
The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo

Abstract
:1. Introduction
2. Results
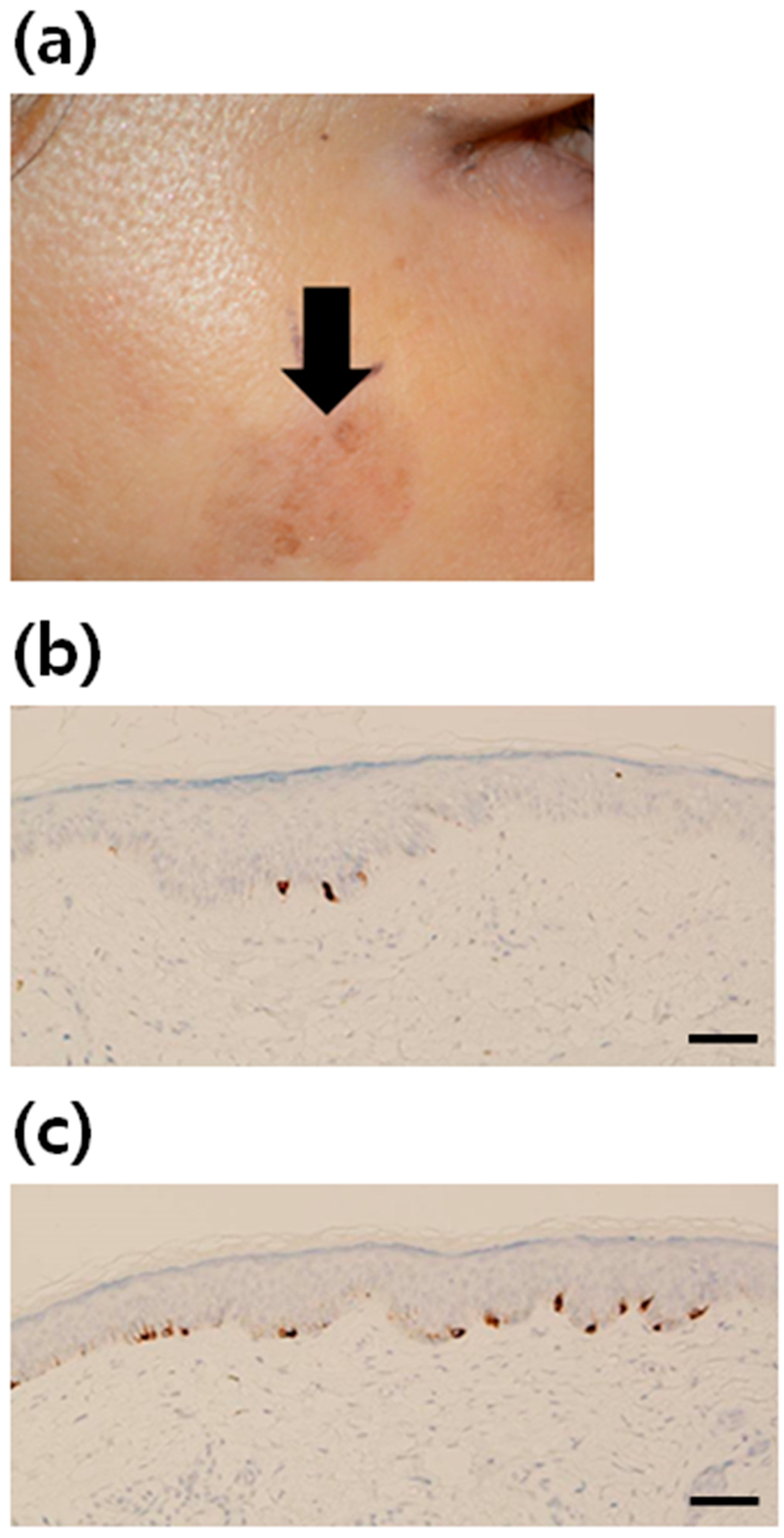
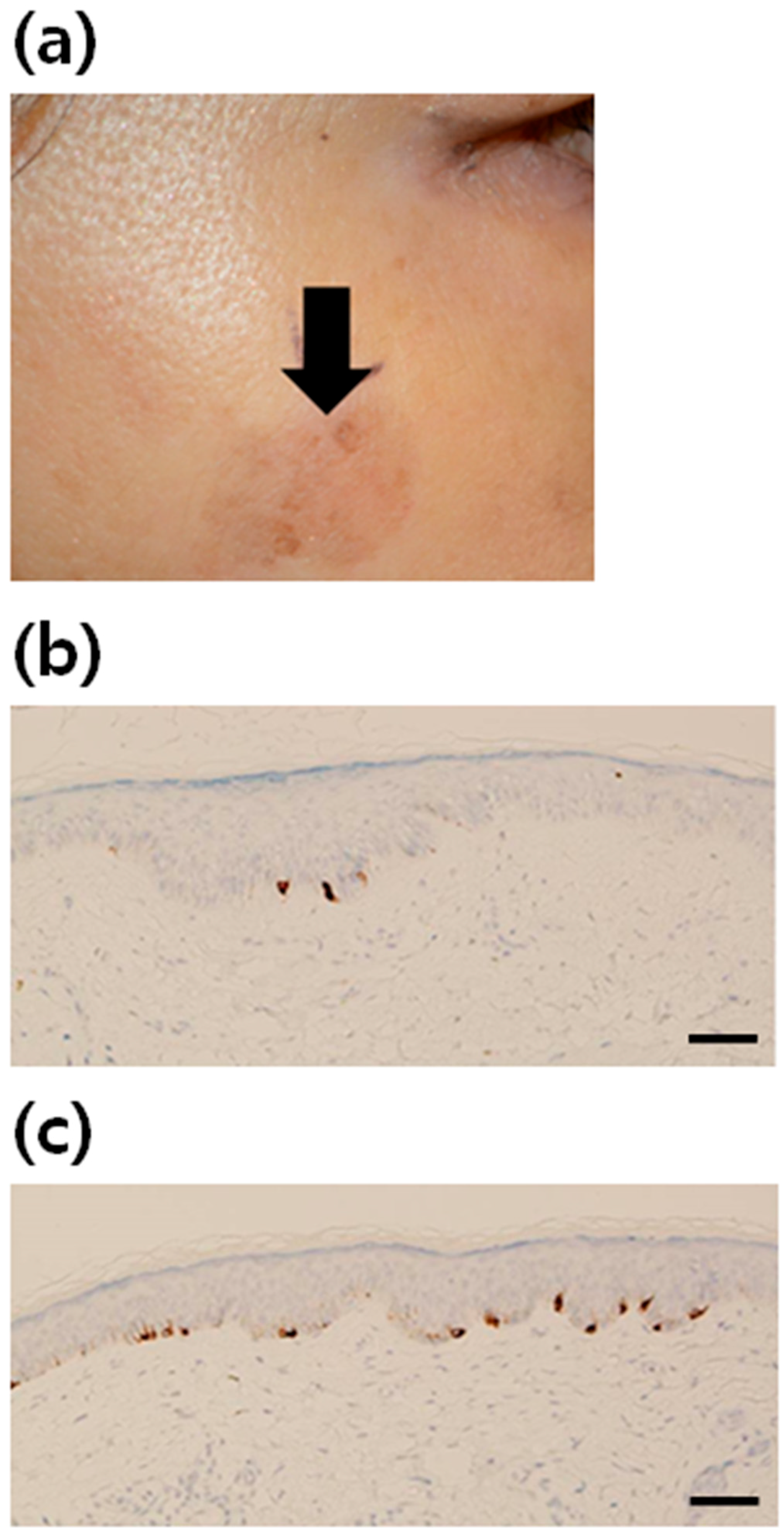
2.1. Increment of Melanocytes in Solar Lentigo (SL) Tissue
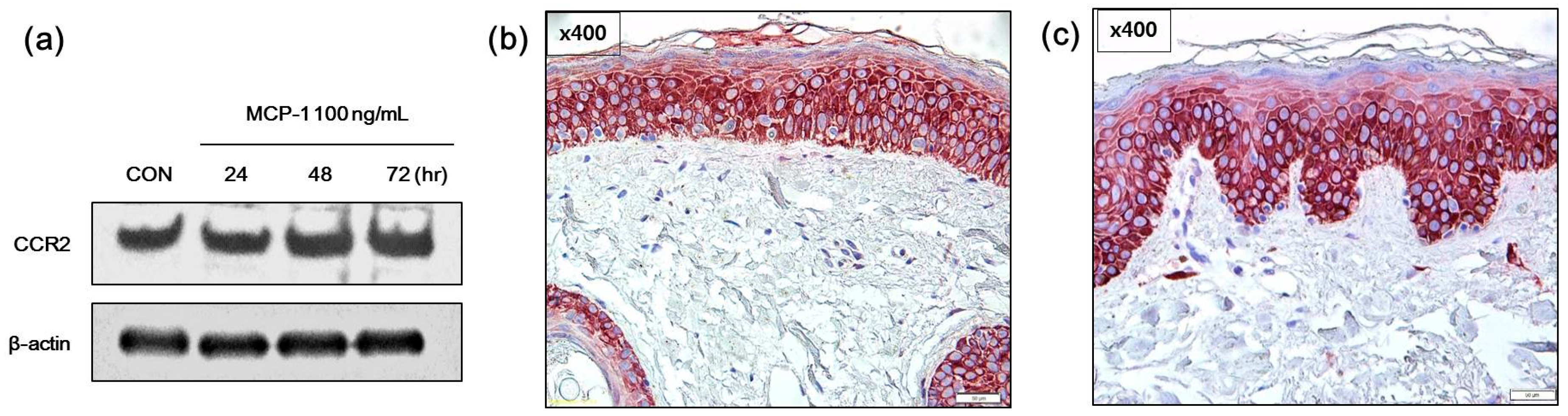
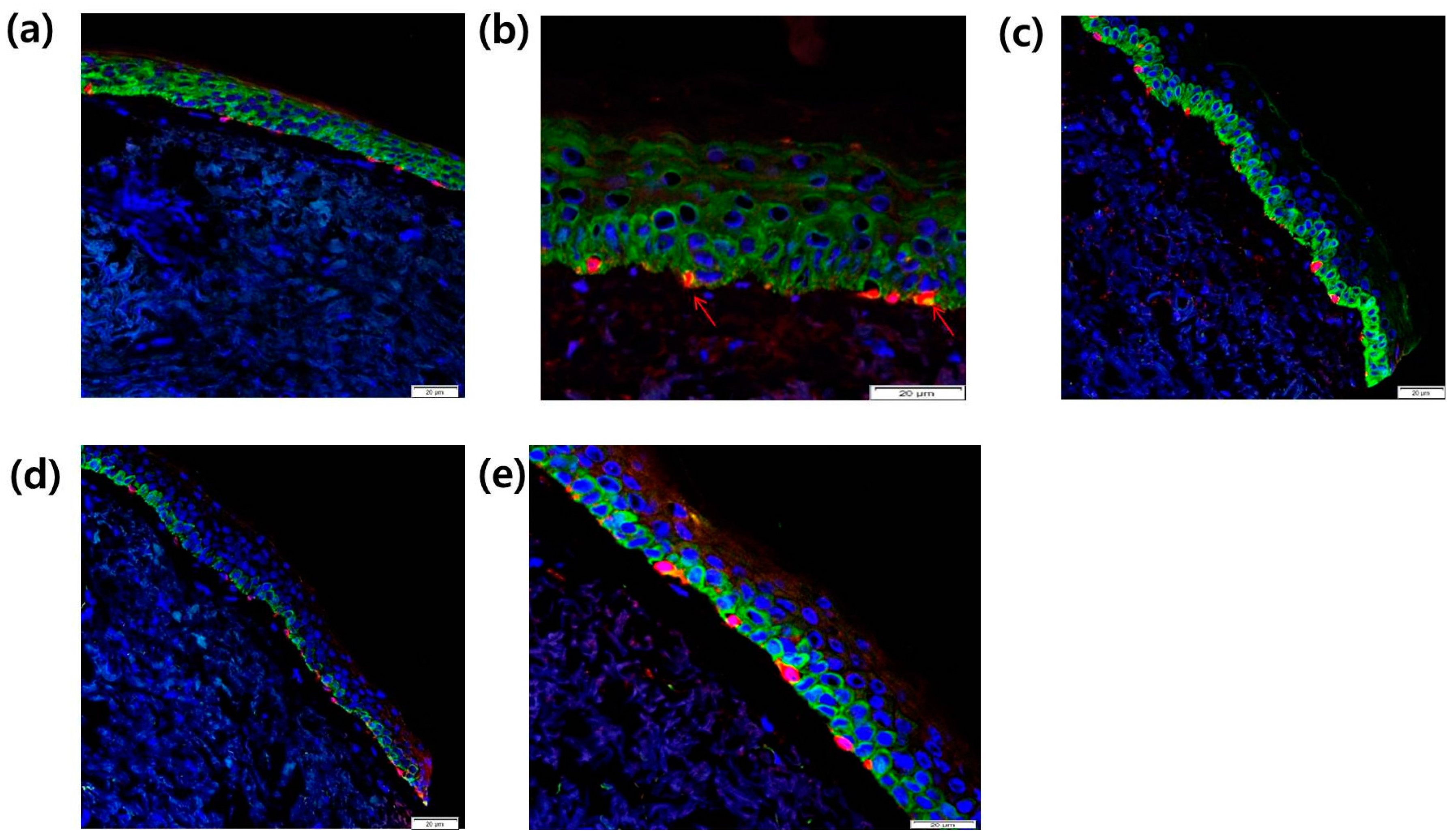
2.2. Expression of CCR2 in Normal Human Melanocytes (NHMs) and SL Skin Tissue
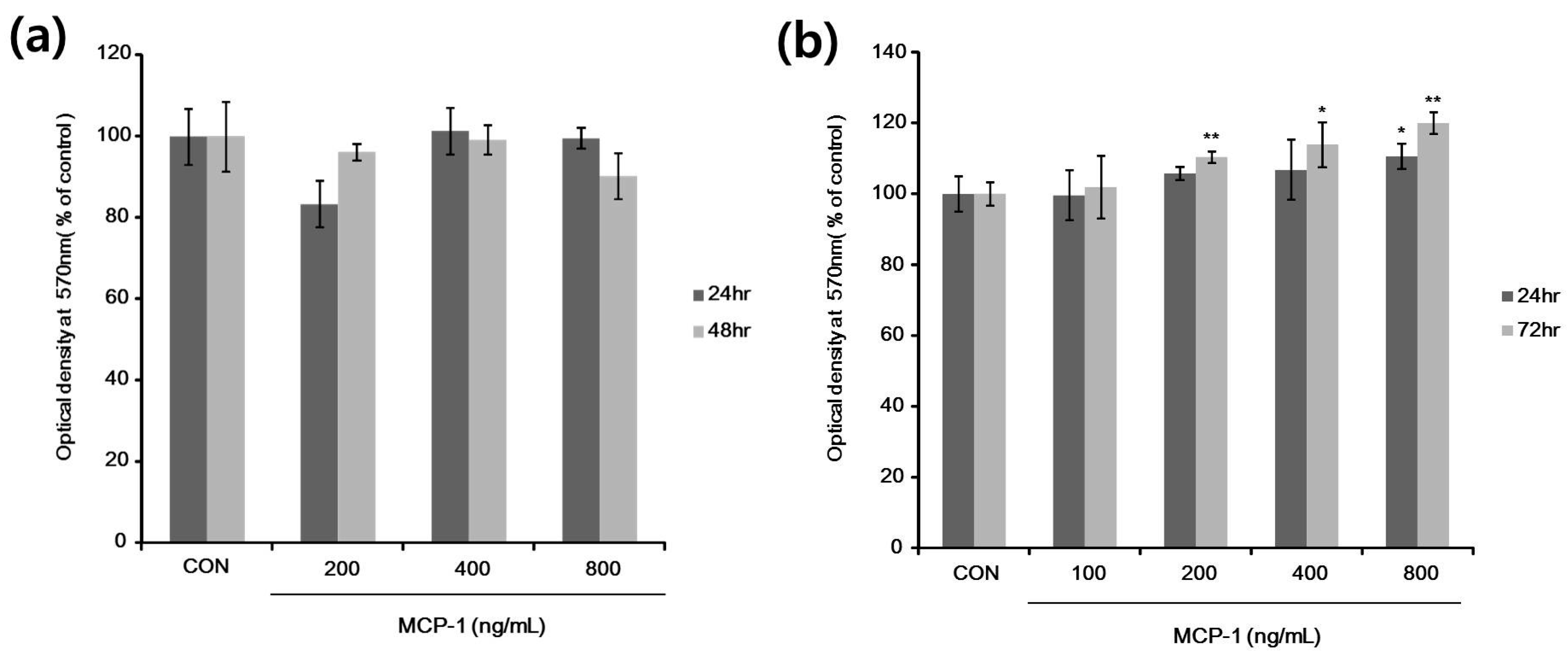
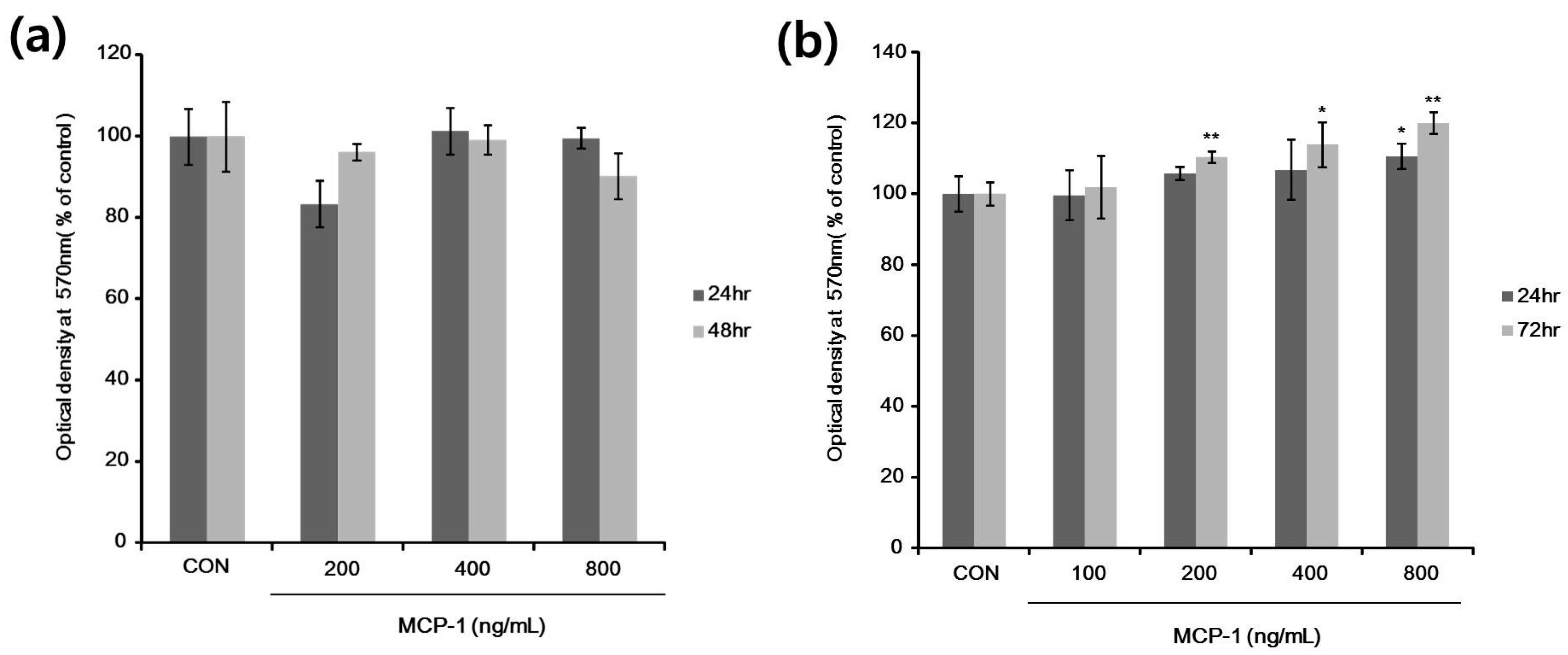
2.3. Cell Viability and Proliferation after MCP-1 Treatment in Normal Human Keratinocytes (NHKs) and NHMs
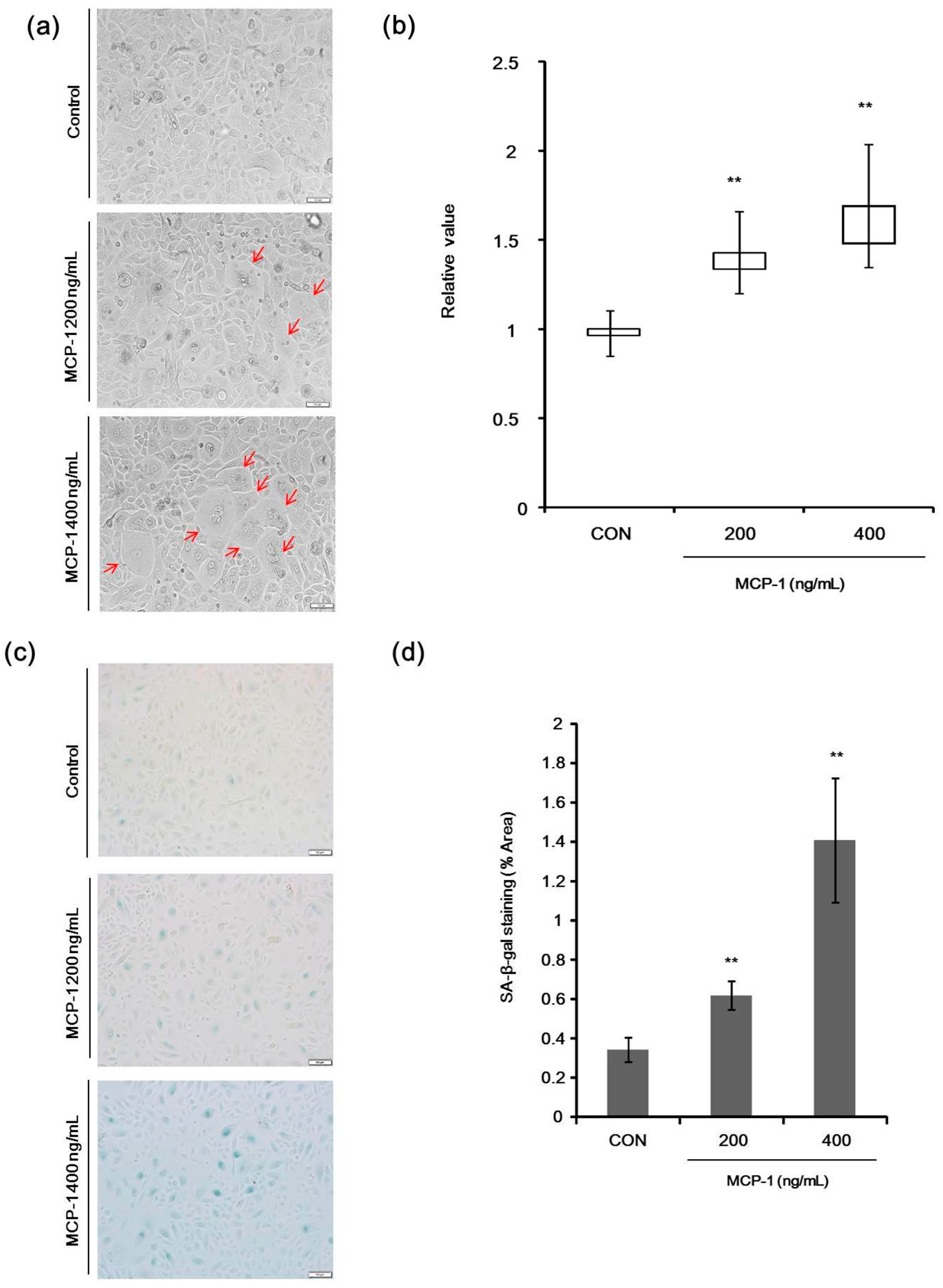
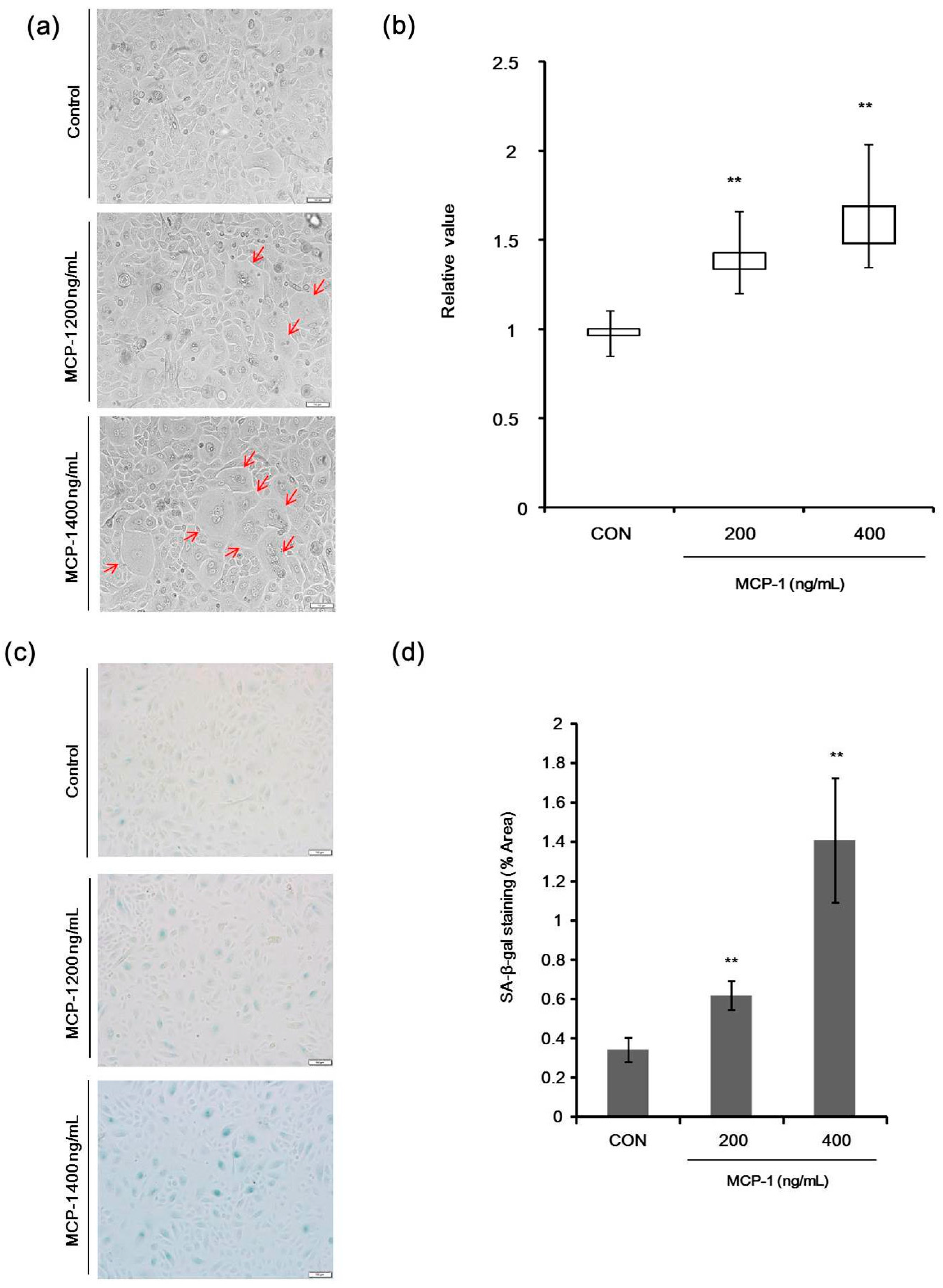
2.4. Morphological Changes and Senescence-Associated Beta-Galactosidase (SA-β-Gal) Staining in NHKs
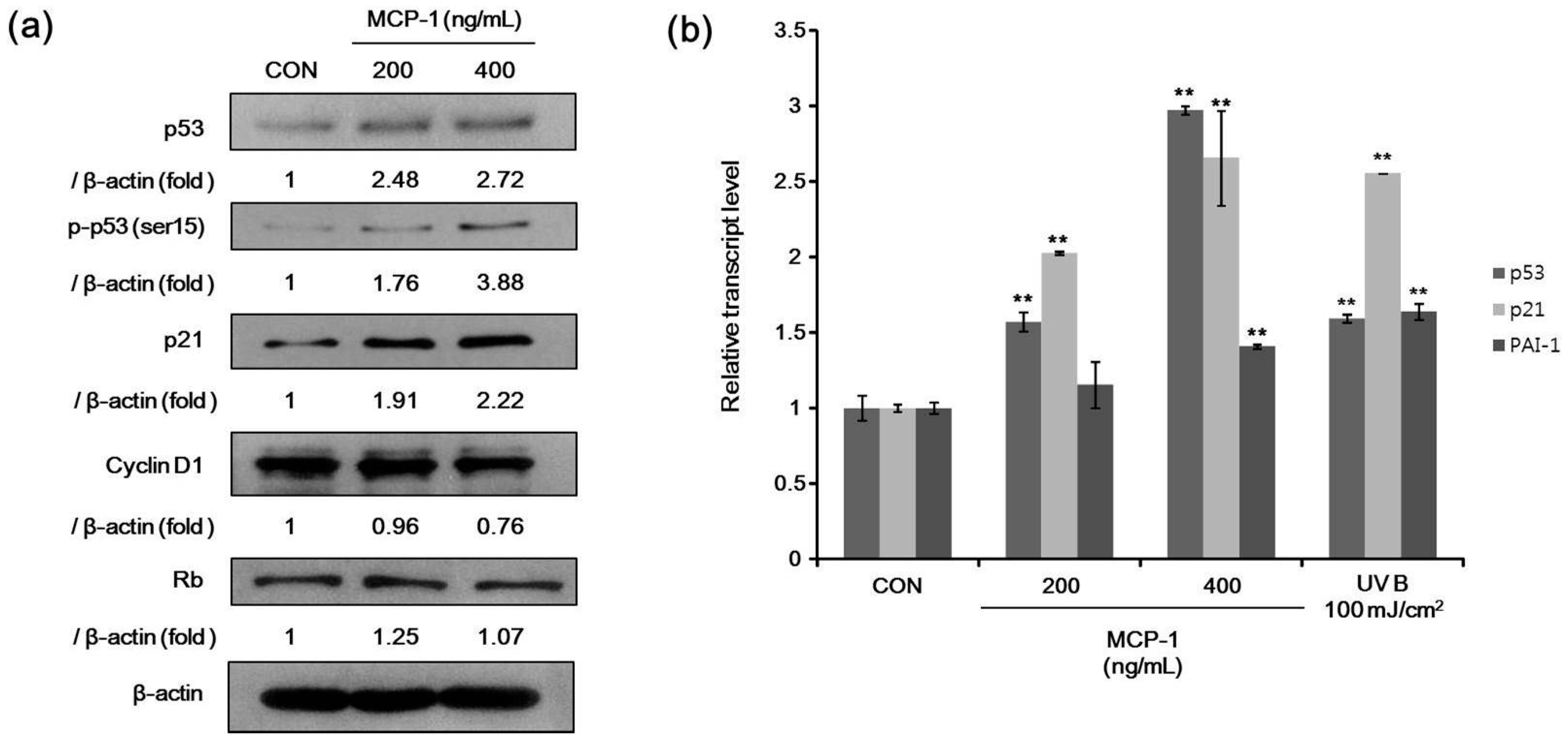
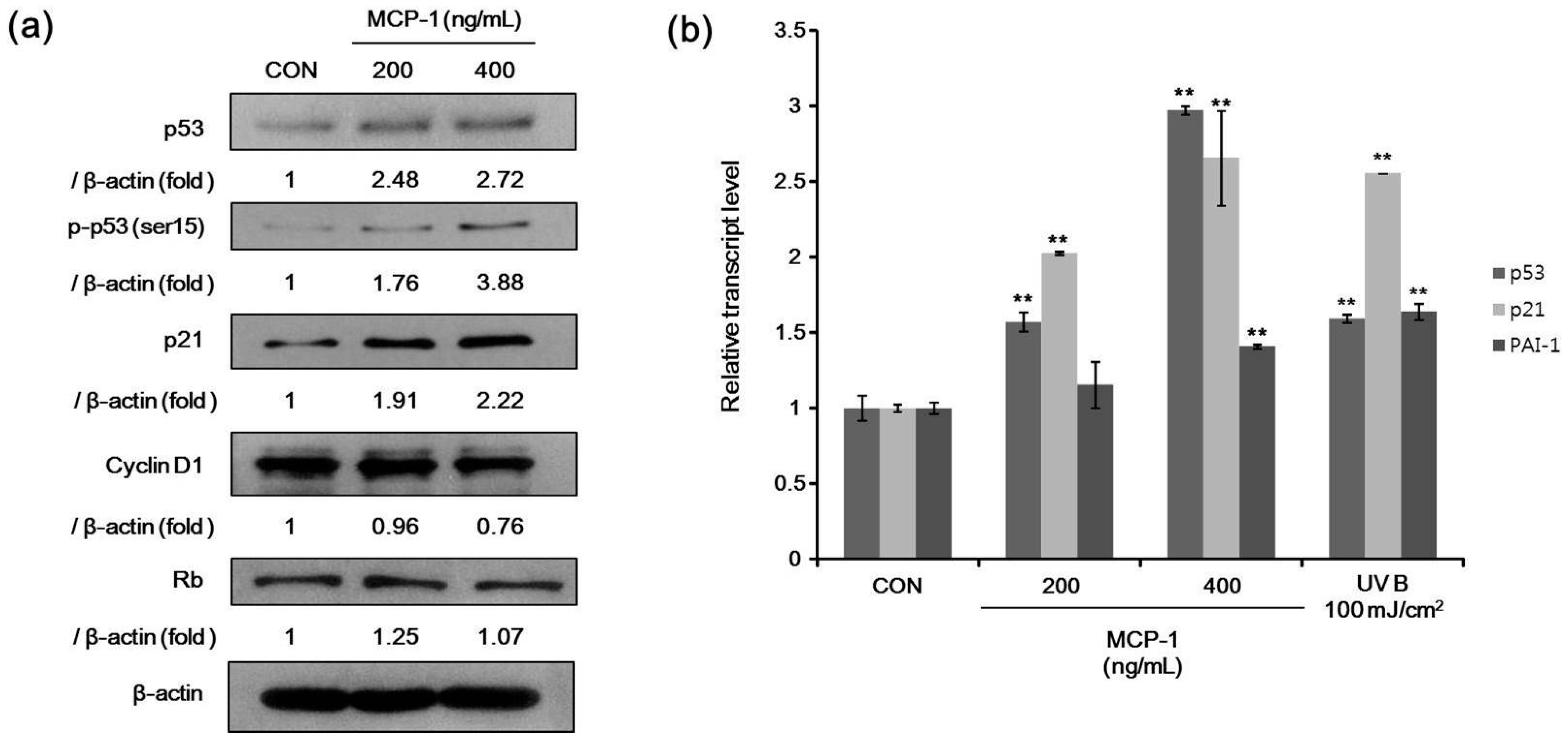
2.5. Senescence-Associated Markers in NHKs
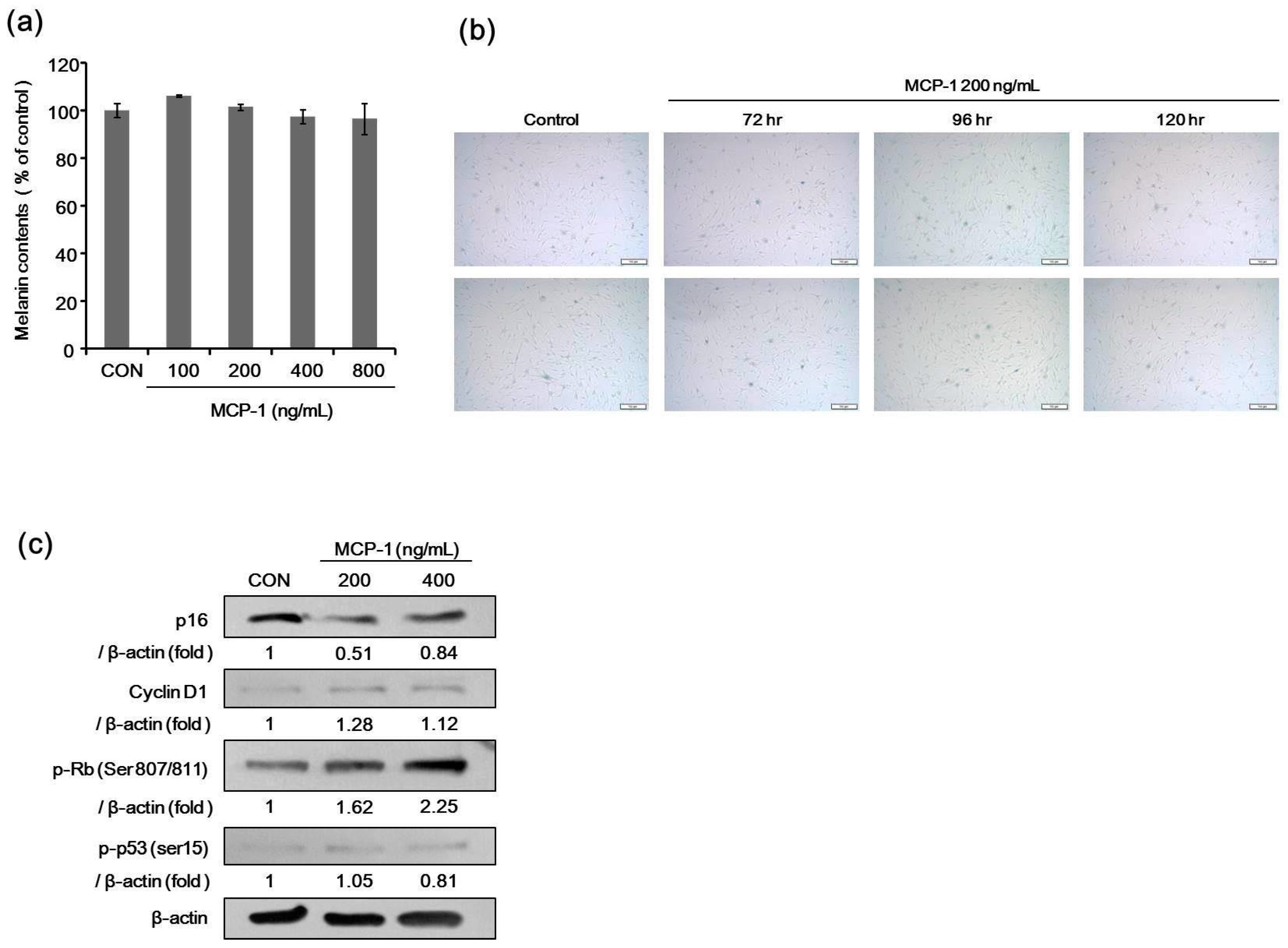
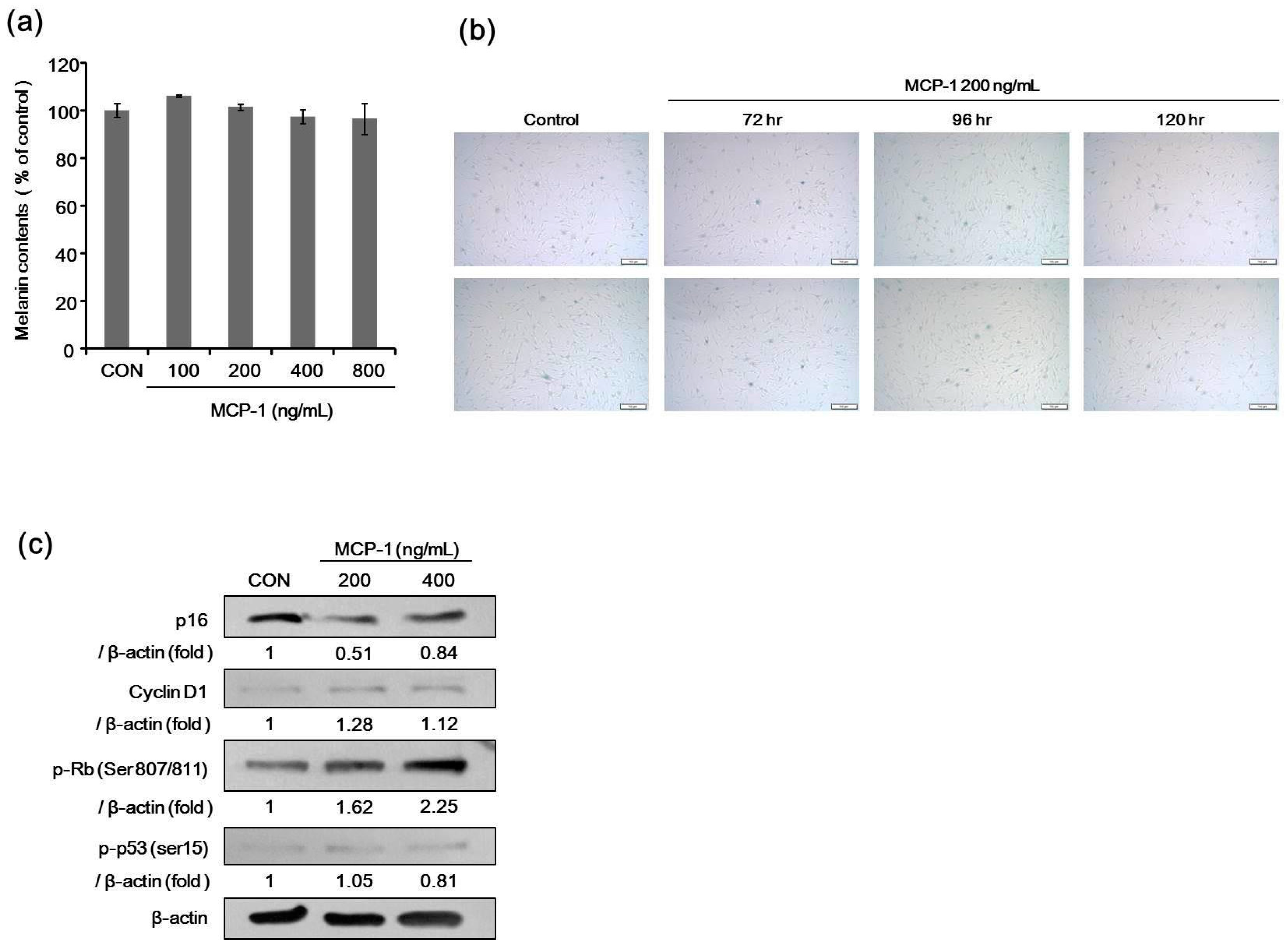
2.6. Melanin Contents and SA-β-Gal Staining after Treatment with MCP-1 in NHMs
2.7. Expression of Proliferation Markers in NHMs Treated with MCP-1
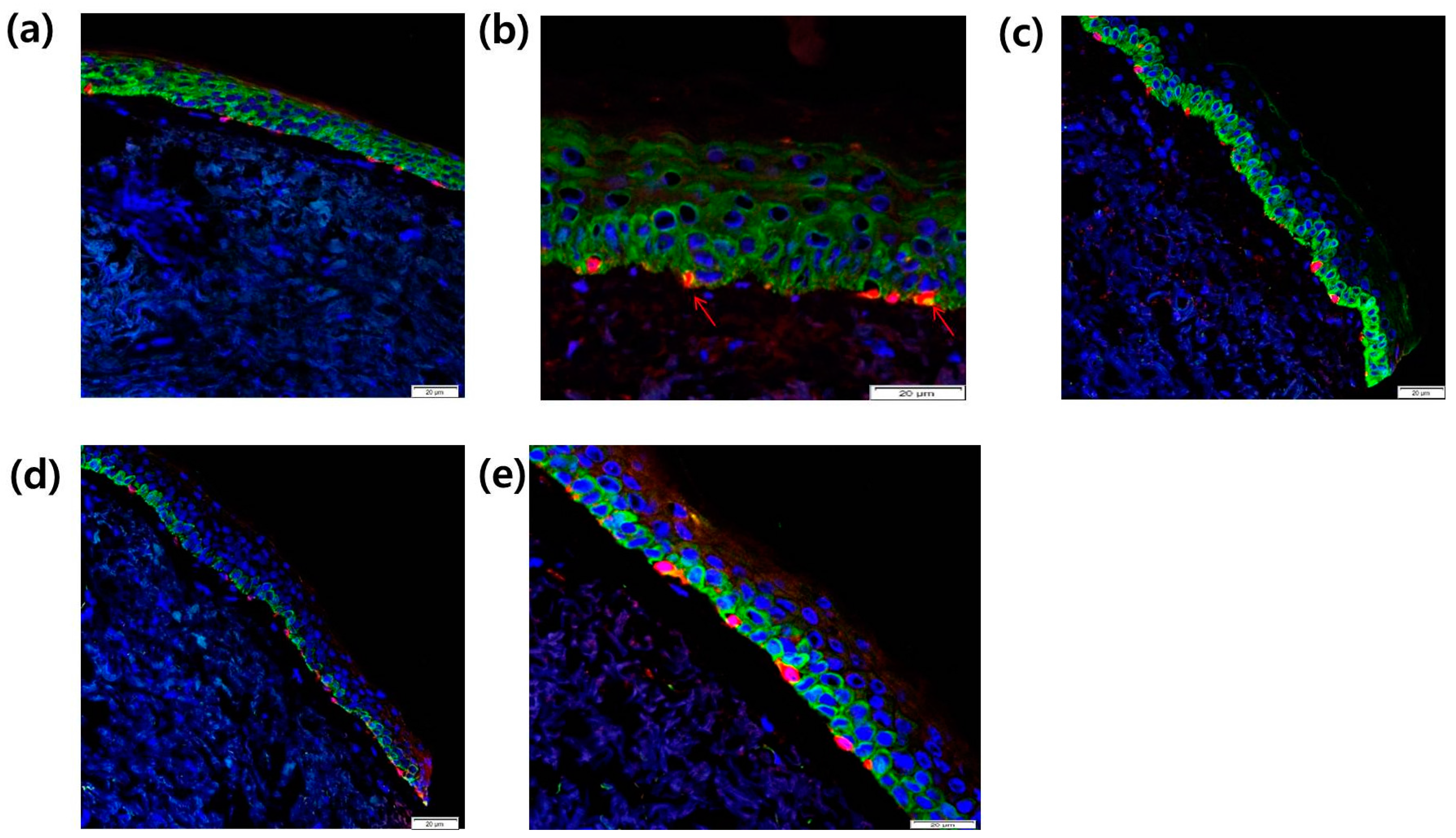
2.8. Intra-Epidermal Distribution of CCR2 and Rb in SL Tissue Evaluated by Confocal Laser Microscopy
3. Discussion
4. Materials and Methods
4.1. Reagent and Antibodies
4.2. Cell Culture
4.3. Cell Proliferation Assay
4.4. Melanin Content Assay
4.5. SA-β-Gal Staining
4.6. Western Blotting
4.7. Real-Time RT-PCR
4.8. Immunohistochemistry
4.9. Immunofluorescence Double Staining
