
Evaluation of Anthelmintic Activity and Composition of Pumpkin (Cucurbita pepo L.) Seed Extracts—In Vitro and in Vivo Studies

 ,
,  ,
,  ,
, 
Abstract
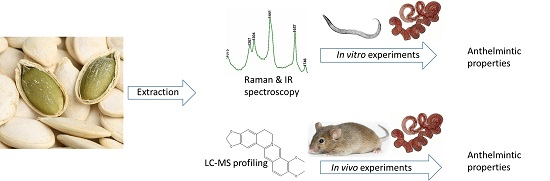
:
1. Introduction
2. Results
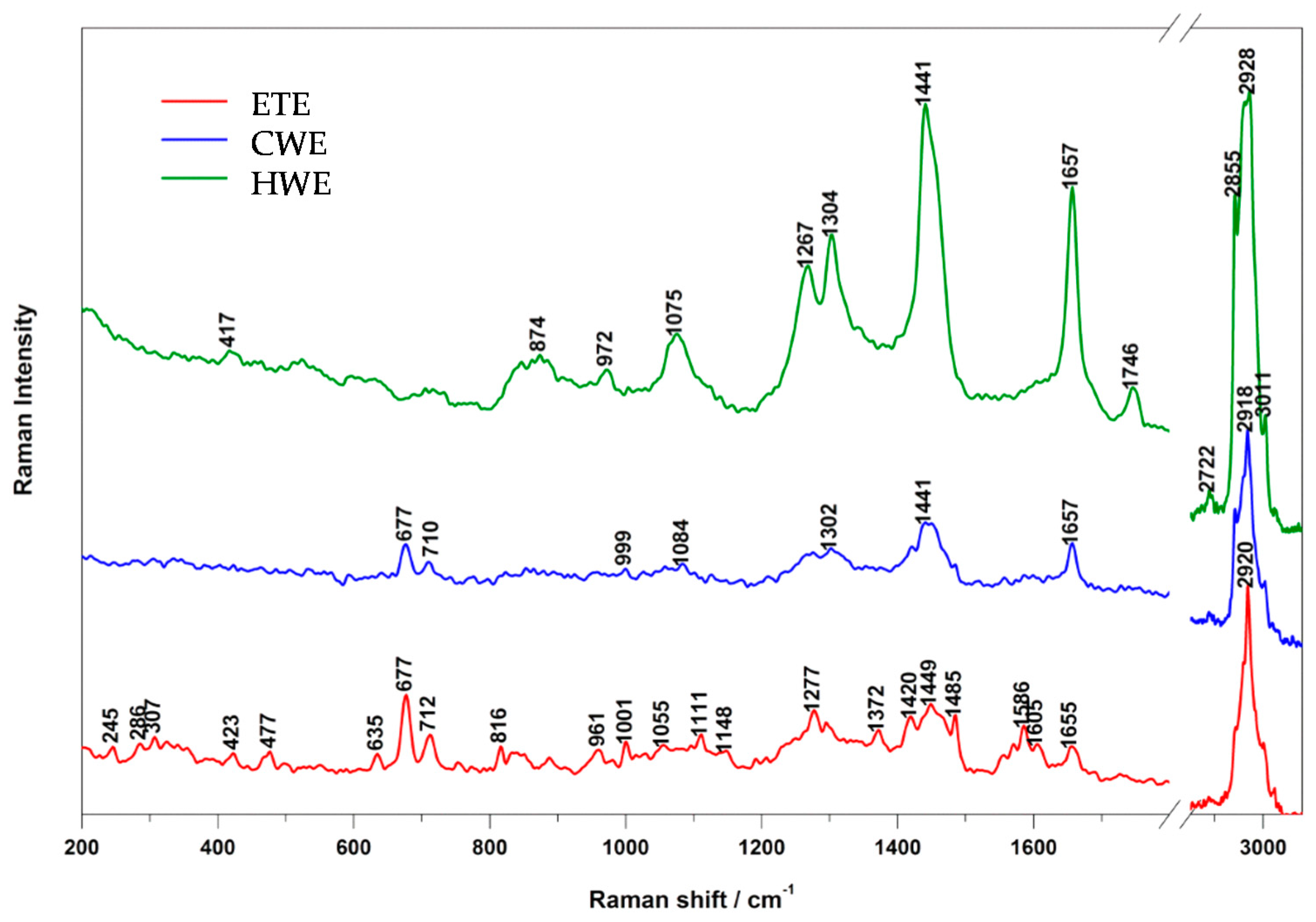
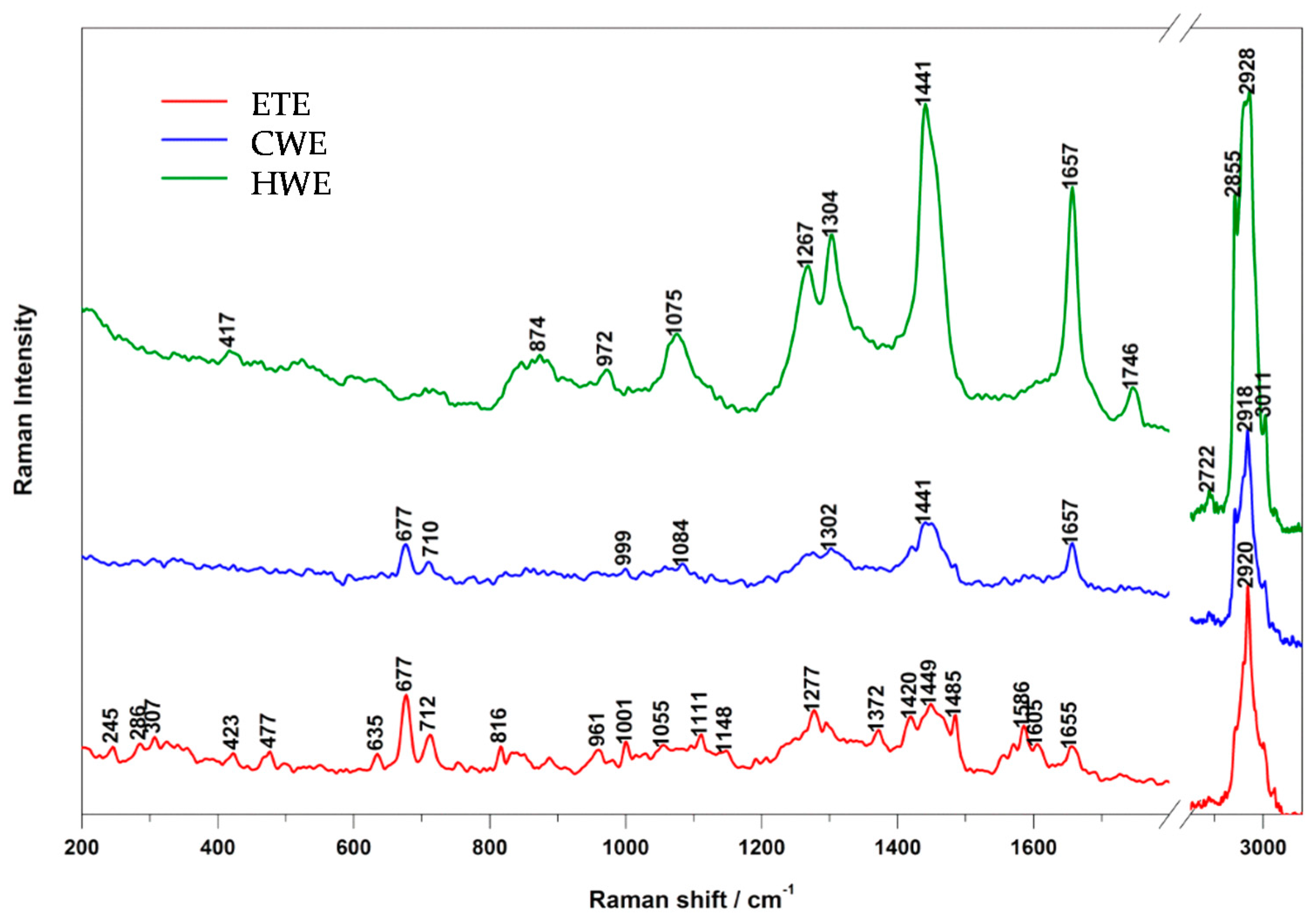
2.1. Raman and IR Spectroscopy
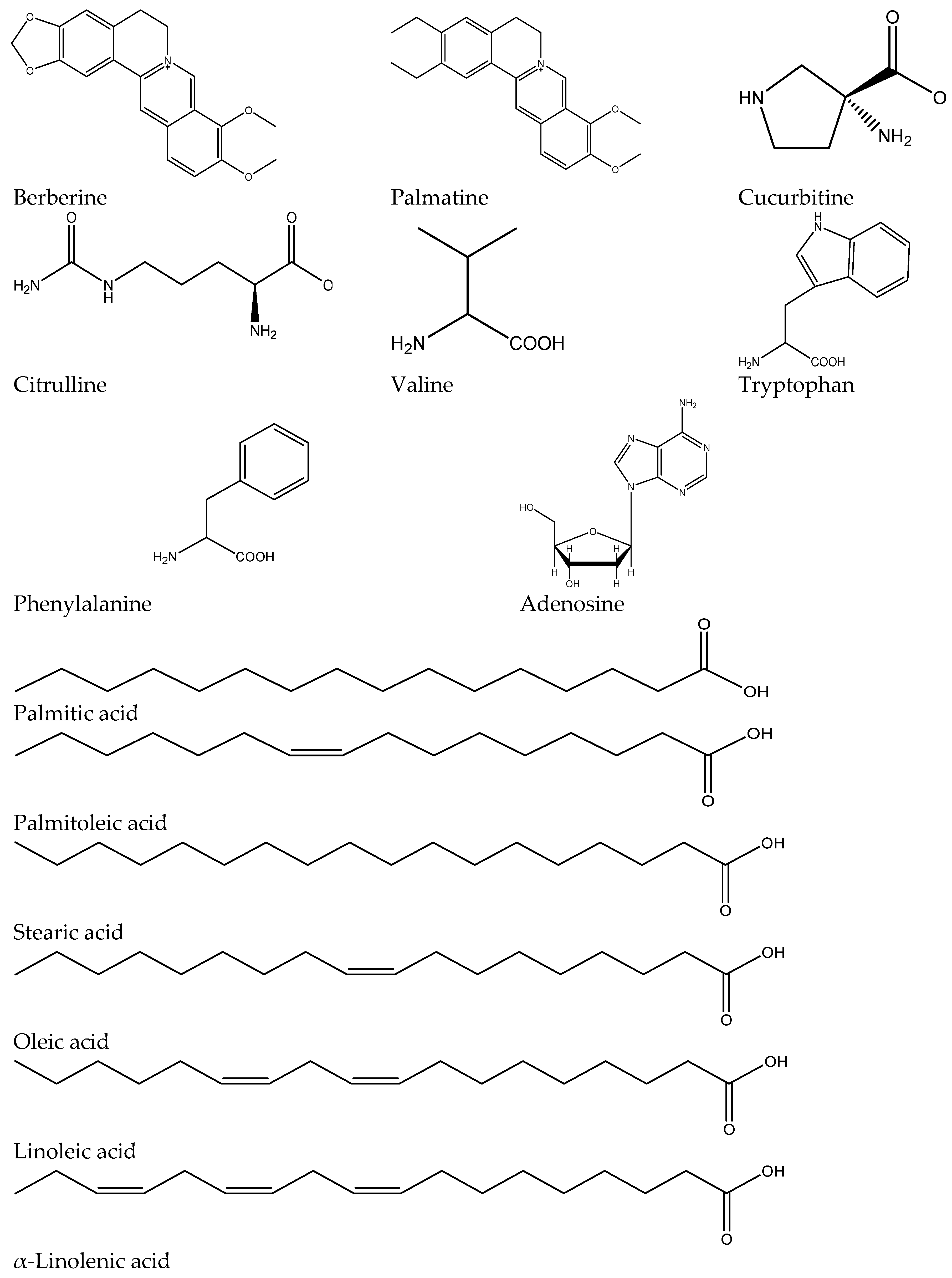
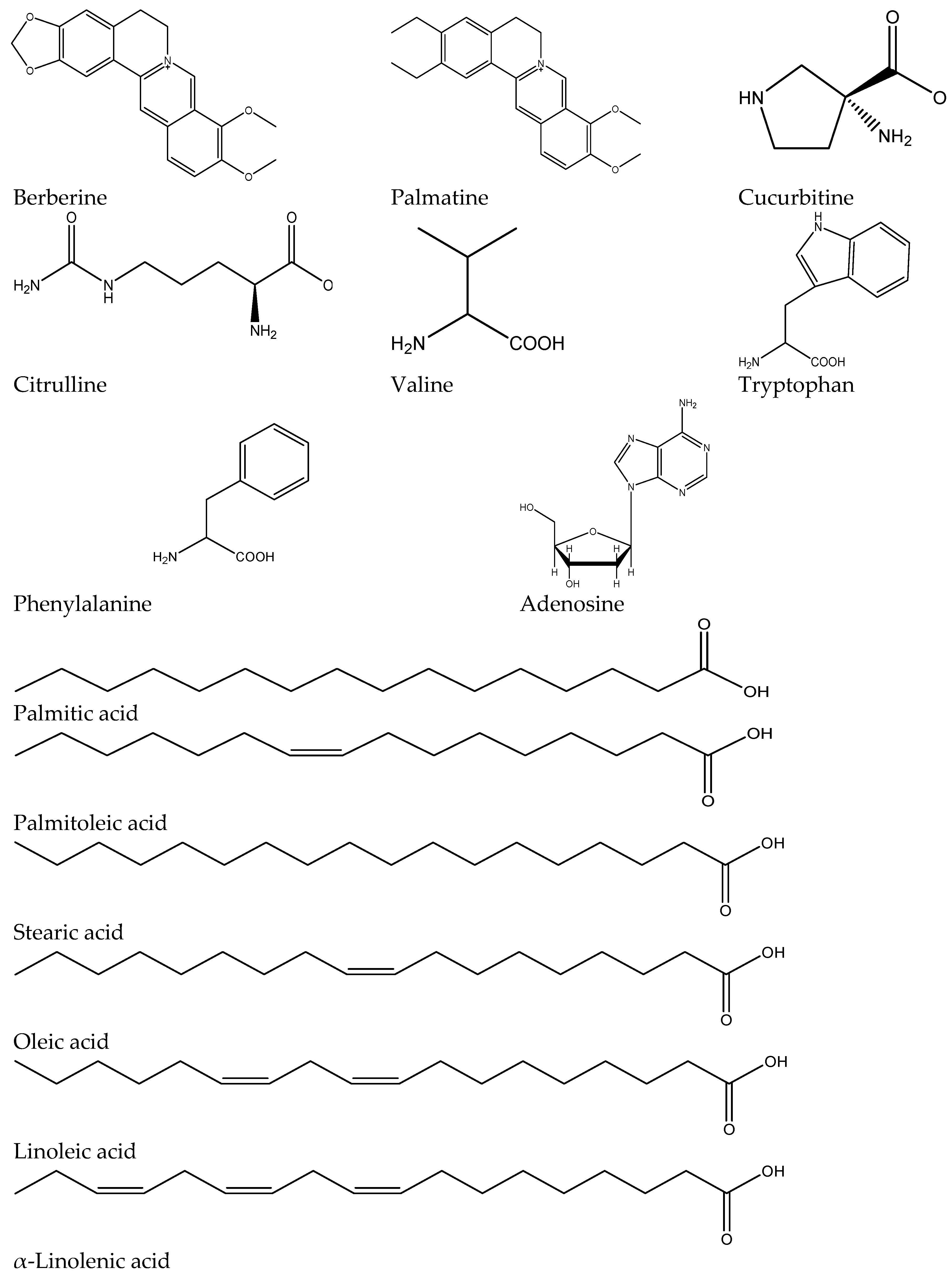
2.2. Quantitative and Qualitative LC-MS Profiling of Extracts
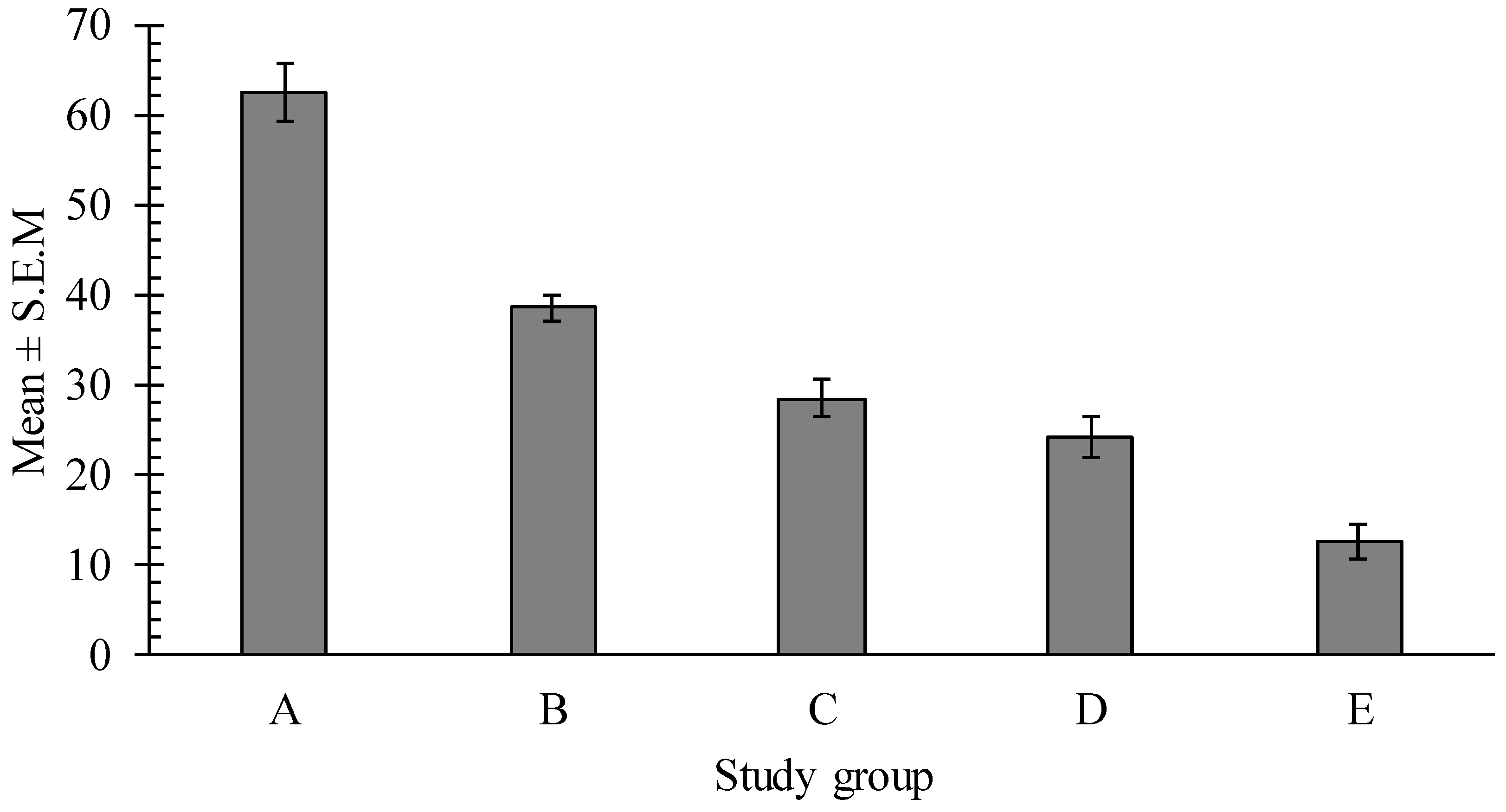
2.3. Effects of Pumpkin Seeds Extracts on Caenorhabditis elegans (C. elegans) Motility
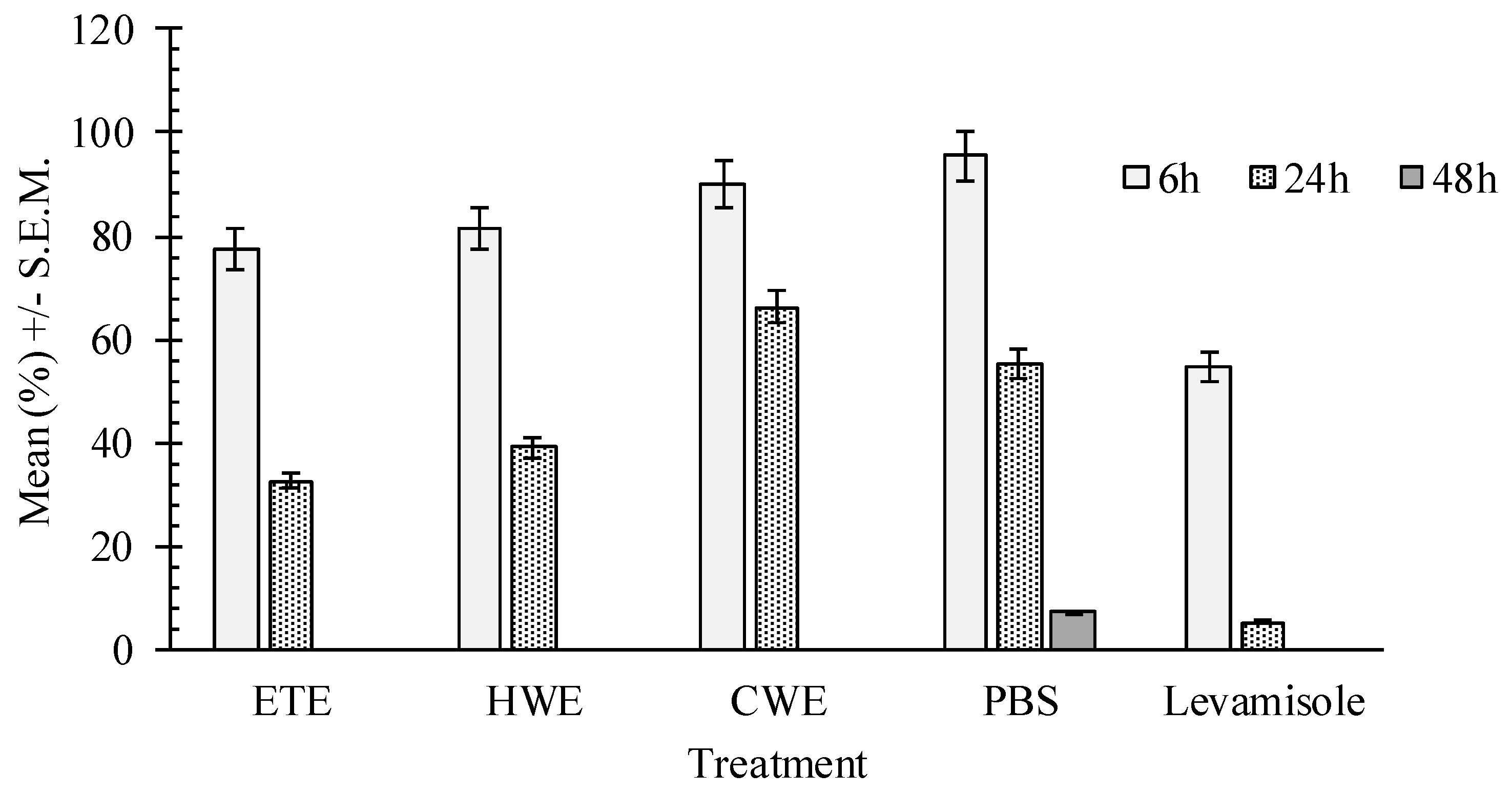
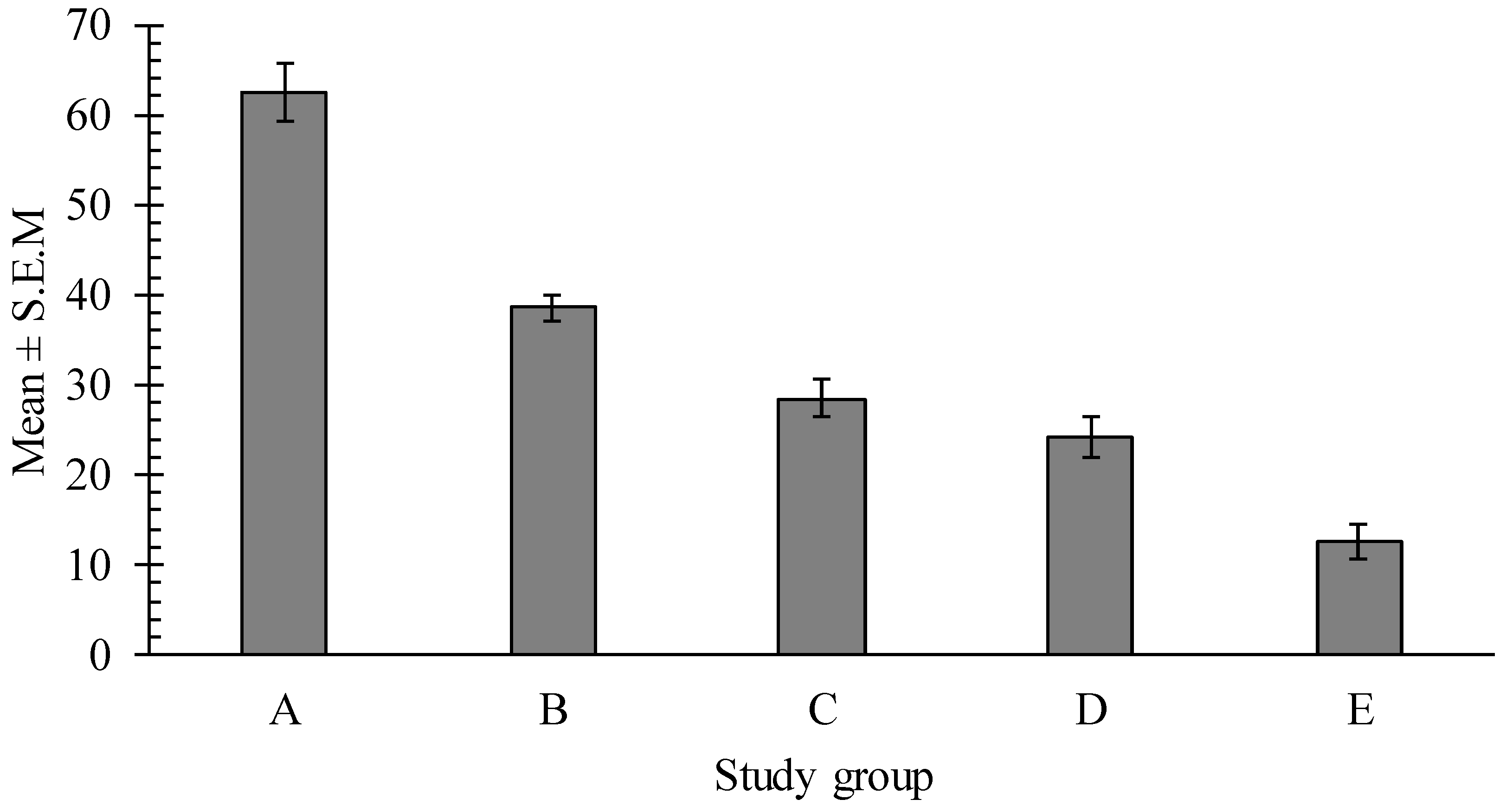
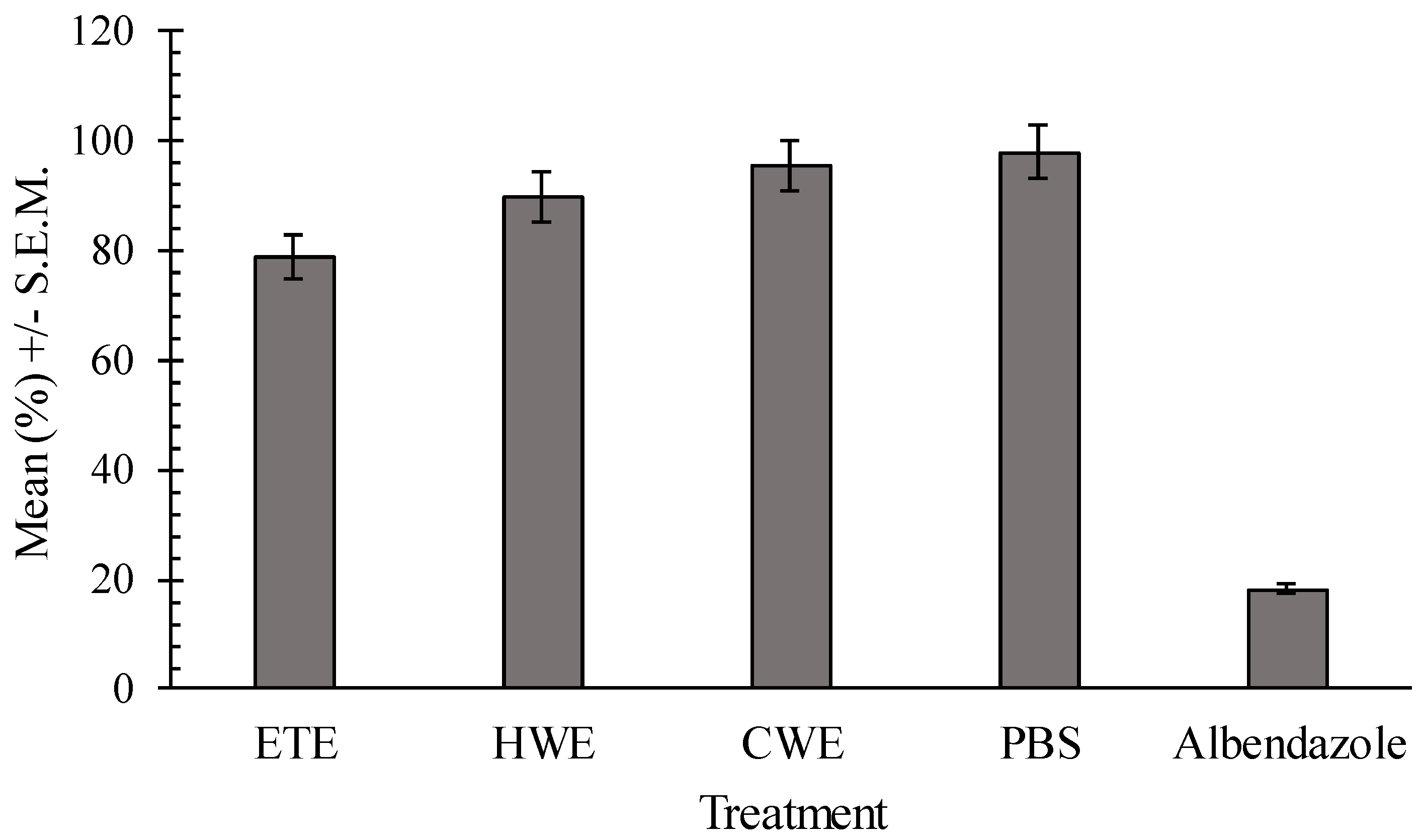
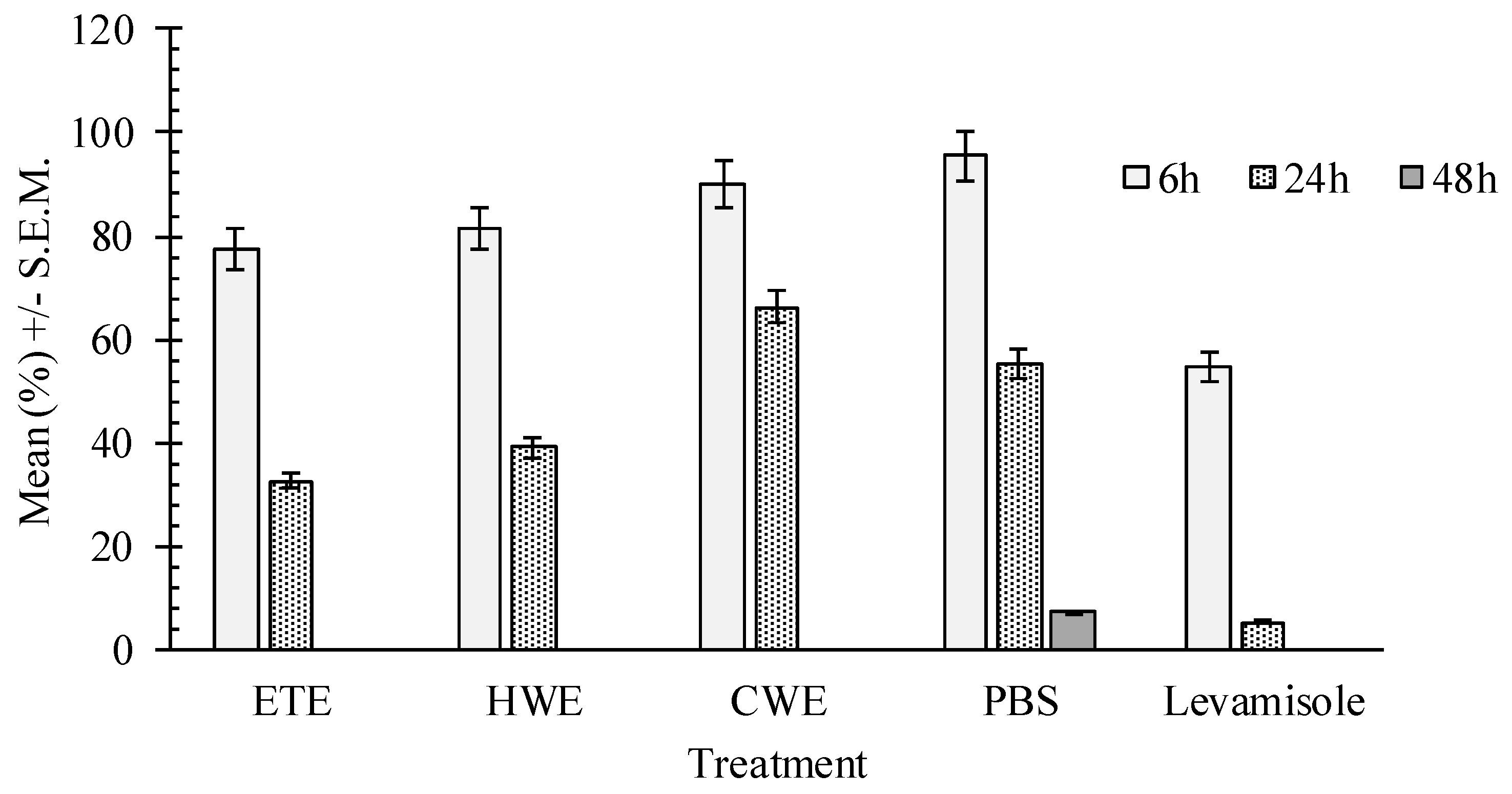
2.4. In Vitro Effects of Pumpkin Seed Extracts on Heligmosoides bakeri (H. bakeri)
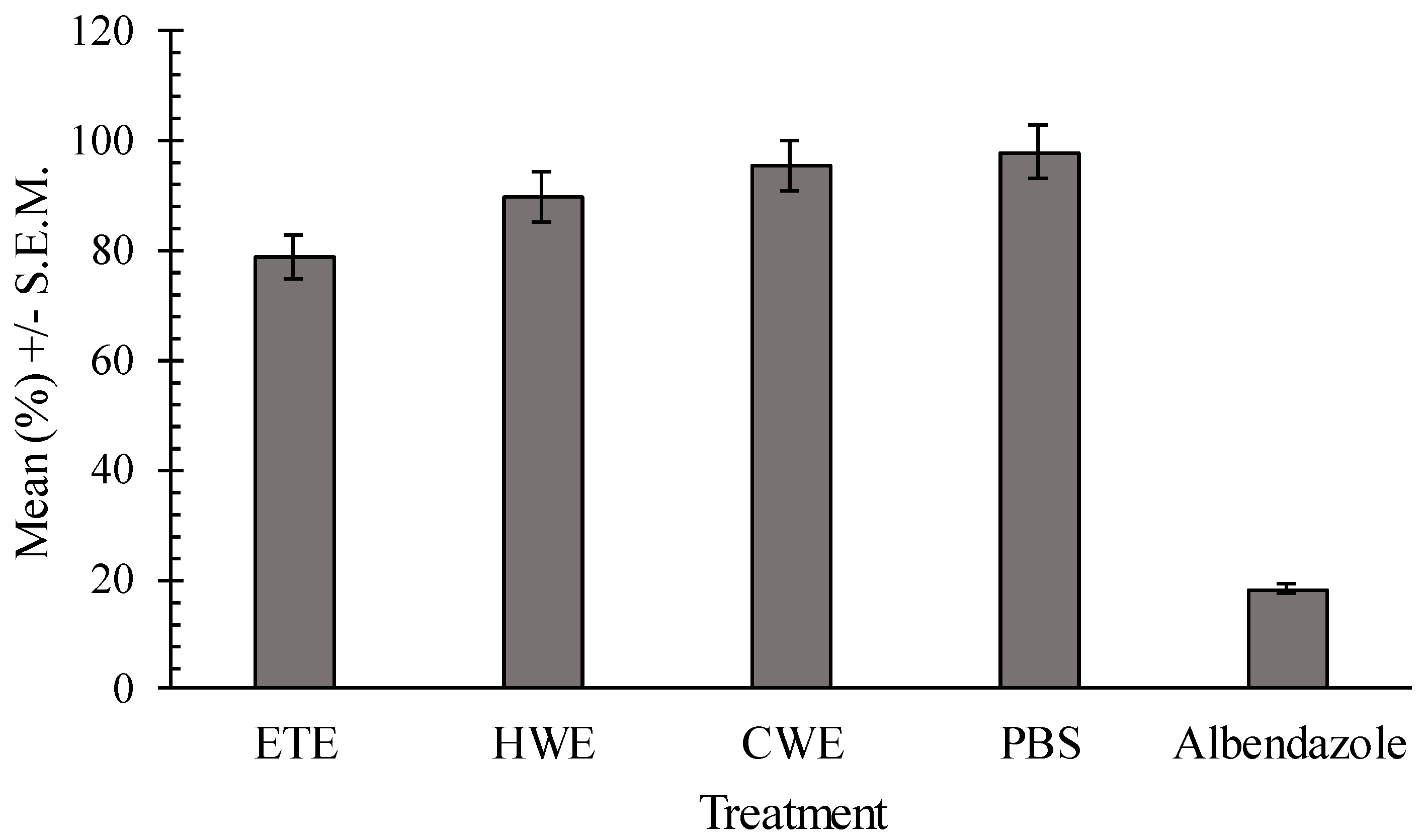
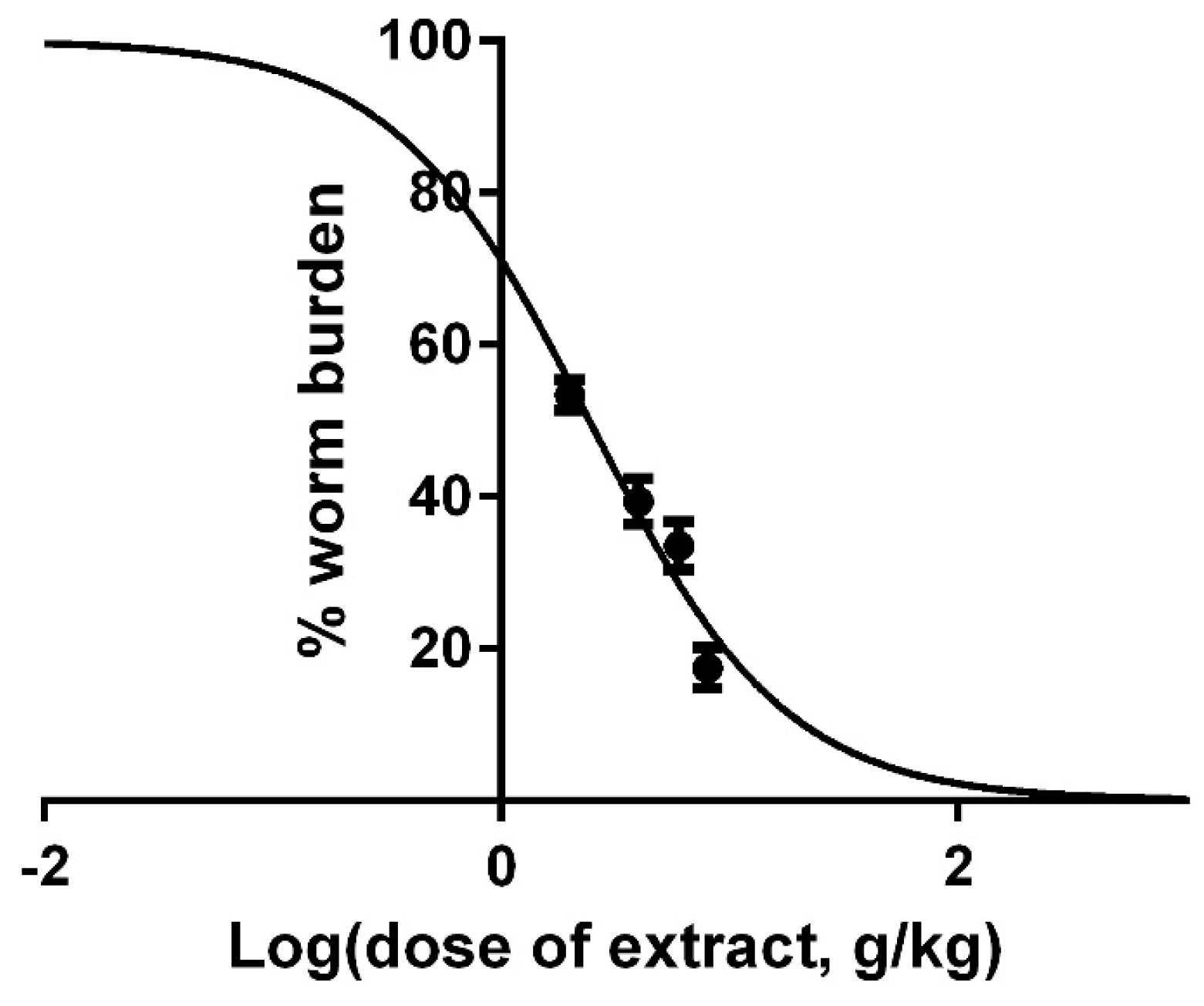
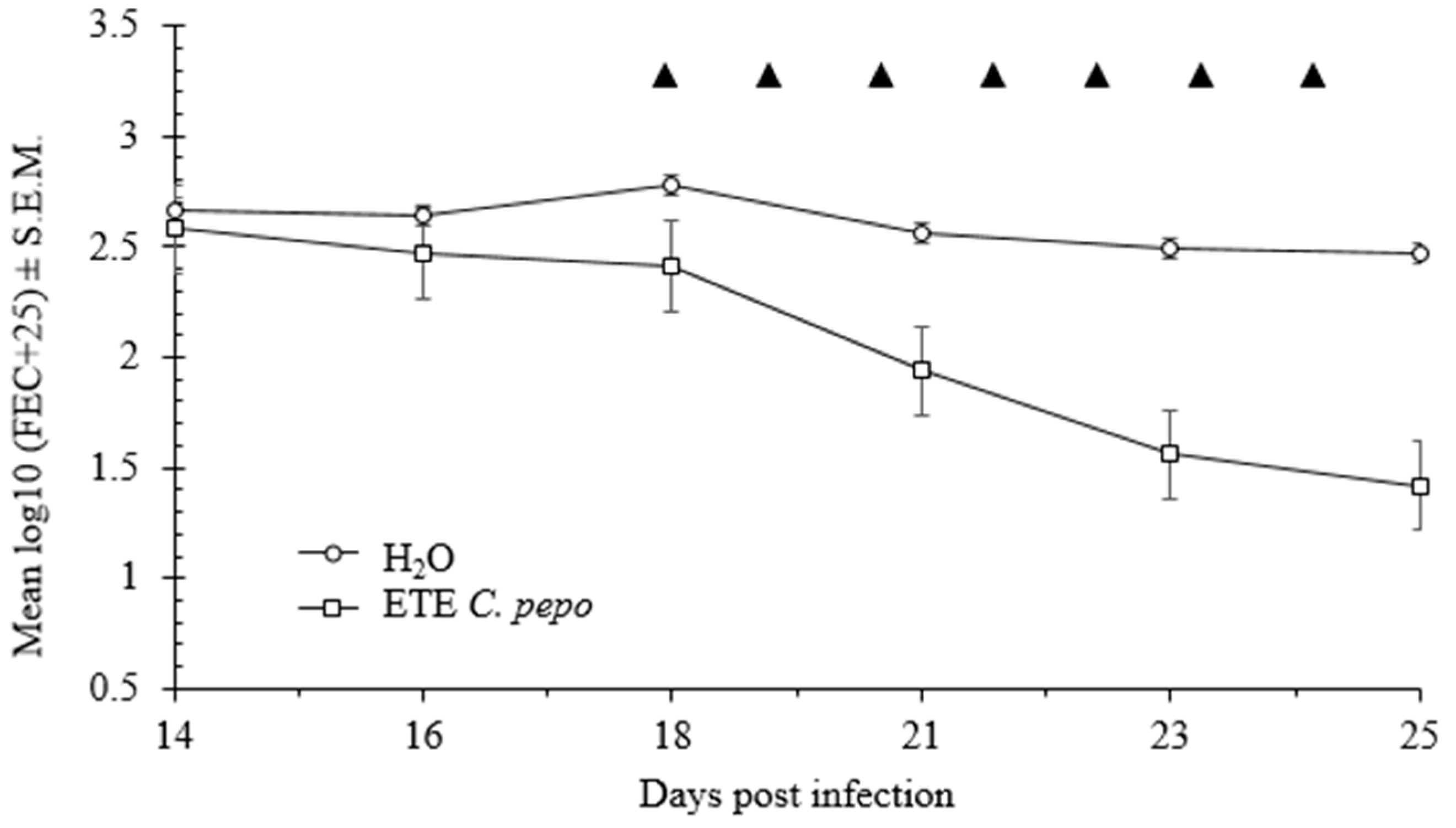
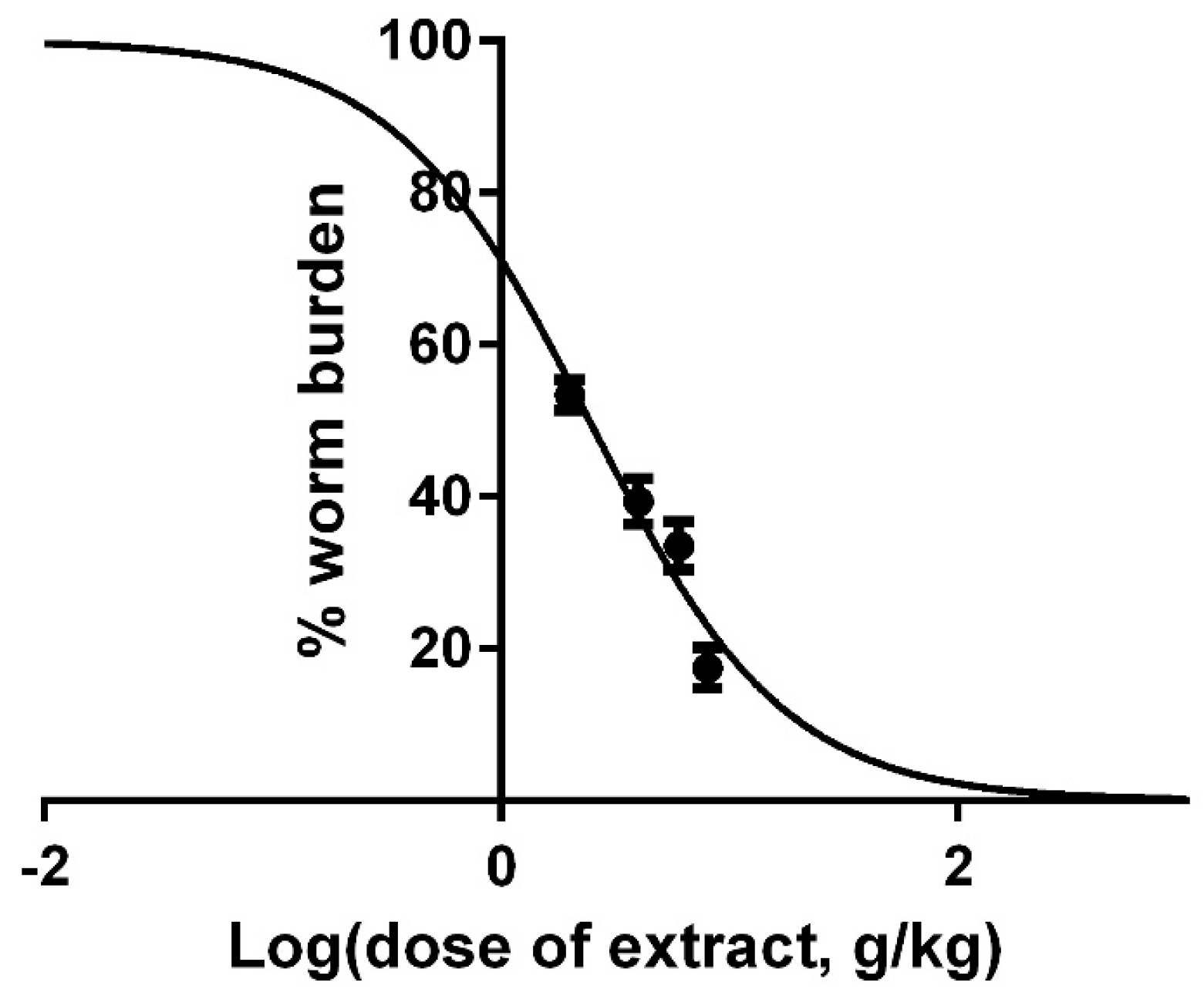
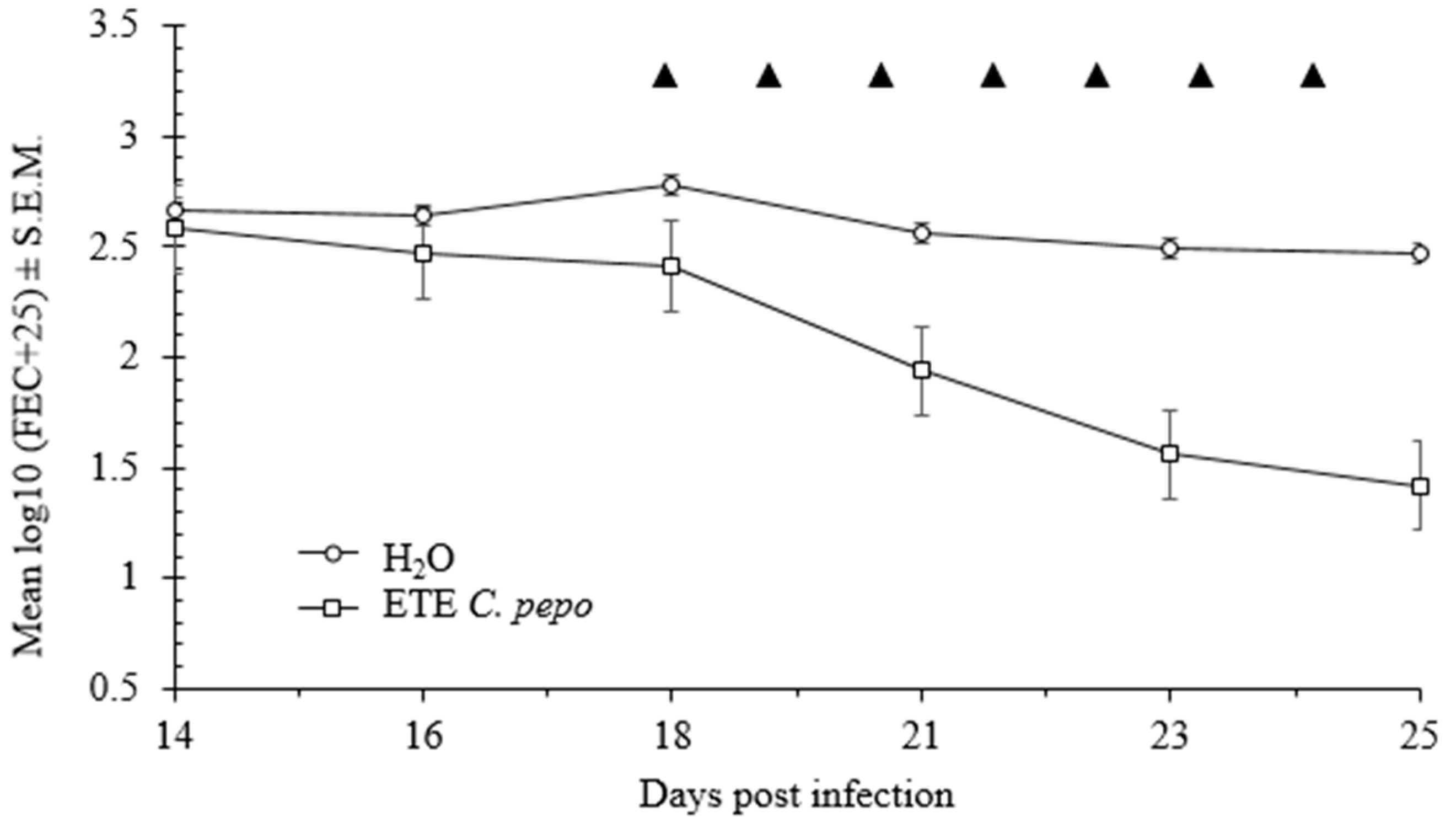
2.5. In Vivo Effects of Pumpkin Seeds Ethanol Extract on H. bakeri
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Plant Material and Preparation of Extracts
4.3. Spectroscopic Analysis of the Extracts
4.3.1. Raman Spectroscopy and IR Spectroscopy
4.3.2. HPLC-ESI-TOF-MS Quantitative and Qualitative Analysis of Extracts
- 0–15 min: a gradient of B in A from 3%–20%;
- 15–18 min: a quick gradient of B in A from 20%–40%;
- 18–21 min: gradient from 40%–95% of B in A;
- 21–23 min: an isocratic run with 95% of B in A;
- 23–24 min: a gradient from 95%–3% of B in A.
4.4. Caenorhabditis elegans Studies
4.4.1. Source and Maintenance of C. elegans Strains
4.4.2. C. elegans Exposure to Extracts and Motility Assay
4.5. Source of the Experimental Mice and Their Maintenance
4.6. In Vitro Anthelmintic Assays Using H. bakeri
4.6.1. Egg Hatch Assay
4.6.2. Larval Development Assay
4.6.3. Adult Worms Motility Assay
4.7. In Vivo Anthelmintic Assay
Experimental Protocol
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Horton, J. Human gastrointestinal helminth infections: Are they now neglected diseases? Trends Parasitol. 2003, 19, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Ndao, M.; Belot, J.; Zinsstag, J.; Pfister, K. Épidémiologie des helminthoses gastro-intestinales des petits Ruminants dans la zone sylvo-pastorale au Sénégal. Vet. Res. 1995, 26, 132–139. [Google Scholar] [PubMed]
- Perry, B.; Randolph, T. Improving the assessment of the economic impact of parasitic diseases and of their control in production animals. Vet. Parasitol. 1999, 84, 145–168. [Google Scholar] [CrossRef]
