Neurotrophic and Neuroregenerative Effects of GH/IGF1


Abstract
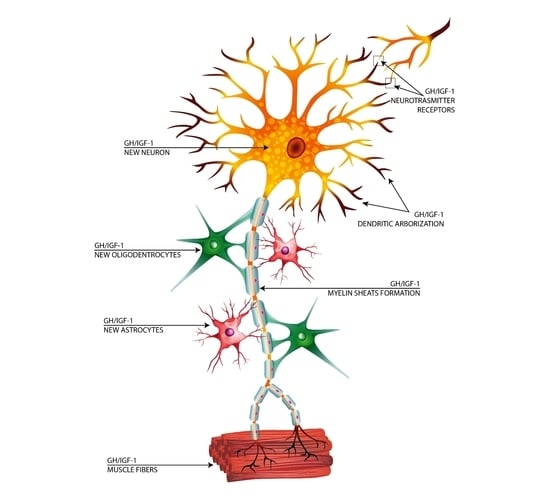
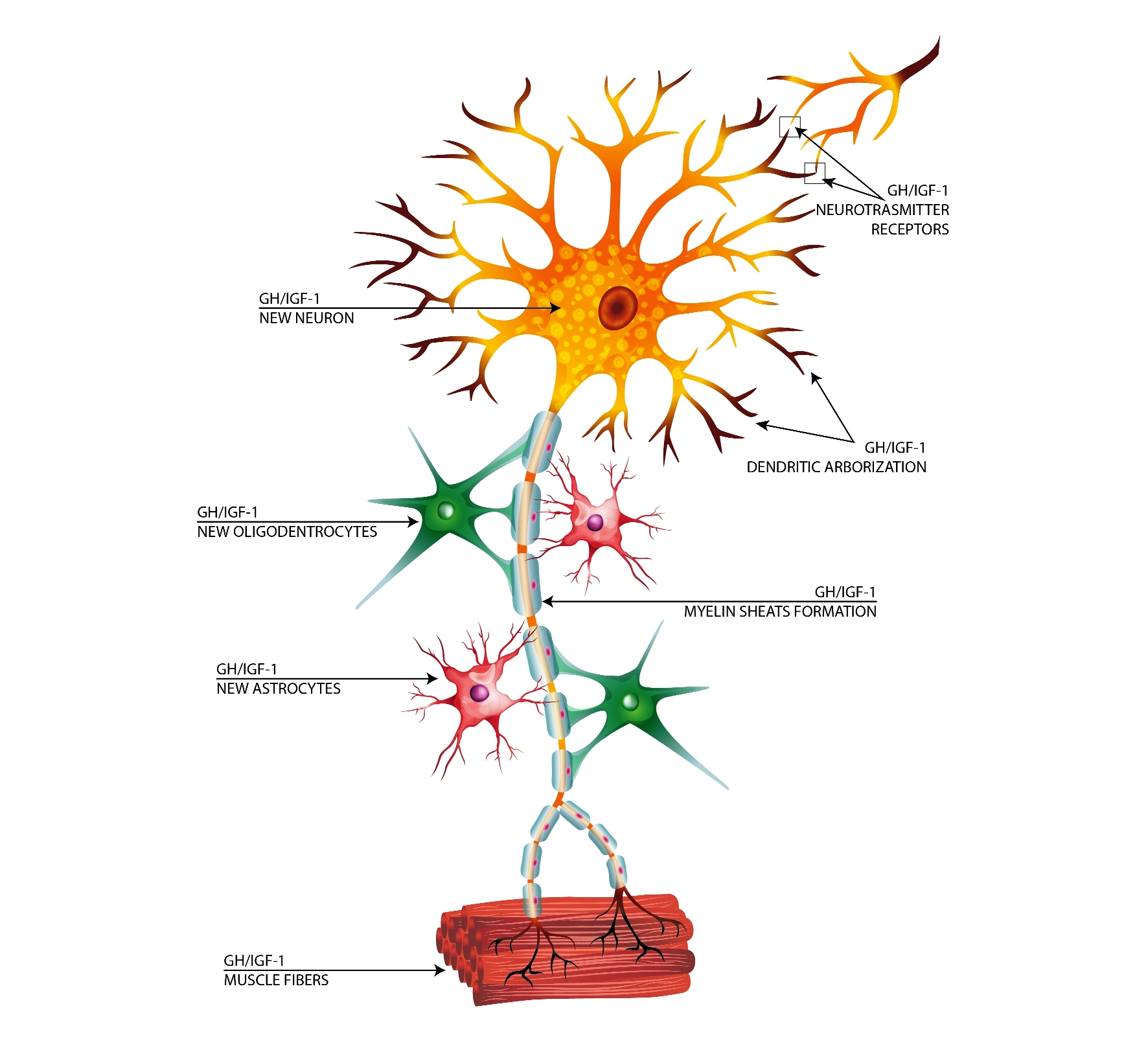
1. Introduction
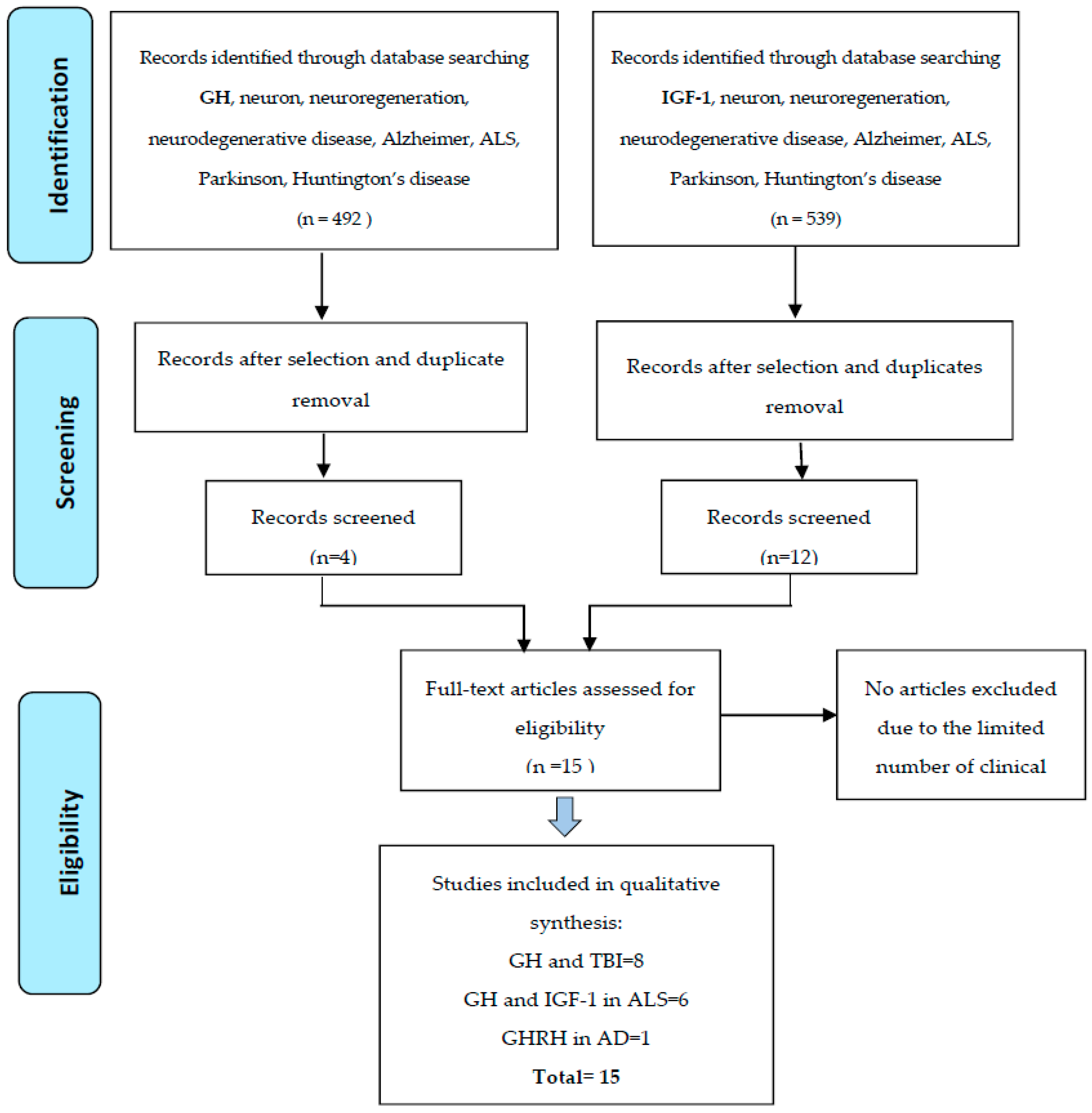
2. Methods
3. Results
3.1. Mechanisms of Action
3.2. GH Therapy in Traumatic Brain Injury (TBI)
3.3. GH/IGF-1 Treatment in Amyotrophic Lateral Sclerosis (ALS)
3.4. Peripheral Nerve Regeneration
3.5. GH/IGF-1 and Alzheimer’s Disease (AD)
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oh, J.; An, J.W.; Oh, S.I.; Oh, K.W.; Kim, J.A.; Lee, J.S.; Kim, S.H. Socioeconomic costs of amyotrophic lateral sclerosis according to staging system. Amyotroph. Lateral Scler. Frontotemporal Degener. 2015, 16, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Dawood, S. Caregiver burden, quality of life and vulnerability towards psychopathology in caregivers of patients with dementia/Alzheimer’s disease. J. Coll. Physicians Surg. Pak. 2016, 26, 892–895. [Google Scholar] [PubMed]
- Bohingamu Mudiyanselage, S.; Watts, J.J.; Abimanyi-Ochom, J.; Lane, L.; Murphy, A.T.; Morris, M.E.; Iansek, R. Cost of living with Parkinson’s disease over 12 months in Australia: A prospective cohort study. Parkinsons Dis. 2017, 2017, 5932675. [Google Scholar] [CrossRef] [PubMed]
- Aberg, N.D.; Brywe, K.G.; Isgaard, J. Aspects of growth hormone and insulin-like growth factor-i related to neuroprotection, regeneration, and functional plasticity in the adult brain. Sci. World J. 2006, 6, 53–80. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Yu, Y.; Cain, C.M.; Nyberg, F.; Couraud, P.O.; Kastin, A.J. Permeation of growth hormone across the blood-brain barrier. Endocrinology 2005, 146, 4898–4904. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.N.; Patel, P.; Gorski, M.; Gorevic, P.D.; Patlak, C.S.; Fenstermacher, J.D. In normal rat, intraventricularly administered insulin-like growth factor-1 is rapidly cleared from csf with limited distribution into brain. Cerebrospinal Fluid Res. 2005, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.S.; Wuarin, L.; Ishii, D.N. Uptake of circulating insulin-like growth factor-i into the cerebrospinal fluid of normal and diabetic rats and normalization of igf-ii mrna content in diabetic rat brain. J. Neurosci. Res. 2000, 59, 649–660. [Google Scholar] [CrossRef]
- Ashpole, N.M.; Sanders, J.E.; Hodges, E.L.; Yan, H.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging brain. Exp. Gerontol. 2015, 68, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Veldhuis, J.D. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocr. Rev. 1998, 19, 717–797. [Google Scholar] [CrossRef] [PubMed]
- Frago, L.M.; Paneda, C.; Dickson, S.L.; Hewson, A.K.; Argente, J.; Chowen, J.A. Growth hormone (gh) and gh-releasing peptide-6 increase brain insulin-like growth factor-i expression and activate intracellular signaling pathways involved in neuroprotection. Endocrinology 2002, 143, 4113–4122. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, M.; Leung, K.C.; Ross, R.J.; Iismaa, T.P.; Ho, K.K. Distribution and abundance of messenger ribonucleic acid for growth hormone receptor isoforms in human tissues. J. Clin. Endocrinol. Metab. 2000, 85, 2865–2871. [Google Scholar] [CrossRef] [PubMed]
- Holly, J.; Perks, C. The role of insulin-like growth factor binding proteins. Neuroendocrinology 2006, 83, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Bondy, C.; Werner, H.; Roberts, C.T., Jr.; LeRoith, D. Cellular pattern of type-i insulin-like growth factor receptor gene expression during maturation of the rat brain: Comparison with insulin-like growth factors i and ii. Neuroscience 1992, 46, 909–923. [Google Scholar] [CrossRef]
- Lobie, P.E.; Garcia-Aragon, J.; Lincoln, D.T.; Barnard, R.; Wilcox, J.N.; Waters, M.J. Localization and ontogeny of growth hormone receptor gene expression in the central nervous system. Brain Res. Dev. Brain Res. 1993, 74, 225–233. [Google Scholar] [CrossRef]
- Bando, H.; Zhang, C.; Takada, Y.; Yamasaki, R.; Saito, S. Impaired secretion of growth hormone-releasing hormone, growth hormone and igf-i in elderly men. Acta Endocrinol. 1991, 124, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.P.; Fernandez, A.M.; Haas, C.; Zimmer, E.; Portela, L.V.; Torres-Aleman, I. Reduced brain insulin-like growth factor i function during aging. Mol. Cell Neurosci. 2012, 49, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Rosenbloom, A.L.; Rivkees, S.A. Off-label use of recombinant igf-i to promote growth: Is it appropriate? J. Clin. Endocrinol. Metab. 2010, 95, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Ajo, R.; Cacicedo, L.; Navarro, C.; Sanchez-Franco, F. Growth hormone action on proliferation and differentiation of cerebral cortical cells from fetal rat. Endocrinology 2003, 144, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.B.; Bartke, A.; O’Callaghan, J.P. Increased glial fibrillary acidic protein (gfap) levels in the brains of transgenic mice expressing the bovine growth hormone (bgh) gene. Exp. Gerontol. 1995, 30, 383–400. [Google Scholar] [CrossRef]
- Oertel, H.; Schneider, H.J.; Stalla, G.K.; Holsboer, F.; Zihl, J. The effect of growth hormone substitution on cognitive performance in adult patients with hypopituitarism. Psychoneuroendocrinology 2004, 29, 839–850. [Google Scholar] [CrossRef]
- McLenachan, S.; Lum, M.G.; Waters, M.J.; Turnley, A.M. Growth hormone promotes proliferation of adult neurosphere cultures. Growth Horm. IGF Res. 2009, 19, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Morisawa, K.; Sugisaki, T.; Kanamatsu, T.; Aoki, T.; Noguchi, T. Factors contributing to cerebral hypomyelination in the growth hormone-deficient little mouse. Neurochem. Res. 1989, 14, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K. Functions of mature mammalian astrocytes: A current view. Neuroscientist 2010, 16, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Baquedano, E.; Chowen, J.A.; Argente, J.; Frago, L.M. Differential effects of gh and gh-releasing peptide-6 on astrocytes. J. Endocrinol. 2013, 218, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.A. Myelination and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tibullo, D.; Giallongo, C.; Raciti, G.; Viola, M.; Avola, R. Modulation of extracellular signal-related kinase, cyclin d1, glial fibrillary acidic protein, and vimentin expression in estradiol-pretreated astrocyte cultures treated with competence and progression growth factors. J. Neurosci. Res. 2015, 93, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tomassoni, D.; Traini, E.; Raciti, G.; Viola, M.; Li Volti, G.; Campisi, A.; Amenta, F.; Avola, R. Effect of growth factors and steroid hormones on heme oxygenase and cyclin d1 expression in primary astroglial cell cultures. J. Neurosci. Res. 2015, 93, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tibullo, D.; Giallongo, C.; Pappa, R.; Brundo, M.V.; Tomassoni, D.; Viola, M.; Amenta, F.; Avola, R. Neuroactive molecules and growth factors modulate cytoskeletal protein expression during astroglial cell proliferation and differentiation in culture. J. Neurosci. Res. 2016, 94, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Grasso, S.; Bramanti, V.; Tomassoni, D.; Bronzi, D.; Malfa, G.; Traini, E.; Napoli, M.; Renis, M.; Amenta, F.; Avola, R. Effect of lipoic acid and alpha-glyceryl-phosphoryl-choline on astroglial cell proliferation and differentiation in primary culture. J. Neurosci. Res. 2014, 92, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Lee, E.; Kim, J.W.; Kwon, B.S.; Jung, M.K.; Jee, Y.H.; Kim, J.; Bae, S.R.; Chang, Y.P. Protective effect of growth hormone on neuronal apoptosis after hypoxia-ischemia in the neonatal rat brain. Neurosci. Lett. 2004, 354, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Li, R.C.; Guo, S.Z.; Raccurt, M.; Moudilou, E.; Morel, G.; Brittian, K.R.; Gozal, D. Exogenous growth hormone attenuates cognitive deficits induced by intermittent hypoxia in rats. Neuroscience 2011, 196, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Pathipati, P.; Gorba, T.; Scheepens, A.; Goffin, V.; Sun, Y.; Fraser, M. Growth hormone and prolactin regulate human neural stem cell regenerative activity. Neuroscience 2011, 190, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Stern, W.C.; Miller, M.; Jalowiec, J.E.; Forbes, W.B.; Morgane, P.J. Effects of growth hormone on brain biogenic amine levels. Pharmacol. Biochem. Behav. 1975, 3, 1115–1118. [Google Scholar] [CrossRef]
