
Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter

Abstract
:1. Introduction
2. Results
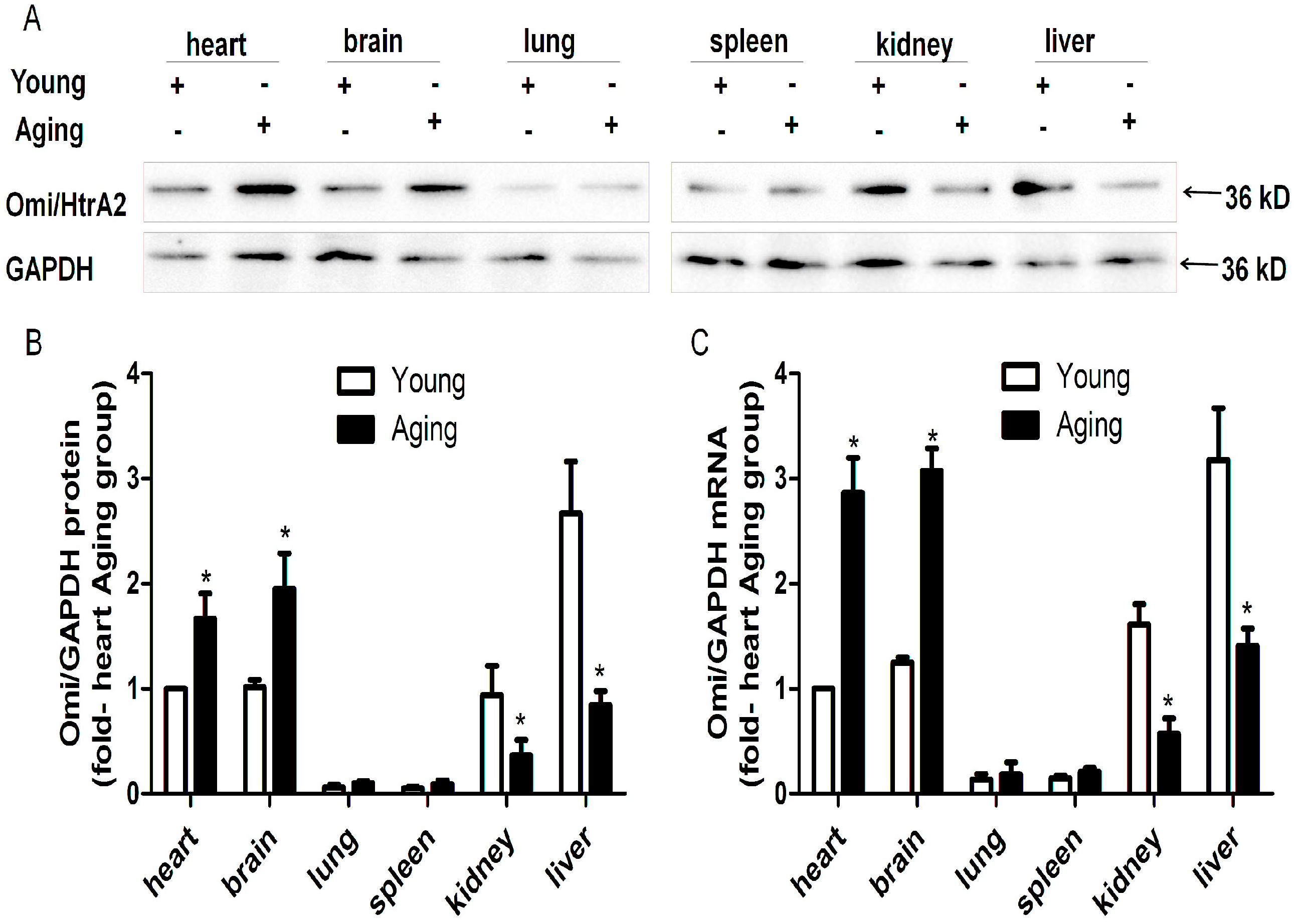
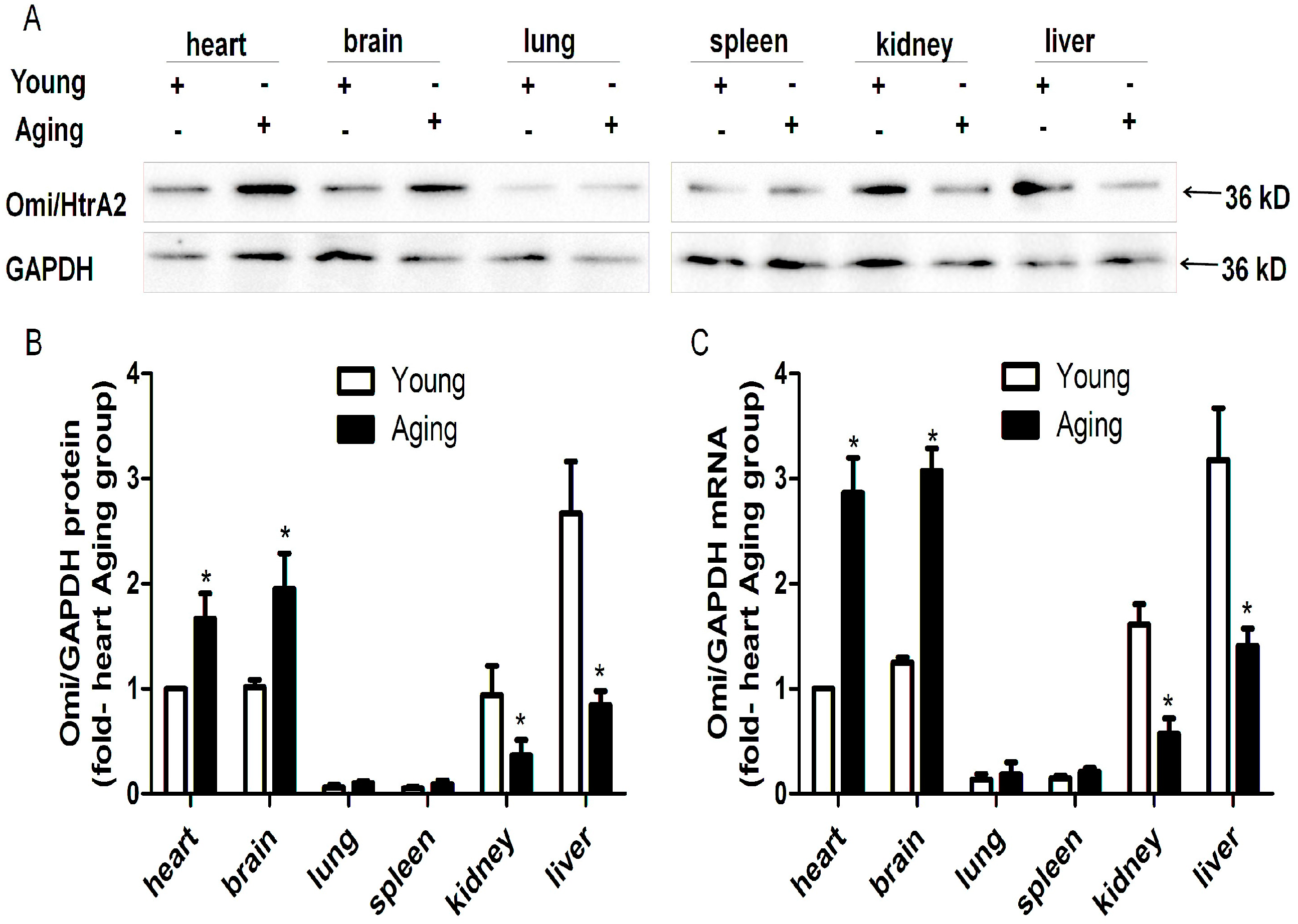
2.1. HtrA Serine