- Prichard, R. Anthelmintic resistance. Vet. Parasitol. 1994, 54, 259–268. [Google Scholar] [CrossRef]
- Nieuwhof, G.J.; Bishop, S.C. Costs of the major endemic diseases of sheep in Great Britain and the potential benefits of reduction in disease impact. Anim. Sci. 2005, 81, 23–29. [Google Scholar] [CrossRef]
- Behnke, J.M.; Buttle, D.J.; Stepek, G.; Lowe, A.; Duce, I.R. Developing novel anthelmintics from plant cysteine proteinases. Parasit. Vectors 2008, 1, 29. [Google Scholar] [CrossRef] [PubMed]
- Waller, P.J. Sustainable helminth control of ruminants in developing countries. Vet. Parasitol. 1997, 71, 195–207. [Google Scholar] [CrossRef]
- Jabbar, A.; Iqbal, Z.; Kerboeuf, D.; Muhammad, G.; Khan, M.N.; Afaq, M. Anthelmintic resistance: The state of play revisited. Life Sci. 2006, 79, 2413–2431. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.M. Drug resistance in nematodes of veterinary importance: A status report. Trends Parasitol. 2004, 20, 477–481. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, D.; Sacko, M.; Behnke, J.; Gilbert, F.; Dorny, P.; Vercruysse, J. Failure of mebendazole in treatment of human hookworm infections in the southern region of Mali. Am. J. Trop. Med. Hyg. 1997, 57, 25–30. [Google Scholar] [PubMed]
- Flohr, C.; Tuyen, L.N.; Lewis, S.; Minh, T.T.; Campbell, J.; Britton, J.; Williams, H.; Hien, T.T.; Farrar, J.; Quinnell, R.J. Low efficacy of mebendazole against hookworm in Vietnam: Two randomized controlled trials. Am. J. Trop. Med. Hyg. 2007, 76, 732–736. [Google Scholar] [PubMed]
- Sangster, N.C. Anthelmintic resistance: past, present and future. Int. J. Parasitol. 1999, 29, 115–124. [Google Scholar] [CrossRef]
- Matthee, S.; Dreyer, F.H.; Hoffmann, W.A.; van Niekerk, F.E. An introductory survey of helminth control practices in south africa and anthelmintic resistance on Thoroughbred stud farms in the Western Cape Province. J. S. Afr. Vet. Assoc. 2002, 73, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A. A larval development test for the detection of anthelmintic resistance in nematodes of sheep. Res. Vet. Sci. 1990, 49, 198–202. [Google Scholar] [PubMed]
