Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli

Abstract
1. Introduction
2. Results
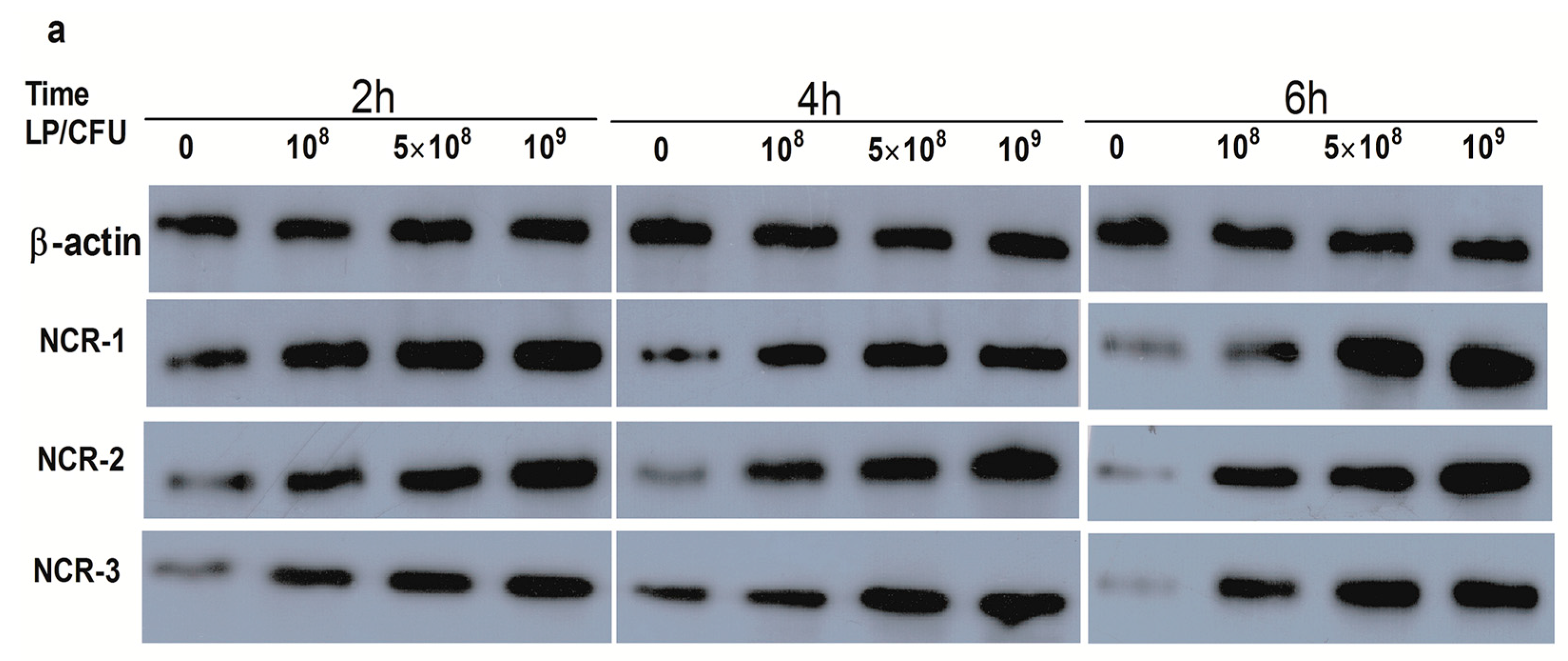
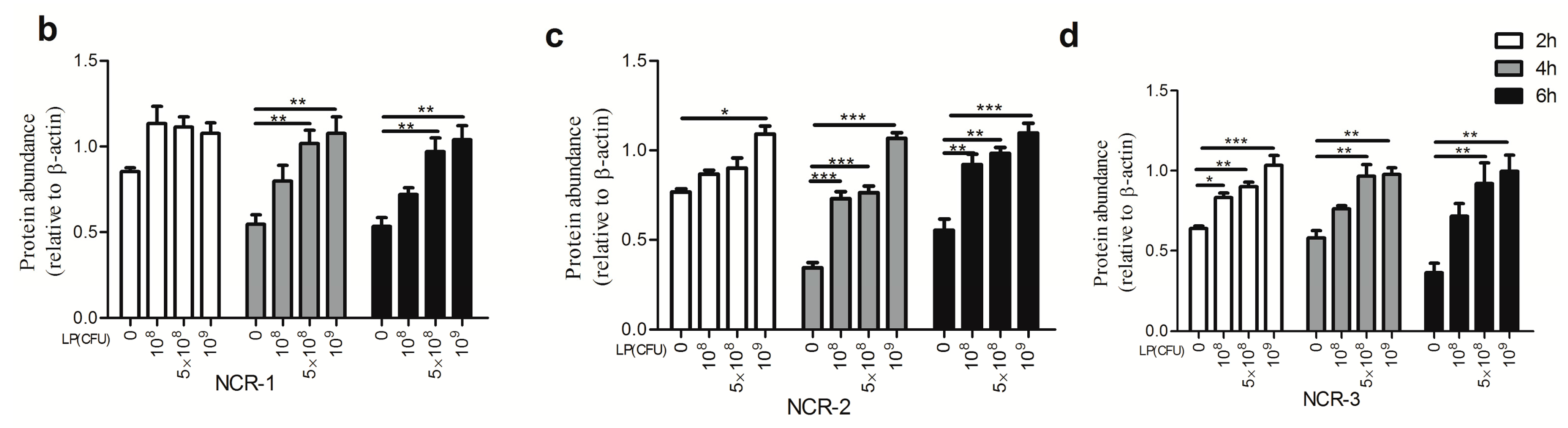
2.1. Effect of Lactobacillus plantarum on Natural Cytotoxicity Receptors (NCRs) Proteins Level in Natural Killer (NK) Cells
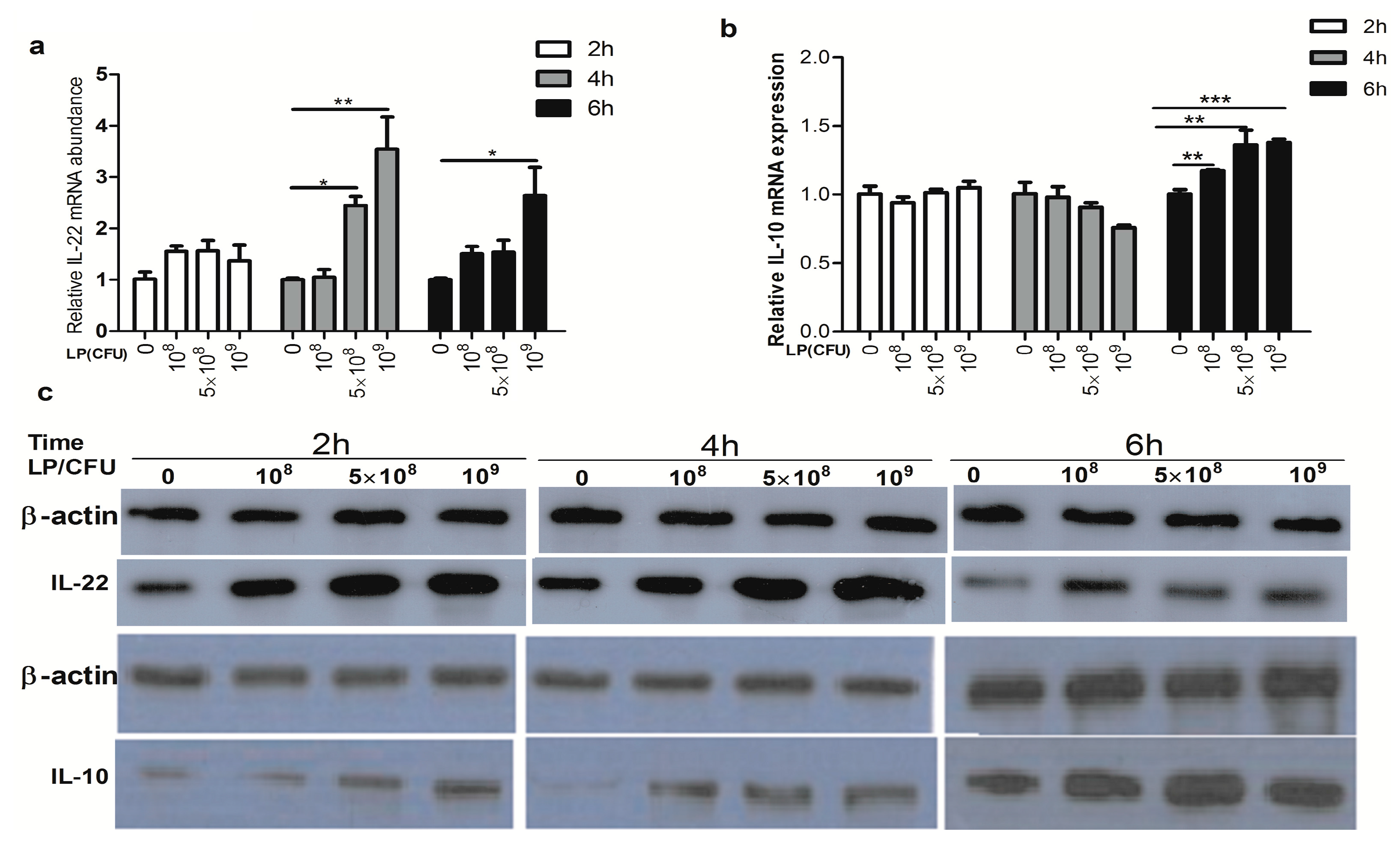
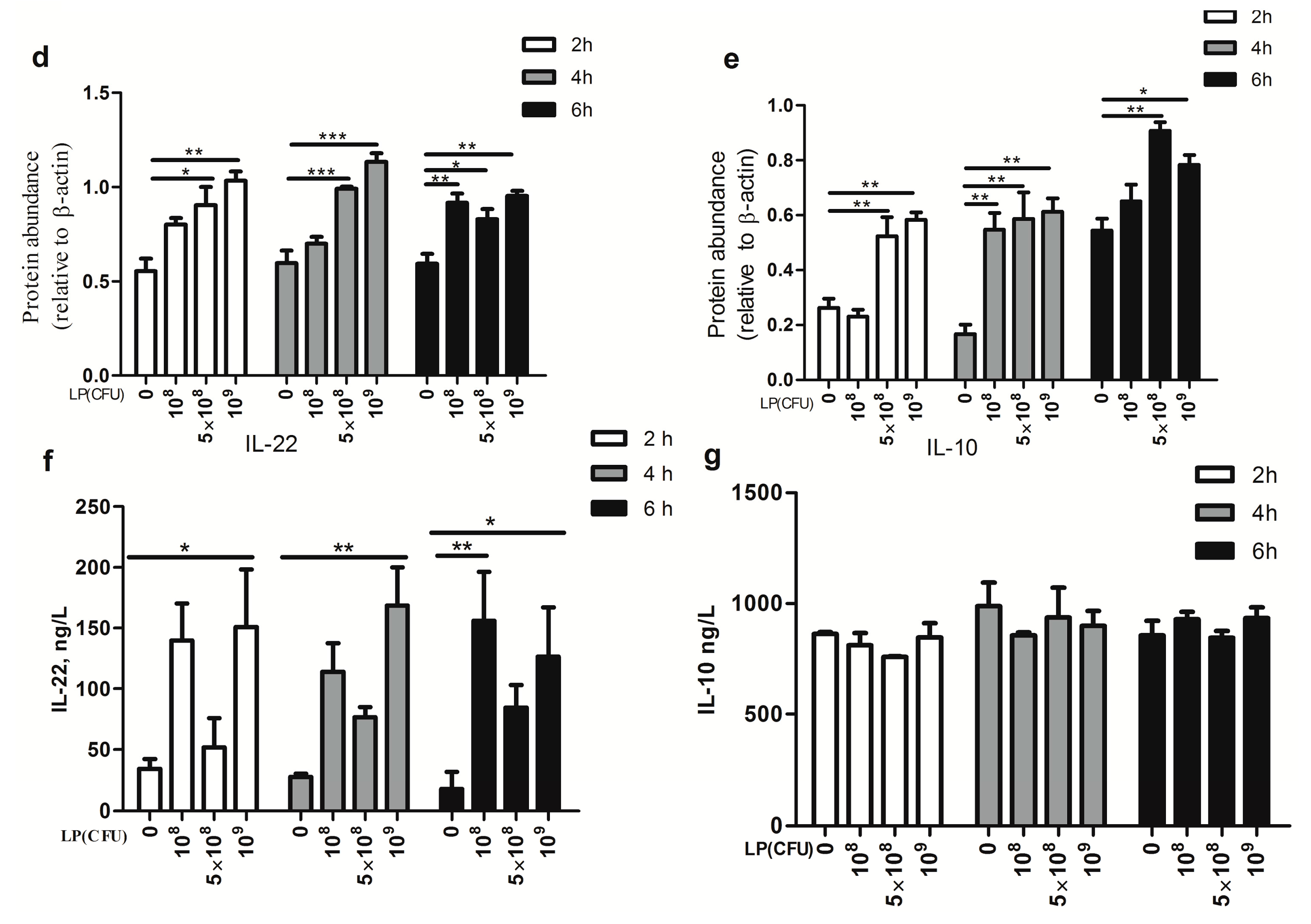
2.2. Effect of Lactobacillus plantarum on Expression of Cytokines by Natural Killer (NK) Cells
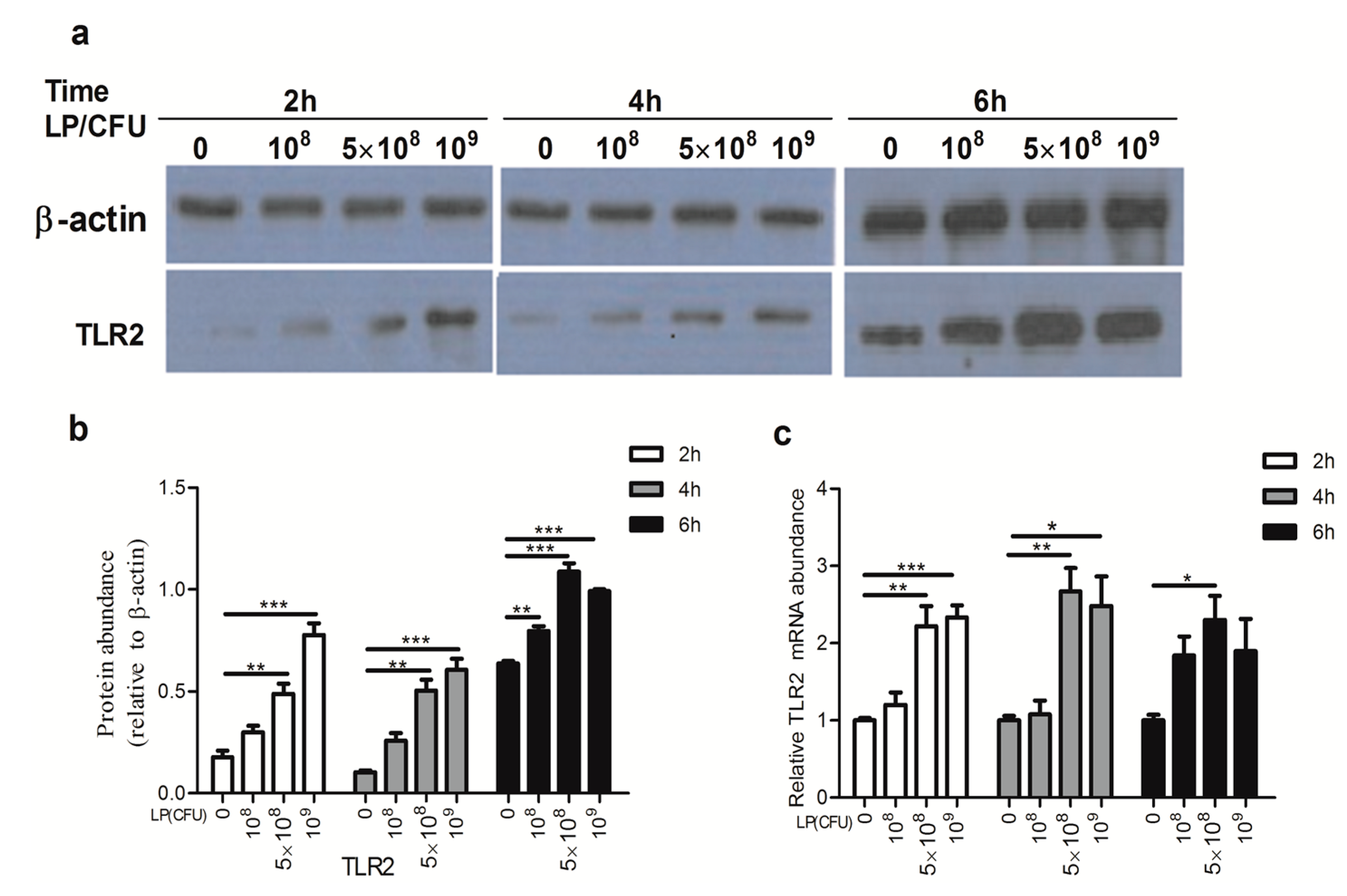
2.3. Lactobacillus plantarum Effectively Increased the Expression of Toll-Like Receptor 2 (TLR2)
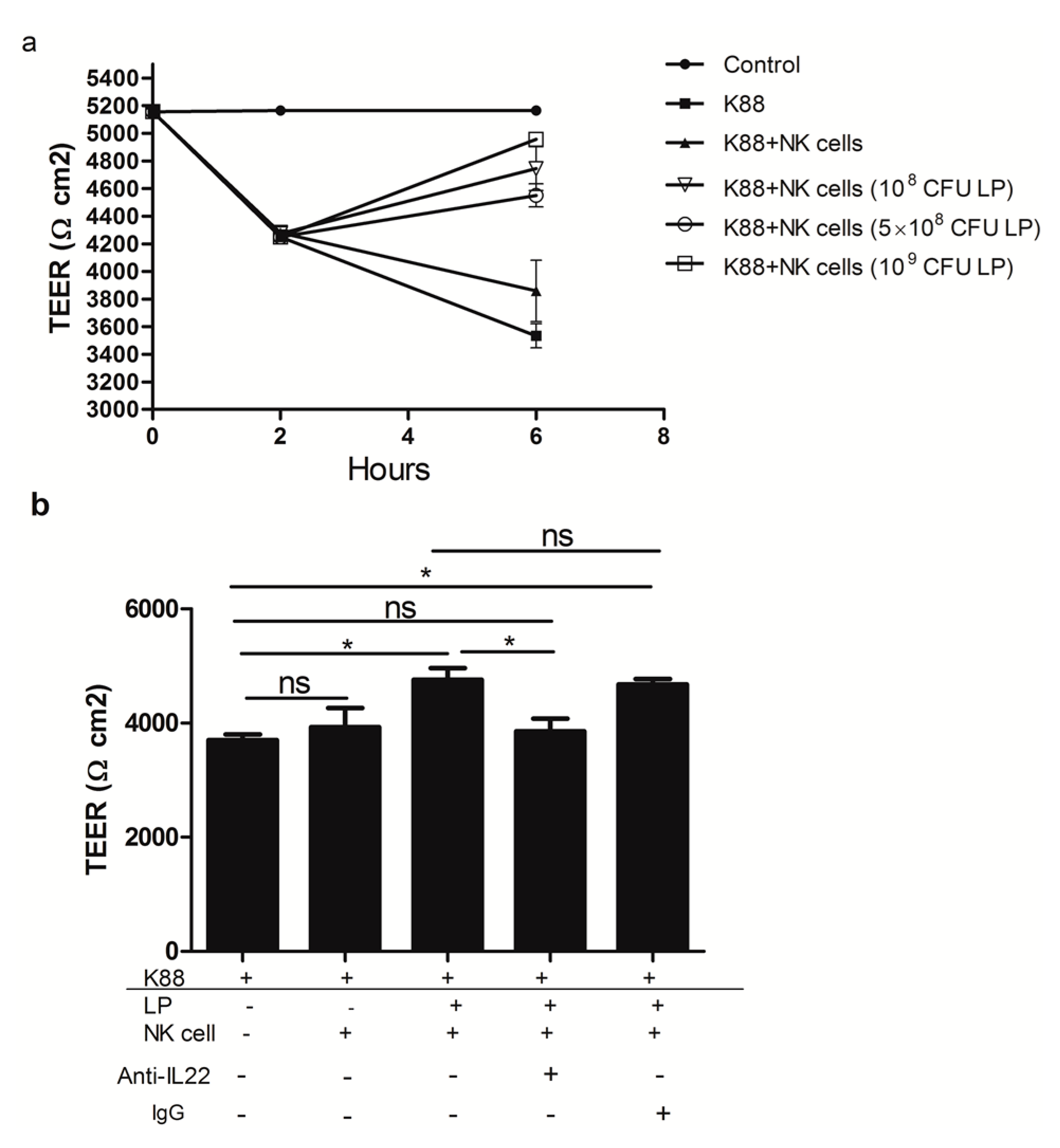
2.4. The Effect of NK Cells Stimulated by Lactobacillus plantarum on the TEER of NCM460 Cells Challenged with Enterotoxigenic Escherichia coli (ETEC) K88
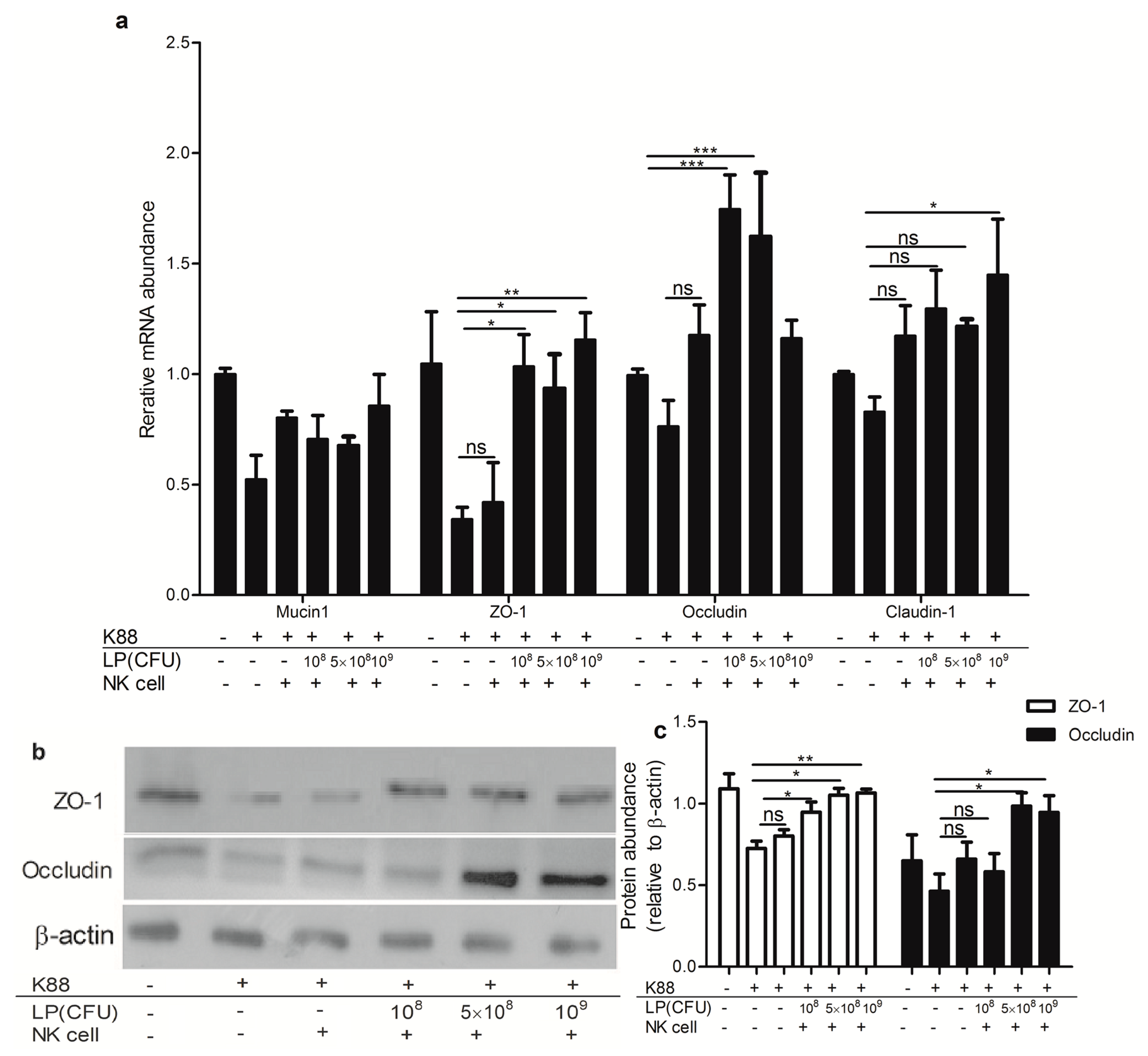
2.5. Lactobacillus plantarum-Treated NK Cells Increased Expression of Tight Junction Components in NCM460 Cells That Were Challenged with ETEC K88
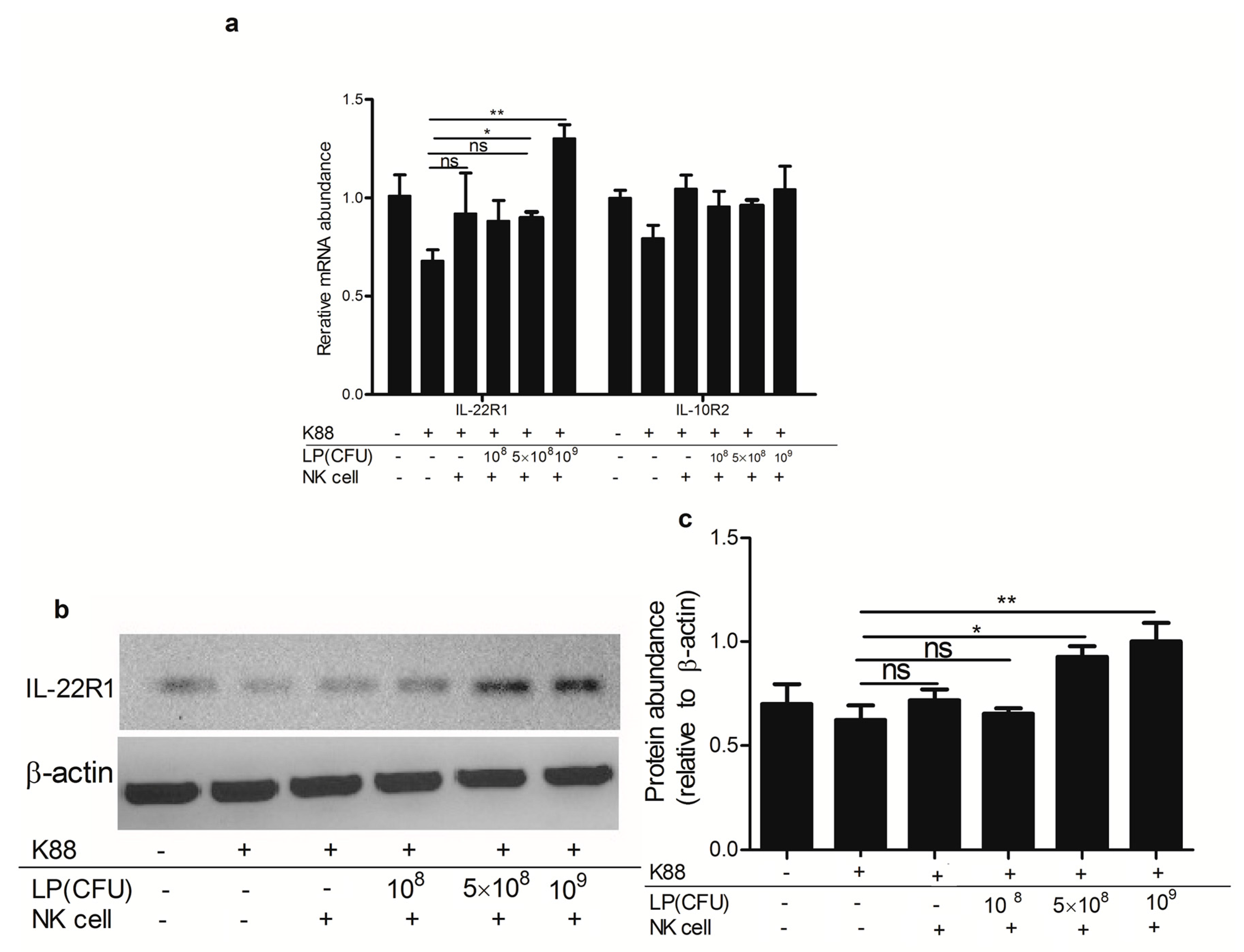
2.6. Lactobacillus plantarum-Treated NK Cells Up-Regulated the Expression of IL-22 Receptor
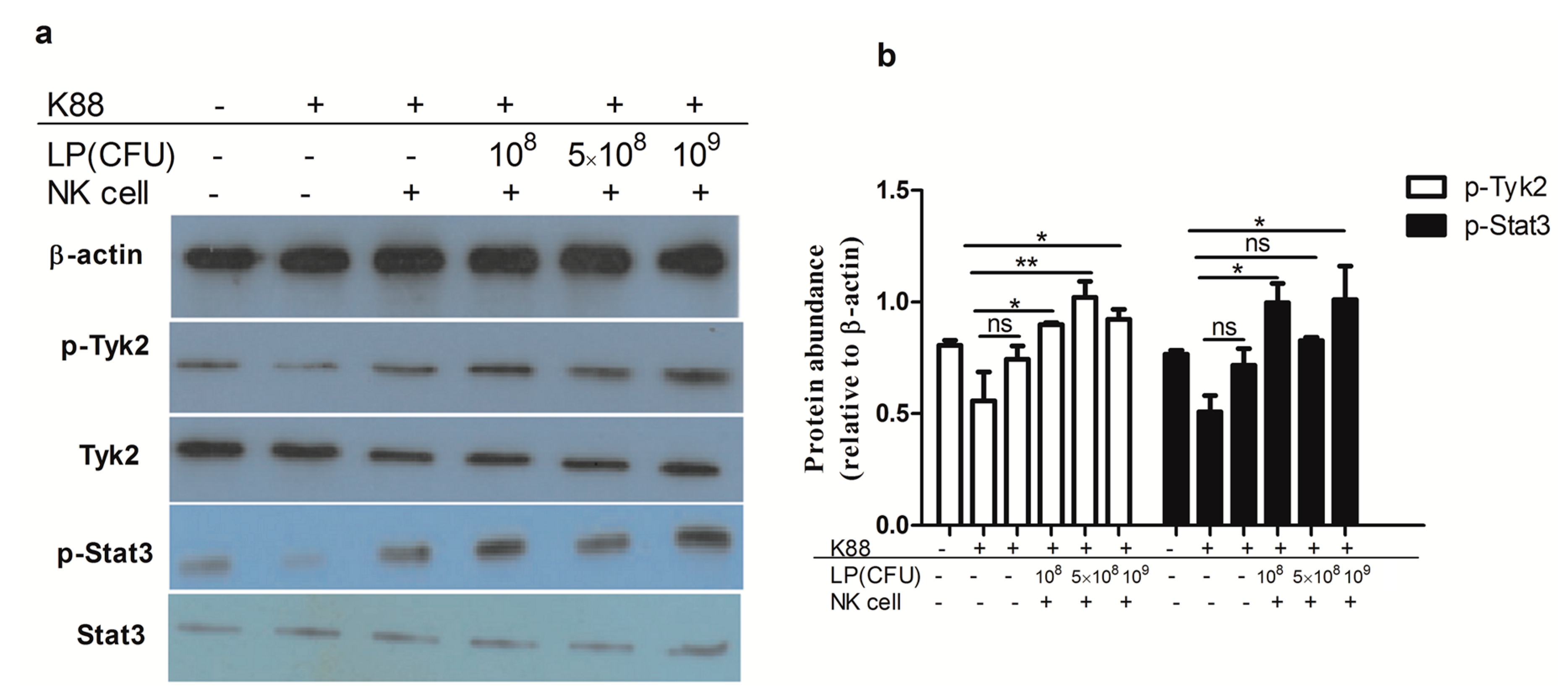
2.7. Lactobacillus plantarum-Treated NK Cells Activated Tyk2 and Stat3 in NCM460 Cells Challenged with ETEC K88
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Bacterial Growth
4.3. Treatment of NK Cells with L. plantarum
4.4. The Effect of NK Cells Stimulated by L. plantarum on the Function of ETEC K88-Infected NCM460 Cells
4.5. Measurement of Trans-Epithelial Electrical Resistance (TEER)
4.6. The Effect of IL-22 on Trans-Epithelial Electrical Resistance (TEER) of NCM460 Cell Monolayer Challenged with ETEC-K88