- Andersson, K.; Fuxe, K.; Eneroth, P.; Isaksson, O.; Nyberg, F.; Roos, P. Rat growth hormone and hypothalamic catecholamine nerve terminal systems. Evidence for rapid and discrete reductions in dopamine and noradrenaline levels and turnover in the median eminence of the hypophysectomized male rat. Eur. J. Pharmacol. 1983, 95, 271–275. [Google Scholar] [CrossRef]
- Johansson, V.; Winberg, S.; Jonsson, E.; Hall, D.; Bjornsson, B.T. Peripherally administered growth hormone increases brain dopaminergic activity and swimming in rainbow trout. Horm. Behav. 2004, 46, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Ishii, D.N.; Glazner, G.W.; Pu, S.F. Role of insulin-like growth factors in peripheral nerve regeneration. Pharmacol. Ther. 1994, 62, 125–144. [Google Scholar] [CrossRef]
- Stewart, H.J.; Bradke, F.; Tabernero, A.; Morrell, D.; Jessen, K.R.; Mirsky, R. Regulation of rat schwann cell po expression and DNA synthesis by insulin-like growth factors in vitro. Eur. J. Neurosci. 1996, 8, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Caroni, P.; Grandes, P. Nerve sprouting in innervated adult skeletal muscle induced by exposure to elevated levels of insulin-like growth factors. J. Cell Biol. 1990, 110, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Glazner, G.W.; Ishii, D.N. Insulinlike growth factor gene expression in rat muscle during reinnervation. Muscle Nerve 1995, 18, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Dore, S.; Kar, S.; Quirion, R. Rediscovering an old friend, igf-i: Potential use in the treatment of neurodegenerative diseases. Trends Neurosci. 1997, 20, 326–331. [Google Scholar] [PubMed]
- Juanes, M.; Guercio, G.; Marino, R.; Berensztein, E.; Warman, D.M.; Ciaccio, M.; Gil, S.; Bailez, M.; Rivarola, M.A.; Belgorosky, A. Three novel igf1r mutations in microcephalic patients with prenatal and postnatal growth impairment. Clin. Endocrinol. (Oxf.) 2015, 82, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Gannage-Yared, M.H.; Klammt, J.; Chouery, E.; Corbani, S.; Megarbane, H.; Abou Ghoch, J.; Choucair, N.; Pfaffle, R.; Megarbane, A. Homozygous mutation of the igf1 receptor gene in a patient with severe pre- and postnatal growth failure and congenital malformations. Eur. J. Endocrinol. 2013, 168, K1–K7. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.P.; Urbonas, A.; Baddoo, A.; Baskin, S.; Malhotra, A.; Meggs, L.G. Igf-1 inhibits the mitochondrial apoptosis program in mesangial cells exposed to high glucose. Am. J. Physiol. Ren. Physiol. 2003, 285, F1013–F1024. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.L.; Randolph, A.; Yee, D.; Delafontaine, P.; Tennekoon, G.; Feldman, E.L. Characterization of insulin-like growth factor-i and its receptor and binding proteins in transected nerves and cultured schwann cells. J. Neurochem. 1996, 66, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Musaro, A.; McCullagh, K.J.; Naya, F.J.; Olson, E.N.; Rosenthal, N. Igf-1 induces skeletal myocyte hypertrophy through calcineurin in association with gata-2 and nf-atc1. Nature 1999, 400, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Musaro, A.; Giacinti, C.; Borsellino, G.; Dobrowolny, G.; Pelosi, L.; Cairns, L.; Ottolenghi, S.; Cossu, G.; Bernardi, G.; Battistini, L.; et al. Stem cell-mediated muscle regeneration is enhanced by local isoform of insulin-like growth factor 1. Proc. Natl. Acad. Sci. USA 2004, 101, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Shavlakadze, T.; White, J.D.; Davies, M.; Hoh, J.F.; Grounds, M.D. Insulin-like growth factor i slows the rate of denervation induced skeletal muscle atrophy. Neuromuscul. Disord. 2005, 15, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Insulin-like growth factors in the brain and their potential clinical implications. Neurology 2012, 79, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Morel, G.R.; Leon, M.L.; Uriarte, M.; Reggiani, P.C.; Goya, R.G. Therapeutic potential of IGF-1 on hippocampal neurogenesis and function during aging. Neurogenesis (Austin) 2017, 4, e1259709. [Google Scholar] [CrossRef] [PubMed]
- Mashayekhi, F.; Azari, M.; Moghadam, L.M.; Yazdankhah, M.; Naji, M.; Salehi, Z. Changes in cerebrospinal fluid nerve growth factor levels during chick embryonic development. J. Clin. Neurosci. 2009, 16, 1334–1337. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.M.; Mobley, B.C.; Hiller, A.; Feldman, E.L. Igf-i prevents glutamate-induced motor neuron programmed cell death. Neurobiol. Dis. 2004, 16, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Bronzi, D.; Bramanti, V.; Tomassoni, D.; Laureanti, F.; Grasso, S.; Li Volsi, G.; Avola, R. Neural markers expression in rat bone marrow mesenchymal stem cell cultures treated with neurosteroids. Neurochem. Res. 2010, 35, 2154–2160. [Google Scholar] [CrossRef] [PubMed]
- Feldman, E.L.; Sullivan, K.A.; Kim, B.; Russell, J.W. Insulin-like growth factors regulate neuronal differentiation and survival. Neurobiol. Dis. 1997, 4, 201–214. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dudek, H.; Datta, S.R.; Franke, T.F.; Birnbaum, M.J.; Yao, R.; Cooper, G.M.; Segal, R.A.; Kaplan, D.R.; Greenberg, M.E. Regulation of neuronal survival by the serine-threonine protein kinase akt. Science 1997, 275, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Bondy, C.A.; Cheng, C.M. Signaling by insulin-like growth factor 1 in brain. Eur. J. Pharmacol. 2004, 490, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.M.; Gordon, T.; Zochodne, D.W.; Power, H.A. Improving peripheral nerve regeneration: From molecular mechanisms to potential therapeutic targets. Exp. Neurol. 2014, 261, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.; Divall, S.; Wu, S. The regulation of reproductive neuroendocrine function by insulin and insulin-like growth factor-1 (igf-1). Front. Neuroendocrinol. 2014, 35, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Philpott, K.L.; Facci, L. Map kinase pathways in neuronal cell death. CNS Neurol. Disord. Drug Targets 2008, 7, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Silverman, S.M.; Liu, Y.; Wordinger, R.J.; Pang, I.H.; Clark, A.F. In vitro and in vivo neuroprotective effects of cjun n-terminal kinase inhibitors on retinal ganglion cells. Mol. Neurodegener. 2016, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Tell, V.; Hilgeroth, A. Recent developments of protein kinase inhibitors as potential ad therapeutics. Front. Cell Neurosci. 2013, 7, 189. [Google Scholar] [CrossRef] [PubMed]