- Cirak, V.Y.; Güleğen, E.; Bauer, C. Benzimidazole resistance in cyathostomin populations on horse farms in western Anatolia, Turkey. Parasitol. Res. 2004, 93, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, A.J.; Fairweather, I.; Prichard, R.; von Samson-Himmelstjerna, G.; Sangster, N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004, 20, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Klimpel, S.; Abdel-Ghaffar, F.; Al-Rasheid, K.A.S.; Aksu, G.; Fischer, K.; Strassen, B.; Mehlhorn, H. The effects of different plant extracts on nematodes. Parasitol. Res. 2011, 108, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghaffar, F.; Semmler, M.; Al-Rasheid, K.A.S.; Strassen, B.; Fischer, K.; Aksu, G.; Klimpel, S.; Mehlhorn, H. The effects of different plant extracts on intestinal cestodes and on trematodes. Parasitol. Res. 2011, 108, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Kocyan, A.; Zhang, L.-B.; Schaefer, H.; Renner, S.S. A multi-locus chloroplast phylogeny for the Cucurbitaceae and its implications for character evolution and classification. Mol. Phylogenet. Evol. 2007, 44, 553–577. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, K.; Gupta, A.; Sharma, D.K.; Gill, N.S.; Goyal, A. A review on the medicinally important plants of the family cucurbitaceae. Asian J. Clin. Nutr. 2012, 4, 16–26. [Google Scholar] [CrossRef]
- Marie-Magdeleine, C.; Hoste, H.; Mahieu, M.; Varo, H.; Archimede, H. In vitro effects of Cucurbita moschata seed extracts on Haemonchus contortus. Vet. Parasitol. 2009, 161, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Guarrera, P.M. Traditional antihelmintic, antiparasitic and repellent uses of plants in Central Italy. J. Ethnopharmacol. 1999, 68, 183–192. [Google Scholar] [CrossRef]
- De Queiroz-Neto, A.; Mataqueiro, M.I.; Santana, A.E.; Alessi, A.C. Toxicologic evaluation of acute and subacute oral administration of Cucurbita maxima seed extracts to rats and swine. J. Ethnopharmacol. 1994, 43, 45–51. [Google Scholar] [CrossRef]
- Mihranian, V.H.; Abou-Chaar, C. Extraction, detection and estimation of cucurbitin in cucurbita seeds. Lloydia 1968, 31, 23–29. [Google Scholar]
- Ponka, R.; Bouba, A.A.; Fokou, E.; Tambe, S.T.; Beaucher, E.; Piot, M.; Leonil, J.; Gaucheron, F. Protein, mineral and amino acid content of some Cameroonian traditional dishes prepared from pumpkin (Cucurbita maxima Duch.). J. Food Compos. Anal. 2015, 43, 169–174. [Google Scholar] [CrossRef]
- Vennerstrom, J.L.; Lovelace, J.K.; Waits, V.B.; Hanson, W.L.; Klayman, D.L. Berberine derivatives as antileishmanial drugs. Antimicrob. Agents Chemother. 1990, 34, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Leitao Da-Cunha, E.V.; Fechine, I.M.; Guedes, D.N. Protoberberine alkaloids. In The Alkaloids; Cordell, G.A., Ed.; Elsevier: New York, NY, USA, 2005; Volume 62, pp. 1–75. [Google Scholar]
- Schiff, P.L. The Thalictrum Alkaloids: Chemistry and Pharmacology. In Alkaloids: Chemical and Biological Perspectives; Pelletier, S.W., Ed.; Springer: New York, NY, USA, 1996; pp. 1–237. [Google Scholar]