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Real-Time Quantitative PCR (QPCR) Analysis
4.9. Western Blotting
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.M.; Kaushik, R.S.; Hardwidge, P.R. Disruption of transepithelial resistance by enterotoxigenic Escherichia coli. Vet. Microbiol. 2010, 141, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ngendahayo, M.C.; Dubreuil, J.D. Escherichia coli heat-stable toxin b impairs intestinal epithelial barrier function by altering tight junction proteins. Infect. Immun. 2013, 81, 2819–2827. [Google Scholar] [CrossRef] [PubMed]
- Lodemann, U.; Strahlendorf, J.; Schierack, P.; Klingspor, S.; Aschenbach, J.R.; Martens, H. Effects of the probiotic enterococcus faecium and pathogenic Escherichia coli strains in a pig and human epithelial intestinal cell model. Scientifica 2015, 2015, 235184. [Google Scholar] [CrossRef]
- Yu, Q.H.; Yuan, L.X.; Deng, J.; Yang, Q. Lactobacillus protects the integrity of intestinal epithelial barrier damaged by pathogenic bacteria. Front. Cell. Infect. Microbiol. 2015, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zuo, Z.X.; Mao, A.P. Effect of probiotics on inducing remission and maintaining therapy in ulcerative colitis, Crohn’s disease, and pouchitis: Meta-analysis of randomized controlled trials. Inflamm. Bowel Dis. 2014, 20, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Saez-Lara, M.J.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: A systematic review of randomized human clinical trials. Biomed. Res. Int. 2015, 505878. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.M.; Jiang, Z.Y.; Zheng, C.T.; Wang, L.; Yang, X.F. Effect of Lactobacillus plantarumon diarrhea and intestinal barrier function of young piglets challenged with enterotoxigenic Escherichia coli K88. J. Anim. Sci. 2014, 92, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Shen, T.Y.; Zhang, P.; Ma, Y.L.; Moyer, M.P.; Qin, H.L. Protective effects of Lactobacillus plantarum against epithelial barrier dysfunction of human colon cell line NCM460. World J. Gastroenterol. 2010, 16, 5759–5765. [Google Scholar] [CrossRef] [PubMed]
- Bär, E.; Whitney, P.G.; Moor, K.; e Sousa, C.R.; LeibundGut-Landmann, S. IL-17 Regulates Systemic Fungal Immunity by Controlling the Functional Competence of NK Cells. Immunity 2014, 40, 117–127. [Google Scholar]
- Ogawa, T.; Asai, Y.; Tamai, R.; Makimura, Y.; Sakamoto, H.; Hashikawa, S.; Yasuda, K. Natural killer cell activities of synbiotic Lactobacillus casei ssp. casei in conjunction with dextran. Cell Host Microbe 2006, 143, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, A.; Yan, H.; Ohashi, S.; Gotoh, Y.; Sato, A.; Tsutsui, H.; Kaisho, T.; Toda, T.; Tsuji, N.M. Lactococcus lactis subsp. Cremoris FC triggers IFN-γ production from NK and T cells via IL-12 and IL-18. Clin. Exp. Immunol. 2012, 14, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Thakar, M.S.; Ouyang, W.; Malarkannan, S. IL-22 from conventional NK cells is epithelial regenerative and inflammation protective during influenza infection. Mucosal Immunol. 