- Tovar, Y.R.L.B.; Ramirez-Jarquin, U.N.; Lazo-Gomez, R.; Tapia, R. Trophic factors as modulators of motor neuron physiology and survival: Implications for als therapy. Front. Cell Neurosci. 2014, 8, 61. [Google Scholar]
- Fernandez, A.M.; Torres-Aleman, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Carro, E.; Trejo, J.L.; Gomez-Isla, T.; LeRoith, D.; Torres-Aleman, I. Serum insulin-like growth factor i regulates brain amyloid-beta levels. Nat. Med. 2002, 8, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Poirier, R.; Fernandez, A.M.; Torres-Aleman, I.; Metzger, F. Early brain amyloidosis in app/ps1 mice with serum insulin-like growth factor-i deficiency. Neurosci. Lett. 2012, 509, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, L.; Gouras, G.K.; Wang, R.; Gross, R.S.; Beal, M.F.; Greengard, P.; Xu, H. Stimulation of beta-amyloid precursor protein trafficking by insulin reduces intraneuronal beta-amyloid and requires mitogen-activated protein kinase signaling. J. Neurosci. 2001, 21, 2561–2570. [Google Scholar] [PubMed]
- Watson, G.S.; Peskind, E.R.; Asthana, S.; Purganan, K.; Wait, C.; Chapman, D.; Schwartz, M.W.; Plymate, S.; Craft, S. Insulin increases csf abeta42 levels in normal older adults. Neurology 2003, 60, 1899–1903. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Speder, P.; Brand, A.H. Control of brain development and homeostasis by local and systemic insulin signalling. Diabetes Obes. Metab. 2014, 16, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, S.W.; Brand, A.H. Cell biology. Insulin finds its niche. Science 2013, 340, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Garwood, C.J.; Ratcliffe, L.E.; Morgan, S.V.; Simpson, J.E.; Owens, H.; Vazquez-Villasenor, I.; Heath, P.R.; Romero, I.A.; Ince, P.G.; Wharton, S.B. Insulin and igf1 signalling pathways in human astrocytes in vitro and in vivo; characterisation, subcellular localisation and modulation of the receptors. Mol. Brain 2015, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Gontier, G.; George, C.; Chaker, Z.; Holzenberger, M.; Aid, S. Blocking igf signaling in adult neurons alleviates Alzheimer’s disease pathology through amyloid-beta clearance. J. Neurosci. 2015, 35, 11500–11513. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.K.; Chen, C.L.; Ting, C.H.; Lin-Chao, S.; Hwu, W.L.; Dodge, J.C.; Passini, M.A.; Cheng, S.H. Systemic administration of a recombinant aav1 vector encoding igf-1 improves disease manifestations in sma mice. Mol. Ther. 2014, 22, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Biondi, O.; Branchu, J.; Ben Salah, A.; Houdebine, L.; Bertin, L.; Chali, F.; Desseille, C.; Weill, L.; Sanchez, G.; Lancelin, C.; et al. IGF-1r reduction triggers neuroprotective signaling pathways in spinal muscular atrophy mice. J. Neurosci. 2015, 35, 12063–12079. [Google Scholar] [CrossRef] [PubMed]
- Amaducci, L.; Tesco, G. Aging as a major risk for degenerative diseases of the central nervous system. Curr. Opin. Neurol. 1994, 7, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Carro, E.; Trejo, J.L.; Gerber, A.; Loetscher, H.; Torrado, J.; Metzger, F.; Torres-Aleman, I. Therapeutic actions of insulin-like growth factor i on app/ps2 mice with severe brain amyloidosis. Neurobiol. Aging 2006, 27, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- White, M.F. Irs2 integrates insulin/igf1 signalling with metabolism, neurodegeneration and longevity. Diabetes Obes. Metab. 2014, 16, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Speder, P.; Liu, J.; Brand, A.H. Nutrient control of neural stem cells. Curr. Opin. Cell Biol. 2011, 23, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.F.; Gonzalo, I.T.; Cohan, P.; Berman, N.; Swerdloff, R.; Wang, C. Hypopituitarism following traumatic brain injury and aneurysmal subarachnoid hemorrhage: A preliminary report. J. Neurosurg. 2000, 93, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Behan, L.A.; Phillips, J.; Thompson, C.J.; Agha, A. Neuroendocrine disorders after traumatic brain injury. J. Neurol. Neurosurg. Psychiatry 2008, 79, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Bondanelli, M.; De Marinis, L.; Ambrosio, M.R.; Monesi, M.; Valle, D.; Zatelli, M.C.; Fusco, A.; Bianchi, A.; Farneti, M.; degli Uberti, E.C. Occurrence of pituitary dysfunction following traumatic brain injury. J. Neurotrauma 2004, 21, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.J.; Schneider, M.; Saller, B.; Petersenn, S.; Uhr, M.; Husemann, B.; von Rosen, F.; Stalla, G.K. Prevalence of anterior pituitary insufficiency 3 and 12 months after traumatic brain injury. Eur. J. Endocrinol. 2006, 154, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.J.; Mattsson, A.F.; Daousi, C.; Korbonits, M.; Koltowska-Haggstrom, M.; Cuthbertson, D.J. Gh deficiency after traumatic brain injury: Improvement in quality of life with gh therapy: Analysis of the kims database. Eur. J. Endocrinol. 2015, 172, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Moreau, O.K.; Cortet-Rudelli, C.; Yollin, E.; Merlen, E.; Daveluy, W.; Rousseaux, M. Growth hormone replacement therapy in patients with traumatic brain injury. J. Neurotrauma 2013, 30, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Sacca, F.; Quarantelli, M.; Rinaldi, C.; Tucci, T.; Piro, R.; Perrotta, G.; Carotenuto, B.; Marsili, A.; Palma, V.; De Michele, G.; et al. A randomized controlled clinical trial of growth hormone in amyotrophic lateral sclerosis: Clinical, neuroimaging, and hormonal results. J. Neurol. 2012, 259, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Reimunde, P.; Quintana, A.; Castanon, B.; Casteleiro, N.; Vilarnovo, Z.; Otero, A.; Devesa, A.; Otero-Cepeda, X.L.; Devesa, J. Effects of growth hormone (gh) replacement and cognitive rehabilitation in patients with cognitive disorders after traumatic brain injury. Brain Inj. 2011, 25, 65–73. [Google Scholar] [CrossRef] [PubMed]
- High, W.M., Jr.; Briones-Galang, M.; Clark, J.A.; Gilkison, C.; Mossberg, K.A.; Zgaljardic, D.J.; Masel, B.E.; Urban, R.J. Effect of growth hormone replacement therapy on cognition after traumatic brain injury. J. Neurotrauma 2010, 27, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Maric, N.P.; Doknic, M.; Pavlovic, D.; Pekic, S.; Stojanovic, M.; Jasovic-Gasic, M.; Popovic, V. Psychiatric and neuropsychological changes in growth hormone-deficient patients after traumatic brain injury in response to growth hormone therapy. J. Endocrinol. Investig. 2010, 33, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Kreitschmann-Andermahr, I.; Poll, E.M.; Reineke, A.; Gilsbach, J.M.; Brabant, G.; Buchfelder, M.; Fassbender, W.; Faust, M.; Kann, P.H.; Wallaschofski, H. Growth hormone deficient patients after traumatic brain injury—Baseline characteristics and benefits after growth hormone replacement—An analysis of the german kims database. Growth Horm. IGF Res. 2008, 18, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Hatton, J.; Kryscio, R.; Ryan, M.; Ott, L.; Young, B. Systemic metabolic effects of combined insulin-like growth factor-i and growth hormone therapy in patients who have sustained acute traumatic brain injury. J. Neurosurg. 2006, 105, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Devesa, J.; Reimunde, P.; Devesa, P.; Barbera, M.; Arce, V. Growth hormone (GH) and brain trauma. Horm. Behav. 2013, 63, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Abs, R.; Mattsson, A.F.; Bengtsson, B.A.; Feldt-Rasmussen, U.; Goth, M.I.; Koltowska-Haggstrom, M.; Monson, J.P.; Verhelst, J.; Wilton, P.; Group, K.S. Isolated growth hormone (gh) deficiency in adult patients: Baseline clinical characteristics and responses to gh replacement in comparison with hypopituitary patients. A sub-analysis of the kims database. Growth Horm. IGF Res. 2005, 15, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Devesa, P.; Reimunde, P.; Gallego, R.; Devesa, J.; Arce, V.M. Growth hormone (GH) treatment may cooperate with locally-produced GH in increasing the proliferative response of hippocampal progenitors to kainate-induced injury. Brain Inj. 2011, 25, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Molitch, M.E.; Clemmons, D.R.; Malozowski, S.; Merriam, G.R.; Vance, M.L.; Endocrine, S. Evaluation and treatment of adult growth hormone deficiency: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1587–1609. [Google Scholar] [CrossRef] [PubMed]
