
The Vitamin E Analog Gamma-Tocotrienol (GT3) and Statins Synergistically Up-Regulate Endothelial Thrombomodulin (TM)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
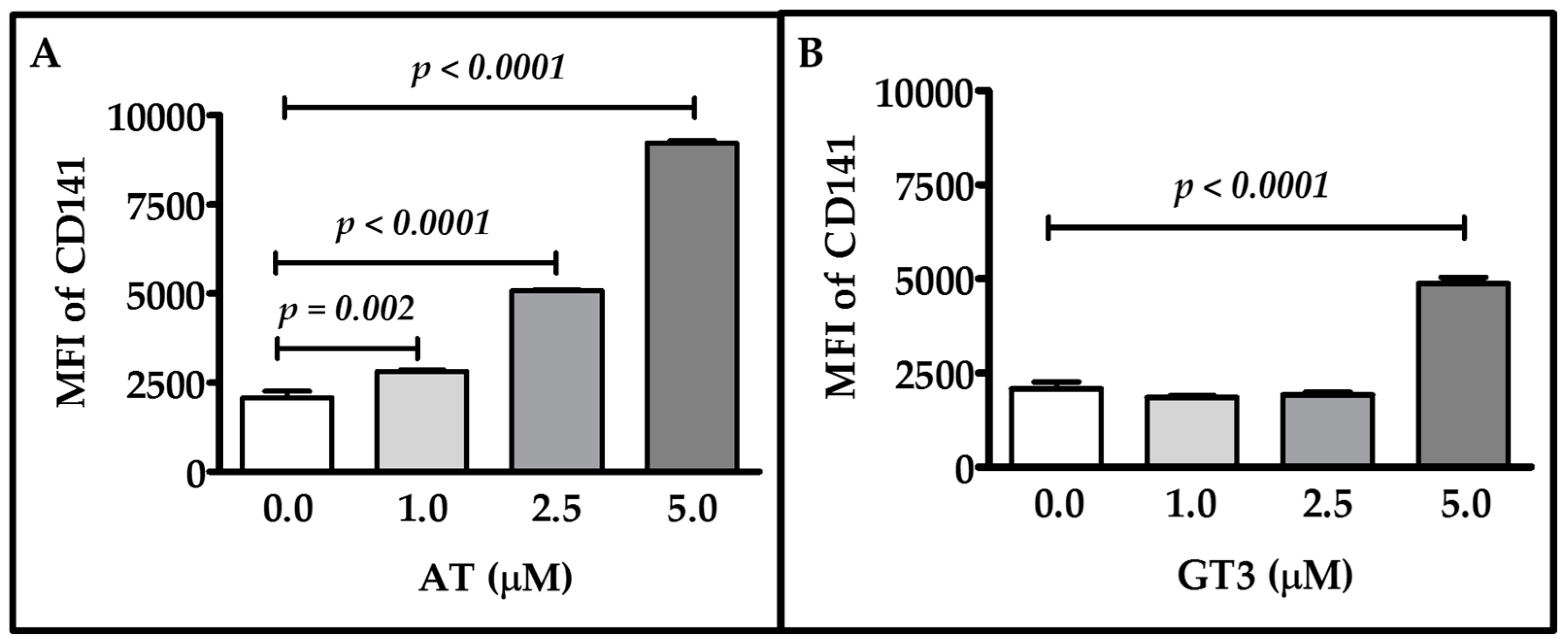
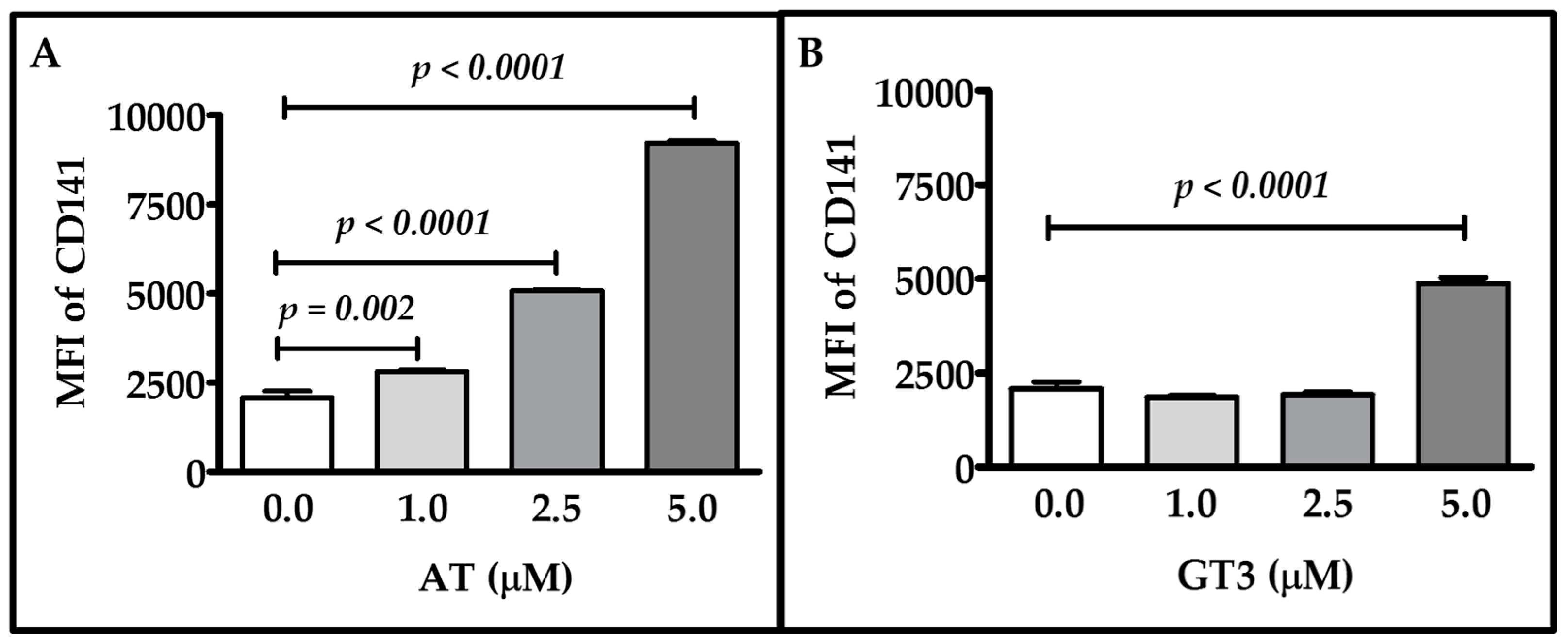
2.1. Dose-Response of Atorvastatin and GT3 on TM Expression
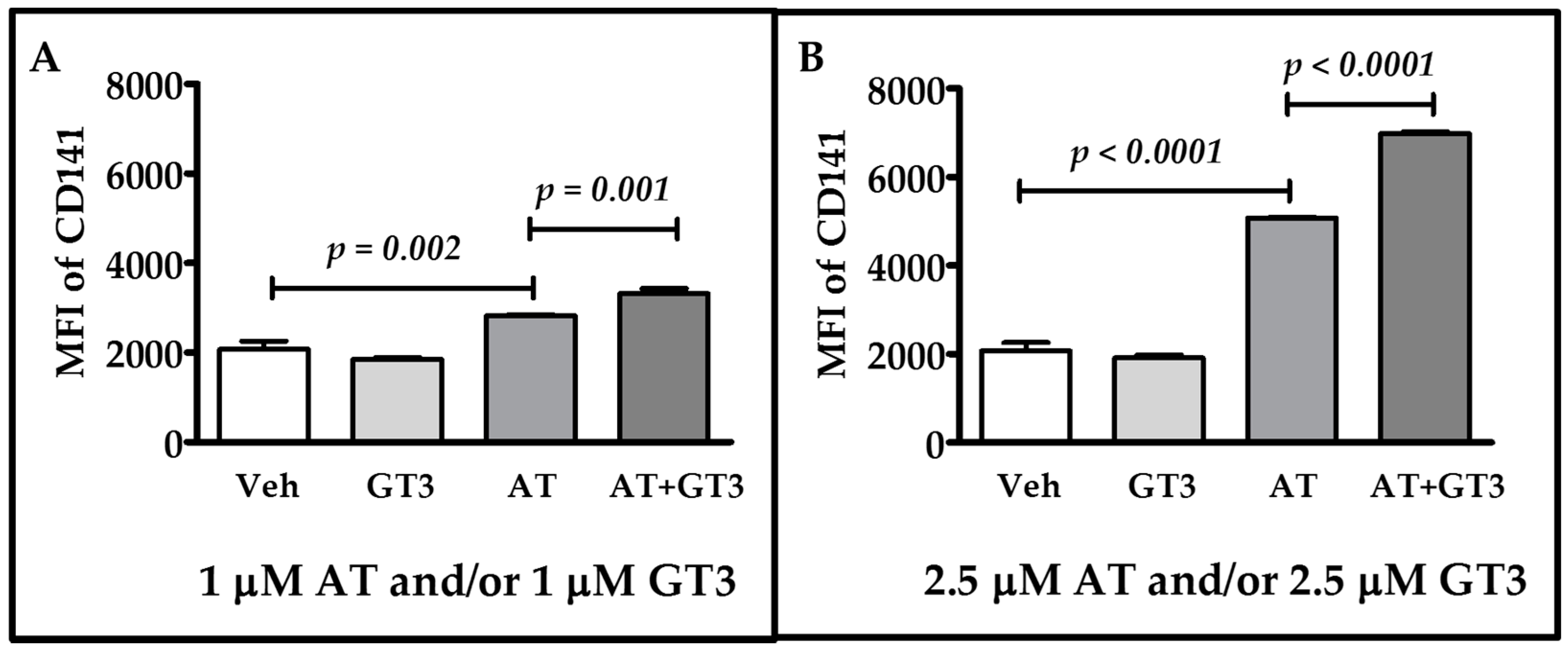
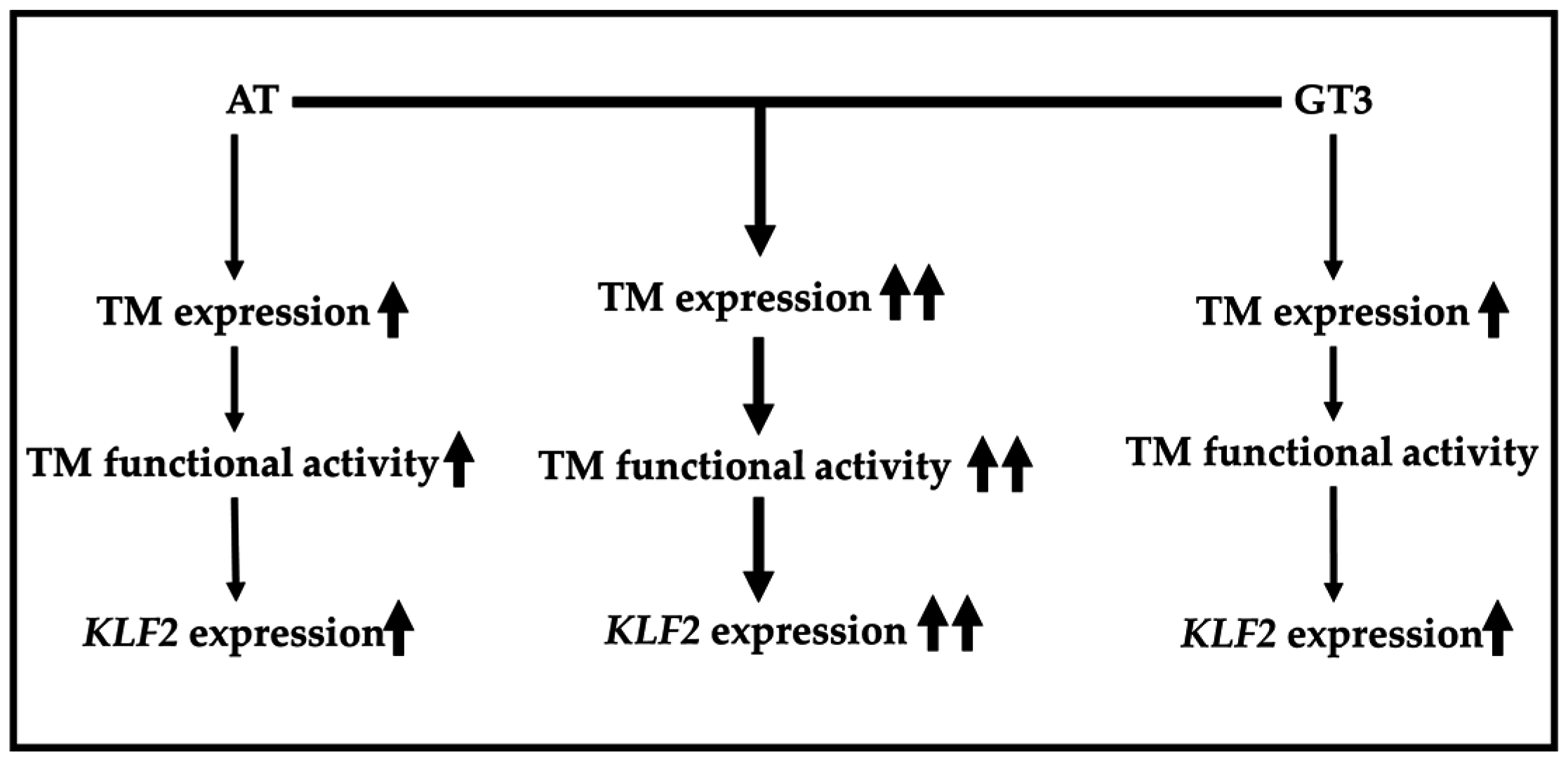
2.2. Atorvastatin and GT3 Synergistically Up-Regulate TM Expression
2.3. Atorvastatin and GT3 Synergistically Induce TM Functional Activity
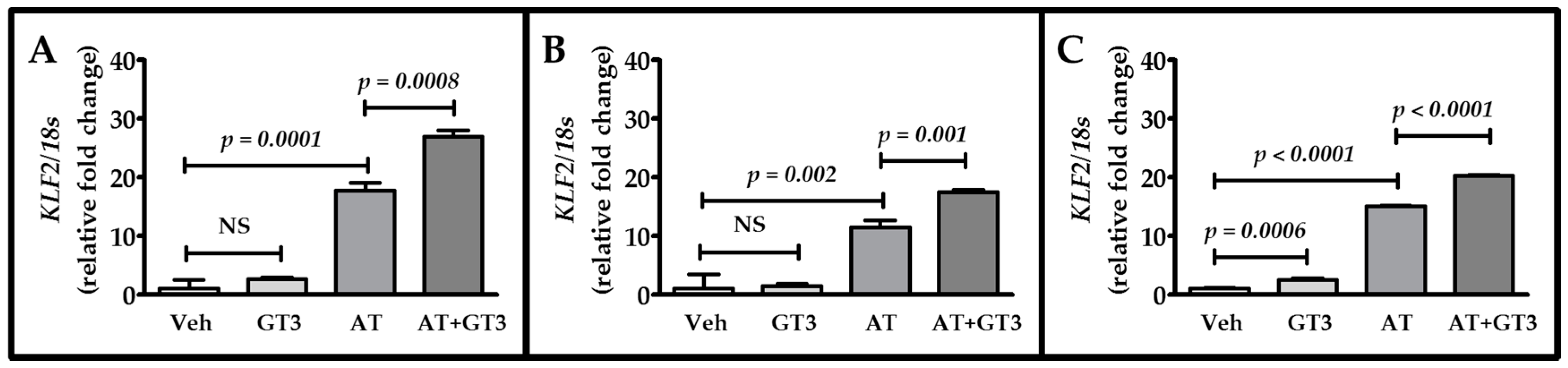
2.4. Atorvastatin and GT3 Synergistically Up-Regulate KLF2 Expression
3. Discussion
4. Materials and Methods
4.1. Cell Line and Reagents
4.2. Cell Culture and Drug Treatment
4.3. Flow Cytometry
4.4. APC Generation Assay
4.5. Total RNA Extraction, cDNA Preparation, and qRT-PCR by Taqman Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial function and dysfunction: Testing and clinical relevance. Circulation 2007, 115, 1285–1295. [Google Scholar] [PubMed]
- Weiler, H.; Isermann, B.H. Thrombomodulin. J. Thromb. Haemost. 2003, 1, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.M. Thrombomodulin and its role in inflammation. Semin. Immunopathol. 2012, 34, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Esmon, C.T. The protein C pathway. Chest 2003, 124, 26–32. [Google Scholar] [CrossRef]
- Van de Wouwer, M.; Conway, E.M. Novel functions of thrombomodulin in inflammation. Crit. Care Med. 2004, 32, 254–261. [Google Scholar] [CrossRef]
- Kim, H.; Hawthorne, W.J.; Kang, H.J.; Lee, Y.J.; Hwang, J.I.; Hurh, S.; Ro, H.; Jeong, J.C.; Cho, B.; Yang, J.; et al. Human thrombomodulin regulates complement activation as well as the coagulation cascade in xeno-immune response. Xenotransplantation 2015, 22, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, K.; Ogura, H.; Fujimi, S.; Morikawa, M.; Ogawa, Y.; Mohri, T.; Nakamori, Y.; Inoue, Y.; Kuwagata, Y.; Tanaka, H.; et