
NOD Scid Gamma Mice Are Permissive to Allogeneic HSC Transplantation without Prior Conditioning



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
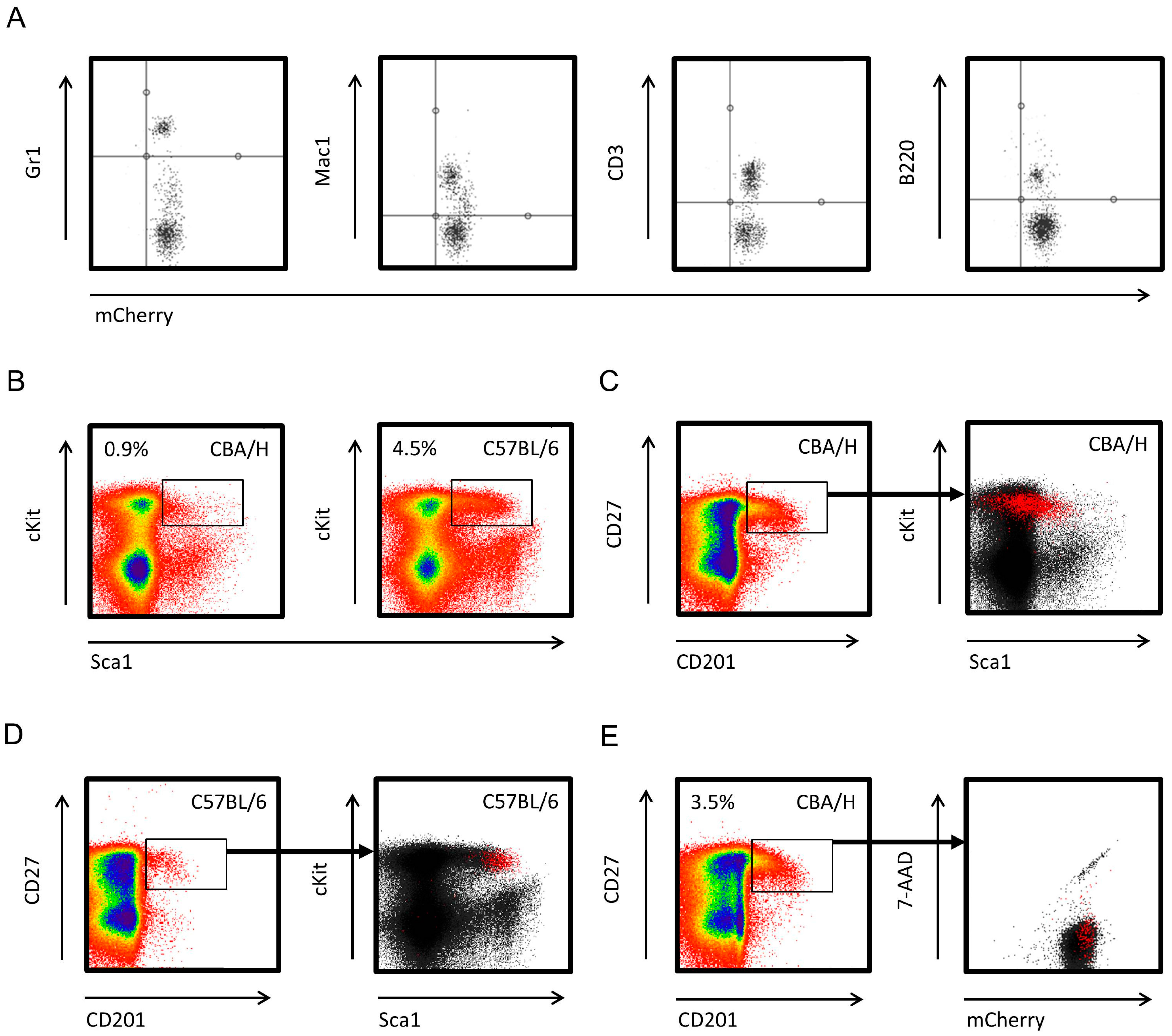
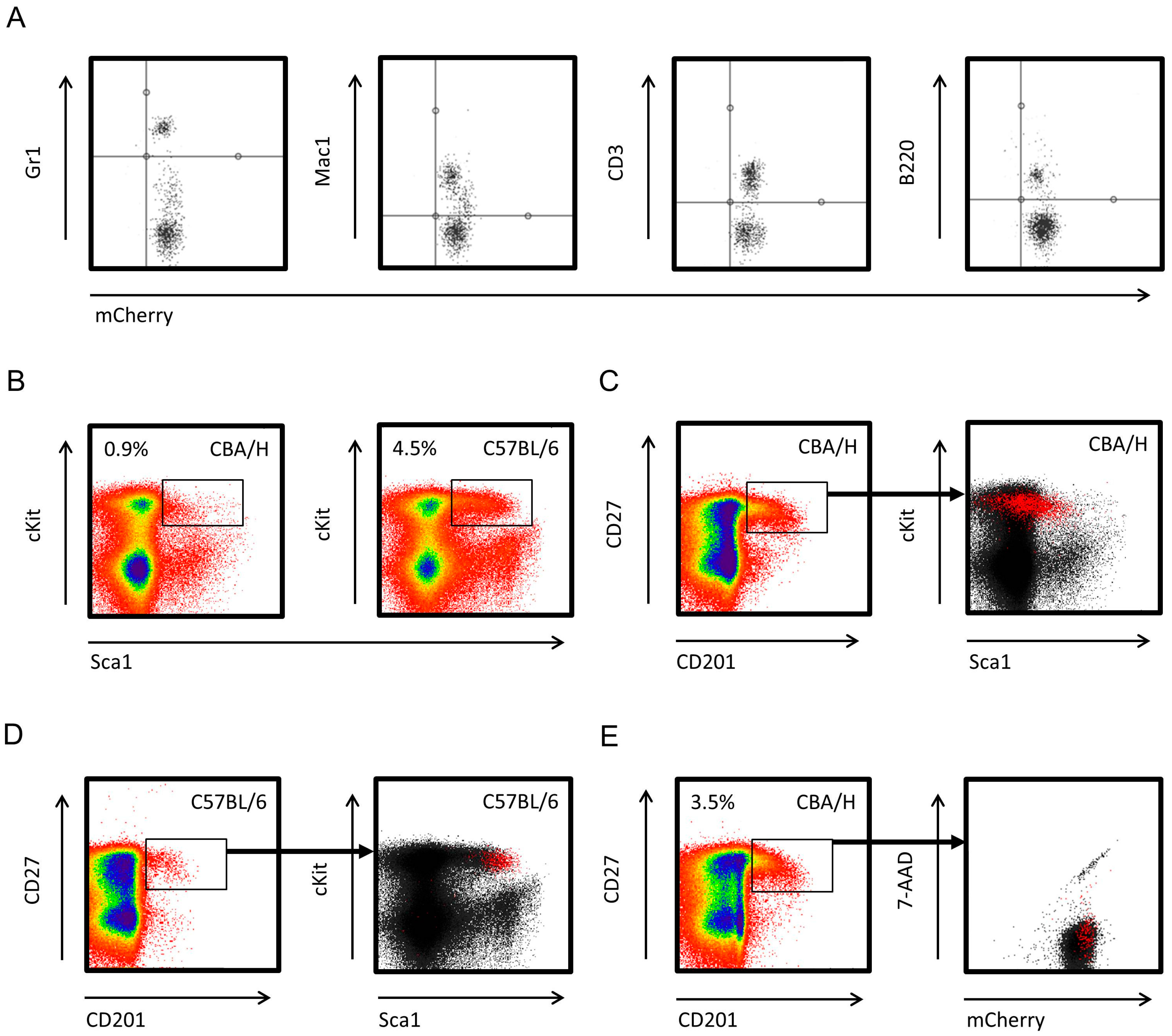
2.1. CBA/HmCherry Mice Constitutively Express mCherry across HSCs and Downstream Hematopoietic Lineages
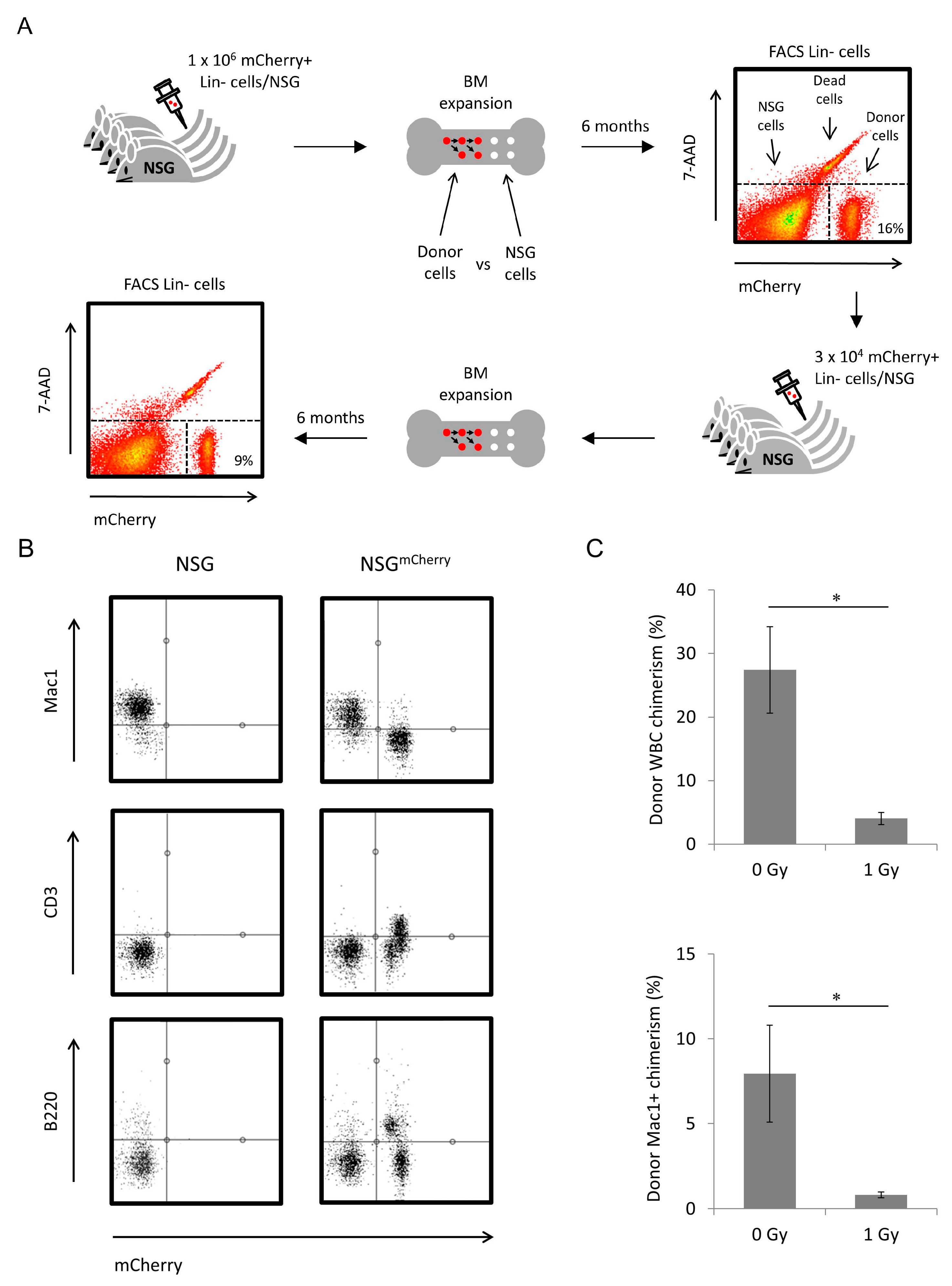
2.2. NOD Scid Gamma Bone Marrow (BM) Microenvironment Supports Allogeneic Hematopoietic Stem Cell (HSC) Engraftment and Differentiation
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Long-Term Transplantation Assays
4.3. Flow Cytometry
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Major, I.R.; Mole, R.H. Myeloid leukaemia in X-ray irradiated CBA mice. Nature 1978, 272, 455–456. [Google Scholar] [CrossRef] [PubMed]
- Verbiest, T.; Bouffler, S.; Nutt, S.L.; Badie, C. PU.1 downregulation in murine radiation-induced acute myeloid leukaemia (AML): From molecular mechanism to human AML. Carcinogenesis 2015, 36, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Mcculloch, E.A.; Till, J.E. The radiation sensitivity of normal mouse bone marrow cells, determined by quantitative marrow transplantation into irradiated mice. Radiat. Res. 1960, 13, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Olme, C.-H.; Brown, N.; Finnon, R.; Bouffler, S.D.; Badie, C. Frequency of acute myeloid leukaemia-associated mouse chromosome 2 deletions in X-ray exposed immature haematopoietic progenitors and stem cells. Mutat. Res. 2013, 756, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Duran-Struuck, R.; Dysko, R.C. Principles of bone marrow transplantation (BMT): Providing optimal veterinary and husbandry care to irradiated mice in BMT studies. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 11–22. [Google Scholar] [PubMed]
- Mathews, J.D.; Forsythe, A.V.; Brady, Z.; Butler, M.W.; Goergen, S.K.; Byrnes, G.B.; Giles, G.G.; Wallace, A.B.; Anderson, P.R.; Guiver, T.A.; et al. Cancer risk in 680,000 people exposed to computed tomography scans in childhood or adolescence: Data linkage study of 11 million Australians. BMJ 2013, 346, f2360. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.S.; Salotti, J.A.; Little, M.P.; McHugh, K.; Lee, C.; Kim, K.P.; Howe, N.L.; Ronckers, C.M.; Rajaraman, P.; Craft, A.W.; et al. Radiation exposure from CT scans in childhood and subsequent risk of leukaemia and brain tumours: A retrospective cohort study. Lancet 2012, 380, 499–505. [Google Scholar] [CrossRef]
- Krestinina, L.Y.; Davis, F.G.; Schonfeld, S.; Preston, D.L.; Degteva, M.; Epifanova, S.; Akleyev, A.V. Leukaemia incidence in the Techa River Cohort: 1953–2007. Br. J. Cancer 2013, 109, 2886–2893. [Google Scholar] [CrossRef] [PubMed]
- Leuraud, K.; Richardson, D.B.; Cardis, E.; Daniels, R.D.; Gillies, M.; O’Hagan, J.A.; Hamra, G.B.; Haylock, R.; Laurier, D.; Moissonnier, M.; et al. Ionising radiation and risk of death from leukaemia and lymphoma in radiation-monitored workers (INWORKS): An international cohort study. Lancet Haematol. 2015, 2, e276–e281. [Google Scholar] [CrossRef]
