
Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Clinical Findings
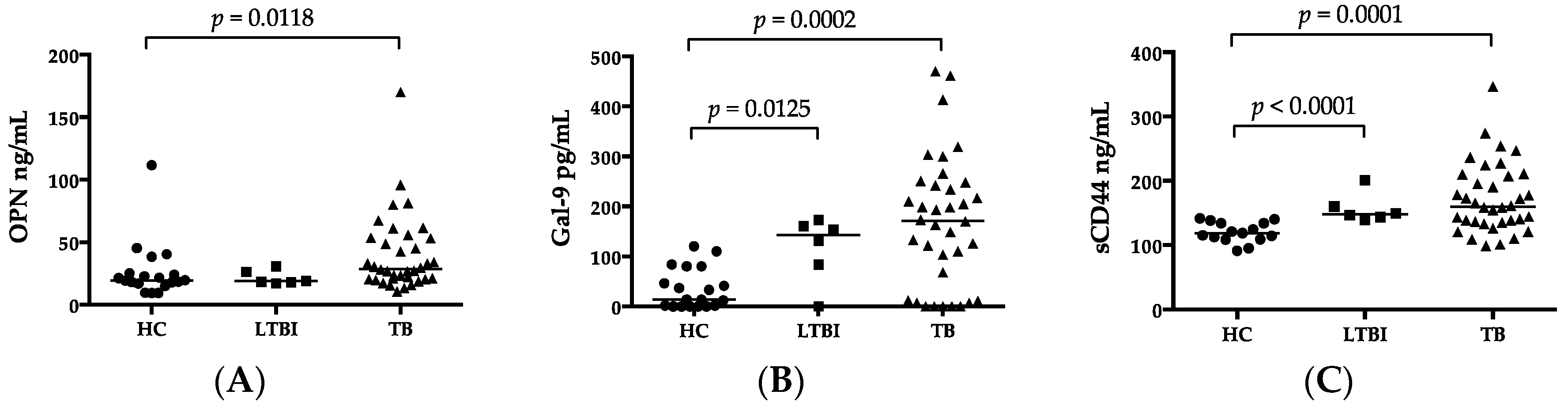
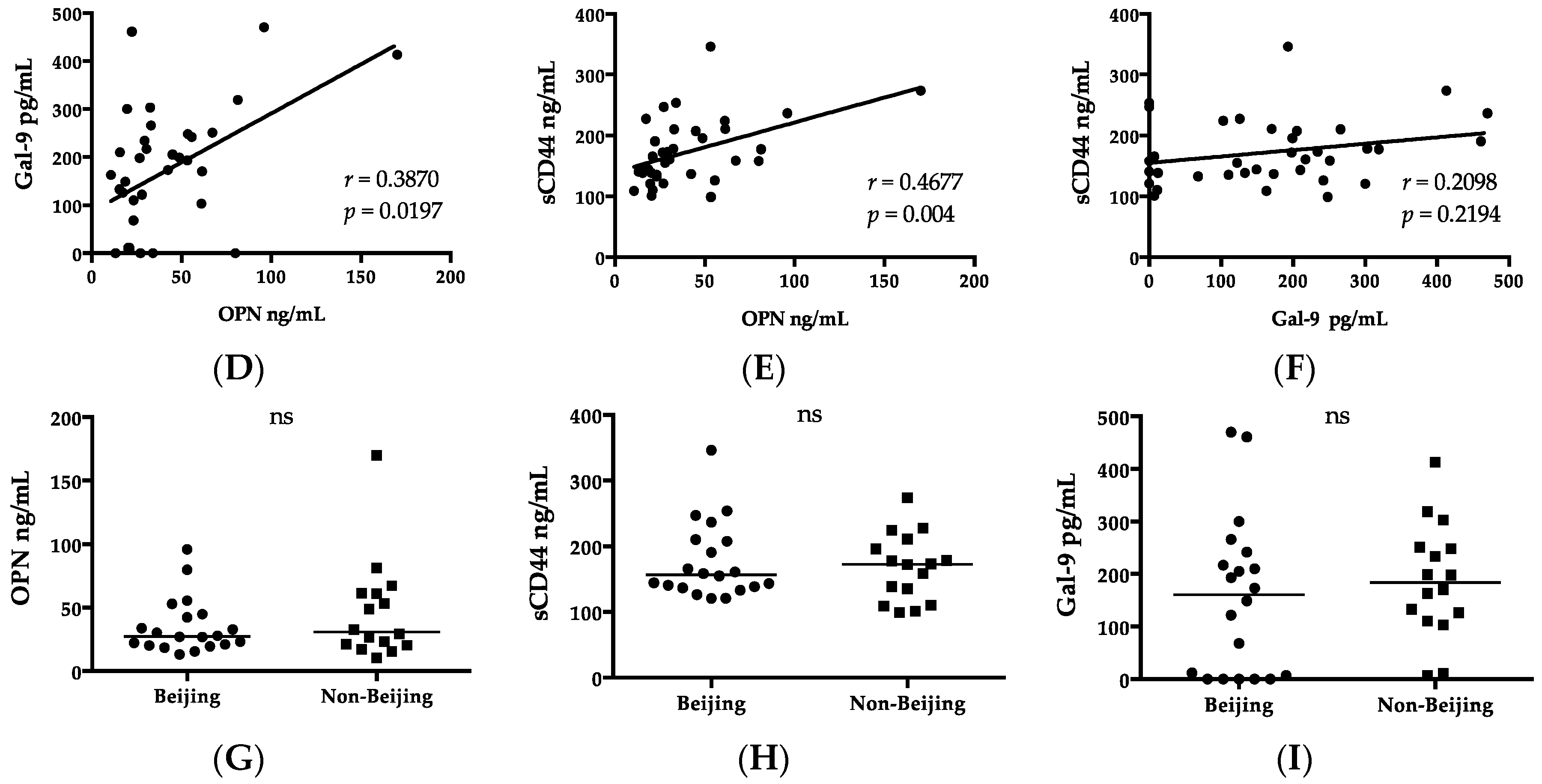
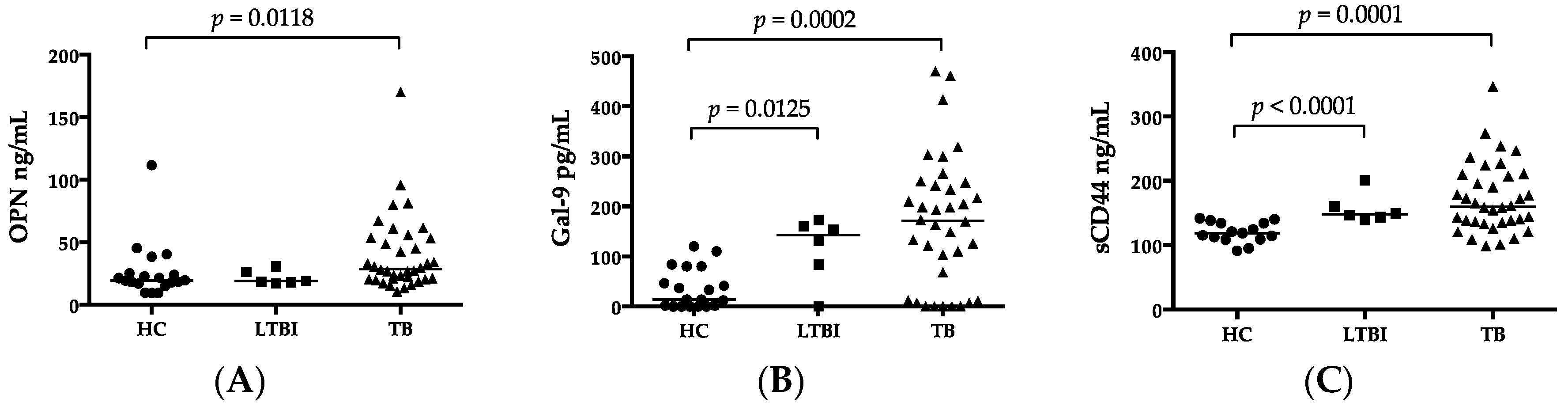
2.2. Luminex and ELISA
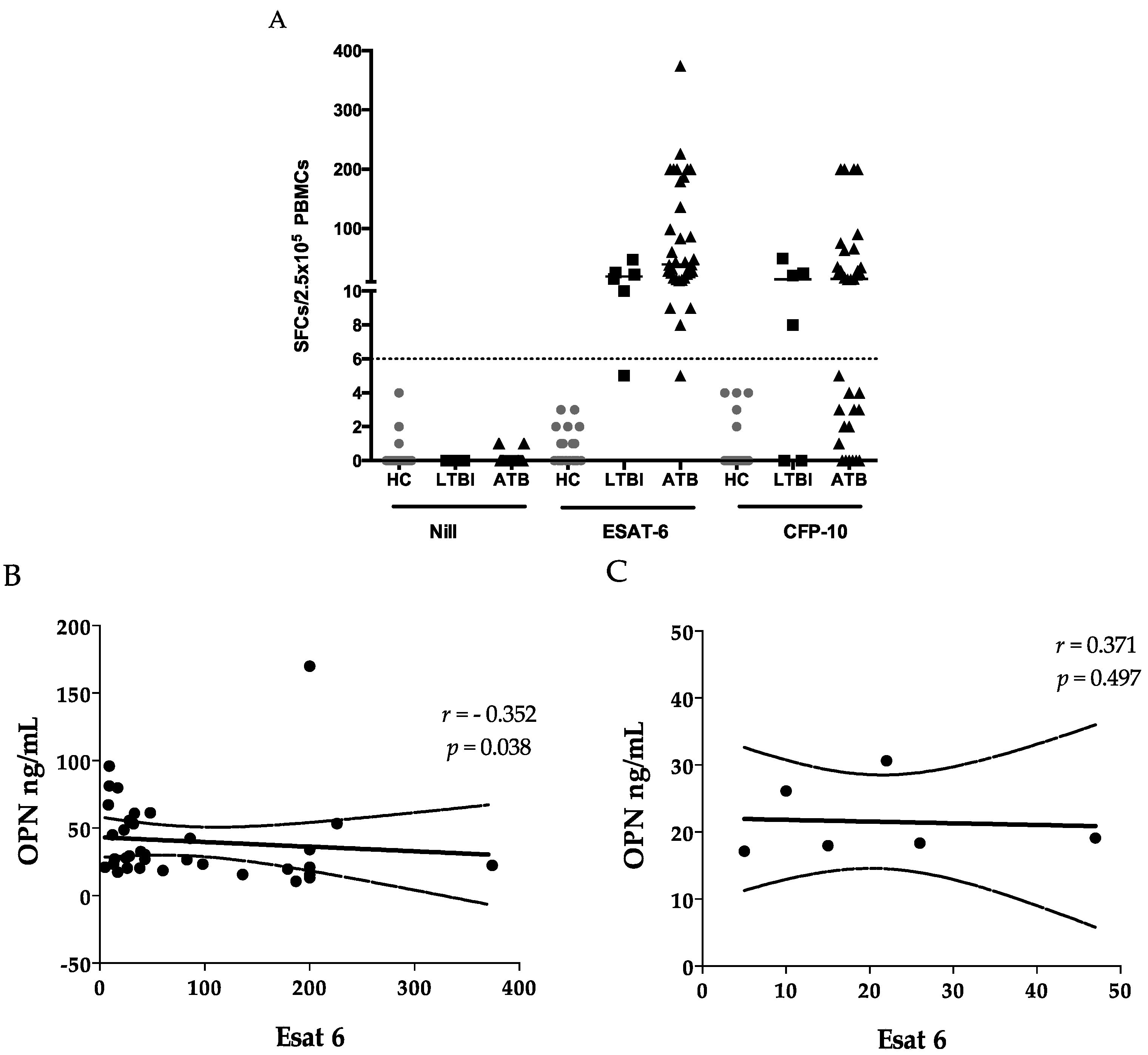
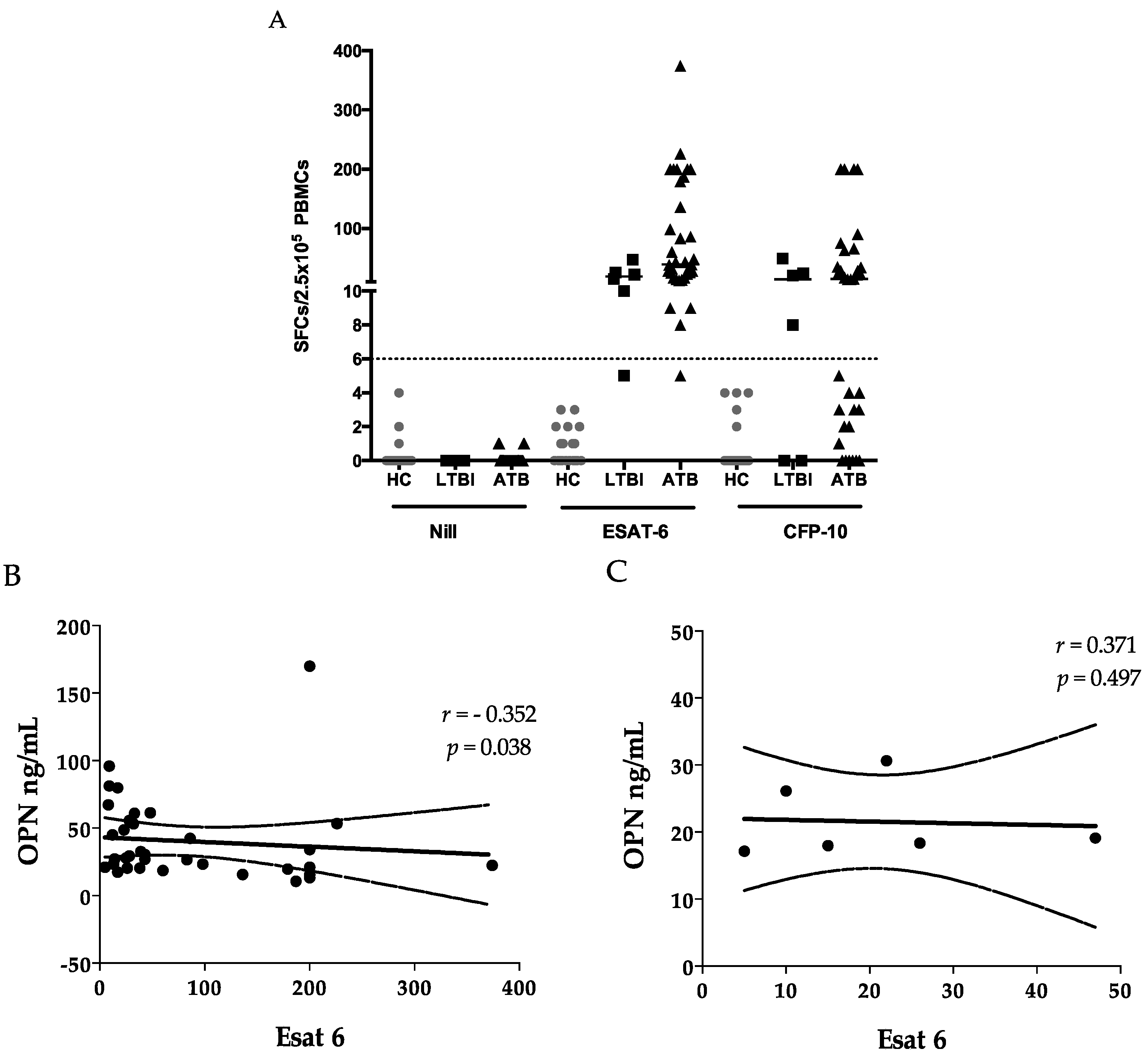
2.3. ELISPOTS
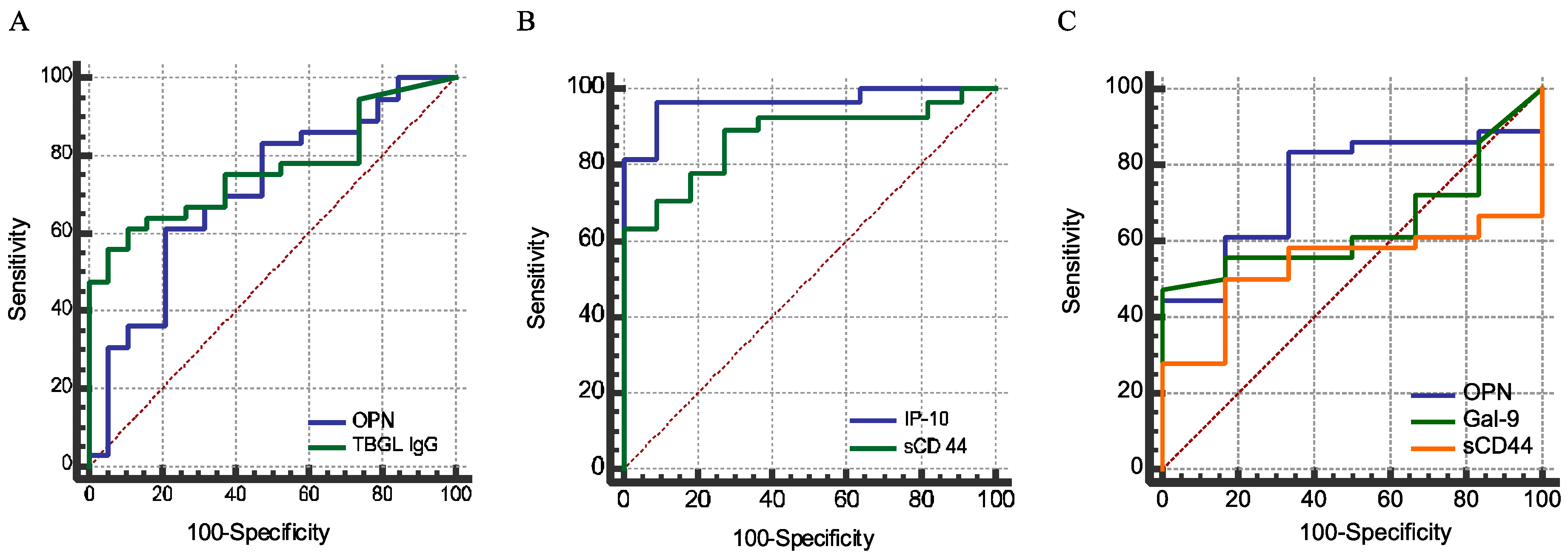
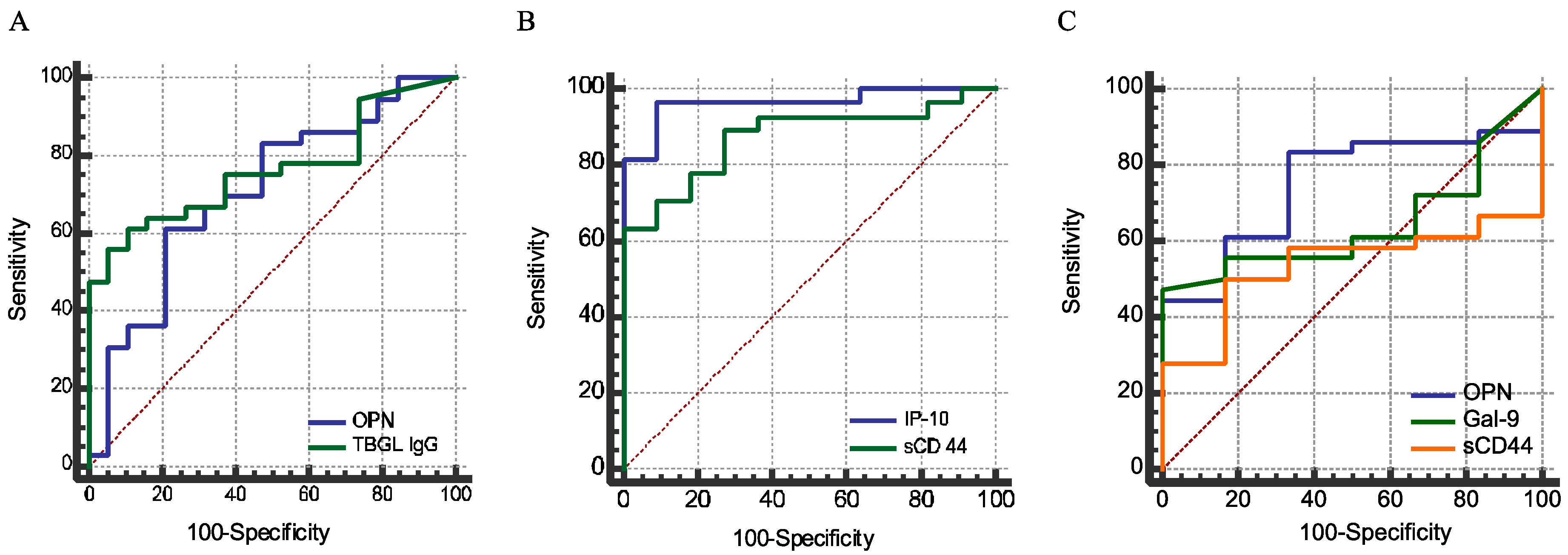
2.4. Sensitivity and Specificity
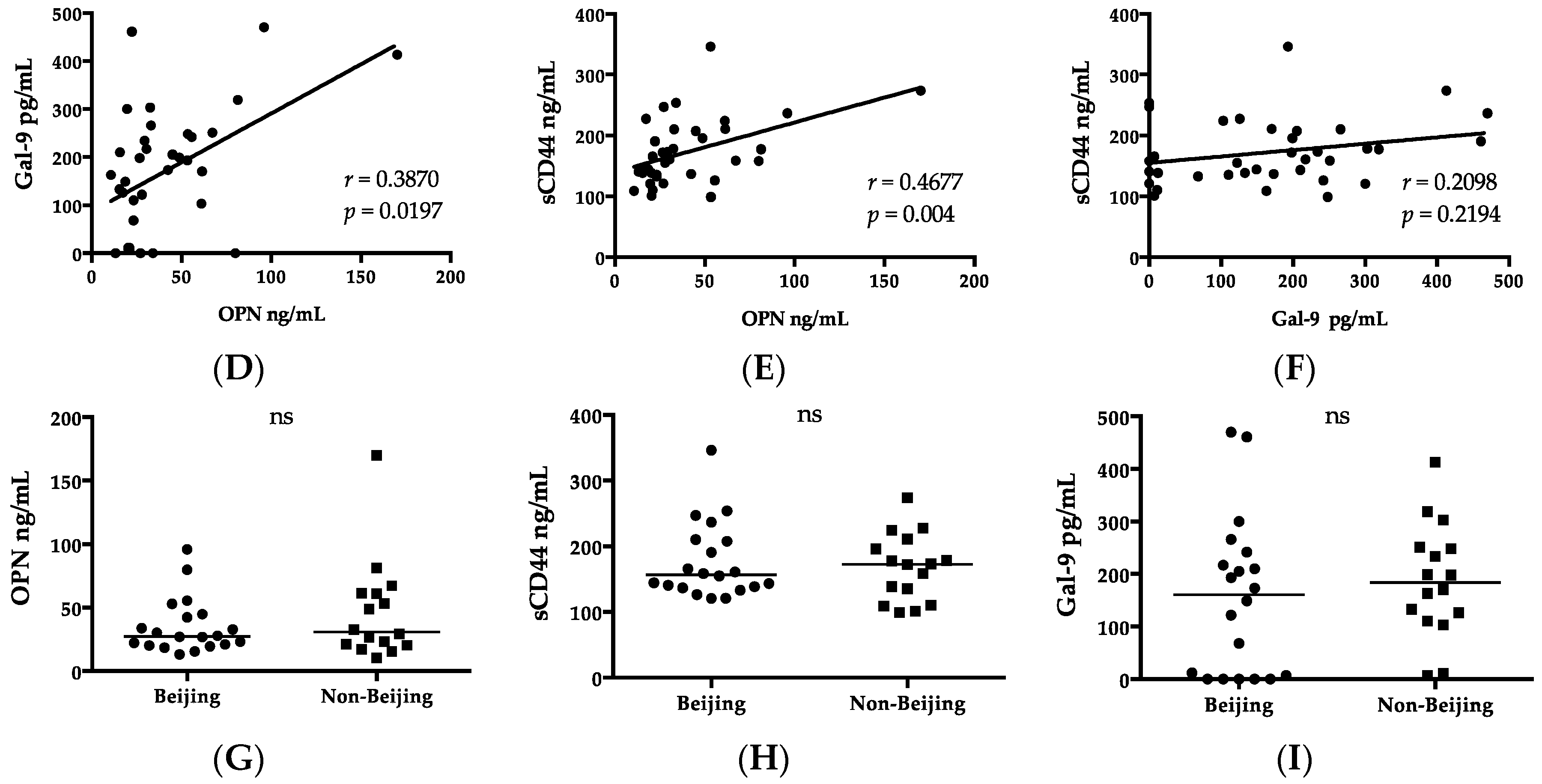
2.5. Clinical Biomarkers, Chest X-rays, and Genotype
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Spoligotyping
4.3. ELISPOTs
4.4. ELISA
4.5. Luminex Assays
4.6. Data Analyses
5. Conclusions
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Bornstein, P.; Sage, E.H. Matricellular proteins: Extracellular modulators of cell function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef]
- Elola, M.T.; Wolfenstein-Todel, C.; Troncoso, M.F.; Vasta, G.R.; Rabinovich, G.A. Galectins: Matricellular glycan-binding proteins linking cell adhesion, migration, and survival. Cell. Mol. Life Sci. CMLS 2007, 64, 1679–1700. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Biol. J. Int. Soc. Matrix Biol. 2014, 37, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Singh, S.; Master, S.; Harris, J.; Roberts, E.; Kyei, G.; Davis, A.; de Haro, S.; Naylor, J.; Lee, H.H.; et al. Mycobacterium tuberculosis inhibition of phagolysosome biogenesis and autophagy as a host defence mechanism. Cell. Microbiol. 2006, 8, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Elkington, P.T.; Emerson, J.E.; Lopez-Pascua, L.D.; O'Kane, C.M.; Horncastle, D.E.; Boyle, J.J.; Friedland, J.S. Mycobacterium tuberculosis up-regulates matrix metalloproteinase-1 secretion from human airway epithelial cells via a p38 MAPK switch. J. Immunol. 2005, 175, 5333–5340. [Google Scholar] [CrossRef] [PubMed]
- Dheda, K.; Booth, H.; Huggett, J.F.; Johnson, M.A.; Zumla, A.; Rook, G.A. Lung remodeling in pulmonary tuberculosis. J. Infect. Dis. 2005, 192, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: Implications for fibrotic diseases and cancer. Dis. Models Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Hasibuan, F.M.; Shiratori, B.; Senoputra, M.A.; Chagan-Yasutan, H.; Koesoemadinata, R.C.; Apriani, L.; Takahashi, Y.; Niki, T.; Alisjahbana, B.; Hattori, T. Evaluation of matricellular proteins in systemic and local immune response to Mycobacterium tuberculosis infection. Microbiol. Immunol. 