
12-Lipoxygenase Inhibition on Microalbuminuria in Type-1 and Type-2 Diabetes Is Associated with Changes of Glomerular Angiotensin II Type 1 Receptor Related to Insulin Resistance

Abstract
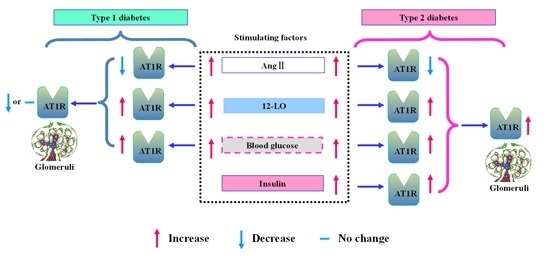
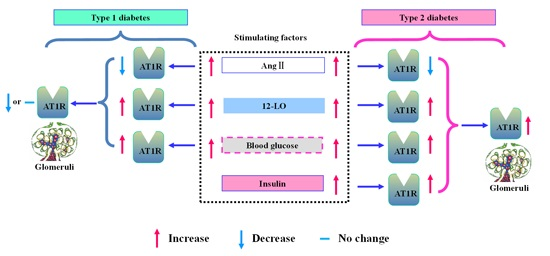{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
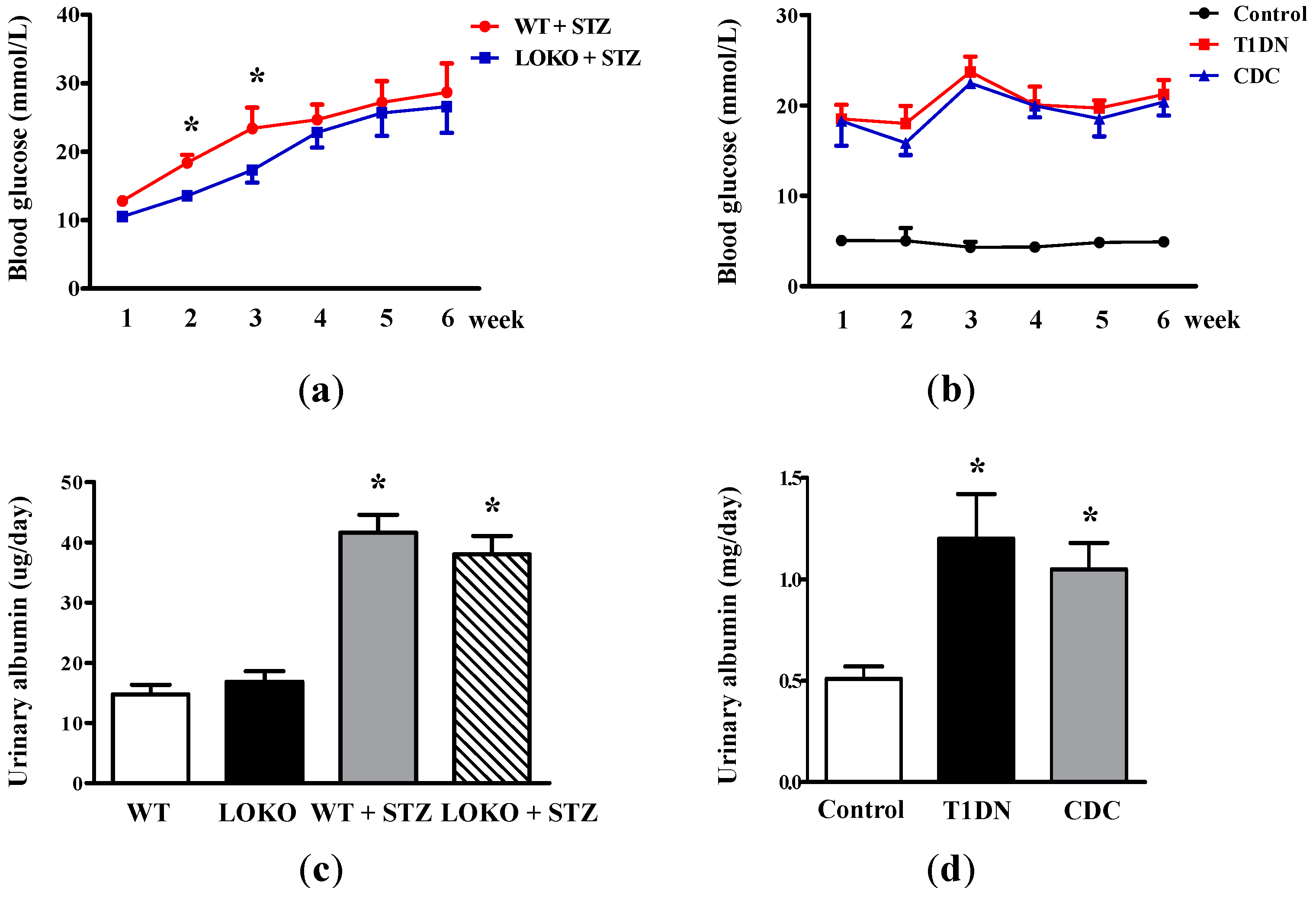
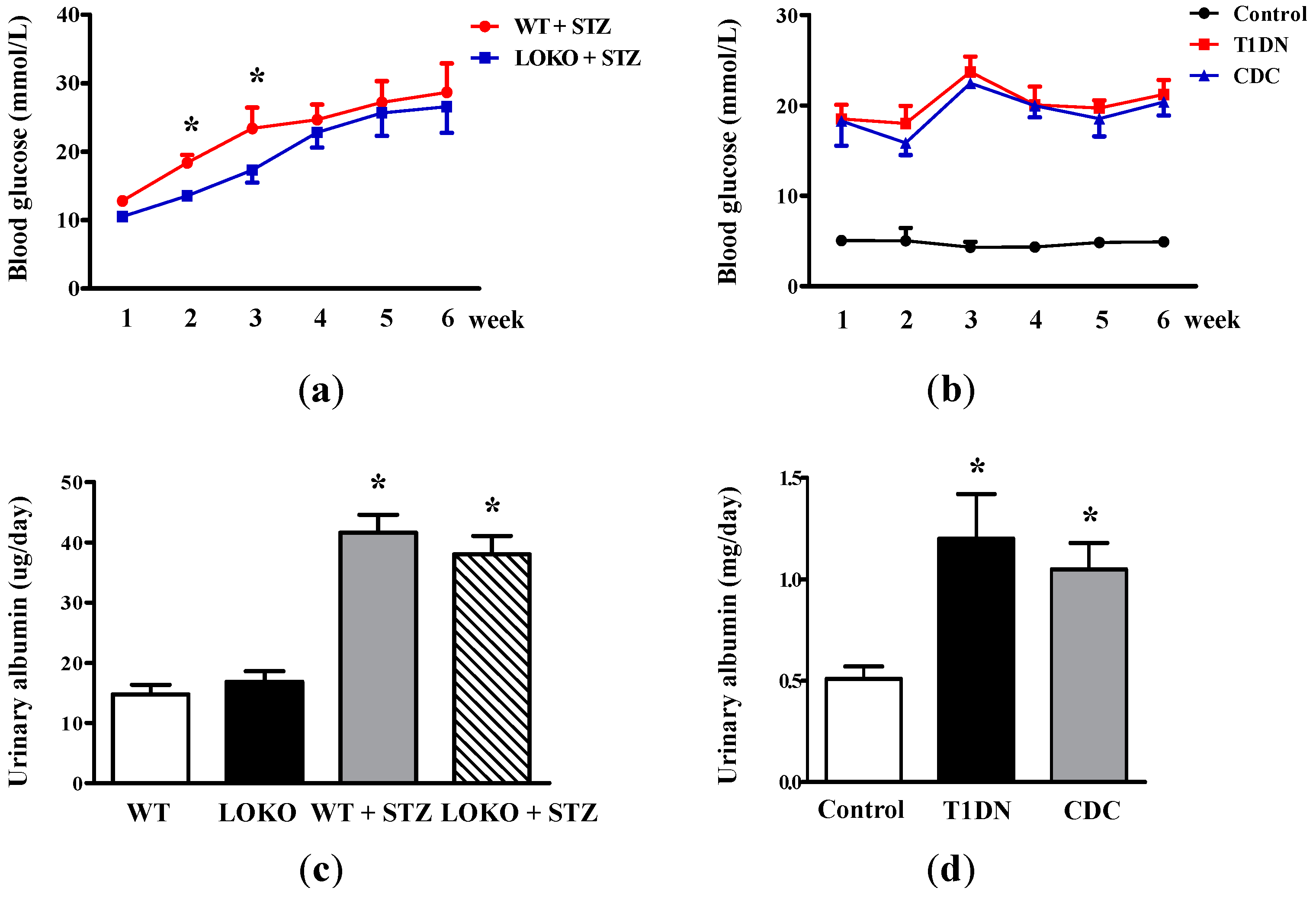
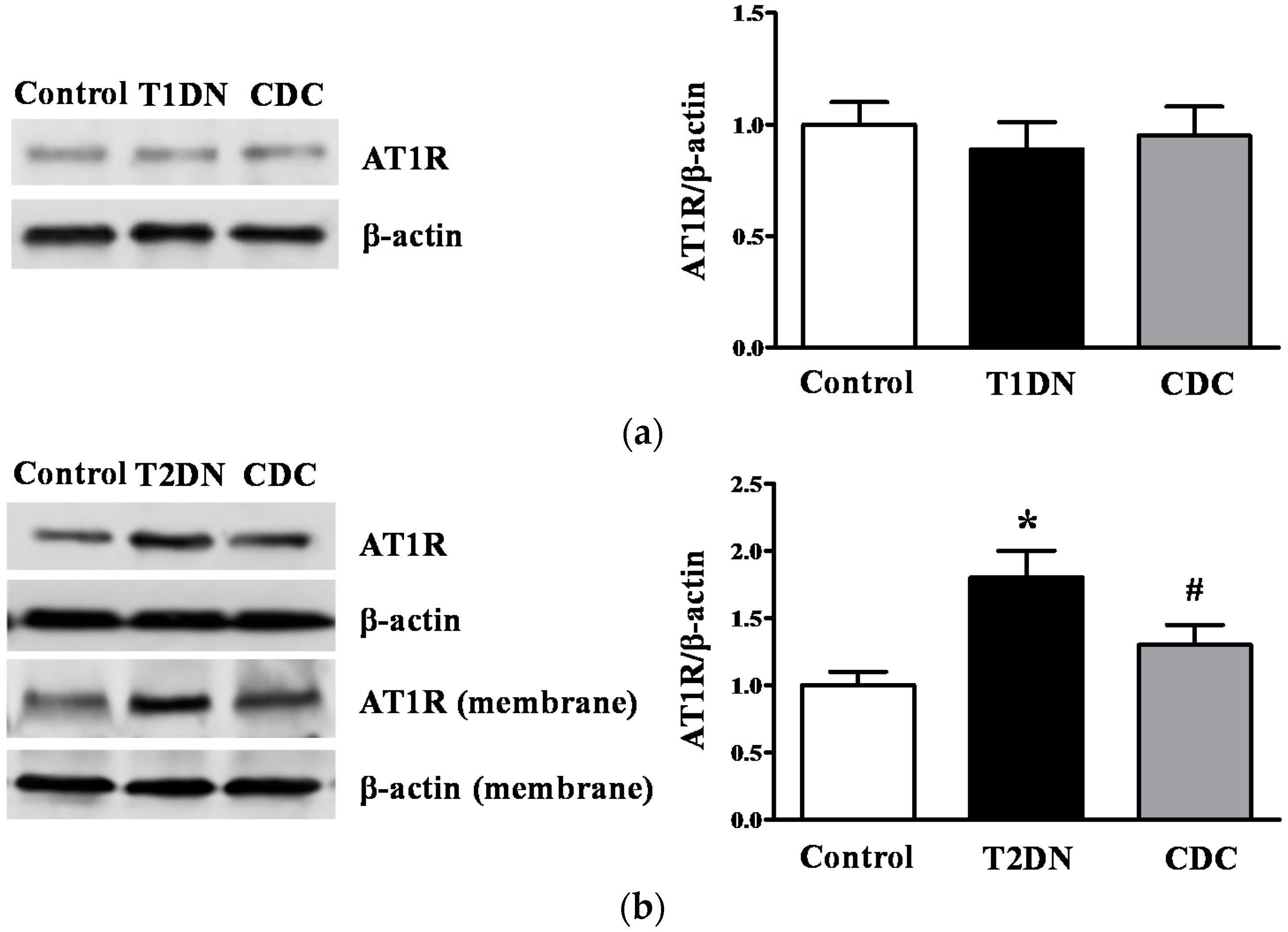
2.1. Roles of 12-LO on Fasting Blood Glucose and MAU in Type-1 Diabetic Models
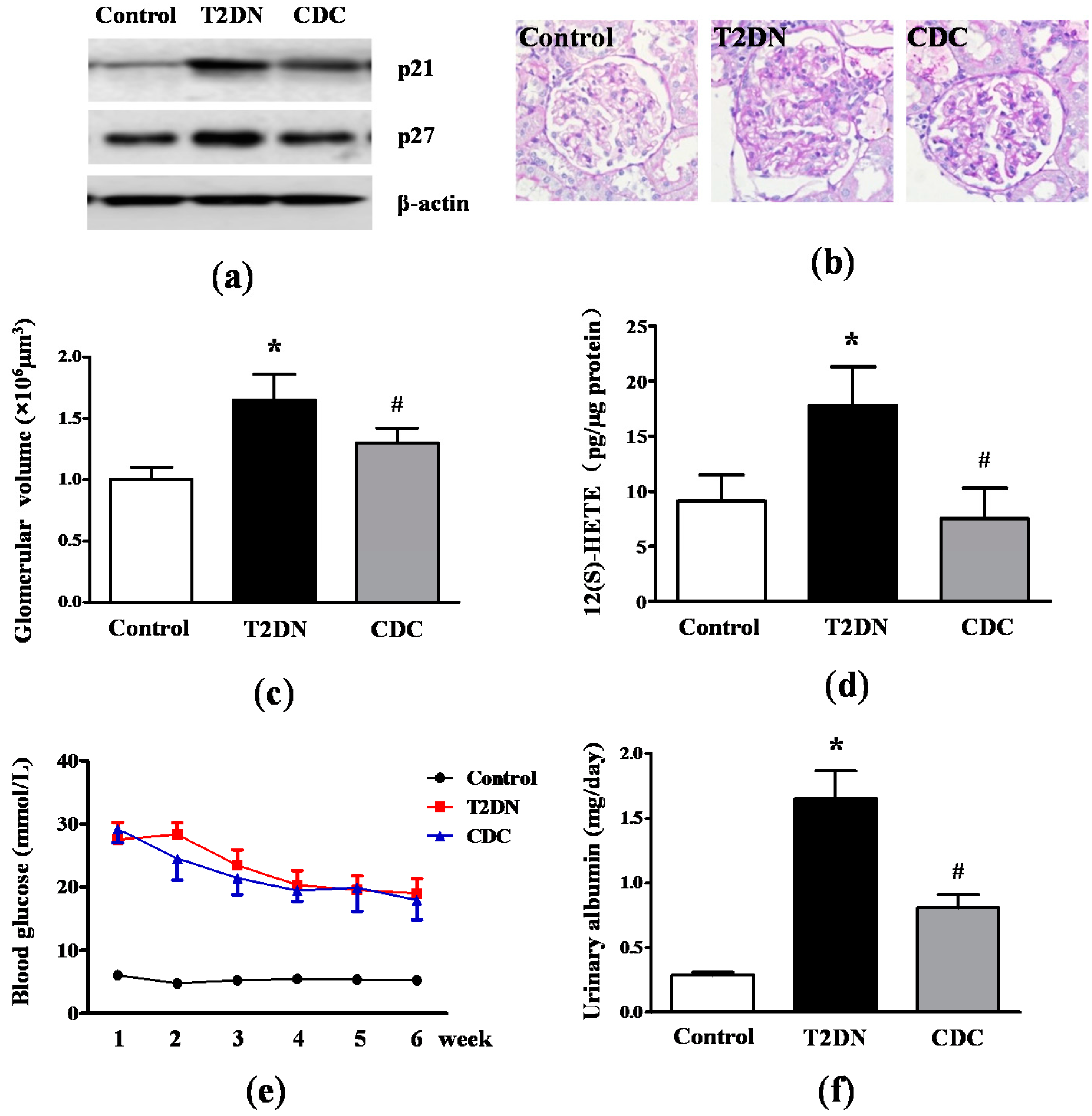
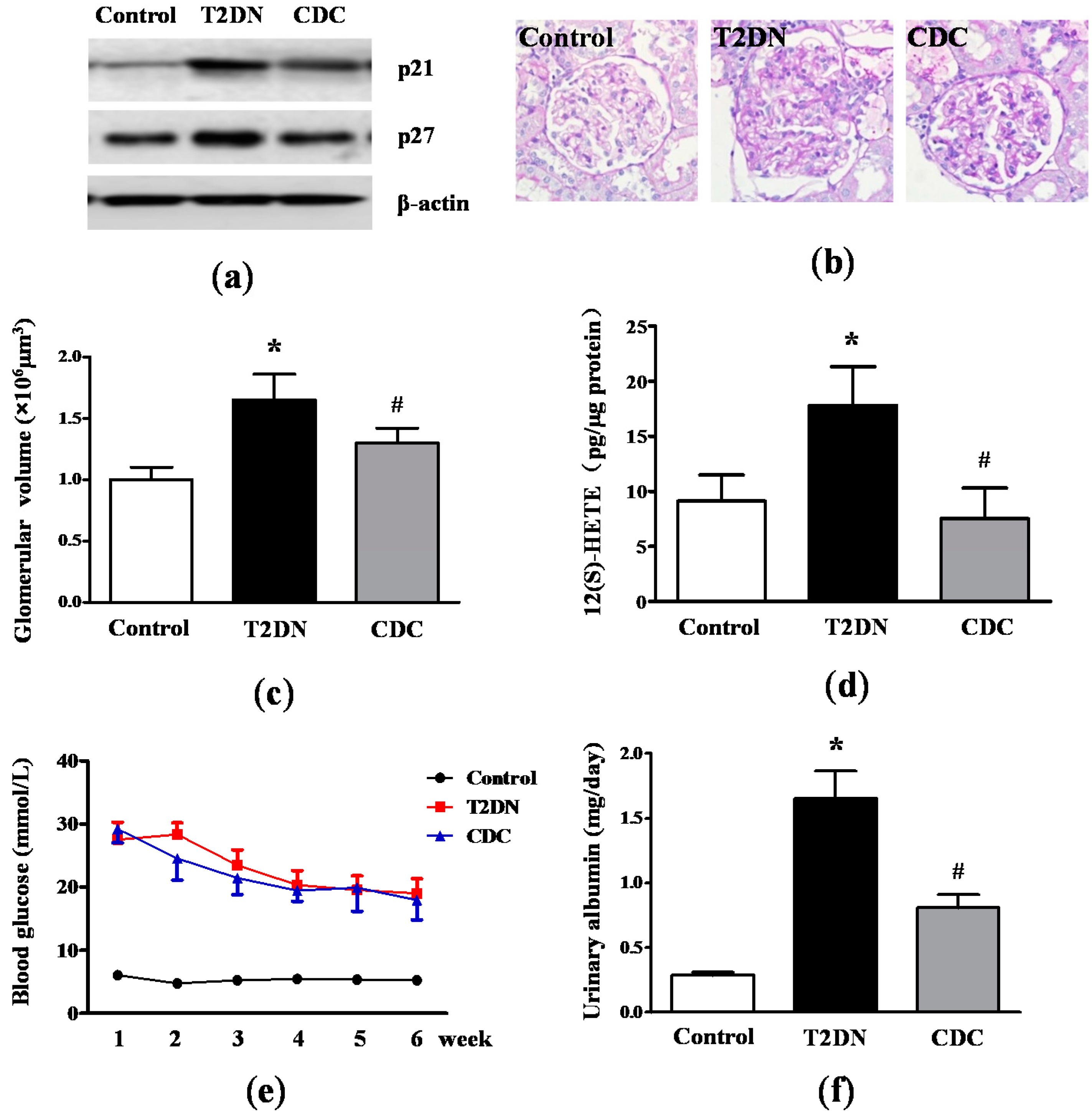
2.2. Kidney Hypertrophy, Glomerular 12(S)-HETE Levels, Fasting Blood Glucose and MAU in Type-2 Diabetic Rats
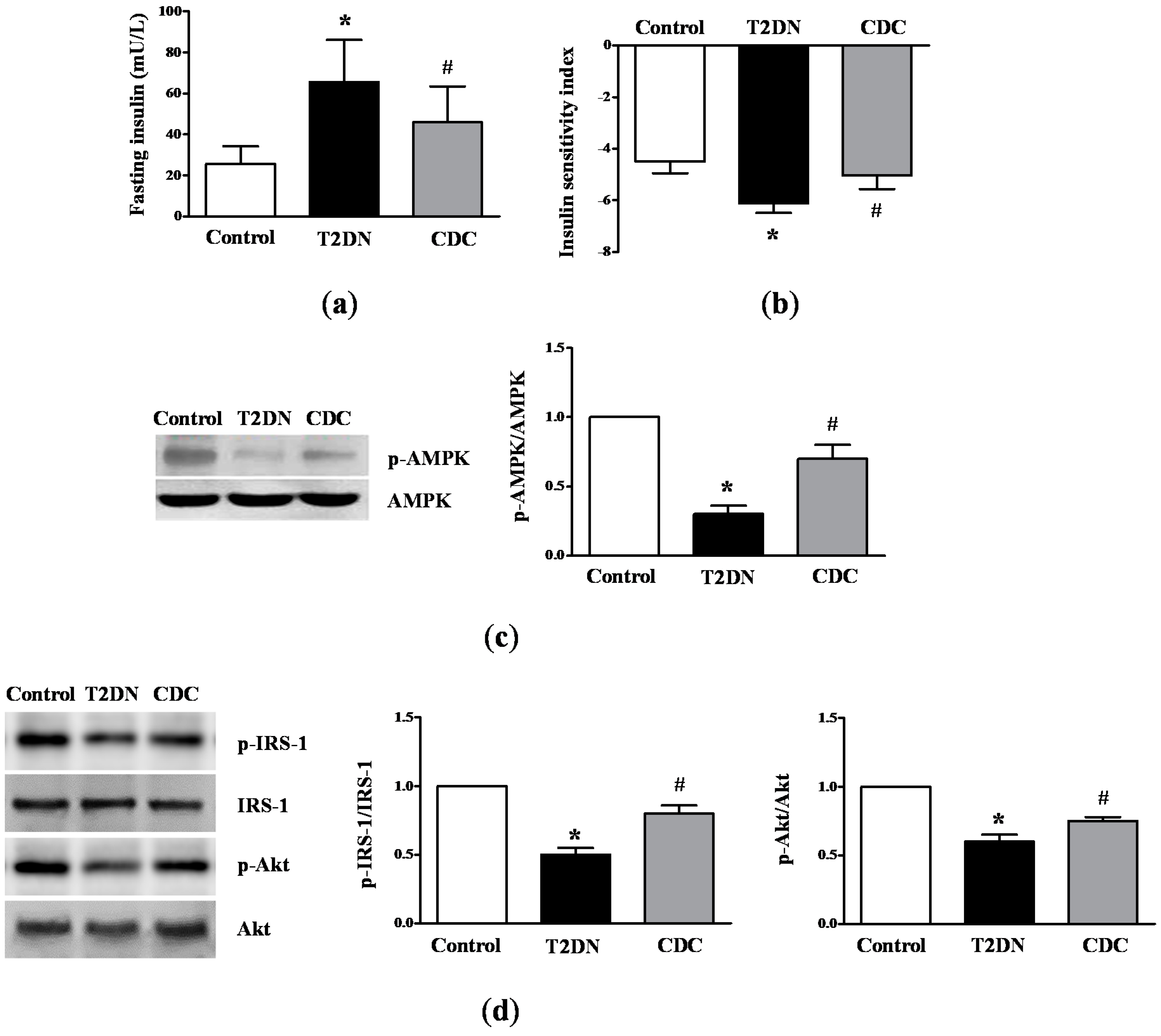
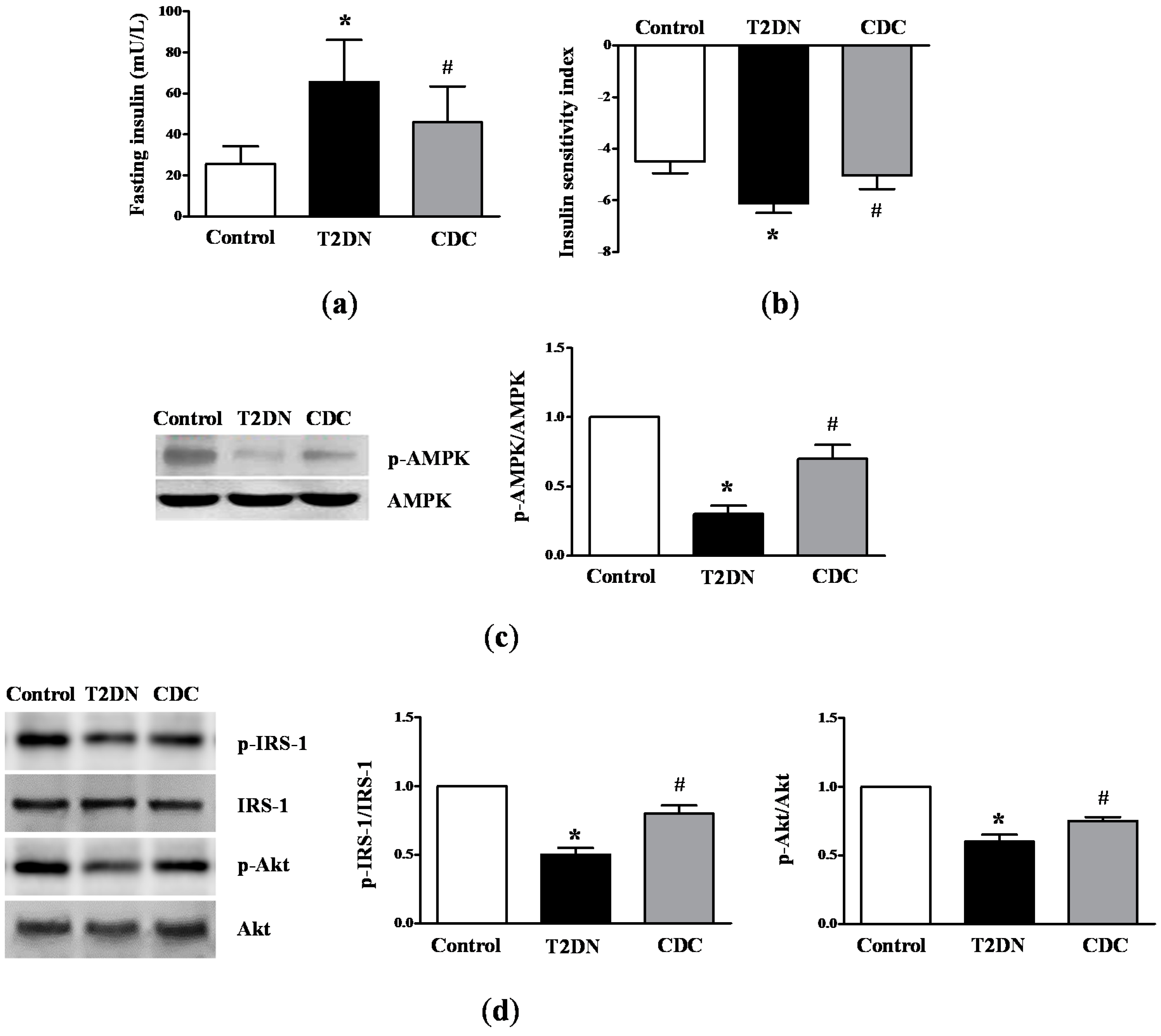
2.3. Roles of 12-LO on IR-Related Parameters in Type-2 Diabetic Rats
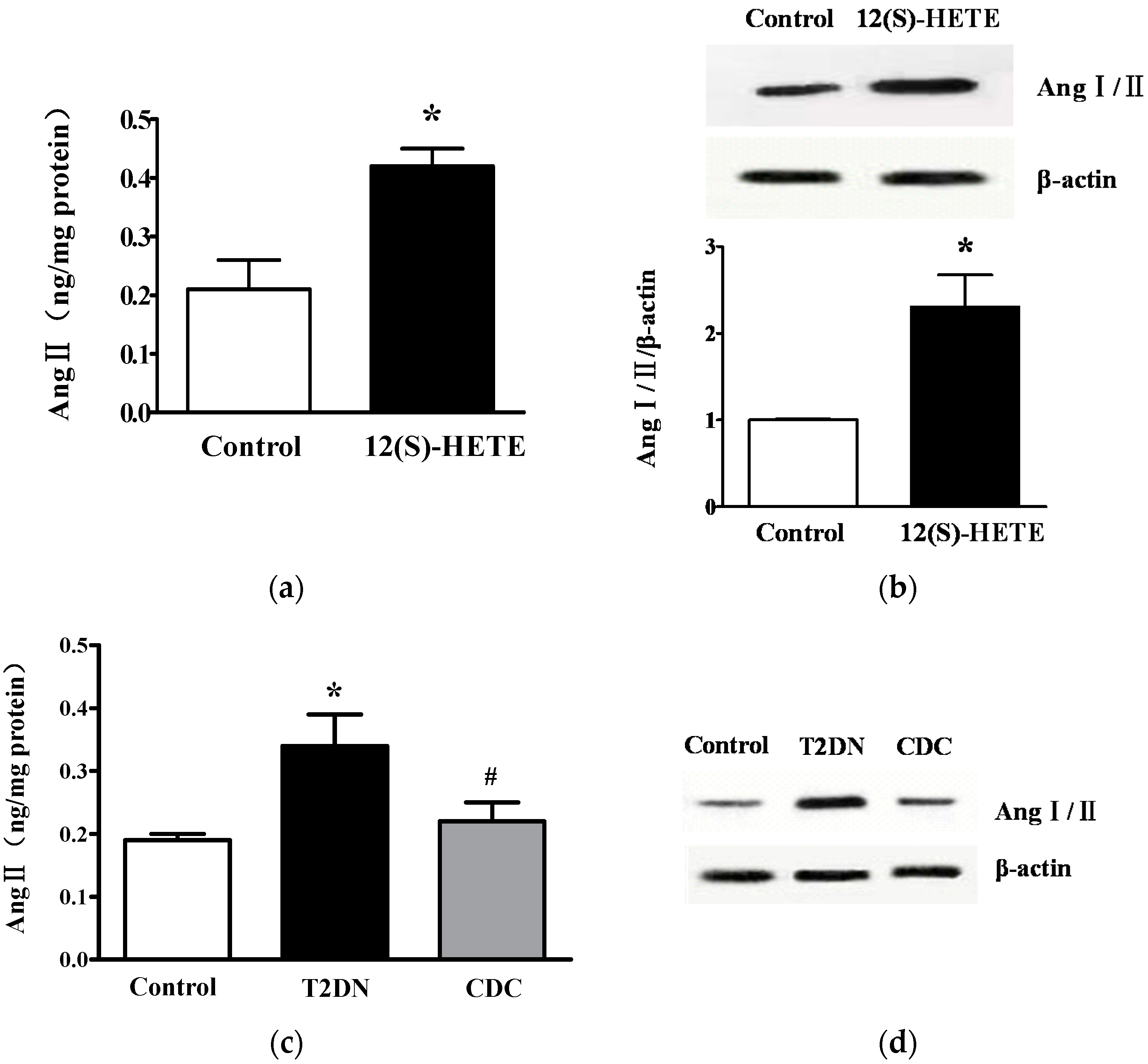
2.4. Effect of 12(S)-HETE on Rat Blood Glucose and Blood Pressure
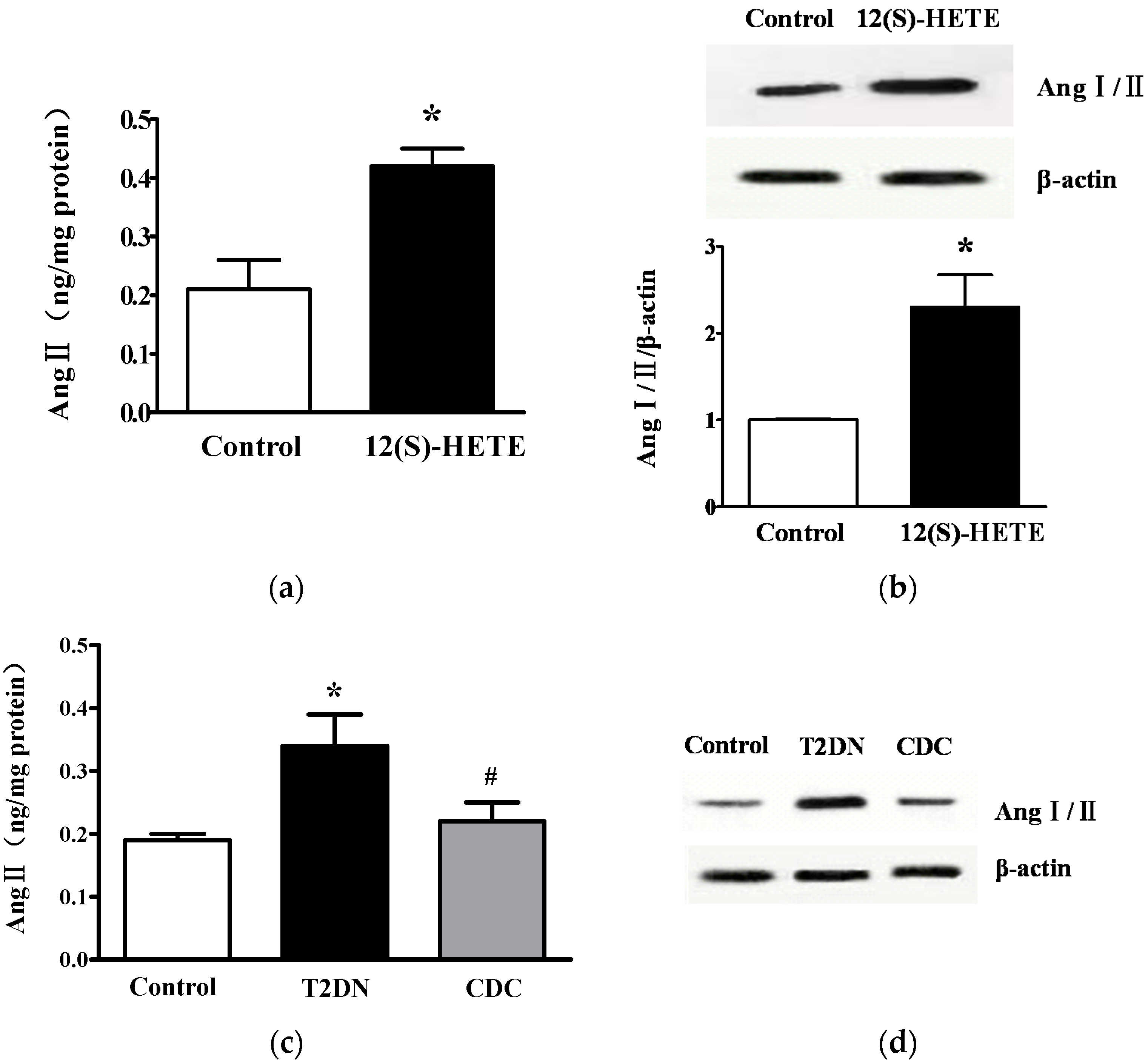
2.5. Effect of 12-LO on Ang II Level in Rat Glomeruli
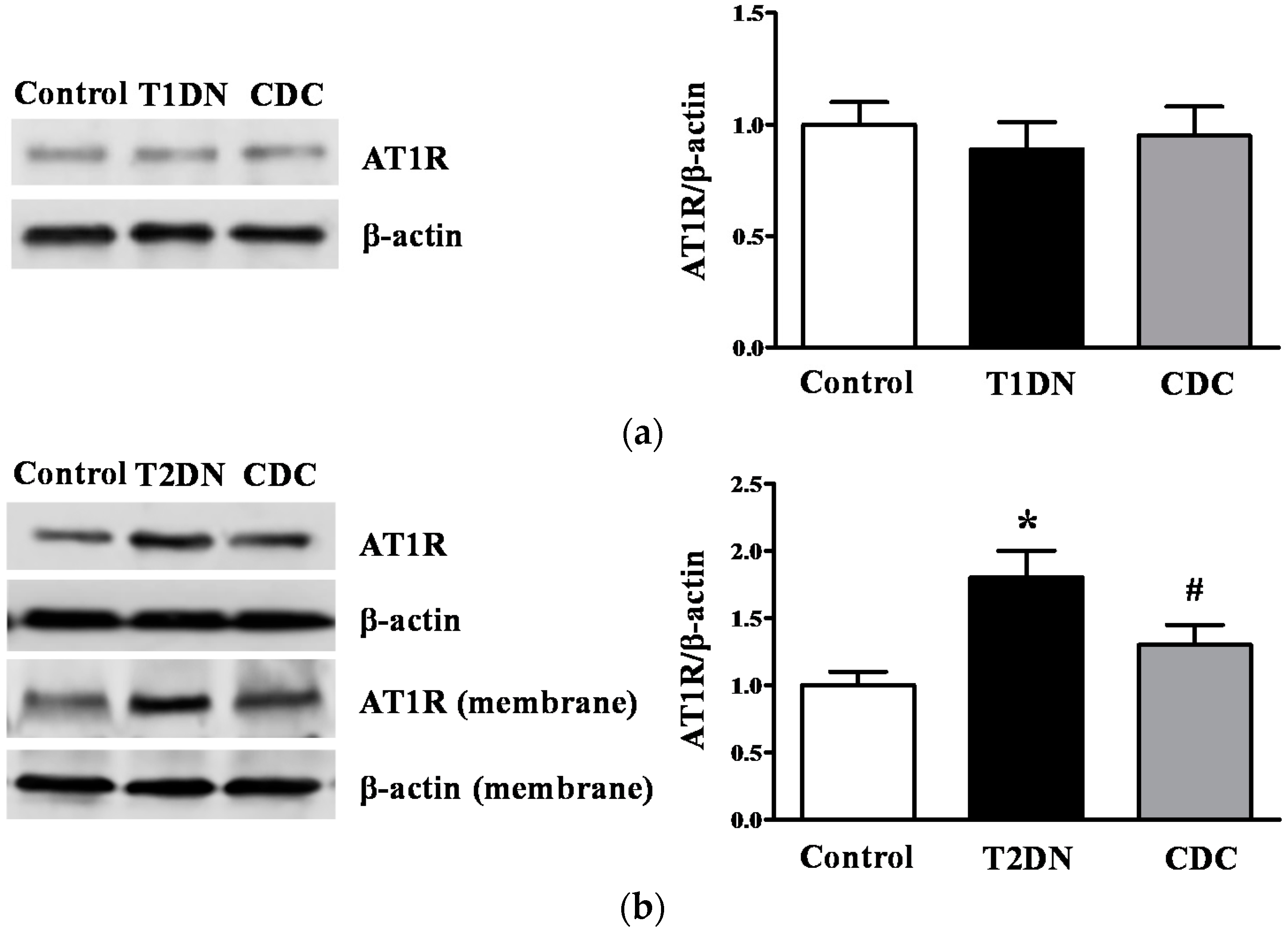
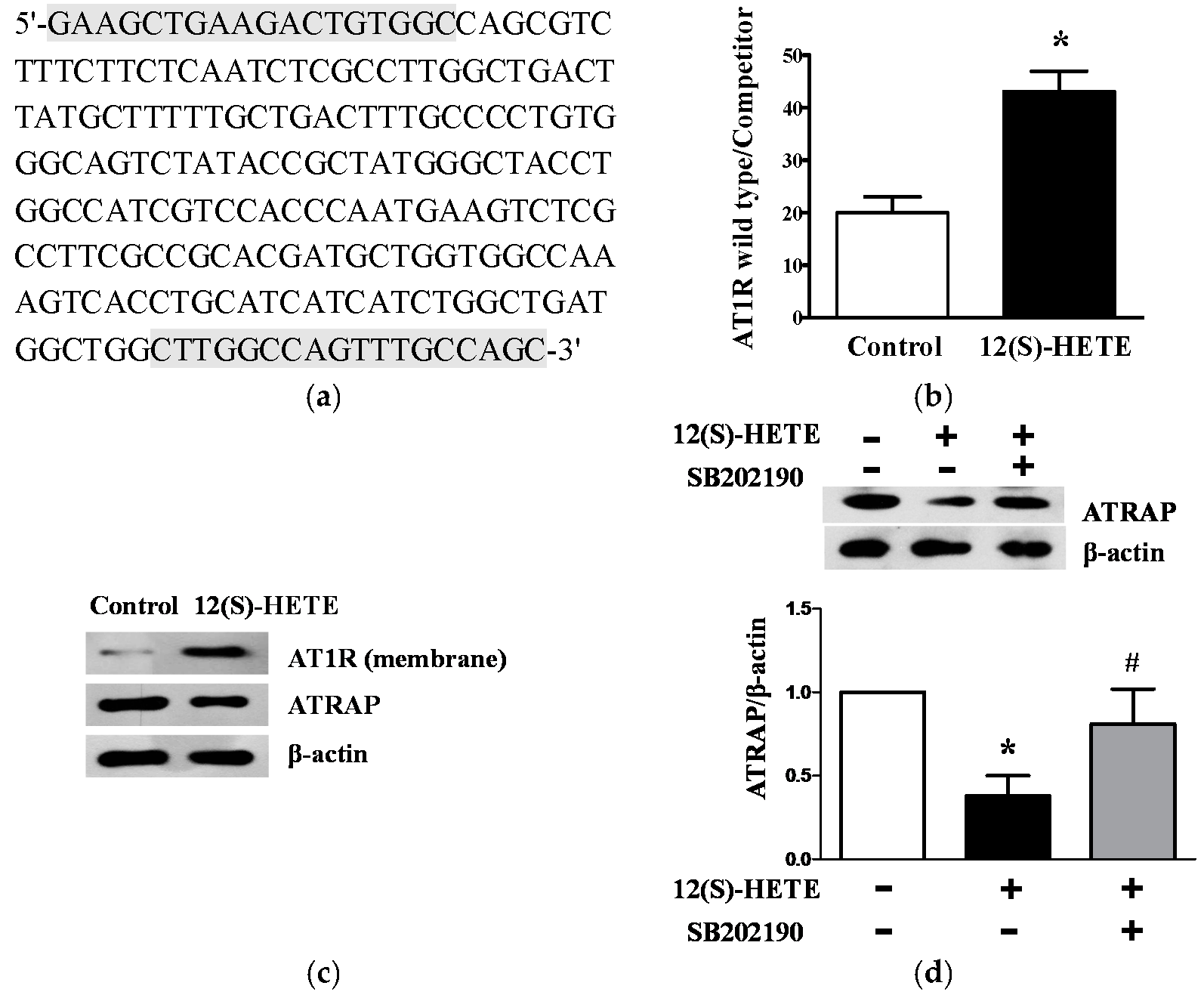
2.6. 12(S)-HETE Induced the Expression of AT1R and AT1R-Associated Protein in Rat Mesangial Cells and Glomeruli
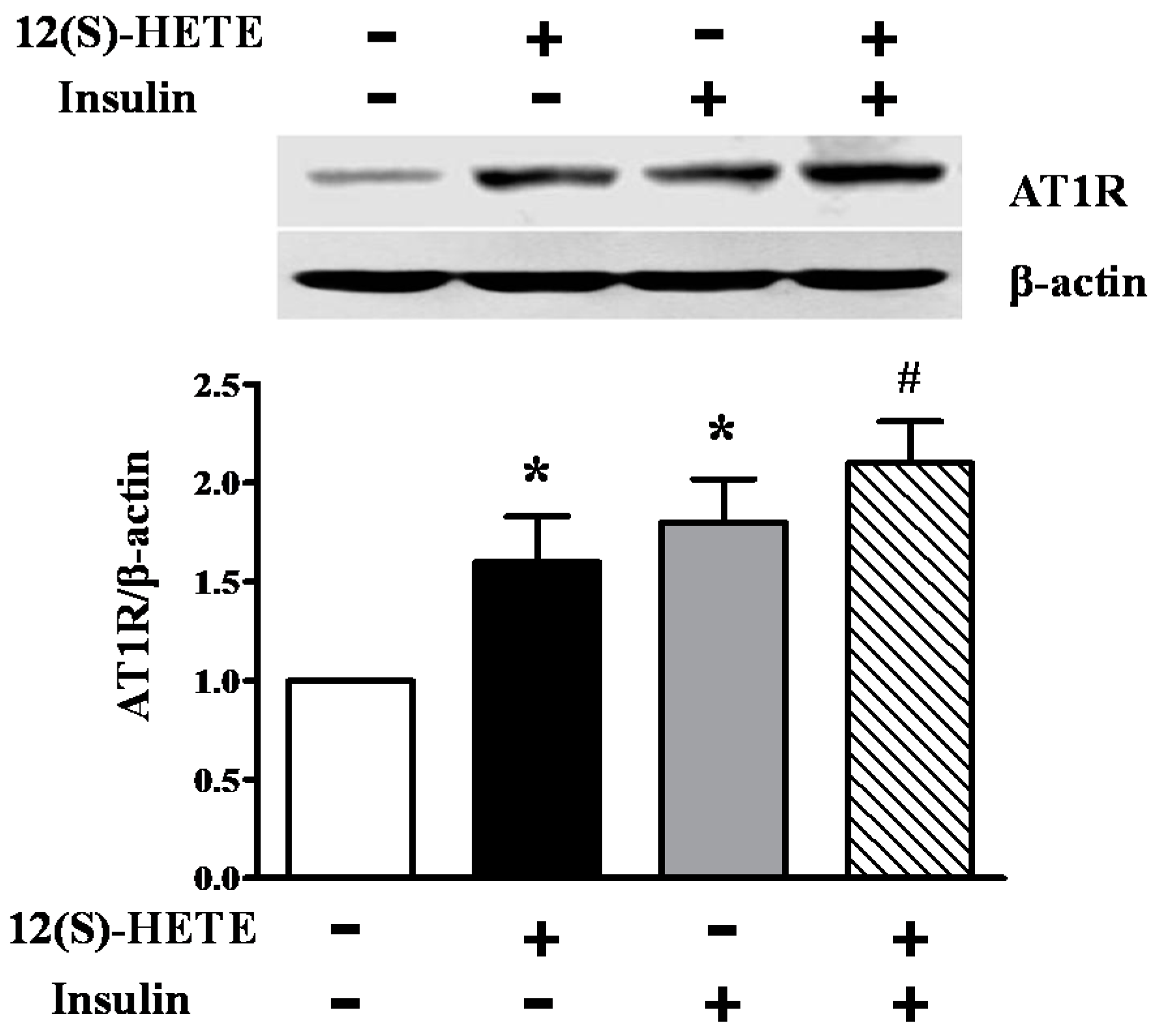
2.7. Effect of 12(S)-HETE and Insulin on AT1R Expression in Rat MCs
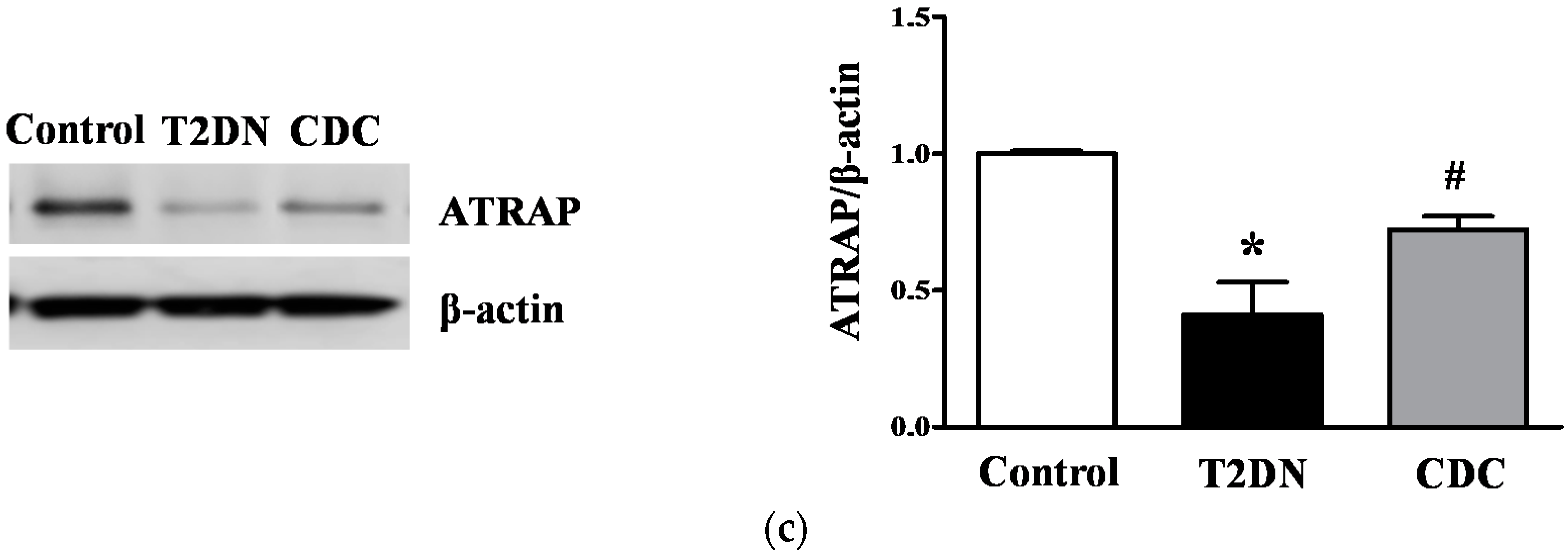
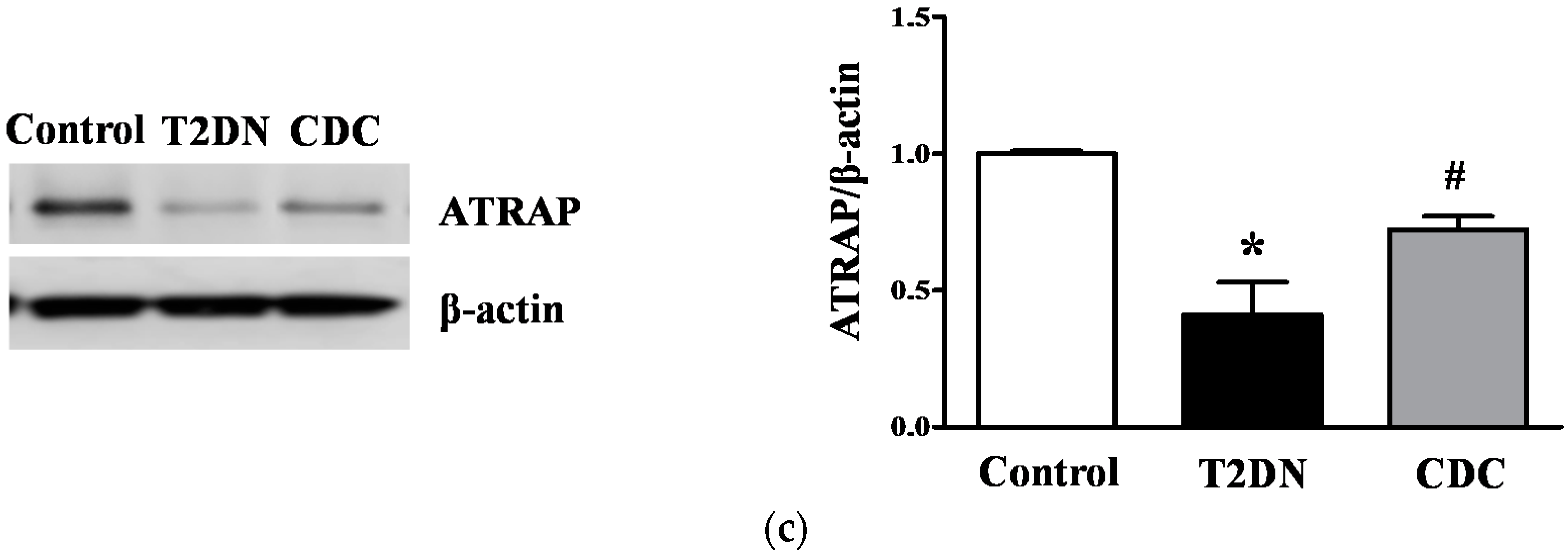
2.8. Effect of 12-LO on AT1R and ATRAP Expression in Glomeruli of Diabetic Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures
4.3. 12(S)-HETE Infusion by the Osmotic Mini-Pump
4.4. Induction of Type-1 and Type-2 Diabetes
4.5. Cell Membrane and Glomeruli Isolation
4.6. Measurement of 12(S)-HETE and Ang II
4.7. Competitive RT-PCR
4.8. Western Blot Analysis
4.9. Histological Evaluation
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 12-LO | 12-lipoxygenase |
| DN | Diabetic nephropathy |
| T2DN | Type-2 diabetic nephropathy |
| Ang II | Angiotensin II |
| AT1R | Angiotensin II type 1 receptor |
| MAU | Microalbuminuria |
| ATRAP | Angiotensin II type 1 receptor-associated protein |