al. Recombinant human soluble thrombomodulin in sepsis-induced disseminated intravascular coagulation: A multicenter propensity score analysis. Intensive Care Med. 2013, 39, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Geiger, H.; Pawar, S.A.; Kerschen, E.J.; Nattamai, K.J.; Hernandez, I.; Liang, H.P.; Fernandez, J.A.; Cancelas, J.A.; Ryan, M.A.; Kustikova, O.; et al. Pharmacological targeting of the thrombomodulin-activated protein C pathway mitigates radiation toxicity. Nat. Med. 2012, 18, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, J.; Zheng, H.; Ling, W.; Joseph, J.; Li, D.; Mehta, J.L.; Ponnappan, U.; Lin, P.; Fink, L.M.; et al. Statins increase thrombomodulin expression and function in human endothelial cells by a nitric oxide-dependent mechanism and counteract tumor necrosis factor α-induced thrombomodulin downregulation. Blood Coagul. Fibrinolysis 2003, 14, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Goralczyk, K.; Soszynska, K.; Haus, O.; Bielis, R.; Rosc, D. The influence of lovastatin on thrombomodulin gene expression in vascular endothelial cells—In vitro study. Folia Histochem. Cytobiol. 2009, 47, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Masamura, K.; Oida, K.; Kanehara, H.; Suzuki, J.; Horie, S.; Ishii, H.; Miyamori, I. Pitavastatin-induced thrombomodulin expression by endothelial cells acts via inhibition of small G proteins of the Rho family. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Boerma, M.; Burton, G.R.; Wang, J.; Fink, L.M.; McGehee, R.E.; Hauer-Jensen, M. Comparative expression profiling in primary and immortalized endothelial cells: Changes in gene expression in response to hydroxy methylglutaryl-coenzyme A reductase inhibition. Blood Coagul. Fibrinolysis 2006, 17, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.M.; Nambudiri, V.; Dai, G.; Larman, H.B.; Gimbrone, M.A.; Garcia-Cardena, G. Statins exert endothelial atheroprotective effects via the KLF2 transcription factor. J. Biol. Chem. 2005, 280, 26714–26719. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Wang, J.; Boerma, M.; Berbee, M.; Qiu, X.; Fink, L.M.; Hauer-Jensen, M. Involvement of heat shock factor 1 in statin-induced transcriptional upregulation of endothelial thrombomodulin. Circ. Res. 2008, 103, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.; Rouleau, L.; Tardif, J.C.; Leask, R.L. Effect of simvastatin on Kruppel-like factor 2, endothelial nitric oxide synthase and thrombomodulin expression in endothelial cells under shear stress. Life Sci. 2010, 87, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Hsieh, F.Y.; Chen, Y.H.; Lin, C.C.; Kuan, I.I.; Wang, S.H.; Wu, C.C.; Chien, H.F.; Lin, F.Y.; Chen, Y.L. Atorvastatin induces thrombomodulin expression in the aorta of cholesterol-fed rabbits and in TNFα-treated human aortic endothelial cells. Histol. Histopathol. 2009, 24, 1147–1159. [Google Scholar] [PubMed]
- Lin, S.J.; Chen, Y.H.; Lin, F.Y.; Hsieh, L.Y.; Wang, S.H.; Lin, C.Y.; Wang, Y.C.; Ku, H.H.; Chen, J.W.; Chen, Y.L. Pravastatin induces thrombomodulin expression in TNFα-treated human aortic endothelial cells by inhibiting Rac1 and Cdc42 translocation and activity. J. Cell. Biochem. 2007, 101, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, P.; Aillaud, M.F.; Habib, G.; Kreitmann, B.; Metras, D.; Luccioni, R.; Bouvenot, G.; Juhan-Vague, I. Fluvastatin decreases soluble thrombomodulin in cardiac transplant recipients. Thromb. Haemost. 2000, 83, 46–48. [Google Scholar] [PubMed]
- Seljeflot, I.; Tonstad, S.; Hjermann, I.; Arnesen, H. Reduced expression of endothelial cell markers after 1 year treatment with simvastatin and atorvastatin in patients with coronary heart disease. Atherosclerosis 2002, 162, 179–185. [Google Scholar] [CrossRef]