4.10. Statistical Analysis
Author Contributions
Conflicts of Interest
Abbreviations
| SL | solar lentigo |
| MCP | monocyte chemoattractant protein |
| UVR | ultraviolet radiation |
| NHMs | normal human melanocytes |
| NHKs | normal human keratinocytes |
| SASP | senescence-associated secretory phenotype |
| IL | interleukin |
| SA-β-Gal | senescence-associated beta-galactosidase |
| PBS | phosphate-buffered saline |
| ELISA | enzyme immunosorbent assay |
| OD | optical density |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| TNF | tumor necrosis factor |
| LSM | laser scanning microscope |
| PAI-1 | plasminogen activator inhibitor-1 |
| POMC | proopiomelanocortin |
| CRH | corticotropin releasing hormone |
| ACTH | adrenocorticotropic hormone |
| MSH | melanocyte-stimulating hormone |
References
- Noh, T.K.; Chung, B.Y.; Yeo, U.C.; Chang, S.; Lee, M.W.; Chang, S.E. Q-Switched 660-nm versus 532-nm Nd: YAG laser for the treatment for facial lentigines in asian patients: A prospective, randomized, double-blinded, split-face comparison pilot study. Dermatol. Surg. 2015, 41, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, G.; Kovacs, D.; Picardo, M. Mechanisms underlying post-inflammatory hyperpigmentation: Lessons from solar lentigo. Ann. Dermatol. Venereol. 2012, 139, 148–152. [Google Scholar] [CrossRef]
