
Rosacea: Molecular Mechanisms and Management of a Chronic Cutaneous Inflammatory Condition


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
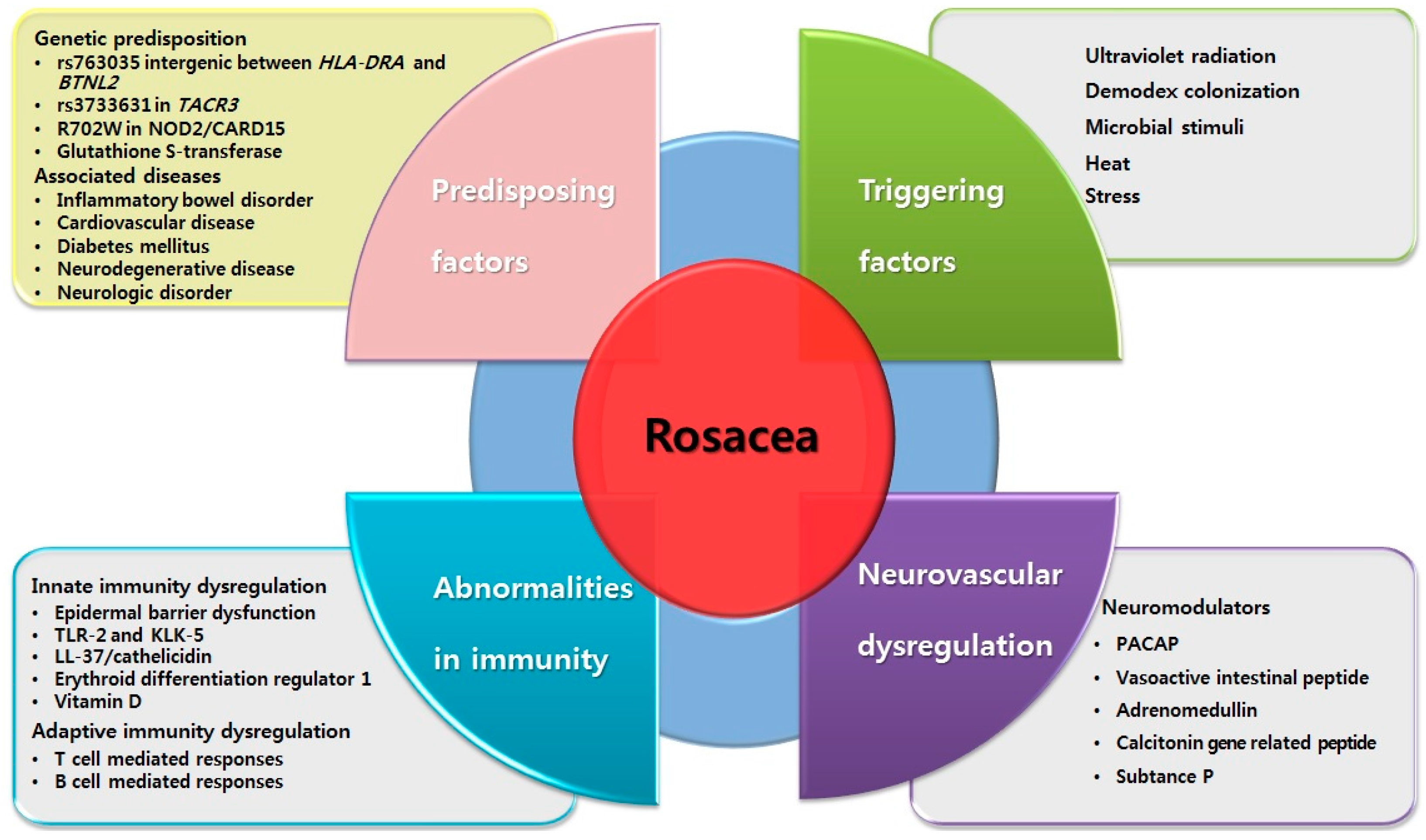
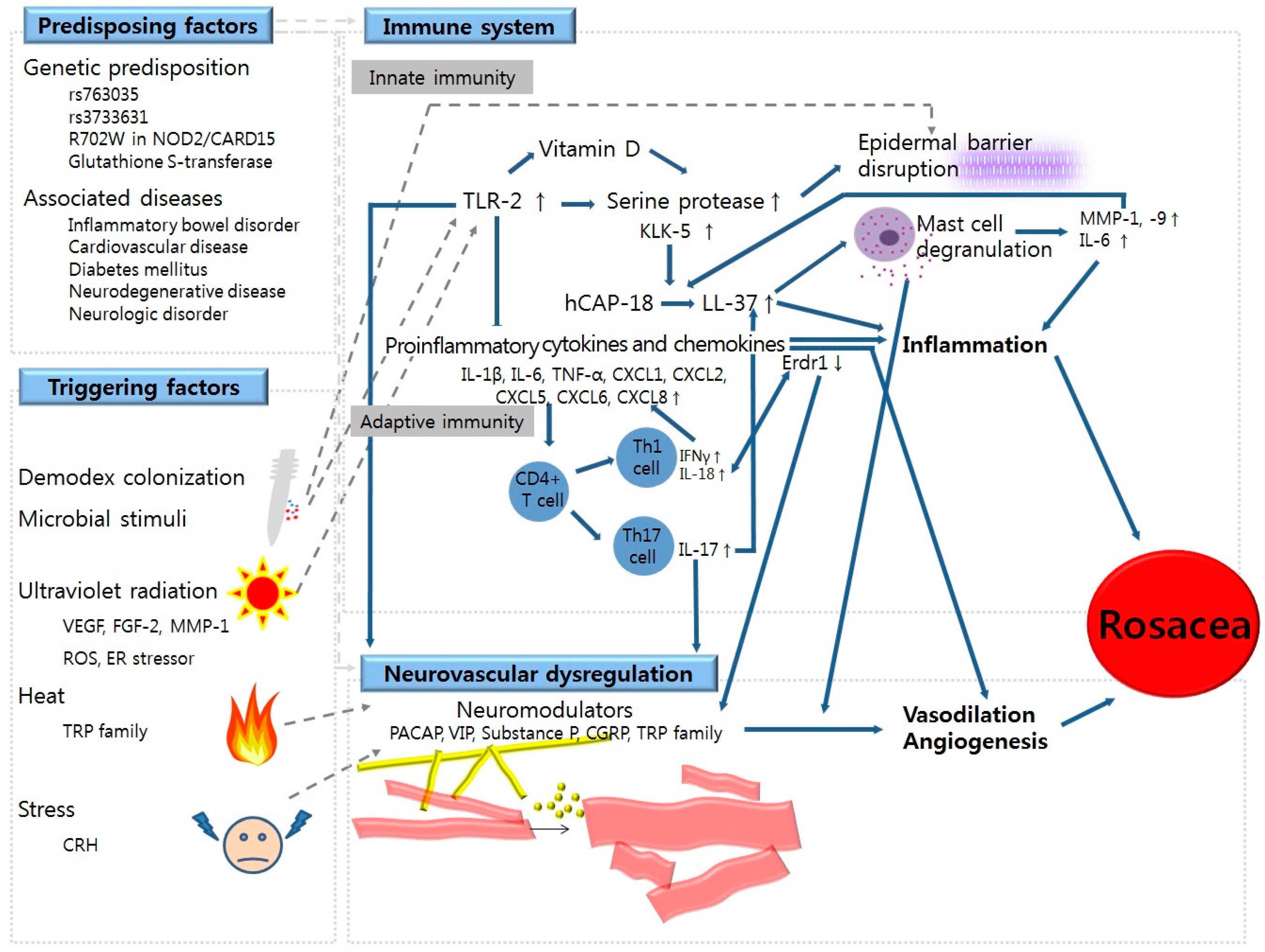
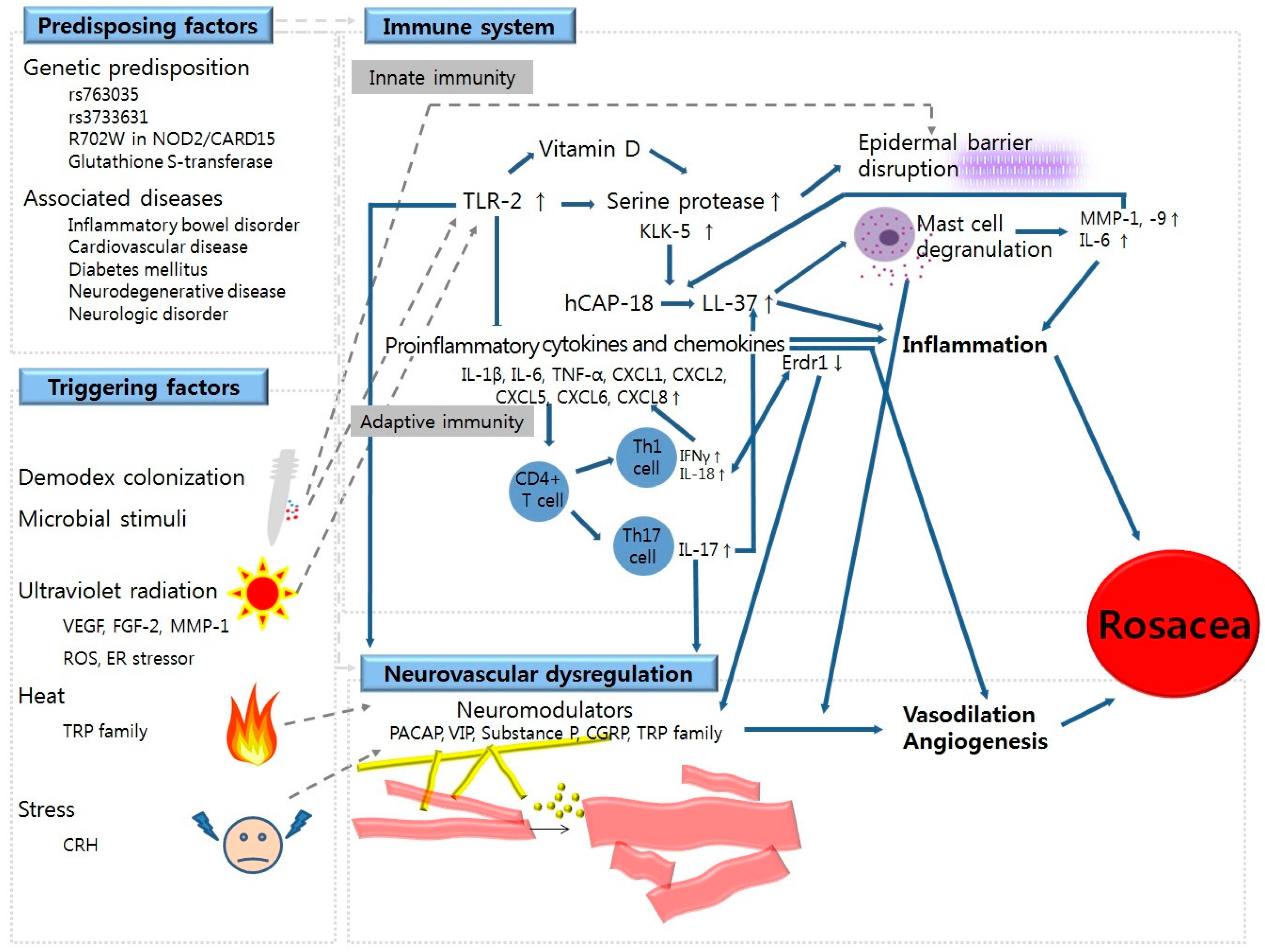
2. Predisposing Factors for Rosacea
2.1. Genetics
2.2. Associated Diseases
3. Triggering Factors for Rosacea
3.1. Ultraviolet (UV) Radiation
3.2. Demodex Colonization
3.3. Microbial Stimuli
3.4. Heat
3.5. Stress
4. Abnormalities in Immune System
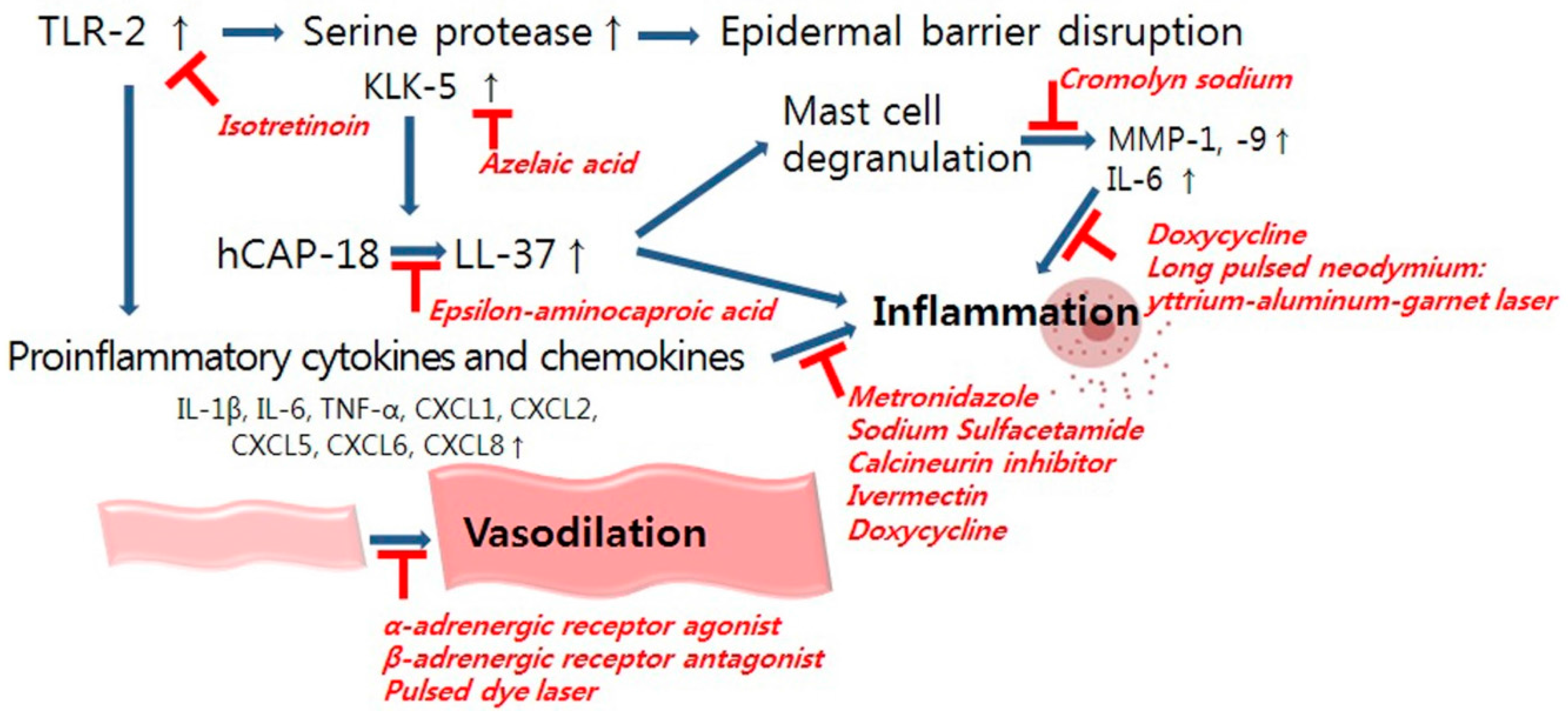
4.1. Dysregulation in Innate Immunity
4.1.1. Epidermal Barrier Disruption
4.1.2. TLR-2 and KLK-5
4.1.3. LL-37/Cathelicidin
4.1.4. Vitamin D
4.2. Dysregulation in Adaptive Immunity
4.2.1. T Cell-Mediated Responses
4.2.2. B Cell-Mediated Responses
5. Neurovascular Dysregulation
6. Treatment Options for Rosacea
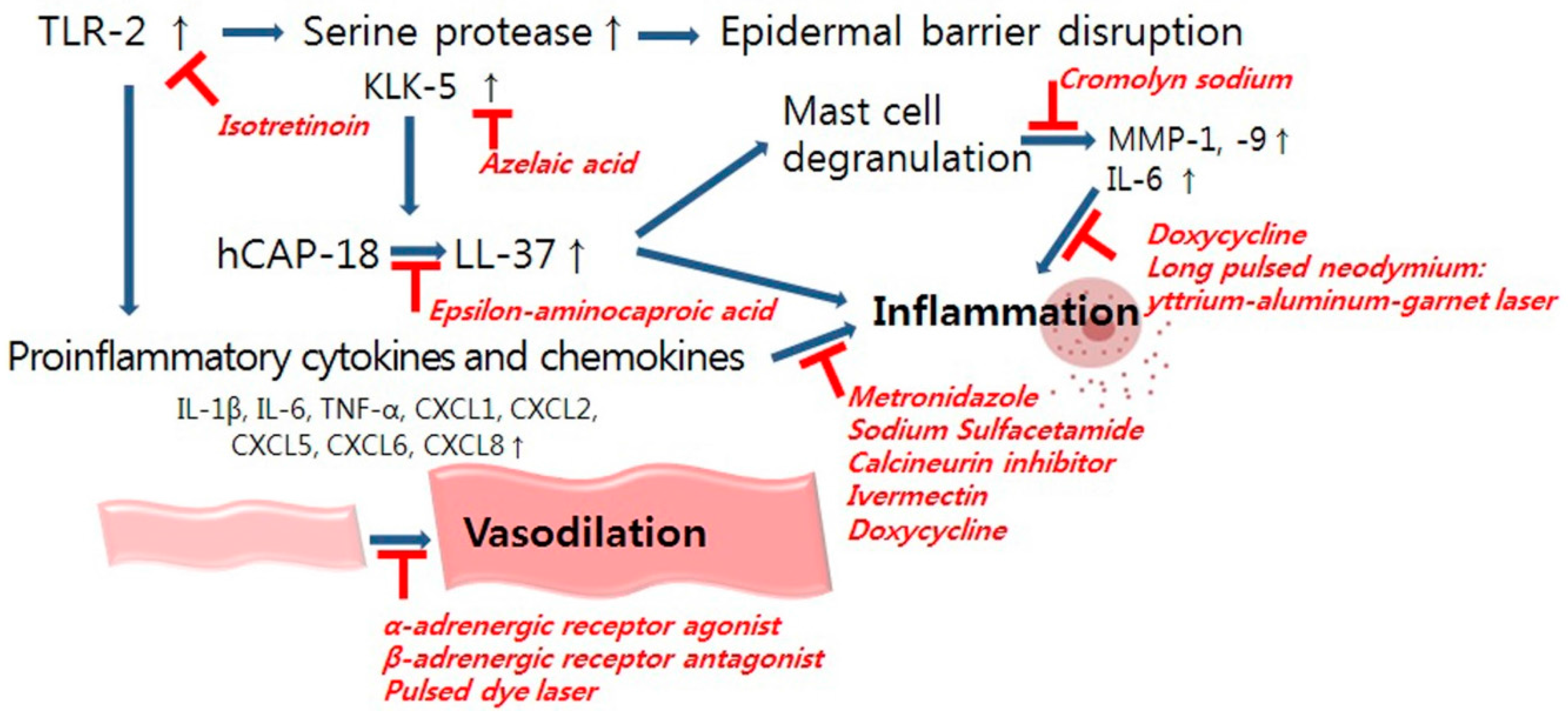
6.1. Management of Inflammation
6.2. Management of Vascular Dysregulation
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AMPs | Antimicrobial peptides |
| BRNL2 | Butyrophilin-like 2 |
| CAMP | Cathelicidin antimicrobial peptide |
| CGRP | Calcitonin gene-related peptide |
| CRH | Corticotrophin- releasing hormone |
| CRH-R | Cortisol releasing hormone receptor |
| ER | Endoplasmic reticulum |
| Erdr1 | Erythroid differentiation regulator 1 |
| GST | Glutathione S-transferase |
| GWAS | Genome-wide association study |
| HLA | Human leukocyte antigen |
| IL | Interleukin |
| IPL | Intense pulsed light |
| KLK | Kallikrein |
| LPND | Long pulsed 1064-nm neodymium: yttrium-aluminum-garnet laser |
| MMP | Matrix metalloproteinase |
| MyD88 | Myeloid differentiation factor 88 |
| NF-κB | Nuclear factor κ-light-chain-enhancers of activated B cells |
| NLRP3 | Nucleotide-binding oligomerization domain-like receptor family, pyrin domain containing 3 |
| NO | Nitric oxid |
| PACAP | Pituitary adenylate cyclase-activating polypeptide |
| PAR-2 | Protease-activated receptor 2 |
| PDL | Pulsed dye laser |
| PERK | Protein kinase R-like endoplasmic reticulum kinase |
| PRRs | Pattern recognition receptors |
| ROS | Reactive oxygen species |
| SNP | Single-nucleotide polymorphism |
| SPINK | Serine protease inhibitor of Kazal-type |
| SSNA | skin sympathetic nerve activity |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
| TRPV1 | Transient receptor potential cation channel subfamily V member 1 |
| UV | Ultraviolet |
| VEGF | Vascular endothelial growth factor |
| VDR | Vitamin D receptor |
| VEGF | Vascular endothelial growth factor |
| VIP | Vasoactive intestinal peptide |
References
- Steinhoff, M.; Schauber, J.; Leyden, J.J. New insights into rosacea pathophysiology: A review of recent findings. J. Am. Acad. Dermatol. 2013, 69, S15–S26. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Berg, M. Rosacea: Current state of epidemiology. J. Am. Acad. Dermatol. 2013, 69, S27–S35. [Google Scholar] [CrossRef] [PubMed]
- Two, A.M.; Wu, W.; Gallo, R.L.; Hata, T.R. Rosacea: Part I. Introduction, categorization, histology, pathogenesis, and risk factors. J. Am. Acad. Dermatol. 2015, 72, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Spoendlin, J.; Voegel, J.J.; Jick, S.S.; Meier, C.R. A study on the epidemiology of rosacea in the U.K. Br. J. Dermatol. 2012, 167, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Chosidow, O.; Cribier, B. Epidemiology of rosacea: Updated data. Ann. Dermatol. Venereol. 2011, 138 (Suppl. 3), S179–S183. [Google Scholar] [CrossRef]
- Hong, E.; Fischer, G. Childhood ocular rosacea: Considerations for diagnosis and treatment. Australas. J. Dermatol. 2009, 50, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Kroshinsky, D.; Glick, S.A. Pediatric rosacea. Dermatol. Ther. 2006, 19, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Allah, K.C.; Kossoko, H.; Yéo, S.; Richard Kadio, M.; Assi Djè Bi Djè, V. Rhinophyma in a black African male patient. Rev. Stomatol. Chir. Maxillofac. 2009, 110, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.I.; Yun, S.J.; Lee, J.B.; Kim, S.J.; Won, Y.H.; Lee, S.C. Clinical evaluation of 168 korean patients with rosacea: The sun exposure correlates with the erythematotelangiectatic subtype. Ann. Dermatol. 2009, 21, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Schmelz, M.; Schauber, J. Facial erythema of rosacea—Aetiology, different pathophysiologies and treatment options. Acta Derm. Venereol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Liu, F.T. Rosacea: Skin innate immunity gone awry? Nat. Med. 2007, 13, 904–906. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2007, 13, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.D.; Steinhoff, M. Integrative concepts of rosacea pathophysiology, clinical presentation, and new therapeutics. Exp. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Buhl, T.; Sulk, M.; Nowak, P.; Buddenkotte, J.; McDonald, I.; Aubert, J.; Carlavan, I.; Deret, S.; Reiniche, P.; Rivier, M.; et al. Molecular and morphological characterization of inflammatory infiltrate in rosacea reveals activation of Th1/Th17 pathways. J. Investig. Dermatol. 2015, 135, 2198–2208. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, N.; Gerstenblith, M.; Fu, P.; Tuttle, M.S.; Varma, P.; Gotow, E.; Cooper, K.D.; Mann, M.; Popkin, D.L. Genetic vs. environmental factors that correlate with rosacea: A cohort-based survey of twins. JAMA Dermatol. 2015, 151, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Akilov, O.E.; Mumcuoglu, K.Y. Association between human demodicosis and HLA class I. Clin. Exp. Dermatol. 2003, 28, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Abram, K.; Silm, H.; Maaroos, H.I.; Oona, M. Risk factors associated with rosacea. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Palleschi, G.M.; Torchia, D. Rosacea in a monozygotic twin. Australas. J. Dermatol. 2007, 48, 132–133. [Google Scholar] [CrossRef] [PubMed]
- Yazici, A.C.; Tamer, L.; Ikizoglu, G.; Kaya, T.I.; Api, H.; Yildirim, H.; Adiguzel, A. GSTM1 and GSTT1 null genotypes as possible heritable factors of rosacea. Photodermatol. Photoimmunol. Photomed. 2006, 22, 208–210. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.L.; Raber, I.; Xu, J.; Li, R.; Spitale, R.; Chen, J.; Kiefer, A.K.; Tian, C.; Eriksson, N.K.; Hinds, D.A.; et al. Assessment of the genetic basis of rosacea by genome-wide association study. J. Investig. Dermatol. 2015, 135, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Van Steensel, M.A.; Badeloe, S.; Winnepenninckx, V.; Vreeburg, M.; Steijlen, P.M.; van Geel, M. Granulomatous rosacea and Crohn’s disease in a patient homozygous for the Crohn-associated NOD2/CARD15 polymorphism R702W. Exp. Dermatol. 2008, 17, 1057–1058. [Google Scholar] [CrossRef] [PubMed]
- Karpouzis, A.; Avgeridis, P.; Tripsianis, G.; Gatzidou, E. Assessment of tachykinin receptor 3′ gene polymorphism rs3733631 in rosacea. Int. Sch. Res. Not. 2015, 2015, 469402. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, M.S.; Cho, J.H.; Rioux, J.D.; McGovern, D.P.; Wu, J.; Annese, V.; Achkar, J.P.; Goyette, P.; Scott, R.; Xu, W.; et al. Ulcerative colitis-risk loci on chromosomes 1p36 and 12q15 found by genome-wide association study. Nat. Genet. 2009, 41, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, L.B.; Steinhoff, M. Rosacea and small intestinal bacterial overgrowth: Prevalence and response to rifaximin. J. Am. Acad. Dermatol. 2013, 68, 875–876. [Google Scholar] [CrossRef] [PubMed]
- Spoendlin, J.; Karatas, G.; Furlano, R.I.; Jick, S.S.; Meier, C.R. Rosacea in patients with ulcerative colitis and crohn’s disease: a population-based case-control study. Inflamm. Bowel Dis. 2016, 22, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, A.; Hansen, P.R.; Gislason, G.H.; Thyssen, J.P. Clustering of autoimmune diseases in patients with rosacea. J. Am. Acad. Dermatol. 2016, 74, 667–672.e1. [Google Scholar] [CrossRef] [PubMed]
- Hua, T.C.; Chung, P.I.; Chen, Y.J.; Wu, L.C.; Chen, Y.D.; Hwang, C.Y.; Chu, S.Y.; Chen, C.C.; Lee, D.D.; Chang, Y.T.; et al. Cardiovascular comorbidities in patients with rosacea: A nationwide case-control study from Taiwan. J. Am. Acad. Dermatol. 2015, 73, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Duman, N.; Ersoy Evans, S.; Atakan, N. Rosacea and cardiovascular risk factors: A case control study. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Rainer, B.M.; Fischer, A.H.; Luz Felipe da Silva, D.; Kang, S.; Chien, A.L. Rosacea is associated with chronic systemic diseases in a skin severity-dependent manner: Results of a case-control study. J. Am. Acad. Dermatol. 2015, 73, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Takci, Z.; Bilgili, S.G.; Karadag, A.S.; Kucukoglu, M.E.; Selek, S.; Aslan, M. Decreased serum paraoxonase and arylesterase activities in patients with rosacea. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.K.; Meher, L.K.; Kota, S.K.; Jammula, S.; Krishna, S.V.; Modi, K.D. Implications of serum paraoxonase activity in obesity, diabetes mellitus, and dyslipidemia. Indian J. Endocrinol. Metab. 2013, 17, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Akin Belli, A.; Ozbas Gok, S.; Akbaba, G.; Etgu, F.; Dogan, G. The relationship between rosacea and insulin resistance and metabolic syndrome. Eur. J. Dermatol. 2016, 26, 260–264. [Google Scholar] [PubMed]
- Spoendlin, J.; Voegel, J.J.; Jick, S.S.; Meier, C.R. Risk of rosacea in patients with diabetes using insulin or oral antidiabetic drugs. J. Investig. Dermatol. 2013, 133, 2790–2793. [Google Scholar] [CrossRef] [PubMed]