- Robberecht, W.; Philips, T. The changing scene of amyotrophic lateral sclerosis. Nat. Rev. Neurosci. 2013, 14, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, M.C.; Vucic, S.; Cheah, B.C.; Turner, M.R.; Eisen, A.; Hardiman, O.; Burrell, J.R.; Zoing, M.C. Amyotrophic lateral sclerosis. Lancet 2011, 377, 942–955. [Google Scholar] [CrossRef]
- Neumann, M.; Sampathu, D.M.; Kwong, L.K.; Truax, A.C.; Micsenyi, M.C.; Chou, T.T.; Bruce, J.; Schuck, T.; Grossman, M.; Clark, C.M.; et al. Ubiquitinated tdp-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 2006, 314, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Valentine, J.S.; Doucette, P.A.; Zittin Potter, S. Copper-zinc superoxide dismutase and amyotrophic lateral sclerosis. Annu. Rev. Biochem. 2005, 74, 563–593. [Google Scholar] [CrossRef] [PubMed]
- Al-Chalabi, A.; Fang, F.; Hanby, M.F.; Leigh, P.N.; Shaw, C.E.; Ye, W.; Rijsdijk, F. An estimate of amyotrophic lateral sclerosis heritability using twin data. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Rosen, D.R.; Siddique, T.; Patterson, D.; Figlewicz, D.A.; Sapp, P.; Hentati, A.; Donaldson, D.; Goto, J.; O’Regan, J.P.; Deng, H.X.; et al. Mutations in cu/zn superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.M.; Nilsson, P.; Forsgren, L.; Marklund, S.L. Cuzn-superoxide dismutase, extracellular superoxide dismutase, and glutathione peroxidase in blood from individuals homozygous for asp90ala cuzu-superoxide dismutase mutation. J. Neurochem. 1998, 70, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Steyn, F.J.; Ngo, S.T.; Lee, J.D.; Leong, J.W.; Buckley, A.J.; Veldhuis, J.D.; McCombe, P.A.; Chen, C.; Bellingham, M.C. Impairments to the gh-igf-i axis in hsod1g93a mice give insight into possible mechanisms of gh dysregulation in patients with amyotrophic lateral sclerosis. Endocrinology 2012, 153, 3735–3746. [Google Scholar] [CrossRef] [PubMed]
- Morselli, L.L.; Bongioanni, P.; Genovesi, M.; Licitra, R.; Rossi, B.; Murri, L.; Rossi, G.; Martino, E.; Gasperi, M. Growth hormone secretion is impaired in amyotrophic lateral sclerosis. Clin. Endocrinol. (Oxf.) 2006, 65, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Pellecchia, M.T.; Pivonello, R.; Monsurro, M.R.; Trojsi, F.; Longo, K.; Piccirillo, G.; Pivonello, C.; Rocco, M.; Di Somma, C.; Colao, A.; et al. The gh-igf system in amyotrophic lateral sclerosis: Correlations between pituitary gh secretion capacity, insulin-like growth factors and clinical features. Eur. J. Neurol. 2010, 17, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Spreux-Varoquaux, O.; Bensimon, G.; Lacomblez, L.; Salachas, F.; Pradat, P.F.; Le Forestier, N.; Marouan, A.; Dib, M.; Meininger, V. Glutamate levels in cerebrospinal fluid in amyotrophic lateral sclerosis: A reappraisal using a new hplc method with coulometric detection in a large cohort of patients. J. Neurol. Sci. 2002, 193, 73–78. [Google Scholar] [CrossRef]
- Lewis, S. Neurodegenerative disease: Expanding neurodegeneration modelling. Nat. Rev. Neurosci. 2015, 16, 376. [Google Scholar] [CrossRef] [PubMed]
- Saenger, S.; Holtmann, B.; Nilges, M.R.; Schroeder, S.; Hoeflich, A.; Kletzl, H.; Spooren, W.; Ostrowitzki, S.; Hanania, T.; Sendtner, M.; et al. Functional improvement in mouse models of familial amyotrophic lateral sclerosis by pegylated insulin-like growth factor i treatment depends on disease severity. Amyotroph. Lateral Scler. 2012, 13, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Kim, H.J.; Kim, M. The protective effect of growth hormone on Cu/Zn superoxide dismutase-mutant motor neurons. BMC Neurosci. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Torres-Aleman, I.; Barrios, V.; Berciano, J. The peripheral insulin-like growth factor system in amyotrophic lateral sclerosis and in multiple sclerosis. Neurology 1998, 50, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.; Melmed, S.; Sherman, B.; Frane, J.; Munsat, T.L.; Festoff, B.W. Recombinant growth hormone treatment of amyotrophic lateral sclerosis. Muscle Nerve 1993, 16, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, B.K.; Frost, L.M.; Christian, L.; Umapathi, P.; Gage, F.H. Synergy of insulin-like growth factor-1 and exercise in amyotrophic lateral sclerosis. Ann. Neurol. 2005, 57, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Lunetta, C.; Serafini, M.; Prelle, A.; Magni, P.; Dozio, E.; Ruscica, M.; Sassone, J.; Colciago, C.; Moggio, M.; Corbo, M.; et al. Impaired expression of insulin-like growth factor-1 system in skeletal muscle of amyotrophic lateral sclerosis patients. Muscle Nerve 2012, 45, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Sorenson, E.J.; Windbank, A.J.; Mandrekar, J.N.; Bamlet, W.R.; Appel, S.H.; Armon, C.; Barkhaus, P.E.; Bosch, P.; Boylan, K.; David, W.S.; et al. Subcutaneous igf-1 is not beneficial in 2-year als trial. Neurology 2008, 71, 1770–1775. [Google Scholar] [CrossRef] [PubMed]
- Nagano, I.; Shiote, M.; Murakami, T.; Kamada, H.; Hamakawa, Y.; Matsubara, E.; Yokoyama, M.; Moritaz, K.; Shoji, M.; Abe, K. Beneficial effects of intrathecal igf-1 administration in patients with amyotrophic lateral sclerosis. Neurol. Res. 2005, 27, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Borasio, G.D.; Robberecht, W.; Leigh, P.N.; Emile, J.; Guiloff, R.J.; Jerusalem, F.; Silani, V.; Vos, P.E.; Wokke, J.H.; Dobbins, T. A placebo-controlled trial of insulin-like growth factor-i in amyotrophic lateral sclerosis. European als/igf-i study group. Neurology 1998, 51, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C.; Felice, K.J.; Festoff, B.W.; Gawel, M.J.; Gelinas, D.F.; Kratz, R.; Murphy, M.F.; Natter, H.M.; Norris, F.H.; Rudnicki, S.A. Effect of recombinant human insulin-like growth factor-i on progression of als. A placebo-controlled study. The north america als/igf-i study group. Neurology 1997, 49, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Beauverd, M.; Mitchell, J.D.; Wokke, J.H.; Borasio, G.D. Recombinant human insulin-like growth factor i (rhIGF-1) for amyotrophic lateral sclerosis/motor neuron disease. Cochrane Database Syst. Rev. 2012, 11, CD002064. [Google Scholar] [CrossRef] [PubMed]