2015, 59, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, B.; Leano, S.; Nakajima, C.; Chagan-Yasutan, H.; Niki, T.; Ashino, Y.; Suzuki, Y.; Telan, E.; Hattori, T. Elevated OPN, IP-10, and neutrophilia in loop-mediated isothermal amplification confirmed tuberculosis patients. Mediat. Inflamm. 2014, 2014, 513263. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Lacuesta, T.L.; Ndhlovu, L.C.; Oguma, S.; Leano, P.S.A.; Telan, E.F.O.; Kubo, T.; Morita, K.; Uede, T.; Dimaano, E.M.; et al. Elevated levels of full-length and thrombin-cleaved osteopontin during acute dengue virus infection are associated with coagulation abnormalities. Thromb. Res. 2014, 134, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Sada-Ovalle, I.; Beladi, S.; Anderson, A.C.; Dardalhon, V.; Hotta, C.; Kuchroo, V.K.; Behar, S.M. Tim3 binding to galectin-9 stimulates antimicrobial immunity. J. Exp. Med. 2010, 207, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Sada-Ovalle, I.; Chavez-Galan, L.; Torre-Bouscoulet, L.; Nava-Gamino, L.; Barrera, L.; Jayaraman, P.; Torres-Rojas, M.; Salazar-Lezama, M.A.; Behar, S.M. The Tim3–galectin 9 pathway induces antibacterial activity in human macrophages infected with Mycobacterium tuberculosis. J. Immunol. 2012, 189, 5896–5902. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Jacques, M.K.; Zhu, C.; Steblenko, K.M.; Stowell, B.L.; Madi, A.; Anderson, A.C.; Kuchroo, V.K.; Behar, S.M. Tim3 mediates T cell exhaustion during Mycobacterium tuberculosis infection. PLoS Pathog. 2016, 12, e1005490. [Google Scholar] [CrossRef] [PubMed]
- Oomizu, S.; Arikawa, T.; Niki, T.; Kadowaki, T.; Ueno, M.; Nishi, N.; Yamauchi, A.; Hattori, T.; Masaki, T.; Hirashima, M. Cell surface galectin-9 expressing th cells regulate Th17 and Foxp3+ treg development by galectin-9 secretion. PLoS ONE 2012, 7, e48574. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Chen, J.; Liao, H.; Zhang, Y.; Wang, H.; Li, S.; Luo, Y.; Fang, D.; Li, G.; Zhou, B.; et al. Tim-3-expressing CD4+ and CD8+ T cells in human tuberculosis (TB) exhibit polarized effector memory phenotypes and stronger anti-TB effector functions. PLoS Pathog. 2012, 8, e1002984. [Google Scholar] [CrossRef] [PubMed]
- Sada-Ovalle, I.; Ocana-Guzman, R.; Perez-Patrigeon, S.; Chavez-Galan, L.; Sierra-Madero, J.; Torre-Bouscoulet, L.; Addo, M.M. Tim-3 blocking rescue macrophage and T cell function against Mycobacterium tuberculosis infection in HIV+ patients. J. Int. AIDS Soc. 2015, 18, 20078. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Ndhlovu, L.C.; Lacuesta, T.L.; Kubo, T.; Leano, P.S.; Niki, T.; Oguma, S.; Morita, K.; Chew, G.M.; Barbour, J.D.; et al. Galectin-9 plasma levels reflect adverse hematological and immunological features in acute dengue virus infection. J. Clin. Virol. 2013, 58, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Dembele, B.P.; Chagan-Yasutan, H.; Niki, T.; Ashino, Y.; Tangpukdee, N.; Shinichi, E.; Krudsood, S.; Kano, S.; Hattori, T. Plasma levels of galectin-9 reflect disease severity in malaria infection. Malar. J. 2016, 15, 403. [Google Scholar] [CrossRef] [PubMed]