2013, 6, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Takayama, N.; Vosshenrich, C.A.; Lesjean-Pottier, S.; Sawa, S.; Lochner, M.; Rattis, F.; Mention, J.J.; Thiam, K.; Cerf-Bensussan, N.; Mandelboim, O.; et al. Microbial flora drives interleukin 22 production in intestinal NKp46+ cells that provide innate mucosal immune defense. Immunity 2008, 29, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Valdez, P.A.; Danilenko, D.M.; Hu, Y.; Sa, S.M.; Gong, Q.; Abbas, A.R.; Modrusan, Z.; Ghilardi, N.; de Sauvage, F.J.; et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat. Med. 2008, 14, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Maroof, A.; Beattie, L.; Zubairi, S.; Svensson, M.; Stager, S.; Kaye, P.M. Posttranscriptional regulation of IL-10 gene expression allows natural killer cells to express immunoregulatory function. Immunity 2008, 29, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Gazit, R.; Gruda, R.; Elboim, M.; Arnon, T.I.; Katz, G.; Achdout, H.; Hanna, J.; Qimron, U.; Landau, G.; Greenbaum, E.; et al. Lethal influenza infection in the absence of the natural killer cell receptor gene Ncr1. Nat. Immunol. 2006, 7, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Walzer, T.; Bléry, M.; Chaix, J.; Fuseri, N.; Chasson, L.; Robbins, S.H.; Jaeger, S.; André, P.; Gauthier, L.; Daniel, L.; et al. Identification, activation, and selective in vivo ablation of mouse NK cells via NKp46. Proc. Natl. Acad. Sci. USA 2007, 104, 3384–3389. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Sanos, S.L.; Bui, V.L.; Mortha, A.; Oberle, K.; Heners, C.; Johner, C.; Diefenbach, A. RORγt and commensal microflora are required for the differentiation of mucosal interleukin 22-producing NKp46+ cells. Nat. Immunol. 2009, 10, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Hoshino, K.; Kawai, T.; Sanjo, H.; Takada, H.; Ogawa, T.; Takeda, K.; Akira, S. Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity 1999, 11, 443–451. [Google Scholar] [CrossRef]