| MAPK | Mitogen-activated protein kinase |
| IR | Insulin resistance |
| RAS | Renin-angiotensin system |
| ACEI | Angiotensin-converting enzyme inhibitor |
| ARB | Angiotensin II type 1 receptor blocker |
| 12(S)-HETE | 12(S)-hydroxyeicosatetraenoic acid |
| CDC | Cinnamyl-3,4-dihydroxy-α-cynanocinnamate |
| AMPK | AMP-activated protein kinase |
| IRS | Insulin receptor substrate |
| ELISA | Enzyme-linked immunosorbent assay |
| RIA | Insulin radioimmunoassay |
| MCs | Mesangial cells |
| STZ | Streptozotocin |
| FBS | Fetal bovine serum |
| RT-PCR | Reverse transcription–polymerase chain reaction |
| ISI | Insulin sensitivity index |
| PAS | Periodic acid-Schiff |
| BHPP | N-benzyl-N-hydroxy-5-phenylpentanamide |
References
- Kahn, S.E.; Zraika, S.; Utzschneider, K.M.; Hull, R.L. The beta cell lesion in type 2 diabetes: There has to be a primary functional abnormality. Diabetologia 2009, 52, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Adler, S. Structure–function relationships associated with extracellular matrix alterations in diabetic glomerulopathy. J. Am. Soc. Nephrol. 1994, 5, 1165–1172. [Google Scholar] [PubMed]
- Mogensen, C.E.; Christensen, C.K.; Vittinghus, E. The stages in diabetic renal disease. With emphasis on the stage of incipient diabetic nephropathy. Diabetes 1983, 32, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Wasada, T.; Katsumori, K.; Saeki, A.; Saito, S.; Omori, Y. Urinary albumin excretion rate is related to insulin resistance in normotensive subjects with impaired glucose tolerance. Diabetes Res. Clin. Pract. 1997, 34, 157–162. [Google Scholar] [CrossRef]
- Forsblom, C.M.; Eriksson, J.G.; Ekstrand, A.; Teppo, A.M.; Taskinen, M.R.; Groop, L.C. Insulin resistance and abnormal albumin excretion in non-diabetic first-degree relatives of patients with NIDMM. Diabetologia 1995, 38, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Favre, G.A.; Esnault, V.L.; Van-Obberghen, E. Modulation of glucose metabolism by the renin-angiotensin-aldosterone system. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E435–E449. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M.; Brown, N.J. The renin-angiotensin-aldosterone system and glucose homeostasis. Trends Pharmacol. Sci. 2011, 32, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Reyes, J.A.; Arellano-Plancarte, A.; Castillo-Hernandez, J.R. Angiotensin II and the development of insulin resistance: Implications for diabetes. Mol. Cell. Endocrinol. 2009, 302, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Perkins, J.M.; Davis, S.N. The renin-angiotensin-aldosterone system: A pivotal role in insulin sensitivity and glycemic control. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.G.; Lanting, L.; Vaziri, N.D.; Li, Z.; Sepassi, L.; Rodriguez-Iturbe, B.; Natarajan, R. Upregulation of angiotensin II type 1 receptor, inflammatory mediators, and enzymes of arachidonate metabolism in obese zucker rat kidney: Reversal by angiotensin II type 1 receptor blockade. Circulation 2005, 111, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.M.; Cooper, M.E.; de-Zeeuw, D.; Keane, W.F.; Mitch, W.E.; Parving, H.H.; Remuzzi, G.; Snapinn, S.M.; Zhang, Z.; Shahinfar, S.; et al. Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathy. N. Engl. J. Med. 2001, 345, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wei, R.B.; Xing, Y.; Tang, L.; Zheng, X.Y.; Wang, Z.C.; Gao, Y.W.; Li, M.X.; Chen, X.M. A meta-analysis of the effect of angiotensin receptor blockers and calcium channel blockers on blood pressure, glycemia and the HOMA-IR index in non-diabetic patients. Metabolism 2013, 62, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wei, R.B.; Wang, Z.C.; Wang, N.; Gao, Y.W.; Li, M.X.; Qiu, Q. A meta-analysis of the effects of angiotensin converting enzyme inhibitors and angiotensin II receptor blockers on insulin sensitivity in hypertensive patients without diabetes. Diabetes Res. Clin. Pract. 2015, 107, 415–423. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.; Davis, W., Jr.; Li, H.; Funk, C.D. Characterization of the murine epidermal 12/15-lipox-ygenase. Prostaglandins Other Lipid Mediat. 2001, 63, 93–107. [Google Scholar] [CrossRef]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [PubMed]