- Spoendlin, J.; Voegel, J.J.; Jick, S.S.; Meier, C.R. Migraine, triptans, and the risk of developing rosacea: A population-based study within the United Kingdom. J. Am. Acad. Dermatol. 2013, 69, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, A.; Hansen, P.R.; Gislason, G.H.; Thyssen, J.P. Association of rosacea with risk for glioma in a Danish nationwide cohort study. JAMA Dermatol. 2016, 152, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, A.; Hansen, P.R.; Gislason, G.H.; Thyssen, J.P. Exploring the association between rosacea and Parkinson disease: A Danish nationwide cohort study. JAMA Neurol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A. Matrix metalloproteinases and their multiple roles in neurodegenerative diseases. Lancet Neurol. 2009, 8, 205–216. [Google Scholar] [CrossRef]
- Egeberg, A.; Hansen, P.R.; Gislason, G.H.; Thyssen, J.P. Patients with rosacea have increased risk of dementia. Ann. Neurol. 2016, 79, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Wlaschek, M.; Bolsen, K.; Herrmann, G.; Schwarz, A.; Wilmroth, F.; Heinrich, P.C.; Goerz, G.; Scharffetter-Kochanek, K. UVA-induced autocrine stimulation of fibroblast-derived-collagenase by IL-6: A possible mechanism in dermal photodamage? J. Investig. Dermatol. 1993, 101, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Cribier, B. Rosacea under the microscope: Characteristic histological findings. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Brauchle, M.; Funk, J.O.; Kind, P.; Werner, S. Ultraviolet B and H2O2 are potent inducers of vascular endothelial growth factor expression in cultured keratinocytes. J. Biol. Chem. 1996, 271, 21793–21797. [Google Scholar] [PubMed]
- Bielenberg, D.R.; Bucana, C.D.; Sanchez, R.; Donawho, C.K.; Kripke, M.L.; Fidler, I.J. Molecular regulation of UVB-induced cutaneous angiogenesis. J. Investig. Dermatol. 1998, 111, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Gray, B.; Metzler-Wilson, K.; Dawes, K.; Wilson, T. A neural link to understanding rosacea: Focusing on flushing triggers. J. Am. Osteopath. Coll. Dermatol. 2003, 33, 11–16. [Google Scholar]
- Jones, D. Reactive oxygen species and rosacea. Cutis 2004, 74, 17–20. [Google Scholar] [PubMed]
- Vemuri, R.C.; Gundamaraju, R.; Sekaran, S.D.; Manikam, R. Major pathophysiological correlations of rosacea: A complete clinical appraisal. Int. J. Med. Sci. 2015, 12, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Öztas, M.; Balk, M.; Ögüs, E.; Bozkurt, M.; Ögüs, I.; Özer, N. The role of free oxygen radicals in the aetiopathogenesis of rosacea. Clin. Exp. Dermatol. 2003, 28, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Tisma, V.S.; Basta-Juzbasic, A.; Jaganjac, M.; Brcic, L.; Dobric, I.; Lipozencic, J.; Tatzber, F.; Zarkovic, N.; Poljak-Blazi, M. Oxidative stress and ferritin expression in the skin of patients with rosacea. J. Am. Acad. Dermatol. 2009, 60, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Endoplasmic reticulum stress: Key promoter of rosacea pathogenesis. Exp. Dermatol. 2014, 23, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Harijith, A.; Ebenezer, D.L.; Natarajan, V. Reactive oxygen species at the crossroads of inflammasome and inflammation. Front. Physiol. 2014, 5, 352. [Google Scholar] [CrossRef] [PubMed]
- Bonnar, E.; Eustace, P.; Powell, F.C. The Demodex mite population in rosacea. J. Am. Acad. Dermatol. 1993, 28, 443–448. [Google Scholar] [CrossRef]
- Forton, F.; Seys, B. Density of Demodex folliculorum in rosacea: A case-control study using standardized skin-surface biopsy. Br. J. Dermatol. 1993, 128, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Casas, C.; Paul, C.; Lahfa, M.; Livideanu, B.; Lejeune, O.; Alvarez-Georges, S.; Saint-Martory, C.; Degouy, A.; Mengeaud, V.; Ginisty, H.; et al. Quantification of Demodex folliculorum by PCR in rosacea and its relationship to skin innate immune activation. Exp. Dermatol. 2012, 21, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Turgut Erdemir, A.; Gurel, M.S.; Koku Aksu, A.E.; Falay, T.; Inan Yuksel, E.; Sarikaya, E. Demodex mites in acne rosacea: Reflectance confocal microscopic study. Australas. J. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Forton, F. Papulopustular rosacea, skin immunity and Demodex: Pityriasis folliculorum as a missing link. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Sattler, E.C.; Hoffmann, V.S.; Ruzicka, T.; Braunmuhl, T.V.; Berking, C. Reflectance confocal microscopy for monitoring the density of Demodex mites in patients with rosacea before and after treatment. Br. J. Dermatol. 2015, 173, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Koller, B.; Muller-Wiefel, A.S.; Rupec, R.; Korting, H.C.; Ruzicka, T. Chitin modulates innate immune responses of keratinocytes. PLoS ONE 2011, 6, e16594. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Sayama, K.; Tohyama, M.; Shirakata, Y.; Hanakawa, Y.; Tokumaru, S.; Yang, L.; Hirakawa, S.; Hashimoto, K. Mite allergen is a danger signal for the skin via activation of inflammasome in keratinocytes. J. Allergy Clin. Immunol. 2011, 127, 806–814.e1-4. [Google Scholar] [CrossRef] [PubMed]