- Rizzello, V.; Bonaccorsi, I.; Dongarrà, M.L.; Fink, L.N.; Ferlazzo, G. Role of natural killer and dendritic cell crosstalk in immunomodulation by commensal bacteria probiotics. J. Biomed. Biotechnol. 2011, 473097. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrède, J.P.; Del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol: Decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharm. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Aujla, S.J.; Chan, Y.R.; Zheng, M.; Fei, M.; Askew, D.J.; Pociask, D.A.; Reinhart, T.A.; McAllister, F.; Edeal, J.; Gaus, K.; et al. IL-22 mediates mucosal host defense against gram-negative bacterial pneumonia. Nat. Med. 2008, 14, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Zenewicz, L.A.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.J.; Stevens, S.; Flavell, R.A. Innate and adaptive interleukin-22 protects mice from inflammatory bowel disease. Immunity 2008, 6, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.Y.; Zhang, B.; He, W.Q.; Zha, J.M.; Odenwald, M.A.; Singh, G.; Tamura, A.; Shen, L.; Sailer, A.; Yeruva, S.; et al. IL-22 upregulates epithelial claudin-2 to drive diarrhea and enteric pathogen clearance. Cell Host Microbe 2017, 21, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.H.; Aggarwal, S.; Ho, W.H.; Foster, J.; Zhang, Z.; Stinson, J.; Wood, W.I.; Goddard, A.D.; Gurney, A.L. Interleukin (IL)-22, a novel human cytokine that signals through the interferon receptor-related proteins CRF2-4 and IL-22R. J. Biol. Chem. 2000, 275, 31335–31339. [Google Scholar] [CrossRef] [PubMed]