- Dkhil, M.A. Role of berberine in ameliorating Schistosoma mansoni-induced hepatic injury in mice. Biol. Res. 2014, 47, 8. [Google Scholar] [CrossRef] [PubMed]
- Krivogorsky, B.; Pernat, J. A.; Douglas, K. A.; Czerniecki, N.J.; Grundt, P. Structure-activity studies of some berberine analogs as inhibitors of Toxoplasma gondii. Bioorg. Med. Chem. Lett. 2012, 22, 2980–2982. [Google Scholar] [CrossRef] [PubMed]
- Ropivia, J.; Derbré, S.; Rouger, C.; Pagniez, F.; Le Pape, P.; Richomme, P. Isoquinolines from the roots of Thalictrum flavum L. and their evaluation as antiparasitic compounds. Molecules 2010, 15, 6476–6484. [Google Scholar] [CrossRef] [PubMed]
- Atjanasuppat, K.; Wongkham, W.; Meepowpan, P.; Kittakoop, P.; Sobhon, P.; Bartlett, A.; Whitfield, P.J. In vitro screening for anthelmintic and antitumour activity of ethnomedicinal plants from Thailand. J. Ethnopharmacol. 2009, 123, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Beloin, N.; Gbeassor, M.; Akpagana, K.; Hudson, J.; de Soussa, K.; Koumaglo, K.; Arnason, J.T. Ethnomedicinal uses of Momordicacharantia (Cucurbitaceae) in Togo and relation to its phytochemistry and biological activity. J. Ethnopharmacol. 2005, 96, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Maciel, M.V.; Morais, S.M.; Bevilaqua, C.M.L.; Camurça-Vasconcelos, A.L.F.; Costa, C.T.C.; Castro, C.M.S. Ovicidal and larvicidal activity of Melia azedarach extracts on Haemonchus contortus. Vet. Parasitol. 2006, 140, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Behnke, J.M.; Menge, D.M.; Noyes, H. Heligmosomoides bakeri: A model for exploring the biology and genetics of resistance to chronic gastrointestinal nematode infections. Parasitology 2009, 136, 1565–1580. [Google Scholar] [CrossRef] [PubMed]
- Bryant, V. The life cycle of Nematospiroides dubius, Baylis, 1926 (Nematoda: Heligmosomidae). J. Helminthol. 1973, 47, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.K.; O’Brien, D.J.; Whitley, N.C.; Burke, J.M.; Miller, J.E.; Barczewski, R.A. Investigation of possible pumpkin seeds and ginger effects on gastrointestinal nematode infection indicators in meat goat kids and lambs. Small Rumin. Res. 2016, 136, 1–6. [Google Scholar] [CrossRef]
- Vandamme, T.F.; Ellis, K.J. Issues and challenges in developing ruminal drug delivery systems. Adv. Drug Deliv. Rev. 2004, 56, 1415–1436. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, E.; Malek, K.; Baranska, M. Rapid approach to analyze biochemical variation in rat organs by ATR FTIR spectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 118, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Stuart, B.H. Infrared Spectroscopy: Fundamentals and Applications, 1st ed.; Wiley: Chichester, UK, 2004; pp. 71–93. [Google Scholar]