- Liao, J.K.; Laufs, U. Pleiotropic effects of statins. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 89–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Liao, J.K. Pleiotropic effects of statins: Basic research and clinical perspectives. Circ. J. 2010, 74, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.Z.; Ran, X.; Zong, Z.W.; Liu, D.Q.; Xiang, G.M.; Su, Y.P.; Zheng, H.E. Protective effect of atorvastatin on radiation-induced vascular endothelial cell injury in vitro. J. Radiat. Res. 2010, 51, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Boerma, M.; Fu, Q.; Kulkarni, A.; Fink, L.M.; Hauer-Jensen, M. Simvastatin ameliorates radiation enteropathy development after localized, fractionated irradiation by a protein C-independent mechanism. Int. J. Radiat. Oncol. Biol. Phys. 2007, 68, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- De Castro, M.L.; Hermo, J.A.; Baz, A.; de, L.C.; Perez, R.; Clofent, J. Acute cholestatic hepatitis after atorvastatin reintroduction. Gastroenterol. Hepatol. 2006, 29, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Pelli, N.; Setti, M.; Ceppa, P.; Toncini, C.; Indiveri, F. Autoimmune hepatitis revealed by atorvastatin. Eur. J. Gastroenterol. Hepatol. 2003, 15, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Billings, F.T.; Hendricks, P.A.; Schildcrout, J.S.; Shi, Y.; Petracek, M.R.; Byrne, J.G.; Brown, N.J. High-dose perioperative atorvastatin and acute kidney injury following cardiac surgery: A randomized clinical trial. JAMA 2016, 315, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Shao, L.; Ghosh, S.P.; Zhou, D.; Boerma, M.; Weiler, H.; Hauer-Jensen, M. Thrombomodulin contributes to γ-tocotrienol-mediated lethality protection and hematopoietic cell recovery in irradiated mice. PLoS ONE 2015, 10, e0122511. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.A.; Pearce, B.C.; Clark, R.W.; Gordon, D.A.; Wright, J.J. Tocotrienols regulate cholesterol production in mammalian cells by post-transcriptional suppression of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Biol. Chem. 1993, 268, 11230–11238. [Google Scholar] [PubMed]