- Kovacs, D.; Cardinali, G.; Aspite, N.; Cota, C.; Luzi, F.; Bellei, B.; Briganti, S.; Amantea, A.; Torrisi, M.R.; Picardo, M. Role of fibroblast-derived growth factors in regulating hyperpigmentation of solar lentigo. Br. J. Dermatol. 2010, 163, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Noh, T.K.; Choi, S.J.; Chung, B.Y.; Kang, J.S.; Lee, J.H.; Lee, M.W.; Chang, S.E. Inflammatory features of melasma lesions in Asian skin. J. Dermatol. 2014, 41, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Barranco, V.P. Multiple benign lichenoid keratoses simulating photodermatoses: Evolution from senile lentigines and their spontaneous regression. J. Am. Acad. Dermatol. 1985, 13, 201–206. [Google Scholar] [CrossRef]
- Salducci, M.; André, N.; Guéré, C.; Martin, M.; Fitoussi, R.; Vié, K.; Cario-André, M. Factors secreted by irradiated aged fibroblasts induce solar lentigo in pigmented reconstructed epidermis. Pigment Cell Melanoma Res. 2014, 27, 502–504. [Google Scholar] [CrossRef] [PubMed]
- Andersen, W.K.; Labadie, R.R.; Bhawan, J. Histopathology of solar lentigines of the face: A quantitative study. J. Am. Acad. Dermatol. 1997, 36, 444–447. [Google Scholar] [CrossRef]
- Yamada, T.; Hasegawa, S.; Inoue, Y.; Date, Y.; Arima, M.; Yagami, A.; Iwata, Y.; Takahashi, M.; Yamamoto, N.; Mizutani, H.; et al. Accelerated differentiation of melanocyte stem cells contributes to the formation of hyperpigmented maculae. Exp. Dermatol. 2014, 23, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Hasegawa, S.; Inoue, Y.; Date, Y.; Arima, M.; Yagami, A.; Iwata, Y.; Abe, M.; Takahashi, M.; Yamamoto, N.; et al. Comprehensive analysis of melanogenesis and proliferation potential of melanocyte lineage in solar lentigines. J. Dermatol. Sci. 2014, 73, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T. Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigment Cell Res. 2005, 18, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G. Autocrine and paracrine regulation of melanocytes in human skin and in pigmentary disorders. Pigment Cell Res. 2004, 17, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.C.; O'Loghlen, A.; Banito, A.; Guijarro, M.V.; Augert, A.; Raguz, S.; Fumagalli, M.; da Costa, M.; Brown, C.; Popov, N.; et al. Chemokine signaling via the CXCR2 receptor reinforce senescence. Cell 2008, 133, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Kortlever, R.M.; Higgins, P.J.; Bernards, R. Plasminogen activator inhibitor-1 is a critical downstream target of p53 in the induction of replicative senescence. Nat. Cell Biol. 2006, 8, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M.; Dewald, B.; Moser, B. Human chemokines: An update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Jayaraman, V.; Huelsmann, E.J.; Bonish, B.; Burgad, D.; Sivaramakrishnan, G.; Qin, S.; DiPietro, L.A.; Zloza, A.; Zhang, C.; et al. Pro-inflammatory chemokine CCL2 (MCP-1) promotes healing in diabetic wounds by restoring the macrophage response. PLoS ONE 2014, 9, e91574. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.M.; Sharma, M.R.; Werth, V.P. UVB and proinflammatory cytokines synergistically activate TNF-α productionin keratinocytes through enhanced gene transcription. J. Investig. Dermatol. 