- Lin-Vien, D.; Colthup, N.B.; Fateley, W.G.; Grasselli, J.G. The Handbook of Infrared and Raman Spectroscopy Chracteristic Frequencies of Organic Molecules; Lin-Vien, B., Colthup, N., Fateley, W., Grasselli, J., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 9–306. [Google Scholar]
- Gravato-Nobre, M.J.; Nicholas, H.R.; Nijland, R.; O’Rourke, D.; Whittington, D.E.; Yook, K.J.; Hodgkin, J. Multiple genes affect sensitivity of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 2005, 171, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Yook, K.; Hodgkin, J. Mos1 mutagenesis reveals a diversity of mechanisms affecting response of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 2007, 175, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Partridge, F.A.; Tearle, A.W.; Gravato-Nobre, M.J.; Schafer, W.R.; Hodgkin, J. The C. elegans glycosyltransferase BUS-8 has two distinct and essential roles in epidermal morphogenesis. Dev. Biol. 2008, 317, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Guven, K.; Duce, J.A.; de Pomerai, D.I. Evaluation of a stress-inducible transgenic nematode strain for rapid aquatic toxicity testing. Aquat. Toxicol. 1994, 29, 119–137. [Google Scholar] [CrossRef]
- Williams, P.L.; Dusenbery, D.B. Aquatic toxicity testing using the nematode, Caenorhabditis elegans. Environ. Toxicol. Chem. 1990, 9, 1285–1290. [Google Scholar] [CrossRef]
- Phiri, A.M.; de Pomerai, D.; Buttle, D.J.; Behnke, J.M.B. Developing a rapid throughput screen for detection of nematicidal activity of plant cysteine proteinases: The role of Caenorhabditis elegans cystatins. Parasitology 2014, 141, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Cheesbrough, M. Medical Laboratory Manual for Tropical Countries, 2nd ed.; Cambridge University Press: Cambridge, UK, 1987; pp. 1–624. [Google Scholar]
- Smyth, J.D. Animal Parasitology; Cambridge University Press: Cambridge, UK, 1996; pp. 36–38. [Google Scholar]
- Pessoa, L.M.; Morais, S.M.; Bevilaqua, C.M.L.; Luciano, J.H.S. Anthelmintic activity of essential oil of Ocimum gratissimum Linn. and eugenol against Haemonchus contortus. Vet. Parasitol. 2002, 109, 59–63. [Google Scholar] [CrossRef]