- Swift, S.N.; Pessu, R.L.; Chakraborty, K.; Villa, V.; Lombardini, E.; Ghosh, S.P. Acute toxicity of subcutaneously administered vitamin E isomers δ- and γ-tocotrienol in mice. Int. J. Toxicol. 2014, 33, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Mukherjee, S.; Lekli, I.; Gurusamy, N.; Bardhan, J.; Raychoudhury, U.; Chakravarty, R.; Banerji, S.; Knowlton, A.A.; Das, D.K. Tocotrienols confer resistance to ischemia in hypercholesterolemic hearts: Insight with genomics. Mol. Cell. Biochem. 2012, 360, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Newaz, M.A.; Nawal, N.N. Effect of γ-tocotrienol on blood pressure, lipid peroxidation and total antioxidant status in spontaneously hypertensive rats (SHR). Clin. Exp. Hypertens. 1999, 21, 1297–1313. [Google Scholar] [PubMed]
- Deng, L.; Ding, Y.; Peng, Y.; Wu, Y.; Fan, J.; Li, W.; Yang, R.; Yang, M.; Fu, Q. γ-Tocotrienol protects against ovariectomy-induced bone loss via mevalonate pathway as HMG-CoA reductase inhibitor. Bone 2014, 67, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.; Fu, Q.; Boerma, M.; Wang, J.; Kumar, K.S.; Hauer-Jensen, M. γ-Tocotrienol ameliorates intestinal radiation injury and reduces vascular oxidative stress after total-body irradiation by an HMG-CoA reductase-dependent mechanism. Radiat. Res. 2009, 171, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Weiler, H. Multiple receptor-mediated functions of activated protein C. Hamostaseologie 2011, 31, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Dahlback, B.; Villoutreix, B.O. The anticoagulant protein C pathway. FEBS Lett. 2005, 579, 3310–3316. [Google Scholar] [CrossRef] [PubMed]