- Merani, S.; Chen, W.; Elahi, S. The bitter side of sweet: The role of galectin-9 in immunopathogenesis of viral infections. Rev. Med. Virol. 2015, 25, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and Tigit: Co-inhibitory receptors with specialized functions in immune regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Tsukasaki, K.; Takahashi, Y.; Oguma, S.; Harigae, H.; Ishii, N.; Zhang, J.; Fukumoto, M.; Hattori, T. Involvement of osteopontin and its signaling molecule CD44 in clinicopathological features of adult T cell leukemia. Leuk. Res. 2011, 35, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yamada, O.; Kida, S.; Matsushita, Y.; Yamaoka, S.; Chagan-Yasutan, H.; Hattori, T. Identification of CD44 as a downstream target of noncanonical NF-kappab pathway activated by human T-cell leukemia virus type 1-encoded tax protein. Virology 2011, 413, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.S.; Yakovlev, S.; Medved, L.; Konstantopoulos, K. Biomolecular characterization of CD44-fibrin(ogen) binding: Distinct molecular requirements mediate binding of standard and variant isoforms of CD44 to immobilized fibrin (ogen). J. Biol. Chem. 2009, 284, 1177–1189. [Google Scholar] [CrossRef] [PubMed]
- Baaten, B.J.; Li, C.R.; Deiro, M.F.; Lin, M.M.; Linton, P.J.; Bradley, L.M. CD44 regulates survival and memory development in Th1 cells. Immunity 2010, 32, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Goletti, D.; Butera, O.; Bizzoni, F.; Casetti, R.; Girardi, E.; Poccia, F. Region of difference 1 antigen-specific CD4+ memory T cells correlate with a favorable outcome of tuberculosis. J. Infect. Dis. 2006, 194, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Thalhamer, T.; Franca, R.F.; Xiao, S.; Wang, C.; Hotta, C.; Zhu, C.; Hirashima, M.; Anderson, A.C.; Kuchroo, V.K. Galectin-9–CD44 interaction enhances stability and function of adaptive regulatory T cells. Immunity 2014, 41, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Zhang, C.L.; Ji, B.Y.; Zhang, L.Z.; Shao, Y.Z.; Jiang, S.L.; Suzuki, Y.; Nakajima, C.; Fan, C.L.; et al. Genotypes and characteristics of clustering and drug susceptibility of Mycobacterium tuberculosis isolates collected in Heilongjiang province, China. J. Clin. Microbiol. 