2009, 129, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- WalderaLupa, D.M.; Kalfalah, F.; Safferling, K.; Boukamp, P.; Poschmann, G.; Volpi, E.; Götz-Rösch, C.; Bernerd, F.; Haag, L.; Huebenthal, U.; et al. Characterization of skin aging-associated secreted proteins (SAASP) produced by dermal fibroblasts isolated from intrinsically aged human skin. J. Investig. Dermatol. 2015, 135, 1954–1968. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Kim, H.N.; Jung da, J.; Kim, J.E.; Mun, G.H.; Kim, Y.S.; Cho, D.; Shin, D.H.; Hwang, Y.I.; Lee, W.J. Regulation of UVB-induced IL-8 and MCP-1 production in skin keratinocytes by increasing vitamin C uptake via the redistribution of SVCT-1 from the cytosol to the membrane. J. Investig. Dermatol. 2007, 127, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Boruah, D.; Manu, V.; Malik, A. A morphometric and immunohistochemical study of melanocytes in periorbital hyperpigmentation. Indian. J. Dermatol. Venereol. Leprol. 2015, 81, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E.; Niu, J. Inflammation, ER stress, autophagy and MCP-1/CCR2 pathway. Circ. Res. 2012, 110, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Hu, Y.; Li, W.H.; Eisinger, M.; Seiberg, M.; Lin, C.B. The role of keratinocyte growth factor in melanogenesis: A possible mechanism for the initiation of solar lentigines. Exp. Dermatol. 2010, 19, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Moro, O. Upregulation of the IFN-γ-stimulated genes in the development of delayed pigmented spots on the dorsal skin of F1 mice of HR-1 × HR/De. J. Investig. Dermatol. 