
Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders


Abstract
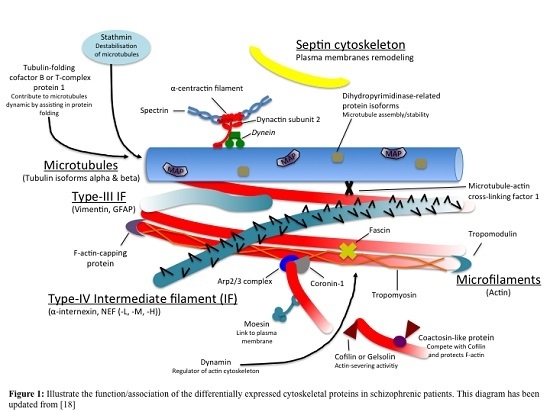
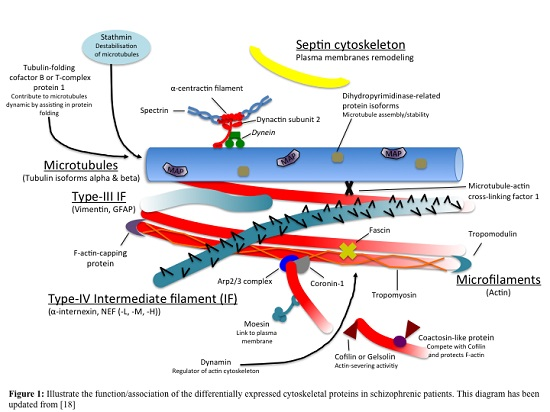
:
1. Introduction
2. The Cytoskeletal Systems
3. The Cytoskeleton in Neuropsychiatric Disorders
4. Proteomic Methodologies Used and Emerging
5. Microscopic Technologies Used and Emerging
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Whiteford, H.A.; Degenhardt, L.; Rehm, J.; Baxter, A.J.; Ferrari, A.J.; Erskine, H.E.; Charlson, F.J.; Norman, R.E.; Flaxman, A.D.; Johns, N.; et al. Global burden of disease attributable to mental and substance use disorders: Findings from the global burden of disease study 2010. Lancet 2013, 382, 1575–1586. [Google Scholar] [CrossRef]
- Bloom, D.E.; Cafiero, E.T.; Jané-Llopis, E.; Abrahams-Gessel, S.; Bloom, L.R.; Fatima, S.; Feigl, A.B.; Gaziano, T.; Mowafi, M.; Pandya, A.; et al. The Global Economic Burden of Non-Communicable Diseases; World Economic Forum: Geneva, Switzerland, 2011. [Google Scholar]
- Dotti, C.G.; Sullivan, C.A.; Banker, G.A. The establishment of polarity by hippocampal neurons in culture. J. Neurosci. 1988, 8, 1454–1468. [Google Scholar] [PubMed]
- English, J.A.; Pennington, K.; Dunn, M.J.; Cotter, D.R. The neuroproteomics of schizophrenia. Biol. Psychiatry 2011, 69, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Hild, G.; Bugyi, B.; Nyitrai, M. Conformational dynamics of actin: Effectors and implications for biological function. Cytoskeleton 2010, 67, 609–629. [Google Scholar] [CrossRef] [PubMed]
- Wasteneys, G.O.; Lechner, B. Microtubules. In Cellular Domains; John Wiley & Sons, Inc.: New York, NY, USA, 2011; pp. 229–243. [Google Scholar]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef]
- Firat-Karalar, E.N.; Welch, M.D. New mechanisms and functions of actin nucleation. Curr. Opin. Cell Biol. 2011, 23, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Shirao, T.; Gonzalez-Billault, C. Actin filaments and microtubules in dendritic spines. J. Neurochem. 2013, 126, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Snider, N.T.; Omary, M.B. Post-translational modifications of intermediate filament proteins: Mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2014, 15, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Perrot, R.; Eyer, J. Neuronal intermediate filaments and neurodegenerative disorders. Brain Res. Bull. 2009, 80, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Dent, E.W.; Gupton, S.L.; Gertler, F.B. The growth cone cytoskeleton in axon outgrowth and guidance. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Benitez-King, G.; Ramirez-Rodriguez, G.; Ortiz, L.; Meza, I. The neuronal cytoskeleton as a potential therapeutical target in neurodegenerative diseases and schizophrenia. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Blanpied, T.A.; Ehlers, M.D. Microanatomy of dendritic spines: Emerging principles of synaptic pathology in psychiatric and neurological disease. Biol. Psychiatry 2004, 55, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Fournet, V.; Schweitzer, A.; Chevarin, C.; Deloulme, J.C.; Hamon, M.; Giros, B.; Andrieux, A.; Martres, M.P. The deletion of STOP/MAP6 protein in mice triggers highly altered mood and impaired cognitive performances. J. Neurochem. 2012, 121, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Martins-De-Souza, D.; Dias-Neto, E.; Schmitt, A.; Falkai, P.; Gormanns, P.; Maccarrone, G.; Turck, C.W.; Gattaz, W.F. Proteome analysis of schizophrenia brain tissue. World J. Biol. Psychiatry 2010, 11, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, R.; Liu, X.; Bosedasgupta, S.; Muller, P.; Zhang, C.L.; Moshous, D.; Studer, V.; Schneider, J.; Genoud, C.; Fossoud, C.; et al. Coronin 1 regulates cognition and behavior through modulation of camp/protein kinase a signaling. PLoS Biol. 2014, 12, e1001820. [Google Scholar] [CrossRef] [PubMed]
- English, J.A.; Dicker, P.; Focking, M.; Dunn, M.J.; Cotter, D.R. 2-d dige analysis implicates cytoskeletal abnormalities in psychiatric disease. Proteomics 2009, 9, 3368–3382. [Google Scholar] [CrossRef] [PubMed]