2011, 49, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Chiacchio, T.; Petruccioli, E.; Vanini, V.; Cuzzi, G.; Pinnetti, C.; Sampaolesi, A.; Antinori, A.; Girardi, E.; Goletti, D. Polyfunctional T-cells and effector memory phenotype are associated with active TB in HIV-infected patients. J. Infect. 2014, 69, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Petruccioli, E.; Petrone, L.; Vanini, V.; Sampaolesi, A.; Gualano, G.; Girardi, E.; Palmieri, F.; Goletti, D. IFNγ/TNFα specific-cells and effector memory phenotype associate with active tuberculosis. J. Infect. 2013, 66, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, S.; Andersson, J. Local immune responses in human tuberculosis: Learning from the site of infection. J. Infect. Dis. 2012, 205, S316–S324. [Google Scholar] [CrossRef] [PubMed]
- Koguchi, Y.; Kawakami, K.; Uezu, K.; Fukushima, K.; Kon, S.; Maeda, M.; Nakamoto, A.; Owan, I.; Kuba, M.; Kudeken, N.; et al. High plasma osteopontin level and its relationship with interleukin-12-mediated type 1 T helper cell response in tuberculosis. Am. J. Respir. Crit. Care Med. 2003, 167, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Uede, T. Osteopontin, intrinsic tissue regulator of intractable inflammatory diseases. Pathol. Int. 2011, 61, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Shinohara, M.L. Cutting edge: Role of osteopontin and integrin α V in T cell-mediated anti-inflammatory responses in endotoxemia. J. Immunol. 2015, 194, 5595–5598. [Google Scholar] [CrossRef] [PubMed]
- Nau, G.J.; Chupp, G.L.; Emile, J.F.; Jouanguy, E.; Berman, J.S.; Casanova, J.L.; Young, R.A. Osteopontin expression correlates with clinical outcome in patients with mycobacterial infection. Am. J. Pathol. 2000, 157, 37–42. [Google Scholar] [CrossRef]
- Nau, G.J.; Guilfoile, P.; Chupp, G.L.; Berman, J.S.; Kim, S.J.; Kornfeld, H.; Young, R.A. A chemoattractant cytokine associated with granulomas in tuberculosis and silicosis. Proc. Natl. Acad. Sci. USA 1997, 94, 6414–6419. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, L.; Gurnani, K.; Dicaire, C.J.; van Faassen, H.; Zafer, A.; Kirschning, C.J.; Sad, S.; Sprott, G.D. Rapid clonal expansion and prolonged maintenance of memory CD8+ T cells of the effector (CD44high CD62llow) and central (CD44high CD62lhigh) phenotype by an archaeosome adjuvant independent of TLR2. J. Immunol. 2007, 178, 2396–2406. [Google Scholar] [CrossRef] [PubMed]
- Gey van Pittius, N.C.; Warren, R.M.; van Helden, P.D. ESAT-6 and CFP-10: What is the diagnosis? Infect. Immunity 2002, 70, 6509–6510. [Google Scholar] [CrossRef]