- Elphick, G.F.; Sarangi, P.P.; Hyun, Y.M.; Hollenbaugh, J.A.; Ayala, A.; Biffl, W.L.; Chung, H.L.; Rezaie, A.R.; McGrath, J.L.; Topham, D.J.; et al. Recombinant human activated protein C inhibits integrin-mediated neutrophil migration. Blood 2009, 113, 4078–4085. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Yang, L.; Rezaie, A.R. Receptors of the protein C activation and activated protein C signaling pathways are colocalized in lipid rafts of endothelial cells. Proc. Natl. Acad. Sci. USA 2007, 104, 2867–2872. [Google Scholar] [CrossRef] [PubMed]
- Ludeman, M.J.; Kataoka, H.; Srinivasan, Y.; Esmon, N.L.; Esmon, C.T.; Coughlin, S.R. PAR1 cleavage and signaling in response to activated protein C and thrombin. J. Biol. Chem. 2005, 280, 13122–13128. [Google Scholar] [CrossRef] [PubMed]
- White, T.C.; Berny, M.A.; Tucker, E.I.; Urbanus, R.T.; de Groot, P.G.; Fernandez, J.A.; Griffin, J.H.; Gruber, A.; McCarty, O.J. Protein C supports platelet binding and activation under flow: Role of glycoprotein Ib and apolipoprotein E receptor 2. J. Thromb. Haemost. 2008, 6, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, S.; Gerlach, H. Activated protein C for sepsis. N. Engl. J. Med. 2009, 361, 2646–2652. [Google Scholar] [CrossRef] [PubMed]
- Sen-Banerjee, S.; Mir, S.; Lin, Z.; Hamik, A.; Atkins, G.B.; Das, H.; Banerjee, P.; Kumar, A.; Jain, M.K. Kruppel-like factor 2 as a novel mediator of statin effects in endothelial cells. Circulation 2005, 112, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Kumar, A.; SenBanerjee, S.; Staniszewski, K.; Parmar, K.; Vaughan, D.E.; Gimbrone, M.A., Jr.; Balasubramanian, V.; Garcia-Cardena, G.; Jain, M.K. Kruppel-like factor 2 (KLF2) regulates endothelial thrombotic function. Circ. Res. 2005, 96, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Shao, L.; Chafekar, S.M.; Feng, W.; Ponnappan, U.; Fink, L.M.; Zhou, D.; Hauer-Jensen, M. IKKbeta regulates endothelial thrombomodulin in a KLF2-dependent manner. J. Thromb. Haemost. 2014, 12, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
