
Acquisition of Flavescence Dorée Phytoplasma by Scaphoideus titanus Ball from Different Grapevine Varieties


 ,
,  and
and 
Abstract
:1. Introduction
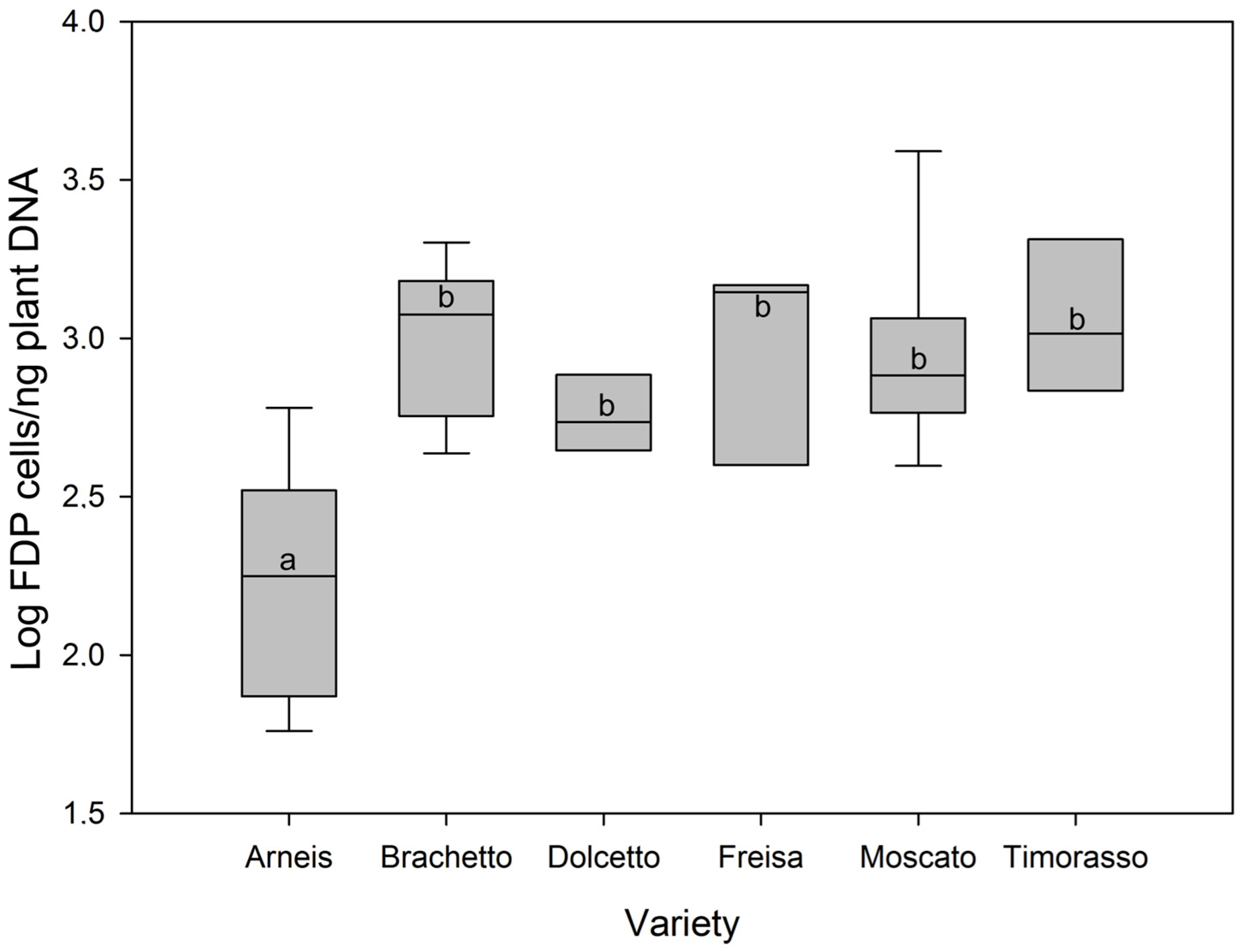
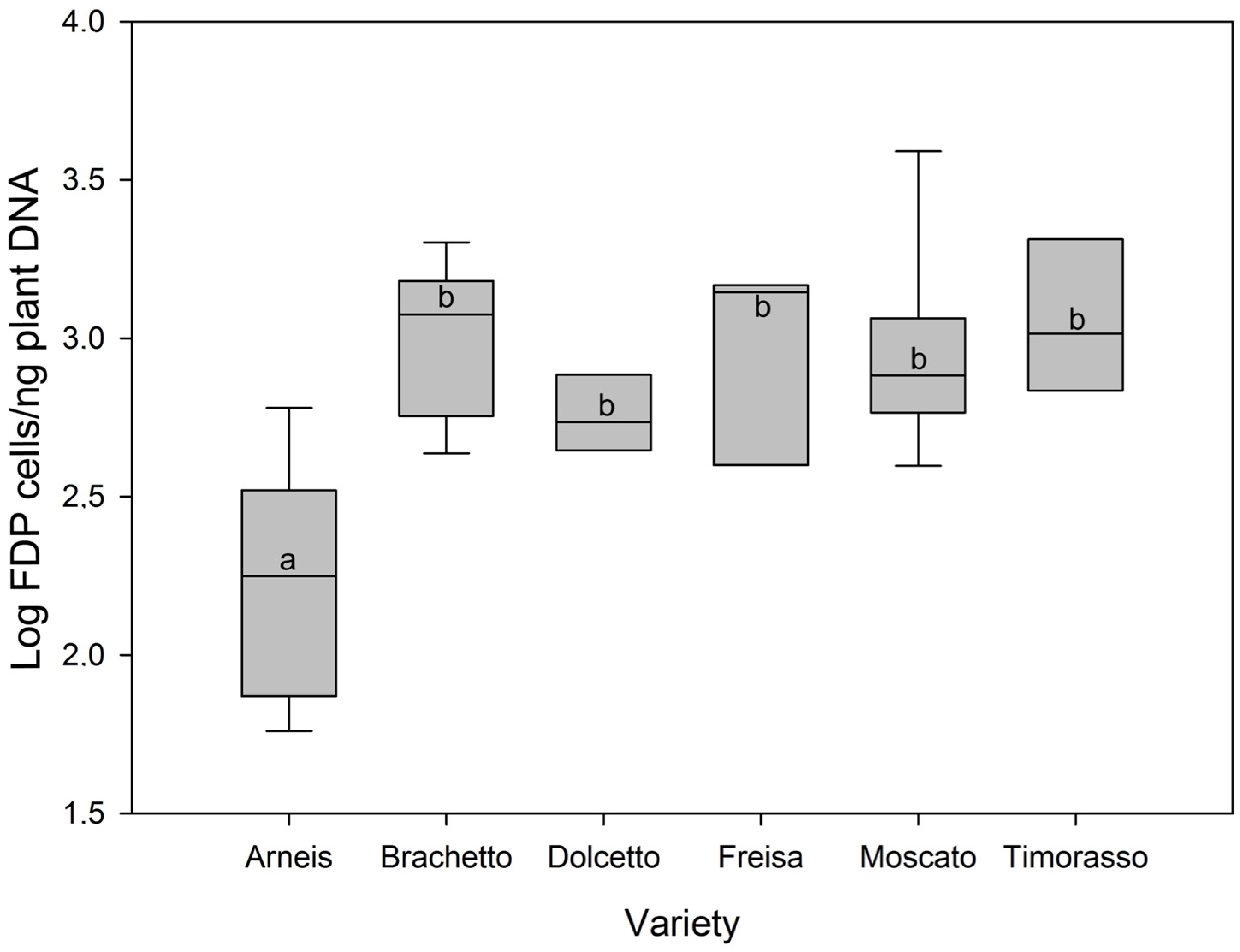
2. Results
3. Discussion
4. Materials and Methods
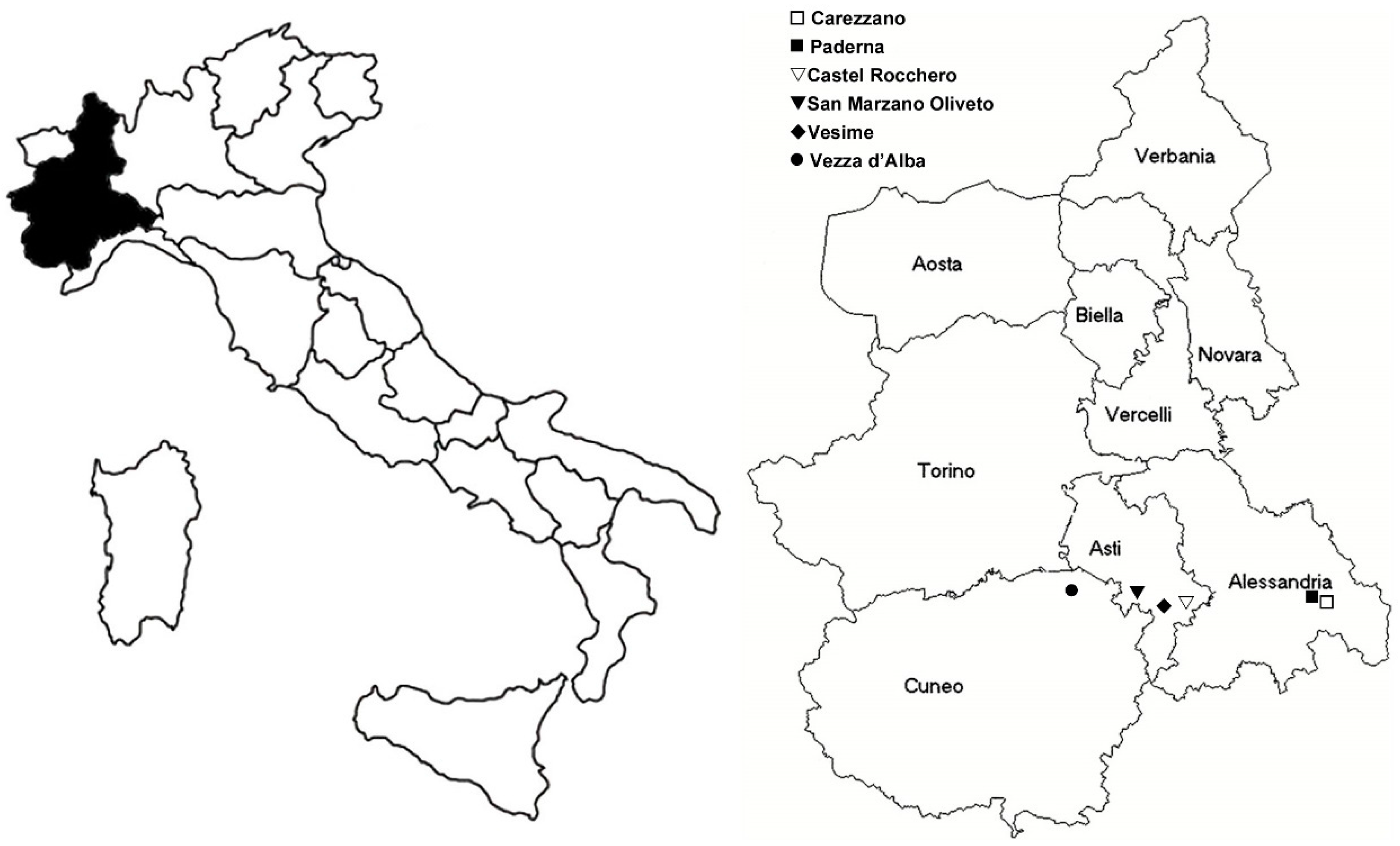
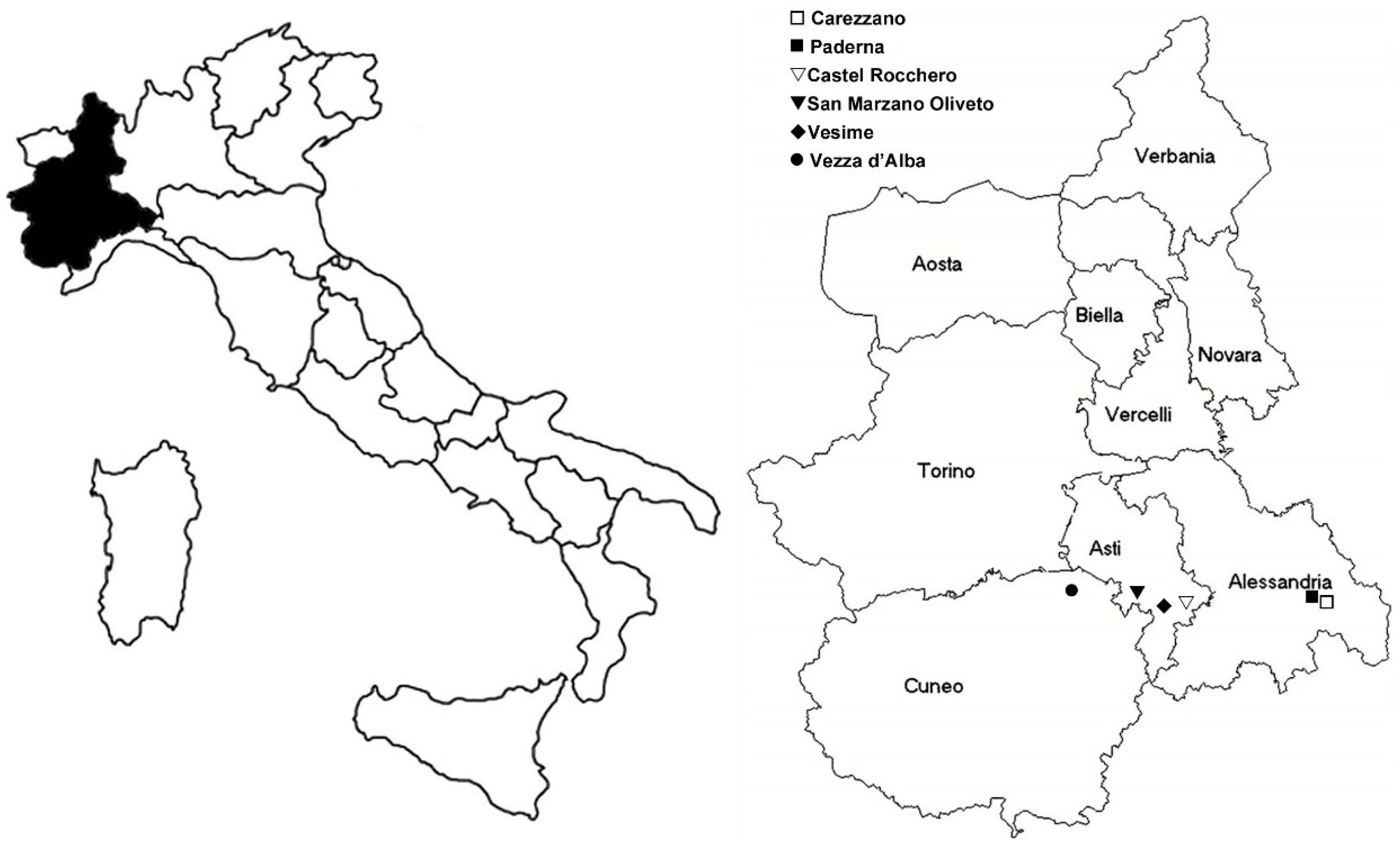