- Dobrian, A.D.; Lieb, D.C.; Cole, B.K.; Taylor-Fishwick, D.A.; Chakrabarti, S.K.; Nadler, J.L. Functional and pathological roles of the 12- and 15-lipoxygenases. Prog. Lipid Res. 2011, 50, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.G.; Li, S.L.; Lanting, L.; Kim, Y.S.; Shanmugam, N.; Reddy, M.A.; Natarajan, R. Relationship between 12/15-lipoxygenase and COX-2 in mesangial cells: Potential role in diabetic nephropathy. Kidney Int. 2006, 69, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.G.; Yuan, H.; Lanting, L.; Li, S.L.; Wang, M.; Shanmugam, N.; Kato, M.; Adler, S.G.; Reddy, M.A.; Natarajan, R. Products of 12/15-lipoxygenase upregulate the angiotensin II receptor. J. Am. Soc. Nephrol. 2008, 19, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Lanting, L.; Xu, Z.G.; Li, S.L.; Swiderski, P.; Putta, S.; Jonnalagadda, M.; Kato, M.; Natarajan, R. Effects of cholesterol-tagged small interfering RNAs targeting 12/15-lipoxygenase on parameters of diabetic nephropathy in a mouse model of type 1 diabetes. Am. J. Physiol. Ren. Physiol. 2008, 295, F605–F617. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.G.; Miao, L.N.; Cui, Y.C.; Jia, Y.; Yuan, H.; Wu, M. Angiotensin II type 1 receptor expression is increased via 12-lipoxygenase in high glucose-stimulated glomerular cells and type 2 diabetic glomeruli. Nephrol. Dial. Transplant. 2009, 24, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.Y.; Miao, L.N.; Li, B.; Ma, F.Z.; Liu, N.; Cai, L.; Xu, Z.G. Role of 12-lipoxygenase in decreasing P-cadherin and increasing angiotensin II type 1 receptor expression according to glomerular size in type 2 diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E708–E716. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Natarajan, R.; LaPage, J.; Lanting, L.; Kim, N.; Becerra, D.; Clemmons, B.; Nast, C.C.; Surya-Prakash, G.K.; Mandal, M.; et al. 12/15-Lipoxygenase inhibitors in diabetic nephropathy in the rat. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Adler, S.G.; Nast, C.C.; LaPage, J.; Gu, J.L.; Nadler, J.L.; Natarajan, R. 12-Lipoxygenase is increased in glucose-stimulated mesangial cells and in experimental diabetic nephropathy. Kidney Int. 2001, 59, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Nunemaker, C.S.; Chen, M.; Pei, H.; Kimble, S.D.; Keller, S.R.; Carter, J.D.; Yang, Z.; Smith, K.M.; Wu, R.; Bevard, M.H.; et al. 12-Lipoxygenase-knockout mice are resistant to inflammatory effects of obesity induced by western diet. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1065–E1075. [Google Scholar] [CrossRef] [PubMed]
- Grzesik, W.J.; Nadler, J.L.; Machida, Y.; Nadler, J.L.; Imai, Y.; Morris, M.A. Expression pattern of 12-lipoxygenases in human islets with type 1 diabetes and type 2 diabetes. J. Clin. Endocrinol. Metab. 2014, 100, E387–E395. [Google Scholar] [CrossRef] [PubMed]
- Tersey, S.A.; Maier, B.; Nishiki, Y.; Maganti, A.V.; Nadler, J.L.; Mirmira, R.G. 12-Lipoxygenase promotes obesity-induced oxidative stress in pancreatic islets. Mol. Cell. Biol. 2014, 34, 3735–3745. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.R.; Holman, T.R.; Imai, Y.; Jadhav, A.; Kenyon, V.; Maloney, D.J.; Nadler, J.L.; Rai, G.; Simeonov, A.; Taylor-Fishwick, D.A. Integration of pro-inflammatory cytokines, 12-lipoxygenase and NOX-1 in pancreatic islet beta cell dysfunction. Mol. Cell. Endocrinol. 2012, 358, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Sears, D.D.; Miles, P.D.; Chapman, J.; Ofrecio, J.M.; Almazan, F.; Thapar, D.; Miller, Y.I. 12/15-Lipoxygenase is required for the early onset of high fat diet-induced adipose tissue inflammation and insulin resistance in mice. PLoS ONE 2009, 4, e7250. [Google Scholar] [CrossRef] [PubMed]
- Ballermann, B.J.; Skorecki, K.L.; Brenner, B.M. Reduced glomerular angiotensin II receptor density in early untreated diabetes mellitus in the rat. Am. J. Physiol. 1984, 247, F110–F116. [Google Scholar] [PubMed]
- Xu, Z.G.; Yoo, T.H.; Ryu, D.R.; Cheon-Park, H.; Ha, S.K.; Han, D.S.; Adler, S.G.; Natarajan, R.; Kang, S.W. Angiotensin II receptor blocker inhibits p27Kip1 expression in glucose-stimulated podocytes and in diabetic glomeruli. Kidney Int. 2005, 67, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Danda, R.S.; Habiba, N.M.; Rincon-Choles, H.; Bhandari, B.K.; Barnes, J.L.; Abboud, H.E.; Pergola, P.E. Kidney involvement in a nongenetic rat model of type 2 diabetes. Kidney Int. 2005, 68, 2562–2571. [Google Scholar] [CrossRef] [PubMed]