- Rosslenbroich, V.; Dai, L.; Baader, S.L.; Noegel, A.A.; Gieselmann, V.; Kappler, J. Collapsin response mediator protein-4 regulates F-actin bundling. Exp. Cell Res. 2005, 310, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Arimura, N.; Menager, C.; Fukata, Y.; Kaibuchi, K. Role of CRMP-2 in neuronal polarity. J. Neurobiol. 2004, 58, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Yoshimura, T.; Tsuboi, D.; Kawabata, S.; Kaneko-Kawano, T.; Shirataki, H.; Takenawa, T.; Kaibuchi, K. CRMP-2 is involved in kinesin-1-dependent transport of the Sra-1/WAVE1 complex and axon formation. Mol. Cell. Biol. 2005, 25, 9920–9935. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Cha, C.; Ye, Y.; Zhang, J.; Li, S.; Wu, F.; Gong, S.; Guo, G. CRMP4 and CRMP2 interact to coordinate cytoskeleton dynamics, regulating growth cone development and axon elongation. Neural. Plast. 2015, 2015, 947423. [Google Scholar] [CrossRef] [PubMed]
- Kedracka-Krok, S.; Swiderska, B.; Jankowska, U.; Skupien-Rabian, B.; Solich, J.; Buczak, K.; Dziedzicka-Wasylewska, M. Clozapine influences cytoskeleton structure and calcium homeostasis in rat cerebral cortex and has a different proteomic profile than risperidone. J. Neurochem. 2015, 132, 657–676. [Google Scholar] [CrossRef] [PubMed]
- Beasley, C.L.; Pennington, K.; Behan, A.; Wait, R.; Dunn, M.J.; Cotter, D. Proteomic analysis of the anterior cingulate cortex in the major psychiatric disorders: Evidence for disease-associated changes. Proteomics 2006, 6, 3414–3425. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Ericson, M.; Soderpalm, B. Chronic phencyclidine increases synapsin-1 and synaptic adaptation proteins in the medial prefrontal cortex. ISRN Psychiatry 2013, 2013, 620361. [Google Scholar] [CrossRef] [PubMed]
- Johnston-Wilson, N.L.; Sims, C.D.; Hofmann, J.P.; Anderson, L.; Shore, A.D.; Torrey, E.F.; Yolken, R.H. Disease-specific alterations in frontal cortex brain proteins in schizophrenia, bipolar disorder, and major depressive disorder. The stanley neuropathology consortium. Mol. Psychiatry 2000, 5, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, S.; Swatton, J.E.; Ryan, M.M.; Huffaker, S.J.; Huang, J.-J.; Griffin, J.L.; Wayland, M.; Freeman, T.; Dudbridge, F.; Lilley, K.S.; et al. Mitochondrial dysfunction in schizophrenia: Evidence for compromised brain metabolism and oxidative stress. Mol. Psychiatry 2004, 9, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.; Dedova, I.; Cordwell, S.; Matsumoto, I. A proteome analysis of the anterior cingulate cortex gray matter in schizophrenia. Mol. Psychiatry 2006, 11, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Sivagnanasundaram, S.; Crossett, B.; Dedova, I.; Cordwell, S.; Matsumoto, I. Abnormal pathways in the genu of the corpus callosum in schizophrenia pathogenesis: A proteome study. Proteom. Clin. Appl. 2007, 1, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.; Dedova, I.; Cordwell, S.; Matsumoto, I. Altered proteins of the anterior cingulate cortex white matter proteome in schizophrenia. Proteom. Clin. Appl. 2007, 1, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Pennington, K.; Beasley, C.L.; Dicker, P.; Fagan, A.; English, J.; Pariante, C.M.; Wait, R.; Dunn, M.J.; Cotter, D.R. Prominent synaptic and metabolic abnormalities revealed by proteomic analysis of the dorsolateral prefrontal cortex in schizophrenia and bipolar disorder. Mol. Psychiatry 2008, 13, 1102–1117. [Google Scholar] [CrossRef] [PubMed]
- Pennington, K.; Dicker, P.; Dunn, M.J.; Cotter, D.R. Proteomic analysis reveals protein changes within layer 2 of the insular cortex in schizophrenia. Proteomics 2008, 8, 5097–5107. [Google Scholar] [CrossRef] [PubMed]