4.1. Vector and Flavescence Dorée Phytoplasma Source Plants
4.2. Flavescence Dorée Phytoplasma Acquisition Experiments
4.3. DNA Extraction and Phytoplasma Detection by PCR
4.4. Flavescence Dorée Phytoplasma Quantification by qPCR
4.5. Data Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Belli, G.; Bianco, P.A.; Conti, M. Grapevine yellows in Italy: Past, present and future. J. Plant Pathol. 2010, 92, 303–326. [Google Scholar]
- Marzachi, C.; Alma, A.; D’Aquilio, M.; Minuto, G.; Boccardo, G. Detection and identification of Phytoplasmas infecting cultivated and wild plants in Liguria (Italian Riviera). J. Plant Pathol. 1999, 81, 127–136. [Google Scholar]
- Marzachì, C.; Boarino, A.; Vischi, A.; Palermo, S.; Morone, C.; Loria, A.; Boccardo, G. Flavescenza dorata, legno nero e giallume dell’astro in vitigni del Piemonte sud orientale. Inf. Fitopatol. 2001, 9, 58–63. [Google Scholar]
- Unesco World Heritage Centre. Vineyard Landscape of Piedmont: Langhe-roero and Monferrato. Available online: http://whc.Unesco.Org/en/list/1390 (accessed on 22 August 2016).