- Ridruechai, C.; Sakurada, S.; Yanai, H.; Yamada, N.; Kantipong, P.; Piyaworawong, S.; Dhepakson, P.; Khusmith, S.; Keicho, N. Association between circulating full-length osteopontin and IFN-γ with disease status of tuberculosis and response to successful treatment. Southeast Asian J. Trop. Med. Public Health 2011, 42, 876–889. [Google Scholar] [PubMed]
- Schnittman, S.M.; Lane, H.C.; Greenhouse, J.; Justement, J.S.; Baseler, M.; Fauci, A.S. Preferential infection of CD4+ memory T cells by human immunodeficiency virus type 1: Evidence for a role in the selective T-cell functional defects observed in infected individuals. Proc. Natl. Acad. Sci. USA 1990, 87, 6058–6062. [Google Scholar] [CrossRef] [PubMed]
- Groot, F.; van Capel, T.M.; Schuitemaker, J.; Berkhout, B.; de Jong, E.C. Differential susceptibility of naive, central memory and effector memory T cells to dendritic cell-mediated HIV-1 transmission. Retrovirology 2006, 3, 52. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Saitoh, H.; Ashino, Y.; Arikawa, T.; Hirashima, M.; Li, S.; Usuzawa, M.; Oguma, S.; EF, O.T.; Obi, C.L.; et al. Persistent elevation of plasma osteopontin levels in hiv patients despite highly active antiretroviral therapy. Tohoku J. Exp. Med. 2009, 218, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lifson, J.D.; Duan, L.; Schacker, T.W.; Reilly, C.; Carlis, J.; Estes, J.D.; Haase, A.T. Potential roles of follicular dendritic cell-associated osteopontin in lymphoid follicle pathology and repair and in B cell regulation in HIV-1 and SIV infection. J. Infect. Dis. 2005, 192, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, J.B.; Corbishley, C.M.; Grange, J.M. Tuberculosis and the kidney. J. Am. Soc. Nephrol. JASN 2001, 12, 1307–1314. [Google Scholar] [PubMed]
- Berry, M.P.; Graham, C.M.; McNab, F.W.; Xu, Z.; Bloch, S.A.; Oni, T.; Wilkinson, K.A.; Banchereau, R.; Skinner, J.; Wilkinson, R.J.; et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature 2010, 466, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.; Lange, C.; Rao, M.; Balaji, K.N.; Lotze, M.; Schito, M.; Zumla, A.I.; Maeurer, M. Progress in tuberculosis vaccine development and host-directed therapies—A state of the art review. Lancet Respir. Med. 2014, 2, 301–320. [Google Scholar] [CrossRef]