- Behan, A.T.; Byrne, C.; Dunn, M.J.; Cagney, G.; Cotter, D.R. Proteomic analysis of membrane microdomain-associated proteins in the dorsolateral prefrontal cortex in schizophrenia and bipolar disorder reveals alterations in LAMP, STXBP1 and BASP1 protein expression. Mol. Psychiatry 2009, 14, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Gattaz, W.F.; Schmitt, A.; Maccarrone, G.; Hunyadi-Gulyas, E.; Eberlin, M.N.; Souza, G.H.; Marangoni, S.; Novello, J.C.; Turck, C.W.; et al. Proteomic analysis of dorsolateral prefrontal cortex indicates the involvement of cytoskeleton, oligodendrocyte, energy metabolism and new potential markers in schizophrenia. J. Psychiatr. Res. 2009, 43, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Gattaz, W.F.; Schmitt, A.; Novello, J.C.; Marangoni, S.; Turck, C.W.; Dias-Neto, E. Proteome analysis of schizophrenia patients wernicke’s area reveals an energy metabolism dysregulation. BMC Psychiatry 2009, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Gattaz, W.F.; Schmitt, A.; Rewerts, C.; Maccarrone, G.; Dias-Neto, E.; Turck, C.W. Prefrontal cortex shotgun proteome analysis reveals altered calcium homeostasis and immune system imbalance in schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Gattaz, W.F.; Schmitt, A.; Rewerts, C.; Marangoni, S.; Novello, J.C.; Maccarrone, G.; Turck, C.W.; Dias-Neto, E. Alterations in oligodendrocyte proteins, calcium homeostasis and new potential markers in schizophrenia anterior temporal lobe are revealed by shotgun proteome analysis. J. Neural Transm. 2009, 116, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Nesvaderani, M.; Matsumoto, I.; Sivagnanasundaram, S. Anterior hippocampus in schizophrenia pathogenesis: Molecular evidence from a proteome study. Aust. N. Z. J. Psychiatry 2009, 43, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Focking, M.; Dicker, P.; English, J.A.; Schubert, K.O.; Dunn, M.J.; Cotter, D.R. Common proteomic changes in the hippocampus in schizophrenia and bipolar disorder and particular evidence for involvement of cornu ammonis regions 2 and 3. Arch. Gen. Psychiatry 2011, 68, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Schubert, K.O.; Focking, M.; Cotter, D.R. Proteomic pathway analysis of the hippocampus in schizophrenia and bipolar affective disorder implicates 14-3-3 signaling, aryl hydrocarbon receptor signaling, and glucose metabolism: Potential roles in gabaergic interneuron pathology. Schizophr. Res. 2015, 167, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Wesseling, H.; Gottschalk, M.G.; Bahn, S. Targeted multiplexed selected reaction monitoring analysis evaluates protein expression changes of molecular risk factors for major psychiatric disorders. Int. J. Neuropsychopharmacol. 2014, 18, 1–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Saia-Cereda, V.M.; Cassoli, J.S.; Schmitt, A.; Falkai, P.; Nascimento, J.M.; Martins-de-Souza, D. Proteomics of the corpus callosum unravel pivotal players in the dysfunction of cell signaling, structure, and myelination in schizophrenia brains. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J. The neuropathology of primary mood disorder. Brain 2002, 125, 1428–1449. [Google Scholar] [CrossRef] [PubMed]
- Berke, J.D.; Hyman, S.E. Addiction, dopamine, and the molecular mechanisms of memory. Neuron 2000, 25, 515–532. [Google Scholar] [CrossRef]
- McKinnon, M.C.; Yucel, K.; Nazarov, A.; MacQueen, G.M. A meta-analysis examining clinical predictors of hippocampal volume in patients with major depressive disorder. J. Psychiatry Neurosci. JPN 2009, 34, 41–54. [Google Scholar] [PubMed]
- Drevets, W.C.; Ongur, D.; Price, J.L. Neuroimaging abnormalities in the subgenual prefrontal cortex: Implications for the pathophysiology of familial mood disorders. Mol. Psychiatry 1998, 3, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.; Hagan, J.J.; Heidbreder, C.A. Neuronal plasticity, stress and depression: Involvement of the cytoskeletal microtubular system? Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.T.; Chang, R.C.; Law, A.C. A breach in the scaffold: The possible role of cytoskeleton dysfunction in the pathogenesis of major depression. Ageing Res. Rev. 2013, 12, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Guest, P.C.; Harris, L.W.; Vanattou-Saifoudine, N.; Webster, M.J.; Rahmoune, H.; Bahn, S. Identification of proteomic signatures associated with depression and psychotic depression in post-mortem brains from major depression patients. Transl. Psychiatry 2012, 2, e87. [Google Scholar] [CrossRef] [PubMed]