- Kendall, G.M.; Little, M.P.; Wakeford, R.; Bunch, K.J.; Miles, J.C.H.; Vincent, T.J.; Meara, J.R.; Murphy, M.F.G. A record-based case-control study of natural background radiation and the incidence of childhood leukaemia and other cancers in Great Britain during 1980–2006. Leukemia 2013, 27, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Spycher, B.D.; Lupatsch, J.E.; Zwahlen, M.; Röösli, M.; Niggli, F.; Grotzer, M.A.; Rischewski, J.; Egger, M.; Kuehni, C.E. Background ionizing radiation and the risk of childhood cancer: A census-based nationwide cohort study. Environ. Health Perspect. 2015, 123, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Nikkilä, A.; Erme, S.; Arvela, H.; Holmgren, O.; Raitanen, J.; Lohi, O.; Auvinen, A. Background radiation and childhood leukemia: A nationwide register-based case-control study. Int. J. Cancer 2016, 139, 1975–1982. [Google Scholar] [CrossRef] [PubMed]
- Qing, Y.; Lin, Y.; Gerson, S.L. An intrinsic BM hematopoietic niche occupancy defect of HSC in scid mice facilitates exogenous HSC engraftment. Blood 2012, 119, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, M.; Gaskins, H.R.; Shultz, L.D.; Leiter, E.H. The nonobese diabetic scid mouse: Model for spontaneous thymomagenesis associated with immunodeficiency. Proc. Natl. Acad. Sci. USA 1992, 89, 3290–3294. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R γ null mice engrafted with mobilized human hemopoietic stem cells. J. Immunol. Baltim. Md 1950 2005, 174, 6477–6489. [Google Scholar] [CrossRef]