- Le, N.T.; Takei, Y.; Izawa-Ishizawa, Y.; Heo, K.S.; Lee, H.; Smrcka, A.V.; Miller, B.L.; Ko, K.A.; Ture, S.; Morrell, C.; et al. Identification of activators of ERK5 transcriptional activity by high-throughput screening and the role of endothelial ERK5 in vasoprotective effects induced by statins and antimalarial agents. J. Immunol. 2014, 193, 3803–3815. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.L.; Kim, J.; Kang, Y.; Shi, H.; Atkins, G.B.; Jain, M.K.; Chun, H.J. Apelin/APJ signaling is a critical regulator of statin effects in vascular endothelial cells—Brief report. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2640–2643. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pathak, R.; Ghosh, S.P.; Zhou, D.; Hauer-Jensen, M. The Vitamin E Analog Gamma-Tocotrienol (GT3) and Statins Synergistically Up-Regulate Endothelial Thrombomodulin (TM). Int. J. Mol. Sci. 2016, 17, 1937. https://doi.org/10.3390/ijms17111937
Pathak R, Ghosh SP, Zhou D, Hauer-Jensen M. The Vitamin E Analog Gamma-Tocotrienol (GT3) and Statins Synergistically Up-Regulate Endothelial Thrombomodulin (TM). International Journal of Molecular Sciences. 2016; 17(11):1937. https://doi.org/10.3390/ijms17111937
Chicago/Turabian StylePathak, Rupak, Sanchita P. Ghosh, Daohong Zhou, and Martin Hauer-Jensen. 2016. "The Vitamin E Analog Gamma-Tocotrienol (GT3) and Statins Synergistically Up-Regulate Endothelial Thrombomodulin (TM)" International Journal of Molecular Sciences 17, no. 11: 1937. https://doi.org/10.3390/ijms17111937
APA StylePathak, R., Ghosh, S. P., Zhou, D., & Hauer-Jensen, M. (2016). The Vitamin E Analog Gamma-Tocotrienol (GT3) and Statins Synergistically Up-Regulate Endothelial Thrombomodulin (TM). International Journal of Molecular Sciences, 17(11), 1937. https://doi.org/10.3390/ijms17111937