- Martins-de-Souza, D.; Guest, P.C.; Vanattou-Saifoudine, N.; Rahmoune, H.; Bahn, S. Phosphoproteomic differences in major depressive disorder postmortem brains indicate effects on synaptic function. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Venkova, K.; Christov, A.; Gunning, W.; Park, J. Collapsin response mediator protein-2: An emerging pathologic feature and therapeutic target for neurodisease indications. Mol. Neurobiol 2011, 43, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Kawano, Y.; Arimura, N.; Kawabata, S.; Kikuchi, A.; Kaibuchi, K. GSK-3β regulates phosphorylation of CRMP-2 and neuronal polarity. Cell 2005, 120, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; Spinhoven, P.; Polak, M.; Bus, B.A.; Penninx, B.W.; Elzinga, B.M. Serum BDNF concentrations as peripheral manifestations of depression: Evidence from a systematic review and meta-analyses on 179 associations (N = 9484). Mol. Psychiatry 2014, 19, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Focking, M.; Opstelten, R.; Prickaerts, J.; Steinbusch, H.W.; Dunn, M.J.; van den Hove, D.L.; Cotter, D.R. Proteomic investigation of the hippocampus in prenatally stressed mice implicates changes in membrane trafficking, cytoskeletal, and metabolic function. Dev. Neurosci. 2014, 36, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.M.; Adams, J.; DelBello, M.P.; Holland, S.K.; Schmithorst, V.; Levine, A.; Jarvis, K.; Strakowski, S.M. Evidence of white matter pathology in bipolar disorder adolescents experiencing their first episode of Mania: A diffusion tensor imaging study. Am. J. Psychiatry 2006, 163, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G.; Halaris, A.; Selemon, L.D. Reductions in neuronal and glial density characterize the dorsolateral prefrontal cortex in bipolar disorder. Biol. Psychiatry 2001, 49, 741–752. [Google Scholar] [CrossRef]
- Zhao, Z.; Xu, J.; Chen, J.; Kim, S.; Reimers, M.; Bacanu, S.A.; Yu, H.; Liu, C.; Sun, J.; Wang, Q.; et al. Transcriptome sequencing and genome-wide association analyses reveal lysosomal function and actin cytoskeleton remodeling in schizophrenia and bipolar disorder. Mol. Psychiatry 2015, 20, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Stepaniants, S.B.; Mao, M.; Weng, L.; Feetham, M.C.; Doyle, M.J.; Yi, E.C.; Dai, H.; Thorsson, V.; Eng, J.; et al. Integrated genomic and proteomic analyses of gene expression in mammalian cells. Mol. Cell. Proteom. MCP 2004, 3, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Abreu Rde, S.; Ko, D.; Le, S.Y.; Shapiro, B.A.; Burns, S.C.; Sandhu, D.; Boutz, D.R.; Marcotte, E.M.; Penalva, L.O. Sequence signatures and mRNA concentration can explain two-thirds of protein abundance variation in a human cell line. Mol. Syst. Biol. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Sanchez, J.C.; Gooley, A.A.; Appel, R.D.; Humphery-Smith, I.; Hochstrasser, D.F.; Williams, K.L. Progress with proteome projects: Why all proteins expressed by a genome should be identified and how to do it. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar] [CrossRef] [PubMed]
- Coumans, J.V.; Gau, D.; Poljak, A.; Wasinger, V.; Roy, P.; Moens, P. Green fluorescent protein expression triggers proteome changes in breast cancer cells. Exp. Cell Res. 2014, 320, 33–45. [Google Scholar] [CrossRef] [PubMed]
- O'Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [PubMed]
- Gianazza, E.; Righetti, P.G. Immobilized PH gradients. Electrophoresis 2009, 30, S112–S121. [Google Scholar] [CrossRef] [PubMed]
- Görg, A.; Drews, O.; Lück, C.; Weiland, F.; Weiss, W. 2-DE with IPGs. Electrophoresis 2009, 30, S122–S132. [Google Scholar] [CrossRef] [PubMed]
- Unlu, M.; Morgan, M.E.; Minden, J.S. Difference gel electrophoresis: A single gel method for detecting changes in protein extracts. Electrophoresis 1997, 18, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Goodlett, D.R. Mass spectrometry in proteomics. Chem. Rev. 2001, 101, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.E.; Gygi, S.P. Target-decoy search strategy for increased confidence in large-scale protein identifications by mass spectrometry. Nat. Methods 2007, 4, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.L.; Huang, Y.N.; Marchese, J.N.; Williamson, B.; Parker, K.; Hattan, S.; Khainovski, N.; Pillai, S.; Dey, S.; Daniels, S.; et al. Multiplexed protein quantitation in saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Mol. Cell. Proteom. MCP 2004, 3, 1154–1169. [Google Scholar] [CrossRef] [PubMed]