- Cudkowicz, M.E.; Shefner, J.M.; Schoenfeld, D.A.; Brown, R.H., Jr.; Johnson, H.; Qureshi, M.; Jacobs, M.; Rothstein, J.D.; Appel, S.H.; Pascuzzi, R.M.; et al. A randomized, placebo-controlled trial of topiramate in amyotrophic lateral sclerosis. Neurology 2003, 61, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Howe, C.L.; Bergstrom, R.A.; Horazdovsky, B.F. Subcutaneous igf-1 is not beneficial in 2-year als trial. Neurology 2009, 73, 1247. [Google Scholar] [CrossRef] [PubMed]
- Wilczak, N.; de Vos, R.A.; De Keyser, J. Free insulin-like growth factor (igf)-i and igf binding proteins 2, 5, and 6 in spinal motor neurons in amyotrophic lateral sclerosis. Lancet 2003, 361, 1007–1011. [Google Scholar] [CrossRef]
- Floyd, S.; Favre, C.; Lasorsa, F.M.; Leahy, M.; Trigiante, G.; Stroebel, P.; Marx, A.; Loughran, G.; O’Callaghan, K.; Marobbio, C.M.; et al. The insulin-like growth factor-i-mtor signaling pathway induces the mitochondrial pyrimidine nucleotide carrier to promote cell growth. Mol. Biol. Cell 2007, 18, 3545–3555. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, H.J.; Spencer, P.S. The fate of schwann cells isolated from axonal contact. J. Neurocytol. 1978, 7, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Aird, R.B.; Naffziger, H.C. The pathology of human striated muscle following denervation. J. Neurosurg. 1953, 10, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, O.A.; Gordon, T. Effects of short- and long-term schwann cell denervation on peripheral nerve regeneration, myelination, and size. Glia 2000, 32, 234–246. [Google Scholar] [CrossRef]
- Scheib, J.; Hoke, A. Advances in peripheral nerve regeneration. Nat. Rev. Neurol. 2013, 9, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Kanje, M.; Skottner, A.; Lundborg, G. Effects of growth hormone treatment on the regeneration of rat sciatic nerve. Brain Res. 1988, 475, 254–258. [Google Scholar] [CrossRef]
- Devesa, P.; Gelabert, M.; Gonzlez-Mosquera, T.; Gallego, R.; Relova, J.L.; Devesa, J.; Arce, V.M. Growth hormone treatment enhances the functional recovery of sciatic nerves after transection and repair. Muscle Nerve 2012, 45, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Nachemson, A.K.; Lundborg, G.; Hansson, H.A. Insulin-like growth factor i promotes nerve regeneration: An experimental study on rat sciatic nerve. Growth Factors 1990, 3, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Bayrak, A.F.; Olgun, Y.; Ozbakan, A.; Aktas, S.; Kulan, C.A.; Kamaci, G.; Demir, E.; Yilmaz, O.; Olgun, L. The effect of insulin like growth factor-1 on recovery of facial nerve crush injury. Clin. Exp. Otorhinolaryngol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.D.; Voskuhl, R.R. Neuroprotective effects of estrogens and androgens in cns inflammation and neurodegeneration. Front. Neuroendocrinol. 2012, 33, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.C.; Copley, P.A.; Harrell, J.R.; Wilhelm, J.C. Estrogen signaling is necessary for exercise-mediated enhancement of motoneuron participation in axon regeneration after peripheral nerve injury in mice. Dev. Neurobiol. 2017, 77, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Tehranipour, M.; Moghimi, A. Neuroprotective effects of testosterone on regenerating spinal cord motoneurons in rats. J. Mot. Behav. 2010, 42, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Tuffaha, S.H.; Budihardjo, J.D.; Sarhane, K.A.; Khusheim, M.; Song, D.; Broyles, J.M.; Salvatori, R.; Means, K.R., Jr.; Higgins, J.P.; Shores, J.T.; et al. Growth hormone therapy accelerates axonal regeneration, promotes motor reinnervation, and reduces muscle atrophy following peripheral nerve injury. Plast. Reconstr. Surg. 2016, 137, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Neff, N.T.; Prevette, D.; Houenou, L.J.; Lewis, M.E.; Glicksman, M.A.; Yin, Q.W.; Oppenheim, R.W. Insulin-like growth factors: Putative muscle-derived trophic agents that promote motoneuron survival. J. Neurobiol. 1993, 24, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. The genetics and molecular pathology of Alzheimer’s disease: Roles of amyloid and the presenilins. Neurol. Clin. 2000, 18, 903–922. [Google Scholar] [CrossRef]
- Solomon, A.; Mangialasche, F.; Richard, E.; Andrieu, S.; Bennett, D.A.; Breteler, M.; Fratiglioni, L.; Hooshmand, B.; Khachaturian, A.S.; Schneider, L.S.; et al. Advances in the prevention of Alzheimer’s disease and dementia. J. Intern. Med. 2014, 275, 229–250. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.S.; Mangialasche, F.; Andreasen, N.; Feldman, H.; Giacobini, E.; Jones, R.; Mantua, V.; Mecocci, P.; Pani, L.; Winblad, B.; et al. Clinical trials and late-stage drug development for Alzheimer’s disease: An appraisal from 1984 to 2014. J. Intern. Med. 2014, 275, 251–283. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Lloret, A.; Orti, R.; Alonso, D. Molecular bases of the treatment of Alzheimer’s disease with antioxidants: Prevention of oxidative stress. Mol. Asp. Med. 2004, 25, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Kryscio, R.J.; Abner, E.L.; Caban-Holt, A.; Lovell, M.; Goodman, P.; Darke, A.K.; Yee, M.; Crowley, J.; Schmitt, F.A. Association of antioxidant supplement use and dementia in the prevention of Alzheimer’s disease by vitamin e and selenium trial (preadvise). JAMA Neurol. 2017, 74, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.M.; Pike, C.J. Sex hormones, aging, and Alzheimer’s disease. Front. Biosci. (Elite Ed.) 2012, 4, 976–997. [Google Scholar] [PubMed]
- Rosario, E.R.; Chang, L.; Head, E.H.; Stanczyk, F.Z.; Pike, C.J. Brain levels of sex steroid hormones in men and women during normal aging and in Alzheimer’s disease. Neurobiol. Aging 2011, 32, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.J.; Carroll, J.C.; Rosario, E.R.; Barron, A.M. Protective actions of sex steroid hormones in Alzheimer’s disease. Front. Neuroendocrinol. 2009, 30, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Obermayr, R.P.; Mayerhofer, L.; Knechtelsdorfer, M.; Mersich, N.; Huber, E.R.; Geyer, G.; Tragl, K.H. The age-related down-regulation of the growth hormone/insulin-like growth factor-1 axis in the elderly male is reversed considerably by donepezil, a drug for Alzheimer’s disease. Exp. Gerontol. 2005, 40, 157–163. [Google Scholar] [CrossRef] [PubMed]
- McGinley, L.M.; Sims, E.; Lunn, J.S.; Kashlan, O.N.; Chen, K.S.; Bruno, E.S.; Pacut, C.M.; Hazel, T.; Johe, K.; Sakowski, S.A.; et al. Human cortical neural stem cells expressing insulin-like growth factor-i: A novel cellular therapy for Alzheimer’s disease. Stem Cells Transl. Med. 2016, 5, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T.; Rogers, J. Inflammation in Alzheimer disease-a brief review of the basic science and clinical literature. Cold Spring Harb. Perspect. Med. 2012, 2, a006346. [Google Scholar] [CrossRef] [PubMed]
- Avila-Munoz, E.; Arias, C. When astrocytes become harmful: Functional and inflammatory responses that contribute to Alzheimer’s disease. Ageing Res. Rev. 2014, 18, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Blasko, I.; Stampfer-Kountchev, M.; Robatscher, P.; Veerhuis, R.; Eikelenboom, P.; Grubeck-Loebenstein, B. How chronic inflammation can affect the brain and support the development of Alzheimer’s disease in old age: The role of microglia and astrocytes. Aging Cell 2004, 3, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Monson, N.L.; Ireland, S.J.; Ligocki, A.J.; Chen, D.; Rounds, W.H.; Li, M.; Huebinger, R.M.; Munro Cullum, C.; Greenberg, B.M.; Stowe, A.M.; et al. Elevated cns inflammation in patients with preclinical Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2014, 34, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Uchoa, M.F.; Moser, V.A.; Pike, C.J. Interactions between inflammation, sex steroids, and Alzheimer’s disease risk factors. Front. Neuroendocrinol. 2016, 43, 60–82. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.D.; Baker, L.D.; Borson, S.; Jensen, J.E.; Barsness, S.M.; Craft, S.; Merriam, G.R.; Otto, R.K.; Novotny, E.J.; Vitiello, M.V. Growth hormone-releasing hormone effects on brain gamma-aminobutyric acid levels in mild cognitive impairment and healthy aging. JAMA Neurol. 2013, 70, 883–890. [Google Scholar] [CrossRef] [PubMed]