- Bressan, A.; Spiazzi, S.; Girolami, V.; Boudon-Padieu, E. Acquisition efficiency of Flavescence dorée phytoplasma by Scaphoideus titanus Ball from infected tolerant or susceptible grapevine cultivars or experimental host plants. VITIS-J. Grapevine Res. 2005, 44, 143. [Google Scholar]
- Galetto, L.; Miliordos, D.; Roggia, C.; Rashidi, M.; Sacco, D.; Marzachi, C.; Bosco, D. Acquisition capability of the grapevine Flavescence doree by the leafhopper vector Scaphoideus titanus Ball correlates with phytoplasma titre in the source plant. J. Pest Sci. 2014, 87, 671–679. [Google Scholar] [CrossRef]
- Marcone, C. Molecular biology and pathogenicity of phytoplasmas. Ann. Appl. Biol. 2014, 165, 199–221. [Google Scholar] [CrossRef]
- Foissac, X.; Wilson, M.R. Current and possible future distributions of phytoplasma diseases and their vectors. In Phytoplasmas: Genomes, Plant Hosts and Vectors; Weintraub, P.G., Jones, P., Eds.; CABI: Wallingford, UK, 2010; pp. 309–324. [Google Scholar]
- Schvester, D.; Carle, P.; Montous, G. Transmission de la Flavescence dorée de la vigne par S. littoralis Ball. Ann. Epiphyt. 1963, 14, 175–198. [Google Scholar]
- Caudwell, A.; Kuszala, C.; Larrue, J.; Bachelier, J.C. Transmission de la Flavescence dorée de la fève à la fève par des cicadelles des genres Euscelis et Euscelidius. Ann. Phytopathol. 1972, 181–189. [Google Scholar]
- Rashidi, M.; D’Amelio, R.; Galetto, L.; Marzachi, C.; Bosco, D. Interactive transmission of two phytoplasmas by the vector insect. Ann. Appl. Biol. 2014, 165, 404–413. [Google Scholar] [CrossRef]
- Salar, P.; Charenton, C.; Foissac, X.; Malembic-Maher, S. Multiplication kinetics of Flavescence dorée phytoplasma in broad bean. Effect of phytoplasma strain and temperature. Eur. J. Plant Pathol. 2013, 135, 371–381. [Google Scholar] [CrossRef]
- De Jong, Y. Fauna Europaea Version 2.6. 2013. Available online: http//www.faunaeur.org (accessed on 23 August 2016).
- Filippin, L.; Jovic, J.; Cvrkovic, T.; Forte, V.; Clair, D.; Tosevski, I.; Boudon-Padieu, E.; Borgo, M.; Angelini, E. Molecular characteristics of phytoplasmas associated with flavescence doree in clematis and grapevine and preliminary results on the role of dictyophara europaea as a vector. Plant Pathol. 2009, 58, 826–837. [Google Scholar] [CrossRef]
- Lessio, F.; Alma, A. Host plants and seasonal presence of dictyophara europea in the vineyard agro-ecosystem. Bull. Insectol. 2008, 61, 199–200. [Google Scholar]
- Gaffuri, F.; Sacchi, S.; Cavagna, B. First detection of the mosaic leafhopper, orientus ishidae, in northern italian vineyards infected by the flavescence dorée phytoplasma. New Disease Rep. 2011, 24. [Google Scholar] [CrossRef]
- Mehle, N.; Seljak, G.; Rupar, M.; Ravnikar, M.; Dermastia, M. The first detection of a phytoplasma from the 16Srv (elm yellows) group in the mosaic leafhopper Orientus ishidae. New Dis. Rep. 2010, 22. [Google Scholar] [CrossRef]
- Trivellone, V.; Filippin, L.; Narduzzi-Wicht, B.; Angelini, E. A regional-scale survey to define the known and potential vectors of grapevine yellow phytoplasmas in vineyards south of swiss alps. Eur. J. Plant Pathol. 2016, 145, 915–927. [Google Scholar] [CrossRef]
- Bosio, G.; Rossi, A. Ciclo biologico in Piemonte di Scaphoideus titanus. L’Inf. Agrar. 2001, 21, 75–78. [Google Scholar]
- Chuche, J.; Thiery, D. Biology and ecology of the Flavescence doree vector Scaphoideus titanus: A review. Agron. Sustain. Dev. 2014, 34, 381–403. [Google Scholar] [CrossRef]
- Prezelj, N.; Nikolic, P.; Gruden, K.; Ravnikar, M.; Dermastia, M. Spatiotemporal distribution of flavescence doree phytoplasma in grapevine. Plant Pathol. 2013, 62, 760–766. [Google Scholar] [CrossRef]