- Gygi, S.P.; Rist, B.; Gerber, S.A.; Turecek, F.; Gelb, M.H.; Aebersold, R. Quantitative analysis of complex protein mixtures using isotope-coded affinity tags. Nat. Biotechnol. 1999, 17, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Kellermann, J.; Lottspeich, F. A novel strategy for quantitative proteomics using isotope-coded protein labels. Proteomics 2005, 5, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.E.; Blagoev, B.; Kratchmarova, I.; Kristensen, D.B.; Steen, H.; Pandey, A.; Mann, M. Stable isotope labeling by amino acids in cell culture, silac, as a simple and accurate approach to expression proteomics. Mol. Cell. Proteom. MCP 2002, 1, 376–386. [Google Scholar] [CrossRef]
- McClatchy, D.B.; Yates, J.R., 3rd. Stable isotope labeling of mammals (SILAM). CSH Protoc. 2008, 2008. [Google Scholar] [CrossRef]
- Chelius, D.; Bondarenko, P.V. Quantitative profiling of proteins in complex mixtures using liquid chromatography and mass spectrometry. J. Proteome Res. 2002, 1, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Neilson, K.A.; Ali, N.A.; Muralidharan, S.; Mirzaei, M.; Mariani, M.; Assadourian, G.; Lee, A.; van Sluyter, S.C.; Haynes, P.A. Less label, more free: Approaches in label-free quantitative mass spectrometry. Proteomics 2011, 11, 535–553. [Google Scholar] [CrossRef] [PubMed]
- Ramsby, M.; Makowski, G. Differential detergent fractionation of eukaryotic cells. Cold Spring Harb. Protoc. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.E.; Souda, P.; Jin, Y.P.; Whitelegge, J.P.; Reed, E.F. Characterization of the endothelial cell cytoskeleton following hla class I ligation. PLoS ONE 2012, 7, e29472. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Wilkins, J.A. Compositional characterization of the cytoskeleton of NK-like cells. J. Proteome Res. 2005, 4, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Coumans, J.V.; dos Remedios, C.G. Actin-binding proteins in mouse C2 myoblasts and myotubes: A combination of affinity chromatography and two-dimensional gel electrophoresis. Electrophoresis 1998, 19, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Coumans, J.V.; Humphery-Smith, I.; dos Remedios, C.G. Two-dimensional gel electrophoresis of actin-binding proteins isolated by affinity chromatography from human skeletal muscle. Electrophoresis 1997, 18, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Chuong, S.D.; Good, A.G.; Taylor, G.J.; Freeman, M.C.; Moorhead, G.B.; Muench, D.G. Large-scale identification of tubulin-binding proteins provides insight on subcellular trafficking, metabolic channeling, and signaling in plant cells. Mol. Cell. Proteom. MCP 2004, 3, 970–983. [Google Scholar] [CrossRef] [PubMed]
- Kozielski, F.; Riaz, T.; DeBonis, S.; Koehler, C.J.; Kroening, M.; Panse, I.; Strozynski, M.; Donaldson, I.M.; Thiede, B. Proteome analysis of microtubule-associated proteins and their interacting partners from mammalian brain. Amino Acids 2010, 41, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Phung-Koskas, T.; Pilon, A.; Pous, C.; Betzina, C.; Sturm, M.; Bourguet-Kondracki, M.L.; Durand, G.; Drechou, A. STAT5B-mediated growth hormone signaling is organized by highly dynamic microtubules in hepatic cells. J. Biol. Chem. 2005, 280, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Asplund, A.; Edqvist, P.H.; Schwenk, J.M.; Ponten, F. Antibodies for profiling the human proteome-the human protein atlas as a resource for cancer research. Proteomics 2012, 12, 2067–2077. [Google Scholar] [CrossRef] [PubMed]
- Reverdatto, S.; Burz, D.S.; Shekhtman, A. Peptide aptamers: Development and applications. Curr. Top. Med. Chem. 2015, 15, 1082–1101. [Google Scholar] [CrossRef] [PubMed]
- Solier, C.; Langen, H. Antibody-based proteomics and biomarker research—Current status and limitations. Proteomics 2014, 14, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.; Hunter, C.L. Quantitative mass spectrometric multiple reaction monitoring assays for major plasma proteins. Mol. Cell. Proteom. MCP 2006, 5, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Nagerl, U.V.; Willig, K.I.; Hein, B.; Hell, S.W.; Bonhoeffer, T. Live-cell imaging of dendritic spines by sted microscopy. Proc. Natl. Acad. Sci. USA 2008, 105, 18982–18987. [Google Scholar] [CrossRef] [PubMed]