- Dumoutier, L.; Van Roost, E.; Colau, D.; Renauld, J.C. Human interleukin-10-related T cell-derived inducible factor: Molecular cloning and functional characterization as an hepatocyte-stimulating factor. Proc. Natl. Acad. Sci. USA 2000, 97, 10144–10149. [Google Scholar] [CrossRef] [PubMed]
- Brand, S.; Beigel, F.; Olszak, T.; Zitzmann, K.; Eichhorst, S.T.; Otte, J.M.; Diepolder, H.; Marquardt, A.; Jagla, W.; Popp, A.; et al. IL-22 is increased in active Crohn’s disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.A.; Clare, S.; Goulding, D.; Arasteh, J.M.; Stares, M.D.; Browne, H.P.; Keane, J.A.; Page, A.J.; Kumasaka, N.; Kane, L.; et al. Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen. Cell Host Microbe 2014, 16, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, G.F.; Fouser, L.A.; Artis, D. Border patrol: Regulation of immunity, inflammation and tissue homeostasis at barrier surfaces by IL-22. Nat. Immunol. 2011, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, D.; Dumoutier, L.; Constantinescu, S.; Kruijer, W.; Schuringa, J.J.; Renauld, J.C. Interleukin-22 (IL-22) activates the JAK/STAT, ERK, JNK, and p38 MAP kinase pathways in a rat hepatoma cell line. Pathways that are shared with and distinct from IL-10. J. Biol. Chem. 2002, 277, 33676–33682. [Google Scholar] [CrossRef] [PubMed]