- McMahon, F.; Banville, N.; Bergin, D.A.; Smedman, C.; Paulie, S.; Reeves, E.; Kavanagh, K. Activation of neutrophils via IP3 pathway following exposure to demodex-associated bacterial proteins. Inflammation 2016, 39, 425–433. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, N.; Gallagher, C.; Katikireddy, K.R.; Clynes, M.; O’Sullivan, F.; Kavanagh, K. Demodex-associated bacillus proteins induce an aberrant wound healing response in a corneal epithelial cell line: Possible implications for corneal ulcer formation in ocular rosaceabacillus proteins and corneal surface disruption. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3250–3259. [Google Scholar] [CrossRef] [PubMed]
- Akilov, O.E.; Mumcuoglu, K.Y. Immune response in demodicosis. J. Eur. Acad. Dermatol. Venereol. 2004, 18, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Delfos, N.M.; Collen, A.F.; Kroon, F.P. Demodex folliculitis: A skin manifestation of immune reconstitution disease. AIDS 2004, 18, 701–702. [Google Scholar] [CrossRef] [PubMed]
- AYRES, S.; Mihan, R. Rosacea-like demodicidosis involving the eyelids: A case report. Arch. Dermatol. 1967, 95, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Akdeniz, S.; Bahceci, M.; Tuzcu, A.; Harman, M.; Alp, S.; Bahceci, S. Is demodex folliculorum larger in diabetic patients? J. Eur. Acad. Dermatol. Venereol. 2002, 16, 539–541. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis—The “accidental” pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Whitfeld, M.; Gunasingam, N.; Leow, L.J.; Shirato, K.; Preda, V. Staphylococcus epidermidis: A possible role in the pustules of rosacea. J. Am. Acad. Dermatol. 2011, 64, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Dahl, M.V.; Ross, A.J.; Schlievert, P.M. Temperature regulates bacterial protein production: Possible role in rosacea. J. Am. Acad. Dermatol. 2004, 50, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Lacey, N.; Delaney, S.; Kavanagh, K.; Powell, F.C. Mite-related bacterial antigens stimulate inflammatory cells in rosacea. Br. J. Dermatol. 2007, 157, 474–481. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, N.; Menezes, N.; Kavanagh, K. Positive correlation between serum immunoreactivity to Demodex-associated Bacillus proteins and erythematotelangiectatic rosacea. Br. J. Dermatol. 2012, 167, 1032–1036. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; O’Reilly, N.; Sheha, H.; Katz, R.; Raju, V.K.; Kavanagh, K.; Tseng, S.C. Correlation between ocular Demodex infestation and serum immunoreactivity to Bacillus proteins in patients with Facial rosacea. Ophthalmology 2010, 117, 870–877.e1. [Google Scholar] [CrossRef] [PubMed]
- Gravina, A.; Federico, A.; Ruocco, E.; Schiavo, A.L.; Masarone, M.; Tuccillo, C.; Peccerillo, F.; Miranda, A.; Romano, L.; de Sio, C. Helicobacter pylori infection but not small intestinal bacterial overgrowth may play a pathogenic role in rosacea. United Eur. Gastroenterol. J. 2015, 3, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Tüzün, Y.; Keskin, S.; Kote, E. The role of Helicobacter pylori infection in skin diseases: Facts and controversies. Clin. Dermatol. 2010, 28, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.Y.; Chan, A.O.O.; Rashid, A.; Wong, D.K.H.; Cho, C.H.; Yuen, M.F. Helicobacter pylori induces promoter methylation of E-cadherin via interleukin-1β activation of nitric oxide production in gastric cancer cells. Cancer 2012, 118, 4969–4980. [Google Scholar] [CrossRef] [PubMed]
- Argenziano, G.; Donnarumma, G.; Arnese, P.; Assunta Baldassarre, M.; Baroni, A. Incidence of anti-Helicobacter pylori and anti-CagA antibodies in rosacea patients. Int. J. Dermatol. 2003, 42, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Ozkol, H.U.; Calka, O.; Akdeniz, N.; Baskan, E.; Ozkol, H. Rosacea and exposure to tandoor heat: Is there an association? Int. J. Dermatol. 2015, 54, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Sanchez, D.A.; Ishiuji, Y.; Patel, T.; Fountain, J.; Chan, Y.H.; Yosipovitch, G. Enhanced skin blood flow and sensitivity to noxious heat stimuli in papulopustular rosacea. J. Am. Acad. Dermatol. 2007, 57, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Minson, C.T.; Berry, L.T.; Joyner, M.J. Nitric oxide and neurally mediated regulation of skin blood flow during local heating. J. Appl. Physiol. 2001, 91, 1619–1626. [Google Scholar] [PubMed]
- Metzler-Wilson, K.; Kube, M.; Mann, S.; Demidova, O.; Sammons, D.L.; Wilson, T.E. Altered vascular endothelial control of facial skin blood flow in rosacea. FASEB J. 2016, 30, 733–736. [Google Scholar]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and expression of non-neuronal transient receptor potential (TRPV) ion channels in rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.D.; Sauder, C.L.; Ray, C.A. Mental stress elicits sustained and reproducible increases in skin sympathetic nerve activity. Physiol. Rep. 2013. [Google Scholar] [CrossRef] [PubMed]
- Black, P.H. Stress and the inflammatory response: A review of neurogenic inflammation. Brain Behav. Immun. 2002, 16, 622–653. [Google Scholar] [CrossRef]