- George, C.; Gontier, G.; Lacube, P.; Francois, J.C.; Holzenberger, M.; Aid, S. The Alzheimer’s disease transcriptome mimics the neuroprotective signature of igf-1 receptor-deficient neurons. Brain 2017, 140, 2012–2027. [Google Scholar] [CrossRef] [PubMed]
- Duan, C. Specifying the cellular responses to igf signals: Roles of igf-binding proteins. J. Endocrinol. 2002, 175, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.S.; Tsai, S.H.; Lee, T.C.; Lin, J.W.; Chang, C.K.; Chiu, W.T. Gene transfer of insulin-like growth factor-i providing neuroprotection after spinal cord injury in rats. J. Neurosurg. Spine 2007, 6, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.S.; Pacut, C.; Backus, C.; Hong, Y.; Johe, K.; Hefferan, M.; Marsala, M.; Feldman, E.L. The pleotrophic effects of insulin-like growth factor-i on human spinal cord neural progenitor cells. Stem Cells Dev. 2010, 19, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Arroba, A.I.; Wallace, D.; Mackey, A.; de la Rosa, E.J.; Cotter, T.G. Igf-i maintains calpastatin expression and attenuates apoptosis in several models of photoreceptor cell death. Eur. J. Neurosci. 2009, 30, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Scheepens, A.; Williams, C.E.; Breier, B.H.; Guan, J.; Gluckman, P.D. A role for the somatotropic axis in neural development, injury and disease. J. Pediatr. Endocrinol. Metab. 2000, 13, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Y.; Evans, M.S.; Hsieh, J.; Panici, J.; Bartke, A. Increased neurogenesis in dentate gyrus of long-lived ames dwarf mice. Endocrinology 2005, 146, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.A.; Smith, G.M.; Randall, D.C.; Brown, D.R.; Rabchevsky, A.G. Genetic manipulation of intraspinal plasticity after spinal cord injury alters the severity of autonomic dysreflexia. J. Neurosci. 2006, 26, 2923–2932. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Devesa, J.; Alonso, B.; Casteleiro, N.; Couto, P.; Castanon, B.; Zas, E.; Reimunde, P. Effects of recombinant growth hormone (gh) replacement and psychomotor and cognitive stimulation in the neurodevelopment of gh-deficient (ghd) children with cerebral palsy: A pilot study. Ther. Clin. Risk Manag. 2011, 7, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Aberg, D.; Jood, K.; Blomstrand, C.; Jern, C.; Nilsson, M.; Isgaard, J.; Aberg, N.D. Serum igf-i levels correlate to improvement of functional outcome after ischemic stroke. J. Clin. Endocrinol. Metab. 2011, 96, E1055–E1064. [Google Scholar] [CrossRef] [PubMed]
- De Smedt, A.; Brouns, R.; Uyttenboogaart, M.; De Raedt, S.; Moens, M.; Wilczak, N.; Luijckx, G.J.; De Keyser, J. Insulin-like growth factor i serum levels influence ischemic stroke outcome. Stroke 2011, 42, 2180–2185. [Google Scholar] [CrossRef] [PubMed]
- Denti, L.; Annoni, V.; Cattadori, E.; Salvagnini, M.A.; Visioli, S.; Merli, M.F.; Corradi, F.; Ceresini, G.; Valenti, G.; Hoffman, A.R.; et al. Insulin-like growth factor 1 as a predictor of ischemic stroke outcome in the elderly. Am. J. Med. 2004, 117, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Ebinger, M.; Ipsen, N.; Leonards, C.O.; Empl, L.; Hanne, L.; Liman, T.; Mai, K.; Strasburger, C.J.; Spranger, J.; Endres, M. Circulating insulin-like growth factor binding protein-3 predicts one-year outcome after ischemic stroke. Exp. Clin. Endocrinol. Diabetes 2015, 123, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.H.; Ma, L.L.; Yu, T.X.; Zheng, J.; Zhang, H.J.; Liang, H.; Shao, P. Insulin-like growth factor-1 as a prognostic marker in patients with acute ischemic stroke. PLoS ONE 2014, 9, e99186. [Google Scholar] [CrossRef] [PubMed]
- Cooney, R.N.; Shumate, M. The inhibitory effects of interleukin-1 on growth hormone action during catabolic illness. Vitam. Horm. 2006, 74, 317–340. [Google Scholar] [PubMed]
- Yumet, G.; Shumate, M.L.; Bryant, D.P.; Lang, C.H.; Cooney, R.N. Hepatic growth hormone resistance during sepsis is associated with increased suppressors of cytokine signaling expression and impaired growth hormone signaling. Crit. Care Med. 2006, 34, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Street, M.E.; Ziveri, M.A.; Spaggiari, C.; Viani, I.; Volta, C.; Grzincich, G.L.; Virdis, R.; Bernasconi, S. Inflammation is a modulator of the insulin-like growth factor (igf)/igf-binding protein system inducing reduced bioactivity of igfs in cystic fibrosis. Eur. J. Endocrinol. 2006, 154, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Farquharson, C. The effect of gh and igf1 on linear growth and skeletal development and their modulation by socs proteins. J. Endocrinol. 2010, 206, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Choukair, D.; Hugel, U.; Sander, A.; Uhlmann, L.; Tonshoff, B. Inhibition of igf-i-related intracellular signaling pathways by proinflammatory cytokines in growth plate chondrocytes. Pediatr. Res. 2014, 76, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in nf-kappab signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Voria, I.; Hauser, J.; Axis, A.; Schenker, M.; Bichet, S.; Kuntzer, T.; Grenningloh, G.; Barakat-Walter, I. Improved sciatic nerve regeneration by local thyroid hormone treatment in adult rat is accompanied by increased expression of scg10. Exp. Neurol. 2006, 197, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Mauras, N.; Rogol, A.D.; Haymond, M.W.; Veldhuis, J.D. Sex steroids, growth hormone, insulin-like growth factor-1: Neuroendocrine and metabolic regulation in puberty. Horm. Res. 1996, 45, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Shughrue, P.J.; Bushnell, C.D.; Dorsa, D.M. Estrogen receptor messenger ribonucleic acid in female rat brain during the estrous cycle: A comparison with ovariectomized females and intact males. Endocrinology 1992, 131, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.H.; McGinnis, M.Y. Androgen receptors in cranial nerve motor nuclei of male and female rats. J. Neurobiol. 2001, 46, 1–10. [Google Scholar] [CrossRef]
- Tabori, N.E.; Stewart, L.S.; Znamensky, V.; Romeo, R.D.; Alves, S.E.; McEwen, B.S.; Milner, T.A. Ultrastructural evidence that androgen receptors are located at extranuclear sites in the rat hippocampal formation. Neuroscience 2005, 130, 151–163. [Google Scholar] [CrossRef] [PubMed]