- Roggia, C.; Caciagli, P.; Galetto, L.; Pacifico, D.; Veratti, F.; Bosco, D.; Marzachi, C. Flavescence dorée phytoplasma titre in field-infected Barbera and Nebbiolo grapevines. Plant Pathol. 2014, 63, 31–41. [Google Scholar] [CrossRef]
- Morone, C.; Gotta, P.; Marzachì, C. Riconoscimento dei sintomi di inizio stagione della flavescenza dorata. L’Inf. Agrar. 2001, 57, 83–86. [Google Scholar]
- Caudwell, A. Epidemiology and characterization of Flavescence dorée (FD) and other grapevine yellows. Agronomie 1990, 10, 655–663. [Google Scholar] [CrossRef]
- Margaria, P.; Palmano, S. Response of the Vitis vinifera L. cv. “Nebbiolo” proteome to Flavescence dorée phytoplasma infection. Proteomics 2011, 11, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Morone, C.; Boveri, M.; Giosué, S.; Gotta, P.; Rossi, V.; Scapin, I.; Marzachì, C. Epidemiology of Flavescence dorée in vineyards in northwestern Italy. Phytopathology 2007, 97, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, M.; Faes, G.; Segala, C.; Stefanini, M.; Salakhutdinov, I.; Zyprian, E.; Toepfer, R.; Grando, M.S.; Velasco, R. Genome diversity and gene haplotypes in the grapevine (Vitis vinifera L.), as revealed by single nucleotide polymorphisms. Mol. Breed. 2004, 14, 385–395. [Google Scholar] [CrossRef]
- Bellomo, C.; Carraro, L.; Ermacora, P.; Pavan, F.; Osler, R.; Frausin, C.; Governatori, G. Recovery phenomena in grapevines affected by grapevine yellows in Friuli Venezia Giulia. Bull. Insectol. 2007, 60, 235–236. [Google Scholar]
- Garau, R.; Tolu, G.; Prota, V.A.; Sechi, A. Differential reactivity of grapevine cultivars to “Bois noir” infections in Sardinia. J. Plant Pathol. 2004, 86, 320. [Google Scholar]
- Osler, R.; Carraro, L.; Ermacora, P.; Ferrini, F.; Loi, N.; Loschi, A.; Martini, M.; Mutton, P.B.; Refatti, R. Roguing: A controversial practice to eradicate grape yellows caused by phytoplasmas. In Proceedings of the 14th Meeting of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine (ICVG), Locorotondo, Italy, 12–17 September 2003; p. 68.
- Speich, P.; Méjean, I.; Noyer, C.; Thomas, C.; Gillet, J.; Cloquemin, G.; Clair, D.; Boudon-Padieu, E. Limited susceptibility of Syrah cv. to Flavescence dorée and Bois noir in South of France. In Proceedings of the 15th Meeting ICVG, Stellenbosch, South Africa, 3–7 April 2006; pp. 171–172.
- Jagoueix-Eveillard, S.; Labroussaa, F.; Salar, P.; Danet, J.L.; Hevin, C.; Perrin, M.; Masson, J.; Foissac, X.; Malembic-Maher, S. Looking for resistance to the Flavescence dorée disease among Vitis vinifera cultivars and other Vitis species. In Proceedings of the 17th Congress of ICVG, Davis, CA, USA, 7–14 October 2012; pp. 234–235.
- Purcell, A.H. Insect vector relationships with prokaryotic plant pathogens. Annu. Rev. Phytopathol. 1982, 20, 397–417. [Google Scholar] [CrossRef]
- Chuche, J.; Boudon-Padieu, E.; Thiéry, D. Host preferences of the leafhopper Scaphoideus titanus, vector of “flavescence dorée” phytoplasma. Phytopathol. Mollicutes 2016, 6, 38–45. [Google Scholar] [CrossRef]
- Pavan, F.; Mori, N.; Bigot, G.; Zandigiacomo, P. Border effect in spatial distribution of Flavescence dorée affected grapevines and outside source of Scaphoideus titanus vectors. Bull. Insectol. 2012, 65, 281–290. [Google Scholar]
- Pegoraro, M.; Rossi, M.; Marzachi, C.; Bosco, D. Scaphoideus titanus Ball e tracciatura delle vie di diffusione della Flavescenza dorata in vigneto. In Proceedings of the XXV Congresso nazionale italiano di entomologia, Padova, Italy, 20–24 June 2016.