- Wilson, T.E.; Toma, K.; Metzler-Wilson, K.; Sammons, D. Augmented supraorbital skin sympathetic nerve activity responses to symptom trigger events in rosacea patients. FASEB J. 2012, 26, 1092.10. [Google Scholar]
- Kim, J.E.; Cho, B.K.; Cho, D.H.; Park, H.J. Expression of hypothalamic-pituitary-adrenal axis in common skin diseases: Evidence of its association with stress-related disease activity. Acta Derm. Venereol. 2013, 93, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; Cruser, D.; Podawiltz, A.; Mummert, D.I.; Jones, H.; Mummert, M.E. Psychological stress and the cutaneous immune response: Roles of the HPA Axis and the sympathetic nervous system in atopic dermatitis and psoriasis. Dermatol. Res. Pract. 2012, 2012, 403908. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Pisarchik, A.; Zbytek, B.; Linton, E.A.; Mazurkiewicz, J.E.; Wei, E.T. Cutaneous expression of corticotropin-releasing hormone (CRH), urocortin, and CRH receptors. FASEB J. 2001, 15, 1678–1693. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.E.; Leung, D.Y. Epidermal barrier in atopic dermatitis. Allergy Asthma Immunol. Res. 2012, 4, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Lv, C.; Man, G.; Song, S.; Elias, P.M.; Man, M.Q. Abnormal epidermal barrier recovery in uninvolved skin supports the notion of an epidermal pathogenesis of psoriasis. J. Investig. Dermatol. 2014, 134, 2843–2846. [Google Scholar] [CrossRef] [PubMed]
- Dirschka, T.; Tronnier, H.; Folster-Holst, R. Epithelial barrier function and atopic diathesis in rosacea and perioral dermatitis. Br. J. Dermatol. 2004, 150, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Ni Raghallaigh, S.; Powell, F.C. Epidermal hydration levels in patients with rosacea improve after minocycline therapy. Br. J. Dermatol. 2014, 171, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Misery, L.; Loser, K.; Stander, S. Sensitive skin. J. Eur. Acad. Dermatol. Venereol. 2016, 30 (Suppl. 1), 2–8. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Kanada, K.; Macleod, D.T.; Borkowski, A.W.; Morizane, S.; Nakatsuji, T.; Cogen, A.L.; Gallo, R.L. TLR2 expression is increased in rosacea and stimulates enhanced serine protease production by keratinocytes. J. Investig. Dermatol. 2011, 131, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Ozlu, E.; Karadag, A.S.; Ozkanli, S.; Oguztuzun, S.; Kilic, M.; Zemheri, E.; Akbulak, O.; Akdeniz, N. Comparison of TLR-2, TLR-4, and antimicrobial peptide levels in different lesions of acne vulgaris. Cutan. Ocul. Toxicol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Hoffert, U.; Schröder, J.M. Epidermal proteases in the pathogenesis of rosacea. J. Investig. Dermatol. Symp. Proc. 2011, 15, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.A.; Buhren, B.A.; Steinhoff, M.; Homey, B. Rosacea: The cytokine and chemokine network. J. Investig. Dermatol. Symp. Proc. 2011, 15, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Segovia, J.; Sabbah, A.; Mgbemena, V.; Tsai, S.Y.; Chang, T.H.; Berton, M.T.; Morris, I.R.; Allen, I.C.; Ting, J.P.; Bose, S. TLR2/MyD88/NF-κB pathway, reactive oxygen species, potassium efflux activates NLRP3/ASC inflammasome during respiratory syncytial virus infection. PLoS ONE 2012, 7, e29695. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Schauber, J.; Coda, A.; Lin, H.; Dorschner, R.A.; Schechter, N.M.; Bonnart, C.; Descargues, P.; Hovnanian, A.; Gallo, R.L. Kallikrein-mediated proteolysis regulates the antimicrobial effects of cathelicidins in skin. FASEB J. 2006, 20, 2068–2080. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.H.; Sim, J.H.; Kang, H.Y.; Kim, Y.C.; Lee, E.S. Immunohistochemical expression of matrix metalloproteinases in the granulomatous rosacea compared with the non-granulomatous rosacea. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.; Grys, K.; Kollet, J.; Sarkany, R.; Young, A.R. Upregulation of MMP12 and its activity by UVA1 in human skin: Potential implications for photoaging. J. Investig. Dermatol. 2014, 134, 2598–2609. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, M.; Majewski, S. The role of antimicrobial peptides in chronic inflammatory skin diseases. Postep. Dermatol. Alergol. 2016, 33, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Cowland, J.B.; Johnsen, A.H.; Borregaard, N. hCAP-18, a cathelin/pro-bactenecin-like protein of human neutrophil specific granules. FEBS Lett. 1995, 368, 173–176. [Google Scholar] [CrossRef]