- Digman, M.A.; Caiolfa, V.R.; Zamai, M.; Gratton, E. The phasor approach to fluorescence lifetime imaging analysis. Biophys. J. 2008, 94, L14–L16. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.H.; Hanley, Q.S.; Verveer, P.J. Graphical representation and multicomponent analysis of single-frequency fluorescence lifetime imaging microscopy data. J. Microsc. 2004, 213, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Jameson, D.M.; Gratton, E.; Hall, R. The measurement and analysis of heterogeneous emissions by multifrequency phase and modulation fluorometry. Appl. Spectrosc. Rev. 1984, 20, 55–106. [Google Scholar] [CrossRef]
- Redford, G.I.; Clegg, R.M. Polar plot representation for frequency-domain analysis of fluorescence lifetimes. J. Fluoresc. 2005, 15, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Eid, J.S. Two-Photon Dual Channel Fluctuation Correlation Spectroscopy: Theory and Application. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Ann Arbor, MI, USA, 2002. [Google Scholar]
- Brown, C.M.; Dalal, R.B.; Hebert, B.; Digman, M.A.; Horwitz, A.R.; Gratton, E. Raster image correlation spectroscopy (rics) for measuring fast protein dynamics and concentrations with a commercial laser scanning confocal microscope. J. Microsc. 2008, 229, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.K.; Zareno, J.; Digman, M.A.; Gratton, E.; Horwitz, A.R. Cross-correlated fluctuation analysis reveals phosphorylation-regulated paxillin-fak complexes in nascent adhesions. Biophys. J. 2011, 100, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Digman, M.A.; Gratton, E. Analysis of diffusion and binding in cells using the rics approach. Microsc. Res. Tech. 2009, 72, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Hell, S.W. Far-field optical nanoscopy. Science 2007, 316, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Calligaris, D.; Verdier-Pinard, P.; Devred, F.; Villard, C.; Braguer, D.; Lafitte, D. Microtubule targeting agents: From biophysics to proteomics. Cell. Mol. Life Sci. CMLS 2010, 67, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Pertsinidis, A.; Mukherjee, K.; Sharma, M.; Pang, Z.P.; Park, S.R.; Zhang, Y.; Brunger, A.T.; Sudhof, T.C.; Chu, S. Ultrahigh-resolution imaging reveals formation of neuronal SNARE/Munc18 complexes in situ. Proc. Natl. Acad. Sci. USA 2013, 110, E2812–E2820. [Google Scholar] [CrossRef] [PubMed]
- Urban, N.T.; Willig, K.I.; Hell, S.W.; Nagerl, U.V. Sted nanoscopy of actin dynamics in synapses deep inside living brain slices. Biophys. J. 2011, 101, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Donnert, G.; Keller, J.; Wurm, C.A.; Rizzoli, S.O.; Westphal, V.; Schonle, A.; Jahn, R.; Jakobs, S.; Eggeling, C.; Hell, S.W. Two-color far-field fluorescence nanoscopy. Biophys. J. 2007, 92, L67–L69. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H. Applying superresolution localization-based microscopy to neurons. Synapse 2015, 69, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Biermann, B.; Sokoll, S.; Klueva, J.; Missler, M.; Wiegert, J.S.; Sibarita, J.B.; Heine, M. Imaging of molecular surface dynamics in brain slices using single-particle tracking. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Groc, L.; Lafourcade, M.; Heine, M.; Renner, M.; Racine, V.; Sibarita, J.B.; Lounis, B.; Choquet, D.; Cognet, L. Surface trafficking of neurotransmitter receptor: Comparison between single-molecule/quantum dot strategies. J. Neurosci. 2007, 27, 12433–12437. [Google Scholar] [CrossRef] [PubMed]
- Alcor, D.; Gouzer, G.; Triller, A. Single-particle tracking methods for the study of membrane receptors dynamics. Eur. J. Neurosci. 2009, 30, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, C.; Gratton, E.; Beltram, F.; Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. USA 2013, 110, 12307–12312. [Google Scholar] [CrossRef] [PubMed]