- Hubert, J.; Kerboeuf, D. A microlarval development assay for the detection of anthelmintic resistance in sheep nematodes. Vet. Rec. 1992, 130, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Ademola, I. A survey on ivermectin resistance in strongyles of sheep in Oyo state Nigeria using larval development. Isr. J. Vet. Med. 2002, 57, 149–151. [Google Scholar]
- Wabo Poné, J.; Bilong Bilong, C.F.; Mpoame, M. In vitro nematicidal activity of extracts of Canthium mannii (Rubiaceae), on different life-cycle stages of Heligmosomoides polygyrus (Nematoda, Heligmosomatidae). J. Helminthol. 2010, 84, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Hounzangbe-Adote, M.S.; Paolini, V.; Fouraste, I.; Moutairou, K.; Hoste, H. In vitro effects of four tropical plants on three life-cycle stages of the parasitic nematode, Haemonchus contortus. Res. Vet. Sci. 2005, 78, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Behnke, J.M.; Parish, H.A. Nematospiroides dubius: Arrested development of larvae in immune mice. Exp. Parasitol. 1979, 47, 116–127. [Google Scholar] [CrossRef]
- Elliot, J.M. Some Methods for the Statistical Analysis of Samples of Benthic Invertebrates, 2nd ed.; Freshwater Biological Association: Ambleside, UK, 1977; pp. 23–68. [Google Scholar]
- Hochberg, Y. A Sharper Bonferroni Procedure for Multiple Tests of Significance. Biometrika 1988, 75, 800. [Google Scholar] [CrossRef]
- Fisher, R.A.; Yates, C.B.E. Statistical Tables for Biological, Agricultural and Medical Research, 6th ed.; Longman: London, UK, 1974; pp. 23–68. [Google Scholar]
- Hammond, J.A.; Fielding, D.; Bishop, S.C. Prospects for plant anthelmintics in tropical veterinary medicine. Vet. Res. Commun. 1997, 21, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Waller, P.J.; Bernes, G.; Thamsborg, S.M.; Sukura, A.; Richter, S.H.; Ingebrigtsen, K.; Höglund, J. Plants as de-worming agents of livestock in the Nordic countries: Historical perspective, popular beliefs and prospects for the future. Acta Vet. Scand. 2001, 42, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Githiori, J.B.; Höglund, J.; Waller, P.J.; Baker, R.L. The anthelmintic efficacy of the plant, Albizia anthelmintica, against the nematode parasites Haemonchus contortus of sheep and Heligmosomoides polygyrus of mice. Vet. Parasitol. 2003, 116, 23–34. [Google Scholar] [CrossRef]
- Tagboto, S.; Townson, S. Antiparasitic properties of medical plants and other naturally occurring products. Adv. Parasitol. 2001, 50, 199–295. [Google Scholar] [PubMed]
- Taylor, S.; Berridge, V. Medicinal plants and malaria: an historical case study of research at the London School of Hygiene and Tropical Medicine in the twentieth century. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 707–714. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