- Bleich, D.; Chen, S.; Zipser, B.; Sun, D.; Funk, C.D.; Nadler, J.L. Resistance to type 1 diabetes induction in 12-lipoxygenase knockout mice. J. Clin. Investig. 1999, 103, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Sarafidis, P.A.; Ruilope, L.M. Insulin resistance, microalbuminuria, and chronic kidney disease. Curr. Hypertens. Rep. 2008, 10, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Mykkänen, L.; Zaccaro, D.J.; Wagenknecht, L.E.; Robbins, D.C.; Gabriel, M.; Haffner, S.M. Microalbuminuria is associated with insulin resistance in nondiabetic subjects: The insulin resistance atherosclerosis study. Diabetes 1998, 47, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chang, H.Y.; Huang, M.C.; Hwang, S.J.; Yang, Y.C.; Tai, T.Y.; Yang, H.J.; Chang, C.T.; Chang, C.J.; Li, Y.S.; et al. Association between insulin resistance and development of microalbuminuria in type 2 diabetes: A prospective cohort study. Diabetes Care 2011, 34, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Catalano, C.; Muscelli, E.; Quiñones-Galvan, A.; Baldi, S.; Masoni, A.; Gibb, I.; Torffvit, O.; Seghieri, G.; Ferrannini, E. Effect of insulin on systemic and renal handling of albumin in nondiabetic and NIDMM subjects. Diabetes 1997, 46, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Freedman, B.I.; Burdon, K.P.; Langefeld, C.D.; Howard, T.; Herrington, D.; Goff, D.C., Jr.; Bowden, D.W.; Wagenknecht, L.E.; Hedrick, C.C.; et al. Association of arachidonate 12-lipoxygenase genotype variation and glycemic control with albuminuria in type 2 diabetes. Am. J. Kidney Dis. 2008, 52, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Cole, B.K.; Morris, M.A.; Grzesik, W.J.; Leone, K.A.; Nadler, J.L. Adipose tissue-specific deletion of 12/15-lipoxygenase protects mice from the consequences of a high-fat diet. Mediat. Inflamm. 2012, 2012, 851798. [Google Scholar] [CrossRef] [PubMed]