- Gudmundsson, G.H.; Agerberth, B.; Odeberg, J.; Bergman, T.; Olsson, B.; Salcedo, R. The human gene FALL39 and processing of the cathelin precursor to the antibacterial peptide LL-37 in granulocytes. Eur. J. Biochem. 1996, 238, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Salzer, S.; Ruzicka, T.; Schauber, J. Face-to-face with anti-inflammatory therapy for rosacea. Exp. Dermatol. 2014, 23, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Koczulla, R.; von Degenfeld, G.; Kupatt, C.; Krotz, F.; Zahler, S.; Gloe, T.; Issbrucker, K.; Unterberger, P.; Zaiou, M.; Lebherz, C.; et al. An angiogenic role for the human peptide antibiotic LL-37/hCAP-18. J. Clin. Investig. 2003, 111, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Schwab, V.D.; Sulk, M.; Seeliger, S.; Nowak, P.; Aubert, J.; Mess, C.; Rivier, M.; Carlavan, I.; Rossio, P.; Metze, D.; et al. Neurovascular and neuroimmune aspects in the pathophysiology of rosacea. J. Investig. Dermatol. Symp. Proc. 2011, 15, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, K.E.; Jung, H.Y.; Jo, H.; Jeong, S.W.; Lee, J.; Kim, C.H.; Kim, H.; Cho, D.; Park, H.J. Recombinant erythroid differentiation regulator 1 inhibits both inflammation and angiogenesis in a mouse model of rosacea. Exp. Dermatol. 2015, 24, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Gallo, R.L. Rosacea as a disease of cathelicidins and skin innate immunity. J. Investig. Dermatol. Symp. Proc. 2011, 15, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Niyonsaba, F.; Ushio, H.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Cathelicidin LL-37 induces the generation of reactive oxygen species and release of human α-defensins from neutrophils. Br. J. Dermatol. 2007, 157, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Bals, R.; Wilson, J.M. Cathelicidins—A family of multifunctional antimicrobial peptides. Cell. Mol. Life Sci. 2003, 60, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, A.; Vitiello, A.; Gallo, R.L. Cutting edge: Mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J. Immunol. 2003, 170, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
- Muto, Y.; Wang, Z.; Vanderberghe, M.; Two, A.; Gallo, R.L.; Di Nardo, A. Mast cells are key mediators of cathelicidin-initiated skin inflammation in rosacea. J. Investig. Dermatol. 2014, 134, 2728–2736. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, Y.J.; Lim, B.J.; Sohn, H.J.; Shin, D.; Oh, S.H. Increased expression of cathelicidin by direct activation of protease-activated receptor 2: Possible implications on the pathogenesis of rosacea. Yonsei Med. J. 2014, 55, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Fioranelli, M.; Roccia, M.G.; di Nardo, V.; Aracena, C.J.; Lotti, T. Vitamin D supplementation for childhood atopic dermatitis. Dermatol. Ther. 2016. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.R.; Jung, K.E.; Koo, D.W.; Lee, J.S. Vitamin D as a marker for disease severity in chronic urticaria and its possible role in pathogenesis. Ann. Dermatol. 2015, 27, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Ekiz, O.; Balta, I.; Sen, B.B.; Dikilitaş, M.C.; Ozuğuz, P.; Rifaioğlu, E.N. Vitamin D status in patients with rosacea. Cutan. Ocul. Toxicol. 2014, 33, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Hallau, J.; Hamann, L.; Schumann, R.R.; Worm, M.; Heine, G. A Promoter polymorphism of the Vitamin D metabolism gene Cyp24a1 is associated with severe atopic dermatitis in adults. Acta Derm. Venereol. 2016, 96, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Schauber, J.; Dorschner, R.A.; Yamasaki, K.; Brouha, B.; Gallo, R.L. Control of the innate epithelial antimicrobial response is cell-type specific and dependent on relevant microenvironmental stimuli. Immunology 2006, 118, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Jansen, T.; Krug, S.; Kind, P.; Plewig, G.; Messer, G. BsmI polymorphism of the vitamin D receptor gene in patients with the fulminant course of rosacea conglobata (rosacea fulminans). J. Dermatol. 2004, 31, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Morizane, S.; Yamasaki, K.; Kabigting, F.D.; Gallo, R.L. Kallikrein expression and cathelicidin processing are independently controlled in keratinocytes by calcium, vitamin D3, and retinoic acid. J. Investig. Dermatol. 2010, 130, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Sakabe, J.; Umayahara, T.; Hiroike, M.; Shimauchi, T.; Ito, T.; Tokura, Y. Calcipotriol increases hCAP18 mRNA expression but inhibits extracellular LL37 peptide production in IL-17/IL-22-stimulated normal human epidermal keratinocytes. Acta Derm. Venereol. 2014, 94, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.T.; Choi, E.-Y.K.; Thomas, D.G.; Hristov, A.C.; Chan, M.P. Comparative analysis of rosacea and cutaneous lupus erythematosus: Histopathologic features, T-cell subsets, and plasmacytoid dendritic cells. J. Am. Acad. Dermatol. 2014, 71, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.B.; Kim, H.J.; Jung, H.Y.; Park, Y.G.; Kim, S.Y.; Cho, B.K.; Cho, D.; Park, H.J. Downregulation of erythroid differentiation regulator 1 as a novel marker of skin tumors. Int. J. Dermatol. 2014, 53, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.K.; Park, Y.; Song, S.B.; Cheon, S.Y.; Park, S.; Houh, Y.; Ha, S.; Kim, H.J.; Park, J.M.; Kim, T.S.; et al. Erythroid differentiation regulator 1, an interleukin 18-regulated gene, acts as a metastasis suppressor in melanoma. J. Investig. Dermatol. 