- DonCarlos, L.L.; Sarkey, S.; Lorenz, B.; Azcoitia, I.; Garcia-Ovejero, D.; Huppenbauer, C.; Garcia-Segura, L.M. Novel cellular phenotypes and subcellular sites for androgen action in the forebrain. Neuroscience 2006, 138, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Styne, D.M. The regulation of pubertal growth. Horm. Res. 2003, 60, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, U.J.; Ho, K.K. Modulation of growth hormone action by sex steroids. Clin. Endocrinol. (Oxf.) 2006, 65, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Bright, G.M.; Rogol, A.D.; Kappelgaard, A.M.; Rosenfeld, R.G.; American Norditropin Clinical Trials Group. Effects of dose and gender on the growth and growth factor response to gh in gh-deficient children: Implications for efficacy and safety. J. Clin. Endocrinol. Metab. 2002, 87, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Carswell, H.V.; Macrae, I.M.; Gallagher, L.; Harrop, E.; Horsburgh, K.J. Neuroprotection by a selective estrogen receptor beta agonist in a mouse model of global ischemia. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1501–H1504. [Google Scholar] [CrossRef] [PubMed]
- Hurn, P.D.; Macrae, I.M. Estrogen as a neuroprotectant in stroke. J. Cereb. Blood Flow Metab. 2000, 20, 631–652. [Google Scholar] [CrossRef] [PubMed]
- Toung, T.J.; Traystman, R.J.; Hurn, P.D. Estrogen-mediated neuroprotection after experimental stroke in male rats. Stroke 1998, 29, 1666–1670. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.; Ghoumari, A.M.; Bielecki, B.; Steibel, J.; Boehm, N.; Liere, P.; Macklin, W.B.; Kumar, N.; Habert, R.; Mhaouty-Kodja, S.; et al. The neural androgen receptor: A therapeutic target for myelin repair in chronic demyelination. Brain 2013, 136, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Tanzer, L.; Jones, K.J. Gonadal steroid regulation of hamster facial nerve regeneration: Effects of dihydrotestosterone and estradiol. Exp. Neurol. 1997, 146, 258–264. [Google Scholar] [CrossRef] [PubMed]
- McCombe, P.A.; Henderson, R.D. Effects of gender in amyotrophic lateral sclerosis. Gend. Med. 2010, 7, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Manjaly, Z.R.; Scott, K.M.; Abhinav, K.; Wijesekera, L.; Ganesalingam, J.; Goldstein, L.H.; Janssen, A.; Dougherty, A.; Willey, E.; Stanton, B.R.; et al. The sex ratio in amyotrophic lateral sclerosis: A population based study. Amyotroph. Lateral Scler. 2010, 11, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Lloret, A. Why women have more Alzheimer’s disease than men: Gender and mitochondrial toxicity of amyloid-beta peptide. J. Alzheimers Dis. 2010, 20, S527–S533. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Singh, M. Sex differences in cognitive impairment and Alzheimer’s disease. Front. Neuroendocrinol. 2014, 35, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Moisan, F.; Kab, S.; Mohamed, F.; Canonico, M.; Le Guern, M.; Quintin, C.; Carcaillon, L.; Nicolau, J.; Duport, N.; Singh-Manoux, A.; et al. Parkinson disease male-to-female ratios increase with age: French nationwide study and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2016, 87, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Kam, G.Y.; Leung, K.C.; Baxter, R.C.; Ho, K.K. Estrogens exert route- and dose-dependent effects on insulin-like growth factor (igf)-binding protein-3 and the acid-labile subunit of the igf ternary complex. J. Clin. Endocrinol. Metab. 2000, 85, 1918–1922. [Google Scholar] [PubMed]
- Cardona-Gomez, G.P.; Mendez, P.; DonCarlos, L.L.; Azcoitia, I.; Garcia-Segura, L.M. Interactions of estrogens and insulin-like growth factor-i in the brain: Implications for neuroprotection. Brain Res. Brain Res. Rev. 2001, 37, 320–334. [Google Scholar] [CrossRef]
- Drummond, E.S.; Harvey, A.R.; Martins, R.N. Androgens and Alzheimer’s disease. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Alsemari, A. Hypogonadism and neurological diseases. Neurol. Sci. 2013, 34, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Little, C.M.; Coons, K.D.; Sengelaub, D.R. Neuroprotective effects of testosterone on the morphology and function of somatic motoneurons following the death of neighboring motoneurons. J. Comp. Neurol. 2009, 512, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Atwi, S.; McMahon, D.; Scharfman, H.; MacLusky, N.J. Androgen modulation of hippocampal structure and function. Neuroscientist 2016, 22, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Galea, L.A.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, hormones and neurogenesis in the hippocampus: Hormonal modulation of neurogenesis and potential functional implications. J. Neuroendocrinol. 2013, 25, 1039–1061. [Google Scholar] [CrossRef] [PubMed]
- Fargo, K.N.; Galbiati, M.; Foecking, E.M.; Poletti, A.; Jones, K.J. Androgen regulation of axon growth and neurite extension in motoneurons. Horm. Behav. 2008, 53, 716–728. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, V.; Marbini, A. Neuroregenerative effect of oxandrolone: A case report. Am. J. Case Rep. 2015, 16, 763–767. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Son, S.W.; Lee, J.S.; Kim, H.G.; Kim, D.W.; Ahn, Y.C.; Son, C.G. Testosterone depletion increases the susceptibility of brain tissue to oxidative damage in a restraint stress mouse model. J. Neurochem. 2016, 136, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Kurth, F.; Luders, E.; Sicotte, N.L.; Gaser, C.; Giesser, B.S.; Swerdloff, R.S.; Montag, M.J.; Voskuhl, R.R.; Mackenzie-Graham, A. Neuroprotective effects of testosterone treatment in men with multiple sclerosis. Neuroimage Clin. 2014, 4, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.; Singh, M.; Su, C.; Cunningham, R.L. Effects of oxidative stress and testosterone on pro-inflammatory signaling in a female rat dopaminergic neuronal cell line. Endocrinology 2016, 157, 2824–2835. [Google Scholar] [CrossRef] [PubMed]
- Maggio, M.; Basaria, S.; Ceda, G.P.; Ble, A.; Ling, S.M.; Bandinelli, S.; Valenti, G.; Ferrucci, L. The relationship between testosterone and molecular markers of inflammation in older men. J. Endocrinol. Investig. 2005, 28, 116–119. [Google Scholar]
- Rettew, J.A.; Huet-Hudson, Y.M.; Marriott, I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Biol. Reprod. 2008, 78, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Cappon, J.; Brasel, J.A.; Mohan, S.; Cooper, D.M. Effect of brief exercise on circulating insulin-like growth factor i. J. Appl. Physiol. (1985) 1994, 76, 2490–2496. [Google Scholar]
- Schwarz, A.J.; Brasel, J.A.; Hintz, R.L.; Mohan, S.; Cooper, D.M. Acute effect of brief low- and high-intensity exercise on circulating insulin-like growth factor (igf) i, ii, and igf-binding protein-3 and its proteolysis in young healthy men. J. Clin. Endocrinol. Metab. 1996, 81, 3492–3497. [Google Scholar] [PubMed]
- Voss, M.W.; Vivar, C.; Kramer, A.F.; van Praag, H. Bridging animal and human models of exercise-induced brain plasticity. Trends Cogn. Sci. 2013, 17, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Trejo, J.L.; Carro, E.; Torres-Aleman, I. Circulating insulin-like growth factor i mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J. Neurosci. 2001, 21, 1628–1634. [Google Scholar] [PubMed]
- Eliakim, A.; Brasel, J.A.; Mohan, S.; Barstow, T.J.; Berman, N.; Cooper, D.M. Physical fitness, endurance training, and the growth hormone-insulin-like growth factor i system in adolescent females. J. Clin. Endocrinol. Metab. 1996, 81, 3986–3992. [Google Scholar] [PubMed]
- Nindl, B.C.; Santtila, M.; Vaara, J.; Hakkinen, K.; Kyrolainen, H. Circulating igf-i is associated with fitness and health outcomes in a population of 846 young healthy men. Growth Horm. IGF Res. 2011, 21, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Chadan, S.G.; Dill, R.P.; Vanderhoek, K.; Parkhouse, W.S. Influence of physical activity on plasma insulin-like growth factor-1 and insulin-like growth factor binding proteins in healthy older women. Mech. Ageing Dev. 1999, 109, 21–34. [Google Scholar] [CrossRef]