- Rogacka, D.; Piwkowska, A.; Audzeyenka, I.; Angielski, S.; Jankowski, M. Involvement of the AMPK-PTEN pathway in insulin resistance induced by high glucose in cultured rat podocytes. Int. J. Biochem. Cell Biol. 2014, 51, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.B.; Carling, D.; Prentki, M.; Cacicedo, J.M. AMPK, insulin resistance, and the metabolic syndrome. J. Clin. Investig. 2013, 123, 2764–2772. [Google Scholar] [CrossRef] [PubMed]
- Lillioja, S.; Mott, D.M.; Howard, B.V.; Bennett, P.H.; Yki-Jarvinen, H.; Freymond, D.; Nyomba, B.L.; Zurlo, F.; Swinburn, B.; Bogardus, C. Impaired glucose tolerance as a disorder of insulin action. Longitudinal and cross-sectional studies in pima indians. N. Engl. J. Med. 1988, 318, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Warram, J.H.; Martin, B.C.; Krolewski, A.S.; Soeldner, J.S.; Kahn, C.R. Slow glucose removal rate and hyperinsulinemia precede the development of type II diabetes in the offspring of diabetic parents. Ann. Intern. Med. 1990, 113, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Tobe, K.; Tamemoto, H.; Ueki, K.; Kaburagi, Y.; Yamamoto-Honda, R.; Takahashi, Y.; Yoshizawa, F.; Aizawa, S.; Akanuma, Y.; et al. Insulin signalling and insulin actions in the muscles and livers of insulin-resistant, insulin receptor substrate 1-deficient mice. Mol. Cell. Biol. 1996, 16, 3074–3084. [Google Scholar] [CrossRef] [PubMed]
- Cozzone, D.; Frojdo, S.; Disse, E.; Debard, C.; Laville, M.; Pirola, L.; Vidal, H. Isoform-specific defects of insulin stimulation of Akt/protein kinase B (PKB) in skeletal muscle cells from type 2 diabetic patients. Diabetologia 2008, 51, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Nickenig, G.; Röling, J.; Strehlow, K.; Schnabel, P.; Böhm, M. Insulin induces upregulation of vascular AT1 receptor gene expression by posttranscriptional mechanisms. Circulation 1998, 98, 2453–2460. [Google Scholar] [CrossRef] [PubMed]
- Wakui, H.; Tamura, K.; Matsuda, M.; Bai, Y.; Dejima, T.; Shigenaga, A.; Masuda, S.; Azuma, K.; Maeda, A.; Hirose, T.; et al. Intrarenal suppression of angiotensin II type 1 receptor binding molecule in angiotensin II-infused mice. Am. J. Physiol. Ren. Physiol. 2010, 299, F991–F1003. [Google Scholar] [CrossRef] [PubMed]