2005, 124, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Han, J.; Kim, K.; Choi, H.; Cho, E.G.; Lee, T.R. CXCR7 mediates SDF-1 induced melanocyte migration. Pigment Cell Melanoma Res. 2013, 26, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell. Biol. 2012, 212, 1–115. [Google Scholar]
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Luger, T.; Paus, R.; Solomon, S. Corticotropin releasing hormone and proopiomelanocortin involvement in the cutaneous response to stress. Physiol. Rev. 2000, 80, 979–1020. [Google Scholar] [PubMed]
- Rousseau, K.; Kauser, S.; Pritchard, L.E.; Warhurst, A.; Oliver, R.L.; Slominski, A.; Wei, E.T.; Thody, A.J.; Tobin, D.J.; White, A.; et al. Proopiomelanocortin (POMC), the ACTH/melanocortin precursor, is secreted by human epidermal keratinocytes and melanocytes and stimulates melanogenesis. FASEB J. 2007, 21, 1844–1856. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. l-tyrosine and l-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Kai, H.; Egami, K.; Murohara, T.; Ikeda, A.; Yasuoka, S.; Egashira, K.; Matsuishi, T.; Kai, M.; Kataoka, Y.; et al. Mutant MCP-1 therapy inhibits tumor angiogenesis and growth of malignant melanoma in mice. Biochem. Biophys. Res. Commun. 2008, 365, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Adini, I.; Adini, A.; Bazinet, L.; Watnick, R.S.; Bielenberg, D.R.; D'Amato, R.J. Melanocyte pigmentation inversely correlates with MCP-1 production and angiogenesis-inducing potential. FASEB J. 2015, 29, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.H.; Lee, H.J.; Heo, J.; Lim, J.; Kim, M.; Kim, M.K.; Nam, H.Y.; Hong, G.H.; Cho, Y.S.; Choi, S.J.; et al. Senescence associated MCP-1 secretion is dependent on a decline in BMI1 in human mesenchymal stromal cells. Antioxid. Redox. Signal. 2016, 24, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.C.; O'Loghlen, A.; Bantio, A.; Raguz, S.; Gil, J. Control of senescence b CXCR2 and its ligands. Cell Cycle 2008, 19, 2956–2959. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age (Years) | Biopsy Site | Epidermal Thickness (μm) | Rete Ridge | No. of Keratinocytes (Per One Melanocyte) | Increment of Melanocytes |
|---|---|---|---|---|---|---|---|
| 1 | F | 53 | cheek | 47 | Normal | 4 | + |
| 2 | F | 59 | cheek | 65 | Elongated | 10 | − |
| 3 | F | 58 | nose | 60 | Flattened | 4 | + |
| 4 | F | 66 | cheek | 125 | Flattened | 6 | + |
| 5 | F | 56 | cheek | 110 | Flattened | 9 | − |
| 6 | F | 75 | cheek | 55 | Normal | 6 | + |
| 7 | F | 57 | cheek | 35 | Normal | 9 | − |
| 8 | F | 66 | temple | 42 | Flattened | 11 | − |
| 9 | F | 58 | temple | 55 | Flattened | 10 | − |
| 10 | F | 58 | nose | 60 | Flattened | 5.5 | + |
| 11 | F | 60 | upper eyelid | 100 | Normal | 4.5 | + |
| 12 | F | 67 | cheek | 43 | Normal | 4 | + |
| 13 | F | 60 | nose | 47 | Flattened | 7.5 | − |
| 14 | F | 61 | cheek | 85 | Normal | 4.5 | + |
| 15 | F | 58 | cheek | 95 | Flattened | 4 | + |
| 16 | F | 55 | cheek | 70 | Flattened | 3 | + |
| 17 | F | 62 | nose | 45 | Elongated | 4 | + |
| 18 | F | 55 | nose | 40 | Normal | 7 | − |