- Ishikawa, F.; Yasukawa, M.; Lyons, B.; Yoshida, S.; Miyamoto, T.; Yoshimoto, G.; Watanabe, T.; Akashi, K.; Shultz, L.D.; Harada, M. Development of functional human blood and immune systems in NOD/scid/IL2 receptor γ chain(null) mice. Blood 2005, 106, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Spangrude, G.J.; Brooks, D.M. Mouse strain variability in the expression of the hematopoietic stem cell antigen Ly-6A/E by bone marrow cells. Blood 1993, 82, 3327–3332. [Google Scholar] [PubMed]
- Vazquez, S.E.; Inlay, M.A.; Serwold, T. CD201 and CD27 identify hematopoietic stem and progenitor cells across multiple murine strains independently of Kit and Sca-1. Exp. Hematol. 2015, 43, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.; Ge, S.; Symbatyan, G.; Rosol, M.S.; Olch, A.J.; Crooks, G.M. Effects of sublethal irradiation on patterns of engraftment after murine bone marrow transplantation. Biol. Blood Marrow Transplant. 2011, 17, NN608–NN619. [Google Scholar] [CrossRef] [PubMed]
- Futrega, K.; Lott, W.B.; Doran, M.R. Direct bone marrow HSC transplantation enhances local engraftment at the expense of systemic engraftment in NSG mice. Sci. Rep. 2016, 6, 23886. [Google Scholar] [CrossRef] [PubMed]
- Brehm, M.A.; Racki, W.J.; Leif, J.; Burzenski, L.; Hosur, V.; Wetmore, A.; Gott, B.; Herlihy, M.; Ignotz, R.; Dunn, R.; et al. Engraftment of human HSCs in nonirradiated newborn NOD-scid IL2rγ null mice is enhanced by transgenic expression of membrane-bound human SCF. Blood 2012, 119, 2778–2788. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, B.E.; Brown, M.E.; Duffin, B.M.; Maufort, J.P.; Vereide, D.T.; Slukvin, I.I.; Thomson, J.A. Nonirradiated NOD,B6.SCID Il2rγ−/− Kit(W41/W41) (NBSGW) mice support multilineage engraftment of human hematopoietic cells. Stem Cell Rep. 2015, 4, 171–180. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verbiest, T.; Finnon, R.; Brown, N.; Finnon, P.; Bouffler, S.; Badie, C. NOD Scid Gamma Mice Are Permissive to Allogeneic HSC Transplantation without Prior Conditioning. Int. J. Mol. Sci. 2016, 17, 1850. https://doi.org/10.3390/ijms17111850
Verbiest T, Finnon R, Brown N, Finnon P, Bouffler S, Badie C. NOD Scid Gamma Mice Are Permissive to Allogeneic HSC Transplantation without Prior Conditioning. International Journal of Molecular Sciences. 2016; 17(11):1850. https://doi.org/10.3390/ijms17111850
Chicago/Turabian StyleVerbiest, Tom, Rosemary Finnon, Natalie Brown, Paul Finnon, Simon Bouffler, and Christophe Badie. 2016. "NOD Scid Gamma Mice Are Permissive to Allogeneic HSC Transplantation without Prior Conditioning" International Journal of Molecular Sciences 17, no. 11: 1850. https://doi.org/10.3390/ijms17111850
APA StyleVerbiest, T., Finnon, R., Brown, N., Finnon, P., Bouffler, S., & Badie, C. (2016). NOD Scid Gamma Mice Are Permissive to Allogeneic HSC Transplantation without Prior Conditioning. International Journal of Molecular Sciences, 17(11), 1850. https://doi.org/10.3390/ijms17111850