- Li, G.W.; Pan, X.R. A new insulin-sensitivity index for the population-based study. Zhonghua Nei Ke Za Zhi 1993, 32, 656–660. [Google Scholar] [PubMed]
- Kim, J.J.; Li, J.J.; Jung, D.S.; Kwak, S.J.; Ryu, D.R.; Yoo, T.H.; Han, S.H.; Choi, H.Y.; Kim, H.J.; Han, D.S.; et al. Differential expression of nephrin according to glomerular size in early diabetic kidney disease. J. Am. Soc. Nephrol. 2007, 18, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R. Numerical density: Shape and size of particles. In Sterological Methods: Theoretical Foundations; Weibel, E.R., Ed.; Academic Press: London, UK, 1980; Volume 2, pp. 149–152. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.-Z.; Cheng, Y.-L.; Wang, W.-N.; Wu, H.; Zhang, Y.-Y.; Zang, C.-S.; Xu, Z.-G. 12-Lipoxygenase Inhibition on Microalbuminuria in Type-1 and Type-2 Diabetes Is Associated with Changes of Glomerular Angiotensin II Type 1 Receptor Related to Insulin Resistance. Int. J. Mol. Sci. 2016, 17, 684. https://doi.org/10.3390/ijms17050684
Xu H-Z, Cheng Y-L, Wang W-N, Wu H, Zhang Y-Y, Zang C-S, Xu Z-G. 12-Lipoxygenase Inhibition on Microalbuminuria in Type-1 and Type-2 Diabetes Is Associated with Changes of Glomerular Angiotensin II Type 1 Receptor Related to Insulin Resistance. International Journal of Molecular Sciences. 2016; 17(5):684. https://doi.org/10.3390/ijms17050684
Chicago/Turabian StyleXu, Hong-Zhao, Yan-Li Cheng, Wan-Ning Wang, Hao Wu, Yuan-Yuan Zhang, Chong-Sen Zang, and Zhong-Gao Xu. 2016. "12-Lipoxygenase Inhibition on Microalbuminuria in Type-1 and Type-2 Diabetes Is Associated with Changes of Glomerular Angiotensin II Type 1 Receptor Related to Insulin Resistance" International Journal of Molecular Sciences 17, no. 5: 684. https://doi.org/10.3390/ijms17050684
APA StyleXu, H.-Z., Cheng, Y.-L., Wang, W.-N., Wu, H., Zhang, Y.-Y., Zang, C.-S., & Xu, Z.-G. (2016). 12-Lipoxygenase Inhibition on Microalbuminuria in Type-1 and Type-2 Diabetes Is Associated with Changes of Glomerular Angiotensin II Type 1 Receptor Related to Insulin Resistance. International Journal of Molecular Sciences, 17(5), 684. https://doi.org/10.3390/ijms17050684