2011, 131, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Woźniacka, A.; Salamon, M.; McCauliffe, D.; Sysa-Jędrzejowska, A. Antinuclear antibodies in rosacea patients. Postep. Dermatol. Alergol. 2013, 30, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Yoo, J.Y.; Kim, M.J.; Hwang, S.G.; Ahn, K.C.; Ryu, J.C.; Choi, M.K.; Joo, J.H.; Kim, C.H.; Lee, S.N.; et al. Distinct TLR-mediated pathways regulate house dust mite-induced allergic disease in the upper and lower airways. J. Allergy Clin. Immunol. 2013, 131, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Pasare, C.; Medzhitov, R. Control of B-cell responses by Toll-like receptors. Nature 2005, 438, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Addor, F.A. Skin barrier in rosacea. An. Bras. Dermatol. 2016, 91, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.H.; Yaar, M.; Eyada, M.M.; Bhawan, J. Lymphangiogenesis and angiogenesis in non-phymatous rosacea. J. Cutan. Pathol. 2007, 34, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, S.; Buddenkotte, J.; Schmidt-Choudhury, A.; Rosignoli, C.; Shpacovitch, V.; von Arnim, U.; Metze, D.; Rukwied, R.; Schmelz, M.; Paus, R.; et al. Pituitary adenylate cyclase activating polypeptide: An important vascular regulator in human skin in vivo. Am. J. Pathol. 2010, 177, 2563–2575. [Google Scholar] [CrossRef] [PubMed]
- Greeno, E.W.; Mantyh, P.; Vercellotti, G.M.; Moldow, C.F. Functional neurokinin 1 receptors for substance P are expressed by human vascular endothelium. J. Exp. Med. 1993, 177, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal control of skin function: The skin as a neuroimmunoendocrine organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef] [PubMed]
- Norrby, K. Mast cells and angiogenesis. APMIS 2002, 110, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, O.; Braun, J.S.; Becker, D.; Halle, A.; Freyer, D.; Dagand, E.; Lehnardt, S.; Weber, J.R. TLR2 mediates neuroinflammation and neuronal damage. J. Immunol. 2007, 178, 6476–6481. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Luo, L.; Pflugfelder, S.C.; Li, D.Q. Doxycycline inhibits TGF-β1-induced MMP-9 via Smad and MAPK pathways in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Golub, L.M.; Lee, H.M.; Ryan, M.E.; Giannobile, W.V.; Payne, J.; Sorsa, T. Tetracyclines inhibit connective tissue breakdown by multiple non-antimicrobial mechanisms. Adv. Dent. Res. 1998, 12, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, A.; Holmes, A.D.; Muto, Y.; Huang, E.Y.; Preston, N.; Winkelman, W.J.; Gallo, R.L. Improved clinical outcome and biomarkers in adults with papulopustular rosacea treated with doxycycline modified-release capsules in a randomized trial. J. Am. Acad. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, P.; Arcoleo, F.; Barbera, C.; di Bella, G.; La Rosa, M.; Misiano, G.; Milano, S.; Brai, M.; Cammarata, G.; Feo, S.; et al. Tetracycline inhibits the nitric oxide synthase activity induced by endotoxin in cultured murine macrophages. Eur. J. Pharmacol. 1998, 346, 283–290. [Google Scholar] [CrossRef]
- Del Rosso, J.Q.; Webster, G.F.; Jackson, M.; Rendon, M.; Rich, P.; Torok, H.; Bradshaw, M. Two randomized phase III clinical trials evaluating anti-inflammatory dose doxycycline (40-mg doxycycline, USP capsules) administered once daily for treatment of rosacea. J. Am. Acad. Dermatol. 2007, 56, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.D. Submicrobial doxycycline and rosacea. Compr. Ther. 2007, 33, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Hünerbein, A.; Getie, M.; Jackel, A.; Neubert, R.H. Scavenging properties of metronidazole on free oxygen radicals in a skin lipid model system. J. Pharm. Pharmacol. 2007, 59, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Cardwell, L.A.; Alinia, H.; Moradi Tuchayi, S.; Feldman, S.R. New developments in the treatment of rosacea—Role of once-daily ivermectin cream. Clin. Cosmet. Investig. Dermatol. 2016, 9, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Coda, A.B.; Hata, T.; Miller, J.; Audish, D.; Kotol, P.; Two, A.; Shafiq, F.; Yamasaki, K.; Harper, J.C.; del Rosso, J.Q.; et al. Cathelicidin, kallikrein 5, and serine protease activity is inhibited during treatment of rosacea with azelaic acid 15% gel. J. Am. Acad. Dermatol. 2013, 69, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Elewski, B.; Draelos, Z.; Dreno, B.; Jansen, T.; Layton, A.; Picardo, M. Rosacea–global diversity and optimized outcome: proposed international consensus from the Rosacea International Expert Group. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Van Zuuren, E.J.; Fedorowicz, Z. Low-Dose isotretinoin: An option for difficult-to-treat papulopustular rosacea. J. Investig. Dermatol. 2016, 136, 1081–1083. [Google Scholar] [CrossRef] [PubMed]