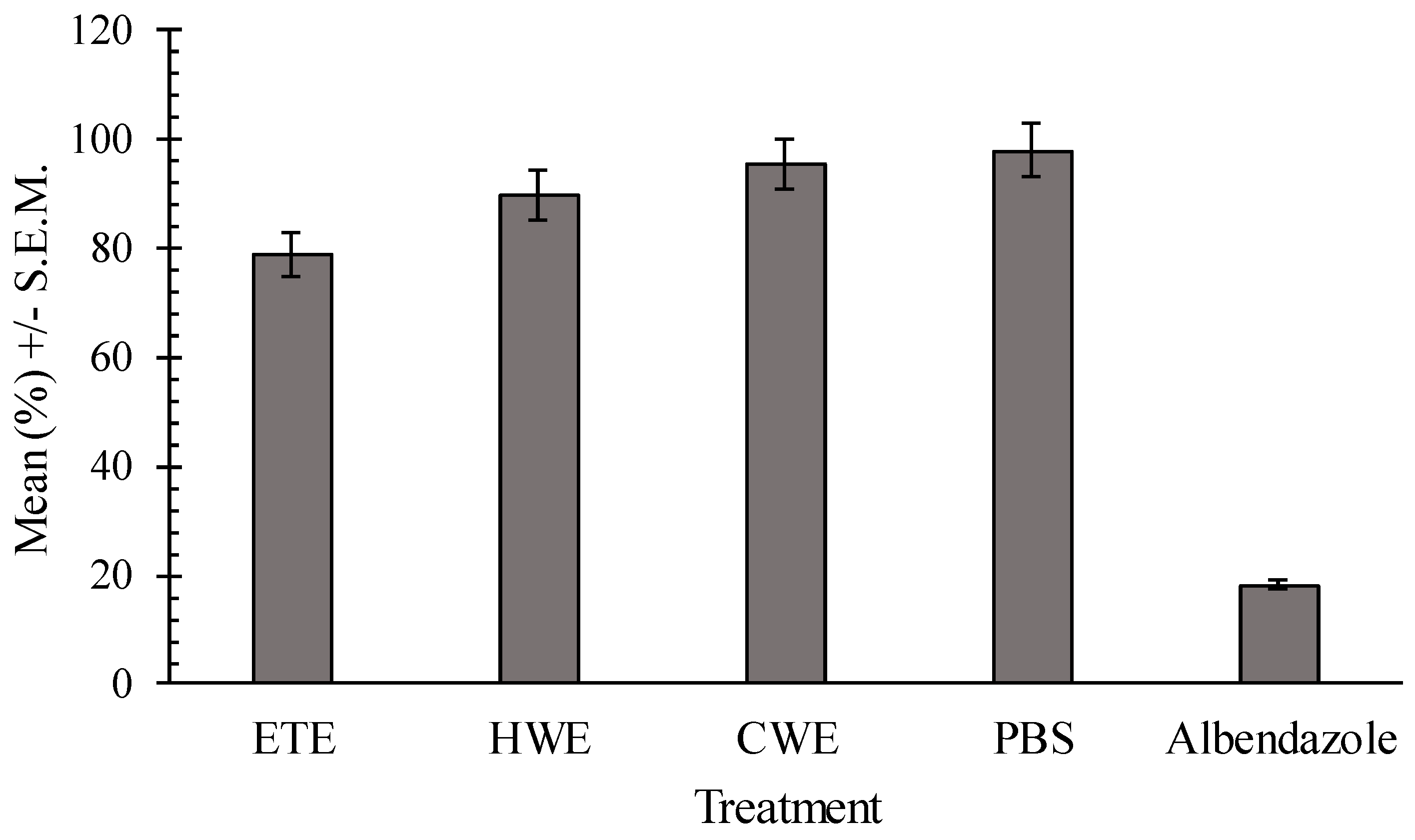{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HWE Band (cm−1) | CWE Band (cm−1) | ETE Band (cm−1) | Assignment |
|---|---|---|---|
| 417 | - | 423 | PO2 |
| - | - | 477 | polisaccharides, amylose, amylopectin |
| - | - | 635 | νC-S (AA: methionin) |
| - | 677 | 677 | DNA: ring, G |
| - | 710 | 712 | νC-S (AA: methionin) |
| - | - | 816 | proline, hydroxyproline, tyrosine, PO2 |
| 874 | - | - | hydroxyproline, tryptophane |
| 972 | - | 961 | fatty acids |
| - | 999 | 1001 | phenylalanine |
| - | - | 1055 | amylose |
| 1075 | - | - | fatty acids |
| - | 1084 | - | DNA |
| - | - | 1111 | cellulose |
| - | - | 1148 | carbohydrates |
| 1267 | - | 1277 | unsaturated fatty acids |
| 1304 | 1302 | 1295 | fatty acids |
| - | - | 1372 | DNA: T, A, G; saccharides |
| - | - | 1420 | DNA: A, G |
| 1441 | 1441 | 1449 | fatty acids |
| - | - | 1485 | DNA: A, G |
| - | - | 1556 | Amide II |
| - | - | 1571 | DNA: A, G |
| - | - | 1586 | phenylalanine, hydroxyproline |
| - | - | 1605 | phenylalanine, tyrosine, NO2 |
| 1657 | 1657 | 1655 | unsaturated fatty acids/proteins (amide I) |
| 1746 | - | - | fatty acids |
| 2722 | - | - | νC-H, νN-H, νO-H |
| 2855 | 2854 | - | saturated fatty acids |
| 2928 | 2918 | 2920 | saturated fatty acids |
| 3011 | 3008 | - | unsaturated fatty acids |
| Mode | Rt (min) | Molecular Formula | m/z exp. | m/z calc. | Δ ppm | RDB | In Source ESI-MS Fragments | Proposed Compound | ETE | HWE | CWE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaloids | |||||||||||
| [M + H]+ | 25.47 | C20H18NO4 | 335.1155 | 335.1158 | 0.7 | 13 | 321, 306, 292, 278 | Berberine | ++ | ++ | + |
| [M + H]+ | 25.87 | C21H22NO4 | 352.1539 | 352.1543 | 1.24 | 12 | 337, 322, 308, 292 | Palmatine | + | ++ | + |
| Amino Acids | |||||||||||
| [M + H]+ | 2.2 | C5H10N2O2 | 131.0832 | 131.0815 | 2.34 | 2 | ND | Cucurbitine | + | + | ++ |
| [M + H]+ | 2.6 | C6H13N3O3 | 175.0978 | 175.0951 | 1.56 | 2 | ND | Citrulline | + | + | + |
| [M + H]+ | 14.20 | C9H11NO2 | 166.0860 | 166.0863 | 1.55 | 5 | 120, 103 | l-Phenylalanine | + | ++ | +++ |
| [M + H]+ | 15.63 | C11H12N2O2 | 205.0974 | 205.0972 | −1.2 | 7 | 188, 146, 118 | l-Tryptophan | +++ | + | +++ |
| [M + H]+ | 8.85 | C5H10NO2 | 118.0855 | 118.0863 | 6.45 | 1 | ND | l-Valine | +++ | + | + |
| Nucleosides | |||||||||||
| [M + H]+ | 15.49 | C10H13N5O4 | 268.1037 | 268.1040 | 1.24 | 7 | 184, 136, 118 | Adenosine | ++ | ++ | + |
| Fatty Acids | |||||||||||