- Pickert, G.; Neufert, C.; Leppkes, M.; Zheng, Y.; Wittkopf, N.; Warntjen, M.; Lehr, H.A.; Hirth, S.; Weigmann, B.; Wirtz, S.; et al. STAT3 links IL-22 signaling in intestinal epithelial cells to mucosal wound healing. J. Exp. Med. 2009, 206, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Moss, A.C.; Anton, P.; Savidge, T.; Newman, P.; Cheifetz, A.S.; Gay, J.; Paraschos, S.; Winter, M.W.; Moyer, M.P.; Karalis, K.; et al. Urocortin II mediates pro-inflammatory effects in human colonocytes via corticotropin-releasing hormone receptor 2α. Gut 2007, 56, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Chen, H.Q.; Zhang, M.; Jiang, Y.Q.; Hang, X.M.; Qin, H.L. Effect of Lactobacillus plantarum LP-Onlly on gut flora and colitis in interleukin-10knockout mice. J. Gastroenterol. Hepatol. 2011, 26, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.Q.; Ma, X.Y.; Yang, X.F.; Wang, L.; Jiang, Z.Y. Effect of sodium butyrate on cell proliferation and cell cycle in porcine intestinal epithelial (IPEC-J2) cells. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 304–311. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′-3′) | Product Size (bp) | % Primer Efficiency (E) | GenBank Accession | |
|---|---|---|---|---|---|
| IL-10 | Forward | AGAGGTCTCCAAAATCGG | 160 | 95.4 | NM_000572.2 |
| Reverse | GGCTTCTTTCTAAATCGTTC | ||||
| IL-22 | Forward | AGTGCTGTTCCCTCAATCT | 203 | 94.8 | NM_020525.4 |
| Reverse | GCAAATCCAGTTCTCCAAT | ||||
| TLR2 | Forward | CTCTACCAGATGCCTCCCT | 128 | 102.2 | NM_001318787.1 |
| Reverse | ATTGCCACCAGCTTCCA | ||||
| IL-10R2 | Forward | AGGGCTGAATTTGCAGATGA | 213 | 98.8 | NM_000628.4 |
| Reverse | CCGTTTTTCCAGTATTGCAC | ||||
| IL-22R1 | Forward | CCCCACTGGGACACTTTCTA | 243 | 96.9 | NM_021258.3 |
| Reverse | TGGCCCTTTAGGTACTGTGG | ||||