- Dispenza, M.C.; Wolpert, E.B.; Gilliland, K.L.; Dai, J.P.; Cong, Z.; Nelson, A.M.; Thiboutot, D.M. Systemic isotretinoin therapy normalizes exaggerated TLR-2-mediated innate immune responses in acne patients. J. Investig. Dermatol. 2012, 132, 2198–2205. [Google Scholar] [CrossRef] [PubMed]
- Del Rosso, J.Q. Evaluating the role of topical therapies in the management of rosacea: Focus on combination sodium sulfacetamide and sulfur formulations. Cutis 2004, 73, 29–33. [Google Scholar] [PubMed]
- Weinkle, A.P.; Doktor, V.; Emer, J. Update on the management of rosacea. Clin. Cosmet. Investig. Dermatol. 2015, 8, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.; Kircik, L.; Fowler, J.; Tan, J.; Draelos, Z.; Fleischer, A.; Appell, M.; Steinhoff, M.; Lynde, C.; Liu, H. Efficacy and safety of ivermectin 1% cream in treatment of papulopustular rosacea: Results of two randomized, double-blind, vehicle-controlled pivotal studies. J. Drugs Dermatol. 2014, 13, 316–323. [Google Scholar] [PubMed]
- Two, A.M.; Wu, W.; Gallo, R.L.; Hata, T.R. Rosacea: Part II. Topical and systemic therapies in the treatment of rosacea. J. Am. Acad. Dermatol. 2015, 72, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Two, A.M.; Hata, T.R.; Nakatsuji, T.; Coda, A.B.; Kotol, P.F.; Wu, W.; Shafiq, F.; Huang, E.Y.; Gallo, R.L. Reduction in serine protease activity correlates with improved rosacea severity in a small, randomized pilot study of a topical serine protease inhibitor. J. Investig. Dermatol. 2014, 134, 1143–1145. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.A.; Weiss, M.A.; Beasley, K.L. Rejuvenation of photoaged skin: 5 Years results with intense pulsed light of the face, neck, and chest. Dermatol. Surg. 2002, 28, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Bae, S.H.; Moon, K.R.; Na, E.Y.; Yun, S.J.; Lee, S.C. Light emitting diodes down-regulates cathelicidin, kallikrein, and Toll-like receptor 2 expressions in keratinocytes and rosacea-like mouse skin. Exp. Dermatol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, M.; Bae, J.M.; Cho, B.K.; Park, H.J. Efficacy of the long-pulsed 1064-nm neodymium:yttrium-aluminum-garnet laser (LPND) (rejuvenation mode) in the treatment of papulopustular rosacea (PPR): A pilot study of clinical outcomes and patient satisfaction in 30 cases. J. Am. Acad. Dermatol. 2015, 73, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.B.; Kang, N.H.; Eun, Y.S.; Cheon, M.S.; Kim, K.M.; Cho, B.K.; Park, H.J. Effects of long-pulsed 1064-nm neodymium-doped yttrium aluminum garnet laser on dermal collagen remodeling in hairless mice. Dermatol. Surg. 2012, 38, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Wollina, U. Recent advances in the understanding and management of rosacea. F1000Prime Rep. 2014, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Cookson, H.; McFadden, J.; White, J.; White, I.R. Allergic contact dermatitis caused by Mirvaso(R), brimonidine tartrate gel 0.33%, a new topical treatment for rosaceal erythema. Contact Dermat. 2015, 73, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Lowe, E.; Lim, S. Paradoxical erythema reaction of long-term topical brimonidine gel for the treatment of facial erythema of rosacea. J. Drugs Dermatol. 2016, 15, 763–765. [Google Scholar] [PubMed]
- Hsu, C.C.; Lee, J.Y. Carvedilol for the treatment of refractory facial flushing and persistent erythema of rosacea. Arch. Dermatol. 2011, 147, 1258–1260. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.J.; Behr, K.L.; Fitzpatrick, R.; Goldman, M.; Ruiz-Esparza, J. Flash lamp pumped dye laser for rosacea-associated telangiectasia and erythema. J. Dermatol. Surg. Oncol. 1991, 17, 522–525. [Google Scholar] [PubMed]
- Clark, S.M.; Lanigan, S.W.; Marks, R. Laser treatment of erythema and telangiectasia associated with rosacea. Lasers Med. Sci. 2002, 17, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Lonne-Rahm, S.; Nordlind, K.; Edstrom, D.W.; Ros, A.M.; Berg, M. Laser treatment of rosacea: A pathoetiological study. Arch. Dermatol. 2004, 140, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, Y.R.; Lim, J.H.; Cho, D.H.; Park, H.J. Rosacea: Molecular Mechanisms and Management of a Chronic Cutaneous Inflammatory Condition. Int. J. Mol. Sci. 2016, 17, 1562. https://doi.org/10.3390/ijms17091562
Woo YR, Lim JH, Cho DH, Park HJ. Rosacea: Molecular Mechanisms and Management of a Chronic Cutaneous Inflammatory Condition. International Journal of Molecular Sciences. 2016; 17(9):1562. https://doi.org/10.3390/ijms17091562
Chicago/Turabian StyleWoo, Yu Ri, Ji Hong Lim, Dae Ho Cho, and Hyun Jeong Park. 2016. "Rosacea: Molecular Mechanisms and Management of a Chronic Cutaneous Inflammatory Condition" International Journal of Molecular Sciences 17, no. 9: 1562. https://doi.org/10.3390/ijms17091562
APA StyleWoo, Y. R., Lim, J. H., Cho, D. H., & Park, H. J. (2016). Rosacea: Molecular Mechanisms and Management of a Chronic Cutaneous Inflammatory Condition. International Journal of Molecular Sciences, 17(9), 1562. https://doi.org/10.3390/ijms17091562