- Zumla, A.; Rao, M.; Dodoo, E.; Maeurer, M. Potential of immunomodulatory agents as adjunct host-directed therapies for multidrug-resistant tuberculosis. BMC Med. 2016, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Dufour, J.H.; Dziejman, M.; Liu, M.T.; Leung, J.H.; Lane, T.E.; Luster, A.D. IFN-γ-inducible protein 10 (IP-10; CXCL10)-deficient mice reveal a role for IP-10 in effector T cell generation and trafficking. J. Immunol. 2002, 168, 3195–3204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.A.; Jia, H.Y.; Chen, J.N.; Cui, G.Y.; Gao, H.; Wei, Y.F.; Lu, C.; Wang, L.; Uede, T.; Diao, H.Y. Decreased osteopontin expression as a reliable prognostic indicator of improvement in pulmonary tuberculosis: Impact of the level of interferon-γ-inducible protein 10. Cell. Physiol. Biochem. 2015, 37, 1983–1996. [Google Scholar] [CrossRef] [PubMed]
- Petrone, L.; Cannas, A.; Vanini, V.; Cuzzi, G.; Aloi, F.; Nsubuga, M.; Sserunkuma, J.; Nazziwa, R.A.; Jugheli, L.; Lukindo, T.; et al. Blood and urine inducible protein 10 as potential markers of disease activity. Int. J. Tuberc. Lung Dis. 2016, 20, 1554–1561. [Google Scholar] [CrossRef] [PubMed]
- Petrone, L.; Chiacchio, T.; Vanini, V.; Petruccioli, E.; Cuzzi, G.; di Giacomo, C.; Pucci, L.; Montalbano, M.; Lionetti, R.; Testa, A.; et al. High urine IP-10 levels associate with chronic HCV infection. J. Infect. 2014, 68, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; di Domenicantonio, A.; Fallahi, P. Autoimmune thyroid disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Bifani, P.J.; Mathema, B.; Kurepina, N.E.; Kreiswirth, B.N. Global dissemination of the Mycobacterium tuberculosis W-Beijing family strains. Trends Microbiol. 2002, 10, 45–52. [Google Scholar] [CrossRef]
- Parwati, I.; van Crevel, R.; van Soolingen, D. Possible underlying mechanisms for successful emergence of the Mycobacterium tuberculosis Beijing genotype strains. Lancet Infect. Dis. 2010, 10, 103–111. [Google Scholar] [CrossRef]
- Zhang, J.; Mi, L.; Wang, Y.; Liu, P.; Liang, H.; Huang, Y.; Lv, B.; Yuan, L. Genotypes and drug susceptibility of Mycobacterium tuberculosis isolates in Shihezi, Xinjiang province, China. BMC Res. Notes 2012, 5, 309. [Google Scholar] [CrossRef] [PubMed]
- Kato-Maeda, M.; Shanley, C.A.; Ackart, D.; Jarlsberg, L.G.; Shang, S.; Obregon-Henao, A.; Harton, M.; Basaraba, R.J.; Henao-Tamayo, M.; Barrozo, J.C.; et al. Beijing sublineages of Mycobacterium tuberculosis differ in pathogenicity in the guinea pig. Clin. Vaccine Immunol. 2012, 19, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, J.; Daniel, D.S.; Yang, C.H.; Sever-Chroneos, Z.; Jagannath, C.; Chroneos, Z.C. Granulocyte-macrophage colony stimulating factor-mediated innate responses in tuberculosis. Tuberculosis 2008, 88, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Who Guidelines on Tuberculosis. Available online: http://www.who.int/publications/guidelines/tuberculosis/en/ (accessed on 24 September 2016).