- Zhu, N.; Jacobs, D.R., Jr.; Schreiner, P.J.; Launer, L.J.; Whitmer, R.A.; Sidney, S.; Demerath, E.; Thomas, W.; Bouchard, C.; He, K.; et al. Cardiorespiratory fitness and brain volume and white matter integrity: The cardia study. Neurology 2015, 84, 2347–2353. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Yang, Y.R.; Wang, P.S.; Kuo, C.H.; Wang, R.Y. Insulin-like growth factor i signaling for brain recovery and exercise ability in brain ischemic rats. Med. Sci. Sports Exerc. 2011, 43, 2274–2280. [Google Scholar] [CrossRef] [PubMed]
- Mattlage, A.E.; Rippee, M.A.; Abraham, M.G.; Sandt, J.; Billinger, S.A. Estimated prestroke peak vo2 is related to circulating igf-1 levels during acute stroke. Neurorehabil. Neural Repair 2017, 31, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.R. Increased premorbid physical activity and amyotrophic lateral sclerosis: Born to run rather than run to death, or a seductive myth? J. Neurol. Neurosurg. Psychiatry 2013, 84, 947. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Nagaoka, U.; Nakayama, Y.; Kawata, A.; Kugimoto, C.; Kuroiwa, Y.; Kawai, M.; Shimohata, T.; Nishizawa, M.; Mihara, B.; et al. Reduction rate of body mass index predicts prognosis for survival in amyotrophic lateral sclerosis: A multicenter study in japan. Amyotroph. Lateral Scler. 2012, 13, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Wark, P.A.; Jenab, M.; Pearce, N.; Brayne, C.; Vermeulen, R.; Andersen, P.M.; Hallmans, G.; Kyrozis, A.; Vanacore, N.; et al. Prediagnostic body fat and risk of death from amyotrophic lateral sclerosis: The epic cohort. Neurology 2013, 80, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Calikoglu, A.; Karayal, A.; D’Ercole, A. Nutritional regulation of igf-i expression during brain development in mice. Pediatr. Res. 2001, 49, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.M.; Kelley, B.; Wang, J.; Strauss, D.; Eagles, D.A.; Bondy, C.A. A ketogenic diet increases brain insulin-like growth factor receptor and glucose transporter gene expression. Endocrinology 2003, 144, 2676–2682. [Google Scholar] [CrossRef] [PubMed]
- Hatton, J.; Rapp, R.P.; Kudsk, K.A.; Brown, R.O.; Luer, M.S.; Bukar, J.G.; Chen, S.A.; McClain, C.J.; Gesundheit, N.; Dempsey, R.J.; et al. Intravenous insulin-like growth factor-i (igf-i) in moderate-to-severe head injury: A phase ii safety and efficacy trial. J. Neurosurg. 1997, 86, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, A.; Lent-Schochet, D.; Pike, C.J. Diet-induced obesity and low testosterone increase neuroinflammation and impair neural function. J. Neuroinflamm. 2014, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Milman, S.; Atzmon, G.; Huffman, D.M.; Wan, J.; Crandall, J.P.; Cohen, P.; Barzilai, N. Low insulin-like growth factor-1 level predicts survival in humans with exceptional longevity. Aging Cell 2014, 13, 769–771. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Authors | Patients | Age | Type of Study | Duration | GH Dose (mg/day) | Clinical Outcome |
|---|---|---|---|---|---|---|
| Gardner, 2015 [82] | 161 | 42.6 | Clinical study | 1 year | 0.37 mg/day | GH therapy achieved clinically relevant, long-term benefit in quality of life |
| Moreau, 2013 [83] | 23 | 37.9 | Clinical trial | 1 year | 0.3 mg/day up to 0.6 mg/day | improve cognition and quality of life |
| Devesa, 2013 [90] | 13 | 6–53 | Follow-up | 8 months | 1 mg/day, 5 days/week, resting 15-days every 2-months | All patients improved during and at the end of treatment. Cognitive and motor improvements also swallowing. Visual performance ameliorated in amaurosis |
| Reimunde, 2011 [85] | 11 M (mean 44.5 months after injury) | 53.3 | Clinical trial | 3 months | 0.5 mg/day for 20 days, then 1 mg/day for 5 days/week | Significative improvement of cognitive parameter, total IQ, and WAIS scale |
| High, 2010 [86] | 23 | 39.1 | Randomized Controlled Trial | 1 year | 0.2 mg/day, increasing 0.2/month up to 0.6 mg/day | Significant improvements of the cognitive impairments that are partially reversible |
| Maric, 2010 [87] | 6 5 M 1 W | 38.6 | Follow-up | 6 months | 0.3 mg for males and 0.4 mg for female sc | Cognitive abilities, (particularly verbal and non-verbal memory) and psychiatric functioning were significantly improved |
| Kreitschmann-Andermahr, 2008 [88] | 854 (28 childhood) | 36.7 | Follow-Up | 1 year | 0.3 mg (starting dose) | Improvement of quality of life |
| Hatton, 2006 [89] | 49 GH/IGF-1 treated | 30 | Randomized double-blind study | 14 days | IGF-1/GH therapy IGF-1 continuous intravenous infusion (0.01 mg/kg/h), and GH (0.05 mg/kg/day) | IGF-I and GH produced sustained improvement in metabolic and nutritional endpoints |
| Authors | Patients | Age | Type of Study | Therapy | Doses of Therapy | Duration | Clinical Effects |
|---|---|---|---|---|---|---|---|
| Sorenson, 2008 [112] | 110 M 57 W (placebo 163) | 53.9 | Clinical Trial | IGF-1 | 0.05 mg/kg body weight twice daily | 2 years | Not provide benefit for patients with amyotrophic lateral sclerosis. |
| Nogano, 2005 [113] | 5 M 4 W | 46 49.5 | Randomized Controlled Trial | IGF-1 | Intrathecal administration high dose = 3 microg/kg body weight low dose = 0.5 microg/kg of body weight of IGF-1 every two weeks | 40 weeks | High-dose treatment slowed a decline of motor functions of the ALS patients in total Norris and limb Norris scales, but not in bulbar Norris or vital capacity. |
| Borasio, 1998 [114] | 183 | 51 | Randomized Controlled Trial | IGF-1 | 0.1 mg/kg/day | 9 months | Treatment showed no significant difference between groups and was safe and well tolerated. |
| Lai, 1997 [115] | 266 | 52.5 | Randomized Controlled Trial | IGF-1 | 0.05 mg/kg/day or 0.10 mg/kg/day | 9 months | Slowed the progression of functional impairment and the decline in health-related quality of life. High and low doses had similar effects. |
| Smith, 1993 [109] | 75 41 M + 34 W | 57.1 | Double-blind controlled trials | GH | 0.1 mg/day three time/week | 12–18 months | Survival analysis at 12 months did not reveal a difference between the treatment and placebo group. No change in IGF-1 plasma level. |
| Saccà [84] | 45 24 M + 16 F | 62.7 | randomized, placebo-controlled, double-blind | GH + riluzole | 0.6 mg (2 IU) s.c. every other day increased up to 2.4 mg (8 IU) | 12 month | No effect on cerebral NAA or clinical improvement Note that IGF-BP3 was reduced after therapy. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, V.E.; Locatelli, V.; Rizzi, L. Neurotrophic and Neuroregenerative Effects of GH/IGF1. Int. J. Mol. Sci. 2017, 18, 2441. https://doi.org/10.3390/ijms18112441
Bianchi VE, Locatelli V, Rizzi L. Neurotrophic and Neuroregenerative Effects of GH/IGF1. International Journal of Molecular Sciences. 2017; 18(11):2441. https://doi.org/10.3390/ijms18112441
Chicago/Turabian StyleBianchi, Vittorio Emanuele, Vittorio Locatelli, and Laura Rizzi. 2017. "Neurotrophic and Neuroregenerative Effects of GH/IGF1" International Journal of Molecular Sciences 18, no. 11: 2441. https://doi.org/10.3390/ijms18112441
APA StyleBianchi, V. E., Locatelli, V., & Rizzi, L. (2017). Neurotrophic and Neuroregenerative Effects of GH/IGF1. International Journal of Molecular Sciences, 18(11), 2441. https://doi.org/10.3390/ijms18112441