| 19 | F | 54 | cheek | 35 | Flattened | 5 | + |
| 20 | F | 50 | forehead | 65 | Elongated | 10 | − |
| 21 | F | 64 | cheek | 47 | Normal | 3 | + |
| mean | 59.6 | - | 63.1 | - | 6.3 | - | |
| % positivity | 13/21 (62%) | ||||||
| Name | Forward | Reverse |
|---|---|---|
| p53 (120 b.p) | TGAGGTTGGCTCTGACTGTA | TTACCACTGGAGTCTTCCAG |
| p21 (113 b.p) | ACAGCAGAGGAAGACCATGTGGACC | CGTTTTCGACCCTGAGAGTCTCCAG |
| PAI-1 (150 b.p) | TGAAGATCGAGGTGAACGAG | GGTCATGTTGCCTTTCCAGT |
| GAPDH | CCCATCACCATCTTCCAGGAG | GTTGTCATGGATGACCTTGGC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, W.J.; Jo, S.Y.; Lee, M.H.; Won, C.H.; Lee, M.W.; Choi, J.H.; Chang, S.E. The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo. Int. J. Mol. Sci. 2016, 17, 948. https://doi.org/10.3390/ijms17060948
Lee WJ, Jo SY, Lee MH, Won CH, Lee MW, Choi JH, Chang SE. The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo. International Journal of Molecular Sciences. 2016; 17(6):948. https://doi.org/10.3390/ijms17060948
Chicago/Turabian StyleLee, Woo Jin, Soo Youn Jo, Mi Hye Lee, Chong Hyun Won, Mi Woo Lee, Jee Ho Choi, and Sung Eun Chang. 2016. "The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo" International Journal of Molecular Sciences 17, no. 6: 948. https://doi.org/10.3390/ijms17060948
APA StyleLee, W. J., Jo, S. Y., Lee, M. H., Won, C. H., Lee, M. W., Choi, J. H., & Chang, S. E. (2016). The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo. International Journal of Molecular Sciences, 17(6), 948. https://doi.org/10.3390/ijms17060948