells infected with E. coli HB101pRI203 | [100] |
| bLf treatment of cultured cells infected with E. coli LF82 and biopsies from patients with Crohn’s disease | ↓IL-6, ↓IL-8 and ↓TNFα mRNA expression | [101] |
| bLf treatment of intestinal cell cultures infected with E. coli LF82 isolated from Crohn’s disease patients | ↑Ferroportin (Fpn) in infected cells suggesting that bLf action on inflammatory response in epithelial cells involves the iron homeostasis | [102] |
| bLF-nanoparticles (bLf-nano) administration to BALB/c mice infected with S. enterica serovar Typhimurium | ↑TNF1α, ↑Interferon (IFN)1β and ↑IFNIII levels (proinflammatory cytokines) | [6] |
| bLf administration to C3H/HeJ mice infected with Entamoeba histolytica (E. histolytica) | ↑IL-4 (Th2), ↑IL-6, ↑IgA ↓damage and ↓Inflammation | [103] |
| bLf treatment to BALB/c mice infected with Helicobacter pylori (H. pylori) | ↓Gastric colonization and ↓inflammation (histopathology score) | [104] |
| bLf treatment of rotavirus infection children | ↔IFNγ, ↔IL-10 and ↔rotavirus incidence in children whether fed or unfed with bLf | [105] |
| Gut-related systemic infections (sepsis) | ||
| Administration of bLf or synthetic LF chimera peptide to BALB/c mice infected with enterohaemorrhagic E. coli (EHEC) 0157:H7 | ↓Mortality (only LF-chimera); ↓kidney damage; ↓fecal bacterial output and ↓sepsis: bLf and LFchimera | [106] |
| Septicemia | ||
| Single-dose bLf administration 2 or 24 h prior intravenous infection of CBA mice with E. coli O55 strain | ↓Blood TNF1α (24 h), or ↑blood TNFα (2 h) | [107] |
| Lf administrated with drinking water (0.5% solution) during 14 days in mice immunosuppressed by cyclophosphamide treatment, and intravenously infected with E. coli | ↓E. coli bacterial load in spleen and liver, ↑blood neutrophils, ↑IL-6 in spleen and peritoneal cells | [108] |
| Endotoxemia | ||
| Prophylactic or therapeutic effect of bLf on systemic inflammation in mice treated with lipopolysaccharide (LPS) | ↓TNFα, ↓IL-6 and ↓IL-10, 1 h prior to the LPS treatment (prophylactic effect). ↓TNFα, ↓Nitric Oxide (NO), ↔IL-6, ↔IL-10, 18 h prior to the LPS treatment (prophylactic effect). ↓NO and ↓post-shock, 2 h after LPS treatment (therapeutic effect) | [109] |
| LF33 peptide administration to mice treated with LPS and to RAW 264.7 cell-line cultures treated with LPS | ↓Limulus amoebocyte lysate coagulation, ↓TNFα secretion by RAW 264.7 cells induced by LPS, ↓TNF-α levels correlated with protection to lethal LPS-induced septic shock | [110] |
| Respiratory tract infections | ||
| Adjuvant effect of Lf mixed with bacillus Calmette-Guerin (BCG) vaccine on mice infected with Mycobacterium tuberculosis (M. tuberculosis) | ↓Lung infection, ↑IFNγ, ↑IL-12 in spleen cell cultures, ↓TNFα and ↓IL-1β correlated with ↓lung pathology. ↑lymphocytic recall response towards BCG | [111,112] |
| Recombinant human Lf mixed with BCG vaccine in mice infected with M. tuberculosis | Early↑ and late↓ of pro-inflammatory cytokines that correlated with the ↓lung pathology | [113] |
| bLf effect in enhancing BCG vaccine by oral route in mice infected with M. tuberculosis | ↓Colony Forming Units (CFU) and ↓inflammation in the lungs, ↑IFNγ producing T CD4 and CD8 cells and ↑Il-17 lymphocytes | [114] |
| bLf effects on cystic fibrosis and bronchial IB3-1 cell cultures infected with Burkholderia cenocepacia (B. cenocepacia) | ↓IL-1β (pro-inflammatory cytokine), ↓IL-11 (anti-inflammatory cytokine) | [115] |
| bLf administration to a murine model of lung injury by LPS | ↓Bronchioalveolar leukocytes, ↓TNF-α, ↓myeloperoxidase (MPO) activity, ↑IL-10, ↓lung edema and inflammation | [116] |
| bLf administration to a murine model of respiratory syncytial virus infection | ↔Viral loads and ↔lung inflammation | [117,118] |
| bLf administration to a murine model of influenza | ↔Viral load and ↔IFNγ, IL-6 and IL-12 in the lungs | [119] |
| Other mucosal and systemic sites | ||
| bLf effects on mammary gland in cows with Staphylococcous aureus (S. aureus) mastitis | ↓Bacterial load, ↑C3 levels, ↓TNFα mRNA expression via Nuclear Factor κB (NFκB) inhibition, ↑curation, ↑proinflammatory cytokines is correlated with ↑peptides derived from bLf-elastase proteolysis | [120,121,122,123] |
| bLf effects on oral candidiasis in immunosuppressed mice infected with Candida albicans (C. albicans) | bLf blocked the suppressive effects of candidiasis in Polymorphonuclear (PMN) neutrophils; ↑IFNγ and TNFα production in cervical lymph nodes | [124] |
| bLf effects on hamsters with amoebic liver abscess by E. histolytica | No damage or inflammation in the liver | [62] |
| Human Lf (hLf) effects on BALB/c mice infected with Listeria monocytogenes (L. monocytogenes) | ↓Bacterial load and ↓necrotic foci in the liver, ↔necrotic foci in the spleen, ↓TNFα, IL-1β and IFNγ mRNA | [125] |
| hLf and peptide-hLf derivatives administration to C3H/TiF mice infected with E. coli O6K5 uropathogenic strain | ↓Bacterial load in the bladder and kidneys, ↓leukocyte in urine, ↓urinary IL-6 levels at 2 h and systemic IL-6 levels at 24 h post-infection | [126] |
| hLf expressing transgenic mice infected with S. aureus | ↓Bacterial growth, ↓septicemia, ↓mortality than congenic litter mates. ↑Th1 polarization in the spleen, given that: ↑TNFα and ↑IFNγ, ↓IL-5 and ↓IL-10 upon stimulation ex vivo with exotoxin toxic shock syndrome toxin-1 compared with congenic controls | [127] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drago-Serrano, M.E.; Campos-Rodríguez, R.; Carrero, J.C.; De la Garza, M. Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections. Int. J. Mol. Sci. 2017, 18, 501. https://doi.org/10.3390/ijms18030501
Drago-Serrano ME, Campos-Rodríguez R, Carrero JC, De la Garza M. Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections. International Journal of Molecular Sciences. 2017; 18(3):501. https://doi.org/10.3390/ijms18030501
Chicago/Turabian StyleDrago-Serrano, Maria Elisa, Rafael Campos-Rodríguez, Julio César Carrero, and Mireya De la Garza. 2017. "Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections" International Journal of Molecular Sciences 18, no. 3: 501. https://doi.org/10.3390/ijms18030501
APA StyleDrago-Serrano, M. E., Campos-Rodríguez, R., Carrero, J. C., & De la Garza, M. (2017). Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections. International Journal of Molecular Sciences, 18(3), 501. https://doi.org/10.3390/ijms18030501