- Huisken, J.; Stainier, D.Y. Selective plane illumination microscopy techniques in developmental biology. Development 2009, 136, 1963–1975. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.B.; Schmitz, H.M.; Santi, P.A. TSLIM imaging and a morphometric analysis of the mouse spiral ganglion. Hear. Res. 2011, 278, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Schroter, T.J.; Johnson, S.B.; John, K.; Santi, P.A. Scanning thin-sheet laser imaging microscopy (STSLIM) with structured illumination and Hilo background rejection. Biomed. Opt. Express 2012, 3, 170–177. [Google Scholar] [CrossRef] [PubMed][Green Version]
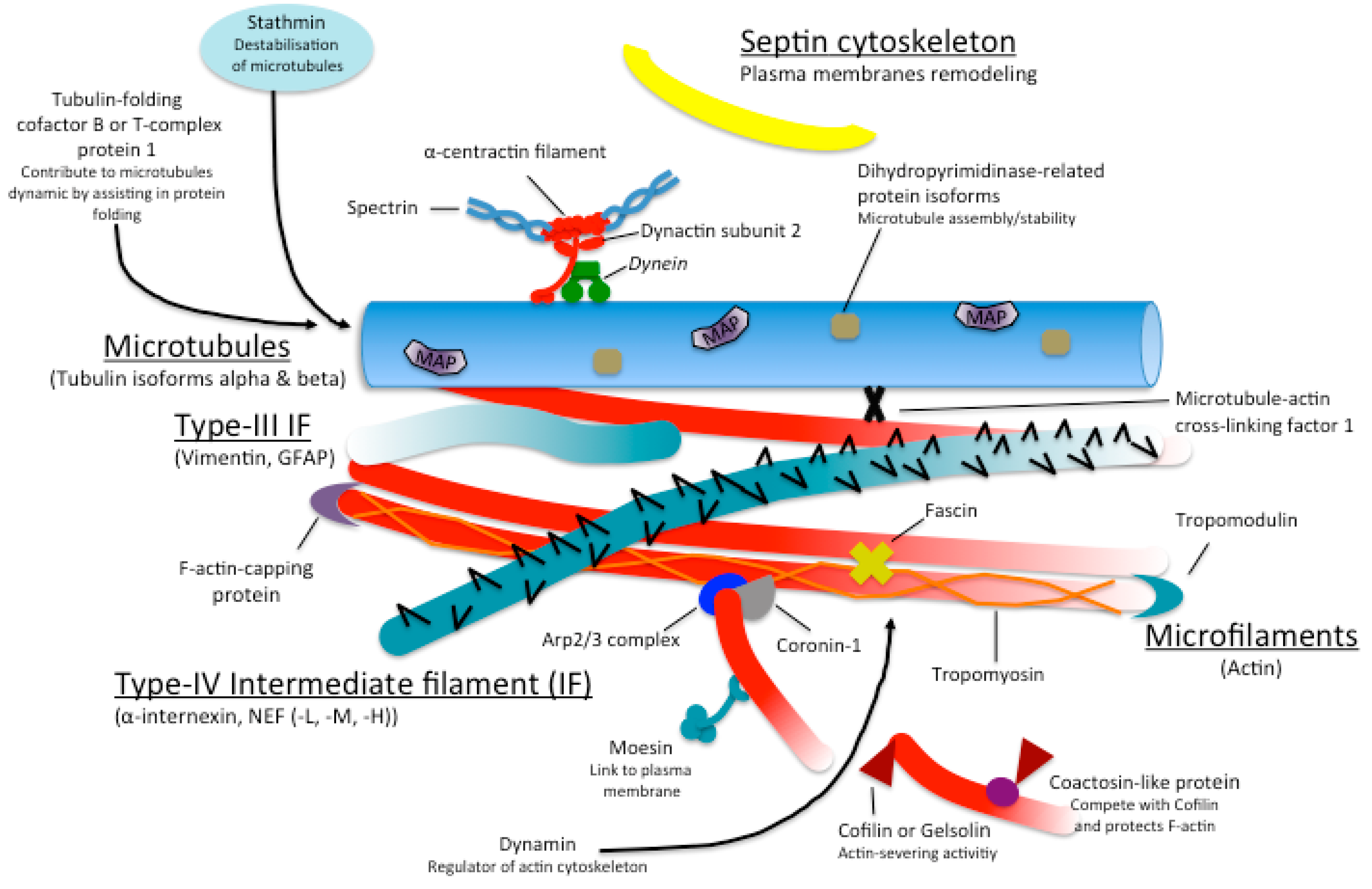

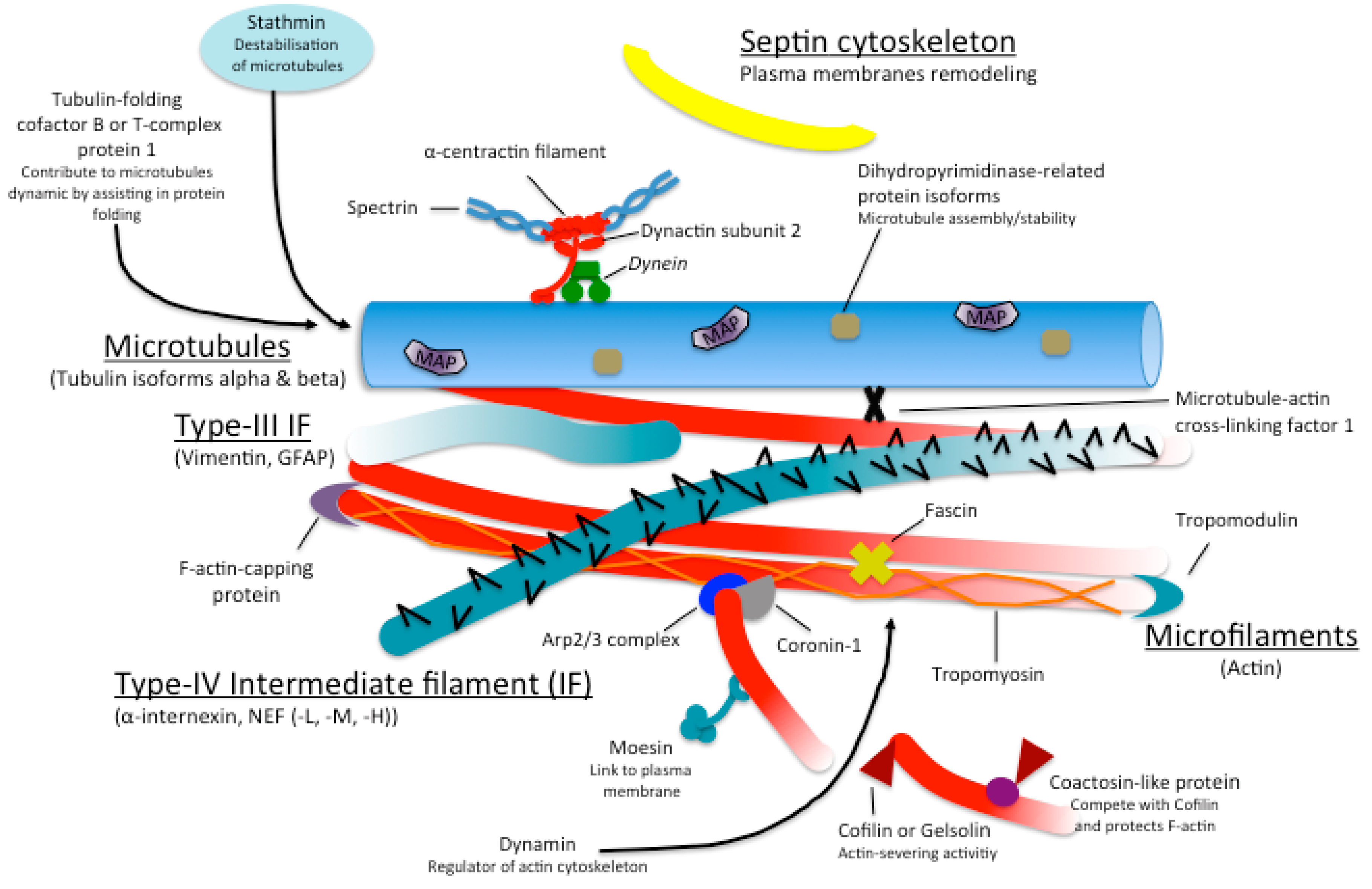
{kind=link}
{kind=link}
| References | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein Name | Gene Symbol | Uniprot acc. No. | [26] | [27] | [24] | [28] | [29] | [30] | [31] | [32] | [18] | [33] | [34] | [35] | [36] | [37] | [38] | [39,40] | [41] | [42] |
| Actin isoforms | ACTG1 | P63261 | ↓GM and WM | ↓ | ‒ | ‒ | ‒ | ‒ | ‒ | |||||||||||
| ACTB | P02570 | ↓ | ↑ | ↓ | ‒ | ↓ | ||||||||||||||
| ACTA1 | P68133 | ↓ | ||||||||||||||||||
| α-centractin | ACTR1A | P61163 | ↑ | |||||||||||||||||
| Actin-related protein 2/3 complex components | ACTR3 | Q92747 | ‒ | ↑ | ↓ | |||||||||||||||
| ARPC1A | P61158 | ↓ GM | ||||||||||||||||||
| ARPC1B | O15143 | |||||||||||||||||||
| F-actin-capping protein subunit α-2 | CAPZA2 | P47755 | ↓ | |||||||||||||||||
| Cofilin-1 | CFL1 | P23528 | ↓ | |||||||||||||||||
| Coronin-1A | CORO1A | P31146 | ↓ | |||||||||||||||||
| Coactosin-like protein | COTL1 | Q14019 | ↑ | |||||||||||||||||
| Dynactin subunit 2 | DCTN2 | Q13561 | ↑ | |||||||||||||||||
| Dynamin-1 | DNM1 | Q05193 | ↑ GM, ↓ WM | ↑ | ↑ | ↑ | ||||||||||||||
| Dihydropyrimidi-nase-related protein isoforms | DPYSL2 | Q16555 | ↓ | ↓ GM and WM | ↑ | ↓ | ↓ | ↓ | ↑ | ↓ | ↓ | ↑ | ↑ | ↓ | ||||||
| DPYSL3 | Q14195 | |||||||||||||||||||
| Fascin | FSCN1 | Q16658 | ↓ WM | ↓ | ↑ | |||||||||||||||
| Glial fibrillary acidic protein | GFAP | P14136 | ↓ | ↑ | ↑ | ↑ | ↑ | ↓ | ↓ | ↓ | ||||||||||
| Gelsolin | GSN | P06396 | ↓ GM and WM | ↑ | ↓ | |||||||||||||||
| Α-internexin | INA | Q16352 | ↑ | ↑ | ↓ | ↓ | ↑ | ↓ | ||||||||||||
| Microtubule-actin cross-linking factor 1 | MACF1 | Q96PK2 | ↑ | |||||||||||||||||
| Microtubule-asso-ciated protein isoforms | MAP1A | P78559 | ↑ | |||||||||||||||||
| MAP2 | P11137 | ↑ | ||||||||||||||||||
| MAP6 | Q96JE9 | ↑ | ||||||||||||||||||
| Moesin | MSN | P26038 | ↑ GM | |||||||||||||||||
| Neurofilament intermediate proteins | NEFL | P07196 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | ‒ | ↓ | ↓ | ↓ | ||||||||
| NEFM | P07197 | ↓ | ‒ | ↓ | ↓ | ↓ | ||||||||||||||
| NEFH | P12036 | ↓ | ↑ | |||||||||||||||||
| Septin isoforms | SEPT3 | Q9UH03 | ↓ GM and WM | ‒ | ↑ | ‒ | ‒ | |||||||||||||
| SEPT5 | Q99719 | ↑ | ‒ | ↓ | ||||||||||||||||
| SEPT11 | Q9NVA2 | ↑ | ↓ | |||||||||||||||||
| Spectrin α chain, non-erythrocytic 1 | SPTAN1 | Q13813 | ↓ GM and WM | ↓ | ||||||||||||||||
| Stathmin | STMN1 | P16949 | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ||||||||||||
| Tubulin isoforms | TUBB | P07437 | ‒ | ‒ | ‒ | ‒ | ‒ | ↑ | ↓ | ‒ | ↑ | ‒ | ↑ | |||||||
| TUBA1B | P68363 | ‒ | ↓ | ‒ | ‒ | ↓ | ‒ | ↓ | ↓ | |||||||||||
| TUBB2A | Q6FGZ8 | ↓ GM and WM | ↓ | ‒ | ‒ | ↑ | ‒ | ‒ | ||||||||||||
| TUBB3 | Q13509 | ‒ | ↑ | ‒ | ↑ | |||||||||||||||
| TUBB4A | P04350 | ↑ | ↓ | ↓ | ||||||||||||||||
| TUBAL3 | ↑ | |||||||||||||||||||
| TUBB2B | ↓ | |||||||||||||||||||
| TUBB4B | ↑ | |||||||||||||||||||
| TUBB6 | ↑ | |||||||||||||||||||
| Tubulin-folding cofactor B | TBCB | Q99426 | ↑ | |||||||||||||||||
| T-complex protein 1 subunit α | TCP1 | P17987 | ↑ | |||||||||||||||||
| Tropomodulin-2 | TMOD2 | Q9NZR1 | ↓ | |||||||||||||||||
| Tropomyosin isoforms | TPM3 | P06753 | ↓ | ‒ | ‒ | |||||||||||||||
| TPM4 | P67936 | ↑ | ↑ | |||||||||||||||||
| Vimentin | VIM | P08670 | ↑ | ↑ | ↑ | ↑ | ||||||||||||||
| References | ||||||
|---|---|---|---|---|---|---|
| Protein Name | Gene Symbol | Uniprot acc. No. | [26] | [24] | [49] | [41] |
| Coronin-1A | CORO1A | P31146 | ↓ | |||
| Dihydropyrimidinase-related protein isoforms | CRMP1 | Q14194 | ‒ | ↑ | ‒ | |
| DPYSL2 | Q16555 | ↓ | ↑ | ↑ | ||