| ZO-1 | Forward | CAACATACAGTGACGCTTCACA | 105 | 105.5 | NM_001301025.2 |
| Reverse | CACTATTGACGTTTCCCCACTC | ||||
| Occludin | Forward | ACAAGCGGTTTTATCCAGAGTC | 89 | 98.3 | NM_001205254.1 |
| Reverse | GTCATCCACAGGCGAAGTTAAT | ||||
| Claudin1 | Forward | AAATCAGAACTTTGGAGGC | 103 | 97.9 | NM_021101.4 |
| Reverse | AAACAAGAGTGCTATGGGTC | ||||
| Mucin1 | Forward | CGCCTGCCTGAATCTGTT | 281 | 104.7 | NM_001018016.2 |
| Reverse | GCTCTTGGTAGTAGTCGGTGC | ||||
| β-actin | Forward | TAGTTGCGTTACACCCTTTC | 153 | 99.9 | NM_001101.3 |
| Reverse | TGTCACCTTCACCGTTCC | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Jiang, Z.; Hu, S.; Wang, L.; Ma, X.; Yang, X. Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli. Int. J. Mol. Sci. 2017, 18, 2409. https://doi.org/10.3390/ijms18112409
Qiu Y, Jiang Z, Hu S, Wang L, Ma X, Yang X. Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli. International Journal of Molecular Sciences. 2017; 18(11):2409. https://doi.org/10.3390/ijms18112409
Chicago/Turabian StyleQiu, Yueqin, Zongyong Jiang, Shenglan Hu, Li Wang, Xianyong Ma, and Xuefen Yang. 2017. "Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli" International Journal of Molecular Sciences 18, no. 11: 2409. https://doi.org/10.3390/ijms18112409
APA StyleQiu, Y., Jiang, Z., Hu, S., Wang, L., Ma, X., & Yang, X. (2017). Lactobacillus plantarum Enhanced IL-22 Production in Natural Killer (NK) Cells That Protect the Integrity of Intestinal Epithelial Cell Barrier Damaged by Enterotoxigenic Escherichia coli. International Journal of Molecular Sciences, 18(11), 2409. https://doi.org/10.3390/ijms18112409