- Ralph, A.P.; Ardian, M.; Wiguna, A.; Maguire, G.P.; Becker, N.G.; Drogumuller, G.; Wilks, M.J.; Waramori, G.; Tjitra, E.; Sandjaja; et al. A simple, valid, numerical score for grading chest X-ray severity in adult smear-positive pulmonary tuberculosis. Thorax 2010, 65, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Grassinger, J.; Haylock, D.N.; Storan, M.J.; Haines, G.O.; Williams, B.; Whitty, G.A.; Vinson, A.R.; Be, C.L.; Li, S.H.; Sorensen, E.S.; et al. Thrombin-cleaved osteopontin regulates hemopoietic stem and progenitor cell functions through interactions with α9β1 and α4β1 integrins. Blood 2009, 114, 49–59. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Ref. | HC (n = 19) | LTBI (n = 6) | TB (n = 36) | p Value |
|---|---|---|---|---|---|
| Antropometric data | |||||
| Age: year; median (range) | 34 (19–67) | 63 (36–71) | 59.5 (19–86) | 0.007 | |
| Gender: male; n (%) | 12 (63) | 5 (83) | 28 (78) | 0.43 | |
| Laboratory findings: median (range) | |||||
| RBC (106/μL) | Male 4.5–5.5; Female 4.0–5.0 | na | 4.65 (4.35–5.12) | 4.44 (2.72–5.57) | 0.443 |
| Hemoglobin (g/dL) | 14–18 | na | 14.6 (14.2–15.3) | 13.1 (8.9–17.5) | 0.064 |
| Hematocrit (%) | 40–48 | na | 43.6 (40.9–46.0) | 39.65 (25.5–51.9) | 0.059 |
| WBC (103/μL) | 4.5–11 | na | 7.64 (5.34–9.96) | 7.33 (3.84–16.22) | 0.945 |
| Neutrophil (%) | 38–80 | na | 57.35(51.9–75.7) | 75.2 (57.0–89.6) | 0.014 |
| Lymphocyte (%) | 15–40 | na | 33.1 (18.2–39.6) | 14.1 (2.8–34.8) | 0.007 |
| Monocyte (%) | 4–7 | na | 6 (4.9–7.7) | 6.9 (3.4–11.2) | 0.035 |
| Eosinophil (%) | 0–8 | na | 1.1 (0.4–4.1) | 1.4 (0–11.4) | 0.902 |
| Platelet (103/μL) | 140–390 | na | 267 (203–280) | 305 (125–564) | 0.228 |
| CRP (mg/dL) | 0–0.3 | na | 0.055 (0.04–0.07) | 6 (0.04–21.5) | 0.002 |
| Genotype (Beijing strain MTB %) | 73.0, year 2010, Japan | na | na | 55.6 | na |
| ALT (Units) | 4–37 | na | 20.5 (11–29) | 15 (8–103) | na |
| Creatinine (mg/dL) | 0.5–1.5 | na | 0.74 (0.42–11) | 0.71 (0.57–0.86) | na |
| Biomarker | HC | TB | p Value |
|---|---|---|---|
| IFNγ (pg/mL) | 4.46 (0.42–19.38) | 8.13 (2.12–41) | 0.0065 |
| IL-8 (pg/mL) | 1.585 (0.38–7.52) | 7.26 (0.6–31.07) | <0.0001 |
| IP-10 (pg/mL) | 235.8 (132.2–472.8) | 864.6 (219.3–3051) | <0.0001 |
| TNFα (pg/mL) | 3.88 (2.83–8.34) | 10.11 (2.19–24.83) | <0.0001 |
| OPN (ng/mL) | 19.63 (9.31–111.64) | 28.62 (10.59–170) | 0.012 |
| Gal-9 (ng/mL) | 14.0 (0–120) | 171.5 (0–470) | 0.0002 |
| sCD44 (ng/mL) | 118.57 (91.02–141.41) | 159.66 (98.89–346.2) | <0.0001 |
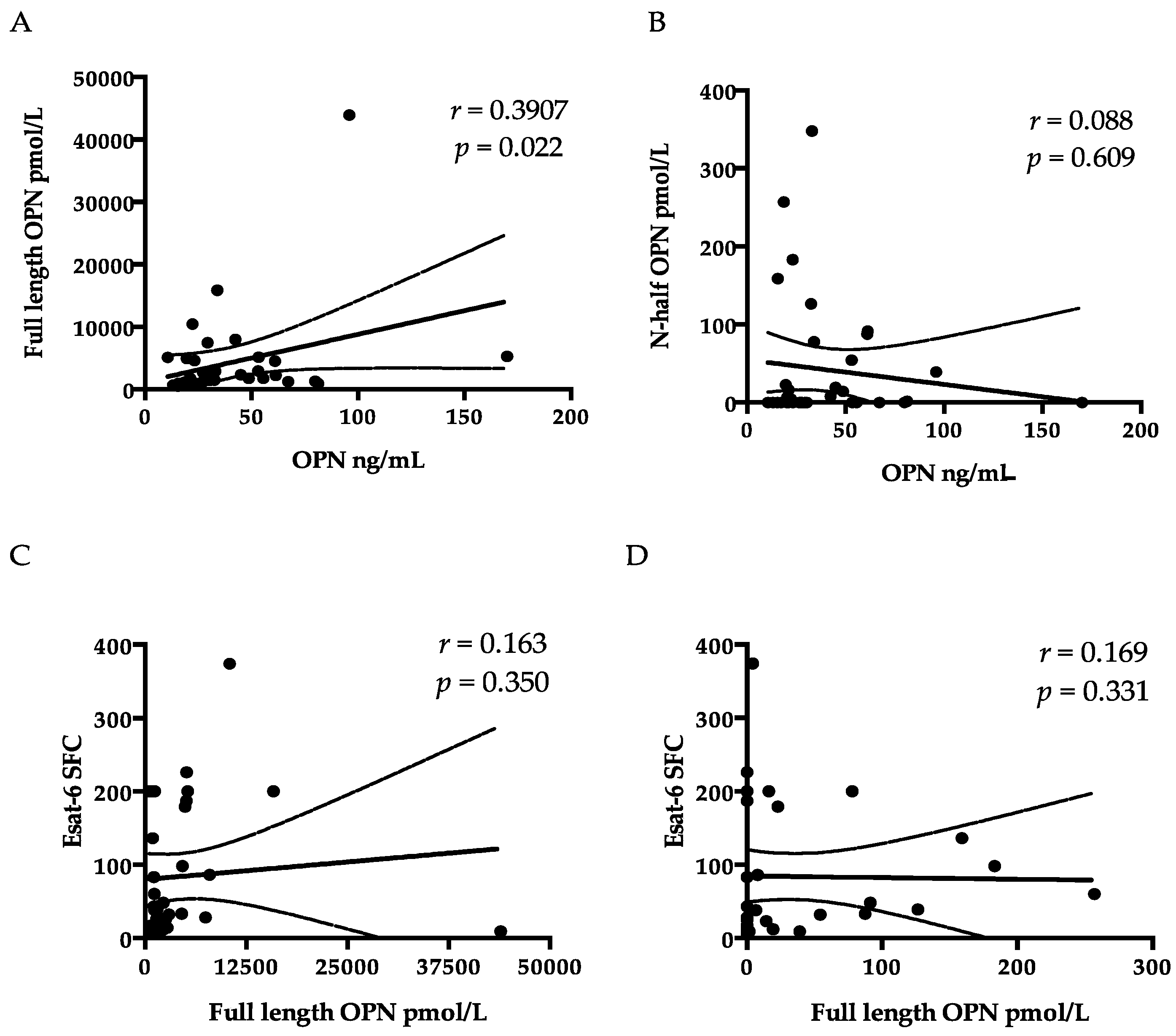
| FL-OPN (nmol/mL) | 5.20 (2.83–14.45) | 1.75 (0.52–43.91) | >0.05 |
| rtOPN (pmol/mL) | 0 (0–26.73) | 0.87 (0–347.95) | >0.05 |
| Measurements | OPN | sCD44 | Gal-9 |
|---|---|---|---|
| r (P) | r (P) | r (P) | |
| WBC (103/μL) | Ns | 0.388 (0.019) | Ns |
| Neutrophil (%) | 0.517 (<0.0001) | Ns | Ns |
| Lymphocyte (%) | −0.569 (<0.0001) | −0.558 (<0.0001) | Ns |
| CRP (mg/dL) | 0.757 (<0.0001) | 0.534 (0.001) | Ns |
| IL-8 (pg/mL) | 0.474 (0.013) | 0.524 (0.005) | Ns |
| IP-10 (pg/mL) | 0.420 (0.029) | 0.542 (0.003) | Ns |
| TNFα | Ns | 0.446 (0.020) | Ns |
| ALT | Ns | Ns | 0.375 (0.024) |
| Cre | Ns | Ns | 0.377 (0.023) |
| Analytes | Youden Index | Cutoff | Sensitivity (%) | Specificity (%) | AUC | Comparison | ||
|---|---|---|---|---|---|---|---|---|
| OPN | 0.4006 | 25.1 | 61.1 | 79 | 0.706 | a | b | |
| Gal-9 | 0.6667 | 120 | 66.7 | 100 | 0.798 | |||