- Malembic-Maher, S.; Salar, P.; Filippin, L.; Carle, P.; Angelini, E.; Foissac, X. Genetic diversity of European phytoplasmas of the 16SrV taxonomic group and proposal of “Candidatus Phytoplasma rubi”. Int. J. Syst. Evol. Microbiol. 2011, 61, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.; Murari, E.; Mori, N.; Bertaccini, A. Identification and epidemic distribution of two flavescence dorée-related phytoplasmas in Veneto (Italy). Plant Dis. 1999, 83, 925–930. [Google Scholar] [CrossRef]
- Galetto, L.; Marzachi, C.; Marques, R.; Graziano, C.; Bosco, D. Effects of temperature and CO2 on phytoplasma multiplication pattern in vector and plant. Bull. Insectol. 2011, 64, S151–S152. [Google Scholar]
- Mori, N.; Bressan, A.; Martini, M.; Guadagnini, M.; Girolami, V.; Bertaccini, A. Experimental transmission by Scaphoideus titanus Ball of two Flavescence dorée-type phytoplasmas. Vitis 2002, 41, 99–102. [Google Scholar]
- Bertin, S.; Guglielmino, C.R.; Karam, N.; Gomulski, L.M.; Malacrida, A.R.; Gasperi, G. Diffusion of the Nearctic leafhopper Scaphoideus titanus Ball in Europe: A consequence of human trading activity. Genetica 2007, 131, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Papura, D.; Burban, C.; van Helden, M.; Giresse, X.; Nusillard, B.; Guillemaud, T.; Kerdelhué, C. Microsatellite and mitochondrial data provide evidence for a single major introduction for the Neartic leafhopper Scaphoideus titanus in Europe. PLoS ONE 2012, 7, e36882. [Google Scholar] [CrossRef] [PubMed]
- Papura, D.; Delmotte, F.; Giresse, X.; Salar, P.; Danet, J.L.; Van Helden, M.; Foissac, X.; Malembic-Maher, S. Comparing the spatial genetic structures of the Flavescence doree phytoplasma and its leafhopper vector Scaphoideus titanus. Infect. Genet. Evol. 2009, 9, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Abba, S.; Palmano, S. Novel aspects of grapevine response to phytoplasma infection investigated by a proteomic and phospho-proteomic approach with data integration into functional networks. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Ferrandino, A.; Caciagli, P.; Kedrina, O.; Schubert, A.; Palmano, S. Metabolic and transcript analysis of the flavonoid pathway in diseased and recovered Nebbiolo and Barbera grapevines (Vitis vinifera L.) following infection by Flavescence dorée phytoplasma. Plant Cell Environ. 2014, 37, 2183–2200. [Google Scholar] [CrossRef] [PubMed]
- Armijo, G.; Schlechter, R.; Agurto, M.; Munoz, D.; Nunez, C.; Arce-Johnosn, P. Grapevine pathogenic microrganisms: Understanding infection strategies and host response scenarios. Front. Plant Sci. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Caudwell, A.; Kuszala, C.; Bachelier, J.C.; Larrue, J. Transmission de la flavescence dorée de la vigne aux plantes herbacées par l’allongement du temps d’utilisation de la cicadelle scaphoideus littoralis ball et l’étude de sa survie sur un grand nombre d’espèces végétales. Ann. Phytopathol. 1970, 2, 415–428. [Google Scholar]
- Galetto, L.; Bosco, D.; Marzachi, C. Universal and group-specific real-time PCR diagnosis of flavescence dorée (16Sr-V), bois noir (16Sr-XII) and apple proliferation (16Sr-X) phytoplasmas from field-collected plant hosts and insect vectors. Ann. Appl. Biol. 2005, 147, 191–201. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenetic classification of plant pathogenic mycoplasma-like organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, R., Tully, J.G., Eds.; Academic Press: San Diego, CA, USA, 1995; Volume I, pp. 369–380. [Google Scholar]