| DPYSL3 | Q14195 | ↑ | ||||
| Glial fibrillary acidic protein | GFAP | P14136 | ↓ | ↓ | ||
| Neurofilament intermediate proteins | NEFL | P07196 | ↑ | |||
| Microtubule-associated protein isoforms | MAP1LC3A | Q9H492 | ↑ | |||
| Tubulin isoforms | TUBA1B | P68363 | ↑ | ‒ | ||
| TUBA4B | Q9H853 | ↑ | ||||
| Transgelin-3 | TAGL3 | Q9UI15 | ↓ | |||
| Profilin | PFN1 | P07737 | ↑ | |||
| PFN2 | P35080 | ↑ | ||||
| References | ||||||||
|---|---|---|---|---|---|---|---|---|
| Protein Name | Gene Symbol | Uniprot acc. No. | [26] | [24] | [31] | [18] | [39,40] | [41] |
| Actin isoforms | ACTB | P60709 | ↑ | |||||
| Actin-related protein 2/3 complex components | ARPC5 | O15511 | ↑ | |||||
| Dihydropyrimidinase-related protein isoforms | CRMP1 | Q14194 | ‒ | ‒ | ‒ | ‒ | ||
| DPYSL2 | Q16555 | ↓ | ‒ | ↑ | ↓ | |||
| DPYSL3 | Q14195 | ↑ | ↑ | ↓ | ||||
| Dynactin subunit 2 | DCTN2 | Q13561 | ↓ | |||||
| Dynamin-1 | DNM1 | Q05193 | ↓ | |||||
| Fascin | FSCN1 | Q16658 | ↓ | |||||
| Glial fibrillary acidic protein | GFAP | P14136 | ↓ | |||||
| Α-internexin | INA | Q16352 | ↓ | ↓ | ||||
| Kinesin light chain 2 | KLC2 | Q9H0B6 | ↑ | |||||
| Microtubule-associated protein isoforms | MAP1LC3A | Q9H492 | ||||||
| Neurofilament intermediate proteins | NEFL | P07196 | ‒ | ‒ | ↓ | ↓ | ||
| NEFM | P07197 | ↓ | ↑ | ↓ | ||||
| NEFH | P12036 | ↓ | ||||||
| Septin isoforms | SEPT3 | Q9UH03 | ‒ | ‒ | ‒ | |||
| SEPT5 | Q99719 | ↑ | ‒ | |||||
| SEPT6 | Q14141 | ↑ | ‒ | |||||
| SEPT11 | Q9NVA2 | ↑ | ↓ | |||||
| Spectrin α chain, non-erythrocytic 1 | SPTAN1 | Q13813 | ↓ | |||||
| Stathmin | STMN1 | P16949 | ↑ | |||||
| Tubulin isoforms | TBBX | P07437 | ↓ | ‒ | ||||
| TUBA1B | P68363 | ↑ | ||||||
| TUBA4B | Q9H853 | ‒ | ||||||
| TUBB1 | Q9H4B7 | ↑ | ||||||
| T-complex protein 1 subunit α | TCP1 | P17987 | ↑ | |||||
| Tropomyosin isoforms | TPM4 | P67936 | ↓ | |||||
| Vimentin | VIM | P08670 | ↑ | ↓ | ||||
| Programmed cell death 6-interacting protein | PDCD6IP | Q8WUM4 | ↑ | |||||
| WD repeat-containing protein 1 | WDR1 | O75083 | ↓ | |||||
| Study (Year) | Region of the Brain Analysed | Technique Used | Ref. |
|---|---|---|---|
| Johnston-Wilson et al. (2000) | FC | 2-DE | [26] |
| Prabakaran et al. (2004)) | DLPFC | 2D-DIGE | [27] |
| Clark et al. (2006) | ACC | 2-DE | [28] |
| Beasley et al. (2006) | ACC | 2-DE | [24] |
| Sivagnanasundaram et al. (2007) | CC | 2-DE | [29] |
| Clark et al. (2007) | ACC | 2-DE | [30] |
| Pennington et al. (2008) | DLPFC | 2-DE/shotgun | [31] |
| Pennington et al. (2008) | IC | 2D-DIGE | [32] |
| English et al. (2009) | DLPFC | 2D-DIGE | [18] |
| Behan et al. (2009) | DLPFC | 2D-DIGE | [33] |
| Martins-de-Souza et al. (2009) | DLPFC | 2-DE | [34] |
| Martins-de-Souza et al. (2009) | WA | 2-DE | [35] |
| Martins-de-Souza et al. (2009) | DLPFC | Shotgun (ICPL) | [36] |
| Martins-de-Souza et al. (2009) | ATL | Shotgun (ICPL) | [37] |
| Nesvaderani et al. (2009) | Hippocampus | 2-DE | [38] |
| Focking et al. (2011) | Hippocampus | 2D-DIGE | [39] |
| Schubert et al. (2015) | Hippocampus | 2D-DIGE | [40] |
| Wesseling et al. (2014) | APFC | SRM | [41] |
| Saia-Cereda et al. (2015) | CC | Shotgun | [42] |
| Martins-de-Souza et al. (2012) | DLPFC | Shotgun | [49] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coumans, J.V.F.; Palanisamy, S.K.A.; McFarlane, J.; Moens, P.D.J. Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders. Int. J. Mol. Sci. 2016, 17, 581. https://doi.org/10.3390/ijms17040581
Coumans JVF, Palanisamy SKA, McFarlane J, Moens PDJ. Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders. International Journal of Molecular Sciences. 2016; 17(4):581. https://doi.org/10.3390/ijms17040581
Chicago/Turabian StyleCoumans, Joëlle V. F., Suresh K. A. Palanisamy, Jim McFarlane, and Pierre D. J. Moens. 2016. "Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders" International Journal of Molecular Sciences 17, no. 4: 581. https://doi.org/10.3390/ijms17040581
APA StyleCoumans, J. V. F., Palanisamy, S. K. A., McFarlane, J., & Moens, P. D. J. (2016). Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders. International Journal of Molecular Sciences, 17(4), 581. https://doi.org/10.3390/ijms17040581