| sCD44 | 0.6389 | 141.4 | 63.9 | 100 | 0.846 | a | ||
| IP-10 | 0.8593 | 400.3 | 92.6 | 93.3 | 0.965 | b | a | |
| Anti-TBGL IgG | 0.5058 | 1.6 | 61.1 | 89.4 | 0.762 | a | ||
| Parameter | Normal Value (Range) | Cavity (−) (n = 24) | Cavity (+) (n = 12) | p Value |
|---|---|---|---|---|
| Hemoglobin (g/dL) | 14–18 | 13.8 (8.9–17.5) | 12.4 (9.9–14.7) | 0.0454 |
| Hematocrit (%) | 40–48 | 42 (25.5–51.9) | 37.5 (28.4–41.4) | 0.0208 |
| Monocyte (%) | 4–7 | 6.2 (3.4–10) | 8.3 (4.9–10.2) | 0.0054 |
| CRP (mg/dL) | 0–0.3 | 3.27 (0.04–21.5) | 8.67 (0.08–17.97) | 0.0478 |
| IP-10 (pg/mL) | 132.2–472.8 | 751.5 (219.3–2735) | 1570 (369.7–3051) | 0.0408 |
| Anti-TBGL IgG (U/mL) | <2 | 13.3 (0.1–62.6) | 1.11 (0–72.6) | 0.0123 |
| CXR Score | 0 | 92.46 (82.4–100) | 131.84 (104.34–140) | 0.0001 |
| Parameter | CXR Scorer (p) | CXR Scorer (p) | |
|---|---|---|---|
| Cavity (+) | Cavity (−) | ||
| Monocytes (%) | 0.502 (0.003) | Ns | Ns |
| OPN | Ns | Ns | Ns |
| sCD44 | Ns | Ns | Ns |
| IP-10 | 0.452 (0.027) | Ns | Ns |
| Gal-9 | Ns | Ns | Ns |
| IL-12p70 | 0.517 (0.01) | Ns | 0.574 (0.032) |
| ESAT-6 SFC | Ns | −0.6185 (0.0425) | Ns |
| Parameter | HC | Beijing Type (n = 20) | Non-Beijing Type (n = 16) | p Value |
|---|---|---|---|---|
| Laboratory findings | ||||
| RBC (106/μL) | na. | 4.28 (2.72–5.16) | 4.64 (3.86–5.57) | 0.0465 |
| Total protein (g/dL) | na. | 7.05 (5.41–8.03) | 7.69 (6.54–8.14) | 0.0323 |
| Biomarker (pg/mL) | ||||
| G-CSF | 48.48 (12.19–90.49) | 56.94 (21.69–106.36) | 38.12 (14.06–73.07) | 0.0388 |
| GM-CSF | 4.24 (0.44–18.98) | 12.29 (0.83–48.8) | 5.84 (0.44–14.6) | 0.0300 |
| IFNα | 11.07 (0–56.49) | 32.87 (1.67–60.81) a | 7.53 (0–40.97) | 0.0034 |
| IFNγ | 4.46 (0.42–19.38) | 16.02 (3.09–41) a | 4.71 (2.12–17.84) | 0.0141 |
| IL-12p70 | 2.74 (1.35–15.06) | 8.22 (1.85–29.77) | 2.62 (0.03–7.13) | 0.0007 |
| IL-1RA | 17.79 (0–81.26) | 51.96 (8.56–217.37) a | 14.02 (0–78.44) | 0.0095 |
| IP-10 | 235.78 (132.2–472.75) | 877.12 (219.29–2814.69) a | 864.57 (369.67–3051.48) b | >0.05 |
| Cavity n (%) | 0 | 8 (40) | 4 (25) | >0.05 |
| CXR score | 0 | 102.23 (82.4–140) | 109.74 (82.77–140) | >0.05 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiratori, B.; Zhao, J.; Okumura, M.; Chagan-Yasutan, H.; Yanai, H.; Mizuno, K.; Yoshiyama, T.; Idei, T.; Ashino, Y.; Nakajima, C.; et al. Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis. Int. J. Mol. Sci. 2017, 18, 19. https://doi.org/10.3390/ijms18010019
Shiratori B, Zhao J, Okumura M, Chagan-Yasutan H, Yanai H, Mizuno K, Yoshiyama T, Idei T, Ashino Y, Nakajima C, et al. Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis. International Journal of Molecular Sciences. 2017; 18(1):19. https://doi.org/10.3390/ijms18010019
Chicago/Turabian StyleShiratori, Beata, Jingge Zhao, Masao Okumura, Haorile Chagan-Yasutan, Hideki Yanai, Kazue Mizuno, Takashi Yoshiyama, Tadashi Idei, Yugo Ashino, Chie Nakajima, and et al. 2017. "Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis" International Journal of Molecular Sciences 18, no. 1: 19. https://doi.org/10.3390/ijms18010019
APA StyleShiratori, B., Zhao, J., Okumura, M., Chagan-Yasutan, H., Yanai, H., Mizuno, K., Yoshiyama, T., Idei, T., Ashino, Y., Nakajima, C., Suzuki, Y., & Hattori, T. (2017). Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis. International Journal of Molecular Sciences, 18(1), 19. https://doi.org/10.3390/ijms18010019