- Lee, I.M.; Gundersen, D.E.; Hammond, R.W.; Davis, R.E. Use of mycoplasmalike organism (MLO) group-specific oligonucleotide primers for nested-PCR assays to detect mixed-MLO infections in a single host-plant. Phytopathology 1994, 84, 559–566. [Google Scholar] [CrossRef]
- Marzachì, C.; Bosco, D. Relative quantification of chrysanthemum yellows (16SrI) phytoplasma in its plant and insect host using real-time polymerase chain reaction. Mol. Biotechnol. 2005, 30, 117–127. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Variety | Number of Plants Used for FDP Quantification | FDP Load * | Number of Plants Used for Acquisition | PCR Positive/Total Tested Insects |
|---|---|---|---|---|
| Arneis | 12 | 230.50 ± 55.88 a | 3 | 45/132 |
| Brachetto | 9 | 1136.19 ± 177.37 b | 2 | 16/41 |
| Dolcetto | 7 | 677.17 ± 171.54 b | 2 | 30/62 |
| Freisa | 3 | 1090.00 ± 346.58 b | 3 | 37/89 |
| Moscato | 12 | 1216.81 ± 386.59 b | 5 | 16/178 |
| Timorasso | 4 | 1294.70 ± 378.72 b | 4 | 22/101 |
| Source | Type III | ||
|---|---|---|---|
| Wald Chi-Square | df | Sign. | |
| (Intercept) | 27.892 | 1 | 0.000 |
| Variety | 26.796 | 5 | 0.000 |
| FDP | 13.788 | 1 | 0.000 |
| Variety× FDP | 19.628 | 5 | 0.001 |
| Parameter | Parameter Estimation | 95% Wald Confidence Interval | Significance | |
|---|---|---|---|---|
| Upper | Lower | |||
| Intercept | 0.432 | −0.544 | 1.408 | 0.385 |
| Arneis | −3.590 | −5.377 | −1.802 | 0.000 |
| Brachetto | −2.801 | −6.339 | 0.738 | 0.121 |
| Freisa | −2.023 | −3.565 | −0.482 | 0.010 |
| Moscato | −2.457 | −3.992 | −0.923 | 0.002 |
| Timorasso | −3.538 | −5.148 | −1.929 | 0.000 |
| Dolcetto | 0 a | – | – | – |
| FDP | −0.001 | −0.003 | 0.001 | 0.248 |
| ARNEIS × FDP | 0.023 | 0.010 | 0.035 | 0.000 |
| BRACHETTO × FDP | 0.003 | −0.001 | 0.007 | 0.109 |
| FREISA × FDP | 0.002 | 6.342 × 10−5 | 0.005 | 0.044 |
| MOSCATO × FDP | 0.001 | −0.002 | 0.003 | 0.554 |
| TIMORASSO × FDP | 0.002 | 0.000 | 0.005 | 0.024 |
| DOLCETTO × FDP | 0 a | – | – | – |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galetto, L.; Miliordos, D.E.; Pegoraro, M.; Sacco, D.; Veratti, F.; Marzachì, C.; Bosco, D. Acquisition of Flavescence Dorée Phytoplasma by Scaphoideus titanus Ball from Different Grapevine Varieties. Int. J. Mol. Sci. 2016, 17, 1563. https://doi.org/10.3390/ijms17091563
Galetto L, Miliordos DE, Pegoraro M, Sacco D, Veratti F, Marzachì C, Bosco D. Acquisition of Flavescence Dorée Phytoplasma by Scaphoideus titanus Ball from Different Grapevine Varieties. International Journal of Molecular Sciences. 2016; 17(9):1563. https://doi.org/10.3390/ijms17091563
Chicago/Turabian StyleGaletto, Luciana, Dimitrios E. Miliordos, Mattia Pegoraro, Dario Sacco, Flavio Veratti, Cristina Marzachì, and Domenico Bosco. 2016. "Acquisition of Flavescence Dorée Phytoplasma by Scaphoideus titanus Ball from Different Grapevine Varieties" International Journal of Molecular Sciences 17, no. 9: 1563. https://doi.org/10.3390/ijms17091563
APA StyleGaletto, L., Miliordos, D. E., Pegoraro, M., Sacco, D., Veratti, F., Marzachì, C., & Bosco, D. (2016). Acquisition of Flavescence Dorée Phytoplasma by Scaphoideus titanus Ball from Different Grapevine Varieties. International Journal of Molecular Sciences, 17(9), 1563. https://doi.org/10.3390/ijms17091563