| [M − H]− | 69.40 | C16H32O2 | 255.2330 | 255.2322 | 2.94 | 1 | ND | Palmitic acid | ++++ | +++ | ++++ |
| [M − H]− | 66.32 | C16H30O2 | 253.2143 | 253.2139 | −3.0 | 2.5 | 112 | Palmitoleic acid | ++ | + | ++ |
| [M − H]− | 63.52 | C18H36O2 | 283.2643 | 283.2642 | 0.19 | 1 | 239 | Stearic acid | - | ++ | ++ |
| [M − H]− | 70.96 | C18H34O2 | 281.2486 | 281.2492 | −2.11 | 2 | 237 | Oleic acid | ++ | +++ | ++++ |
| [M − H]− | 67.06 | C18H32O2 | 279.2330 | 279.2323 | 2.33 | 3 | 235 | Linoleic acid | +++ | ++++ | +++++ |
| [M − H]− | 64.33 | C18H30O2 | 277.2173 | 277.2179 | −2.14 | 4 | 233, 205 | α-Linolenic acid | ++ | + | + |
| Strain | Control | 1000 µg/mL CWE | 1000 µg/mL HWE | 1000 µg/mL ETE | 75 µg/mL HWE | 75 µg/mL ETE |
|---|---|---|---|---|---|---|
| Wild-type | 5 | 5 | 5 | 5 | 5 | 5 |
| 4 | 5 | 5 | 5 | 5 | 5 | |
| 4 | 4 | 4 | 4 | 5 | 5 | |
| 4 | 4 | 4 | 5 | 5 | 5 | |
| cpi-1-/- | 4 | 5 | 5 | 5 | 5 | 5 |
| 4 | 5 | 5 | 5 | 5 | 5 | |
| 4 | 4 | 4 | 5 | 5 | 5 | |
| 4 | 4 | 5 | 5 | 3 | 5 | |
| CB 7031 | 4 | 5 | 5 | 5 | 5 | 5 |
| 4 | 5 | 5 | 5 | 5 | 5 | |
| 4 | 4 | 5 | 5 | 5 | 4 | |
| 4 | 4 | 4 | 5 | 5 | 5 | |
| CB 7014 | 5 | 5 | 5 | 5 | 5 | 5 |
| 4 | 5 | 5 | 5 | 5 | 5 | |
| 4 | 4 | 4 | 5 | 5 | 4 | |
| 4 | 5 | 5 | 5 | 5 | 5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzybek, M.; Kukula-Koch, W.; Strachecka, A.; Jaworska, A.; Phiri, A.M.; Paleolog, J.; Tomczuk, K. Evaluation of Anthelmintic Activity and Composition of Pumpkin (Cucurbita pepo L.) Seed Extracts—In Vitro and in Vivo Studies. Int. J. Mol. Sci. 2016, 17, 1456. https://doi.org/10.3390/ijms17091456
Grzybek M, Kukula-Koch W, Strachecka A, Jaworska A, Phiri AM, Paleolog J, Tomczuk K. Evaluation of Anthelmintic Activity and Composition of Pumpkin (Cucurbita pepo L.) Seed Extracts—In Vitro and in Vivo Studies. International Journal of Molecular Sciences. 2016; 17(9):1456. https://doi.org/10.3390/ijms17091456
Chicago/Turabian StyleGrzybek, Maciej, Wirginia Kukula-Koch, Aneta Strachecka, Aleksandra Jaworska, Andrew M. Phiri, Jerzy Paleolog, and Krzysztof Tomczuk. 2016. "Evaluation of Anthelmintic Activity and Composition of Pumpkin (Cucurbita pepo L.) Seed Extracts—In Vitro and in Vivo Studies" International Journal of Molecular Sciences 17, no. 9: 1456. https://doi.org/10.3390/ijms17091456
APA StyleGrzybek, M., Kukula-Koch, W., Strachecka, A., Jaworska, A., Phiri, A. M., Paleolog, J., & Tomczuk, K. (2016). Evaluation of Anthelmintic Activity and Composition of Pumpkin (Cucurbita pepo L.) Seed Extracts—In Vitro and in Vivo Studies. International Journal of Molecular Sciences, 17(9), 1456. https://doi.org/10.3390/ijms17091456