Peptidase 2 (Omi/HtrA2) is Ubiquitously Expressed but Its Protein and mRNA Levels Vary in Different Tissues
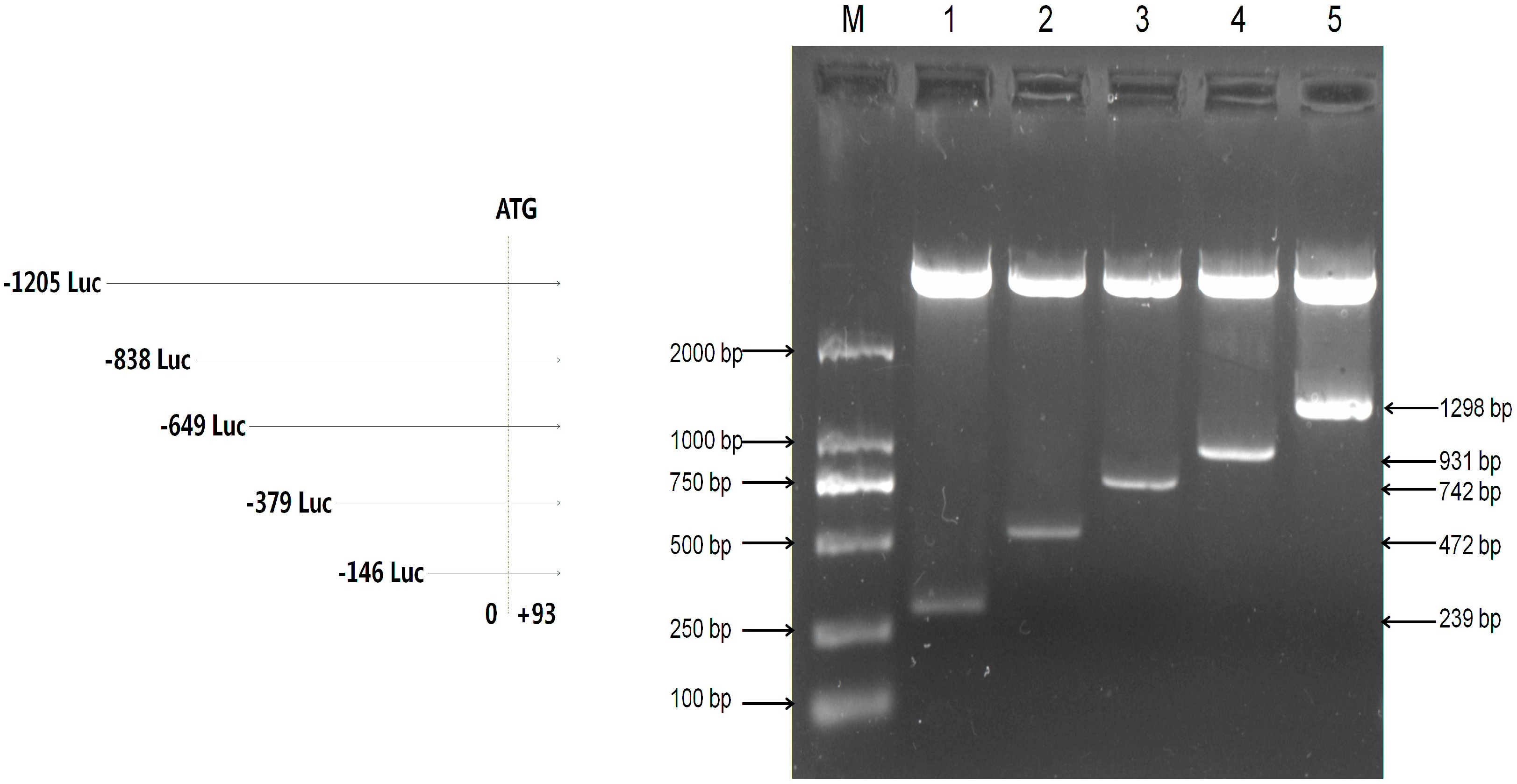
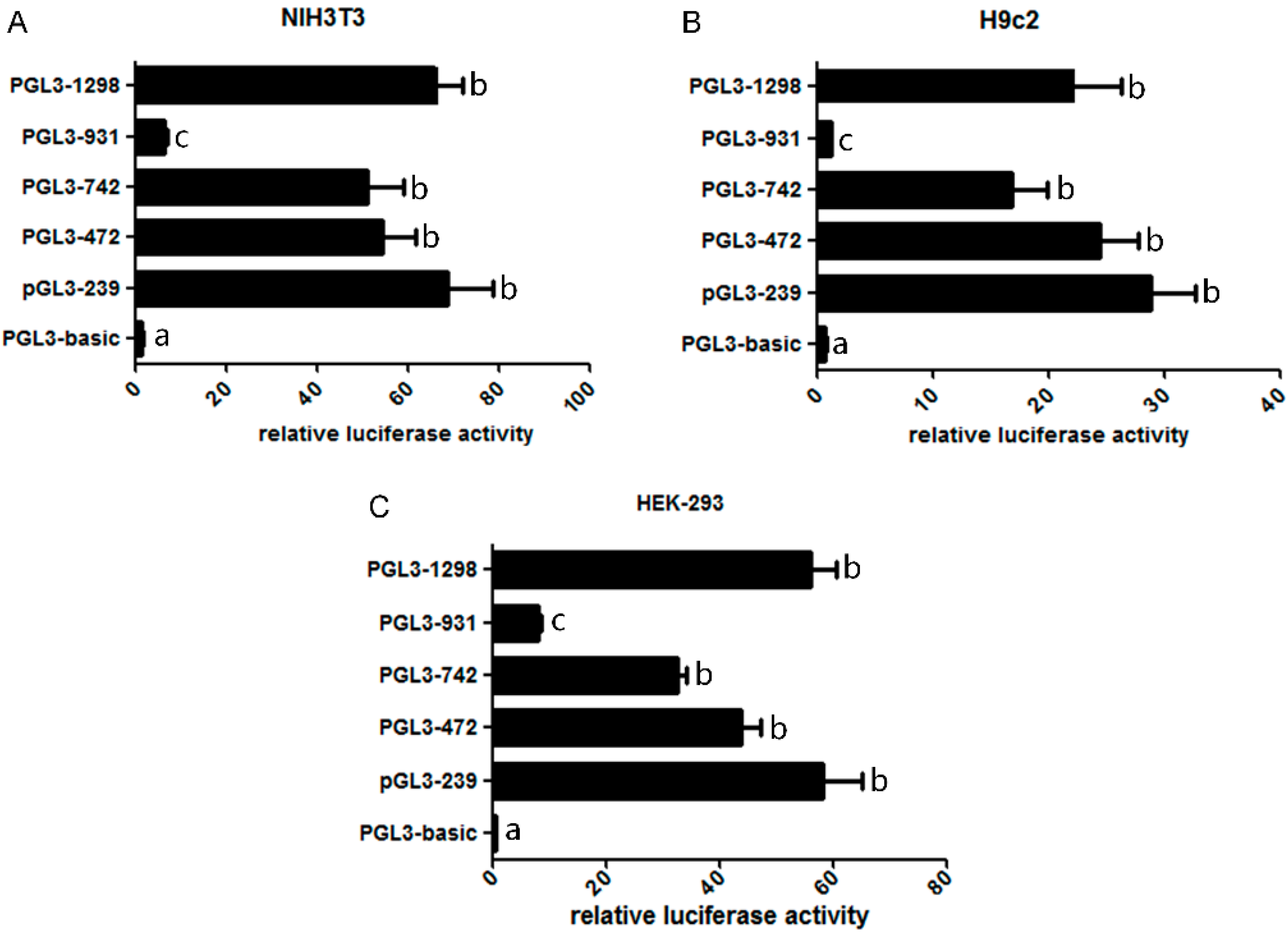
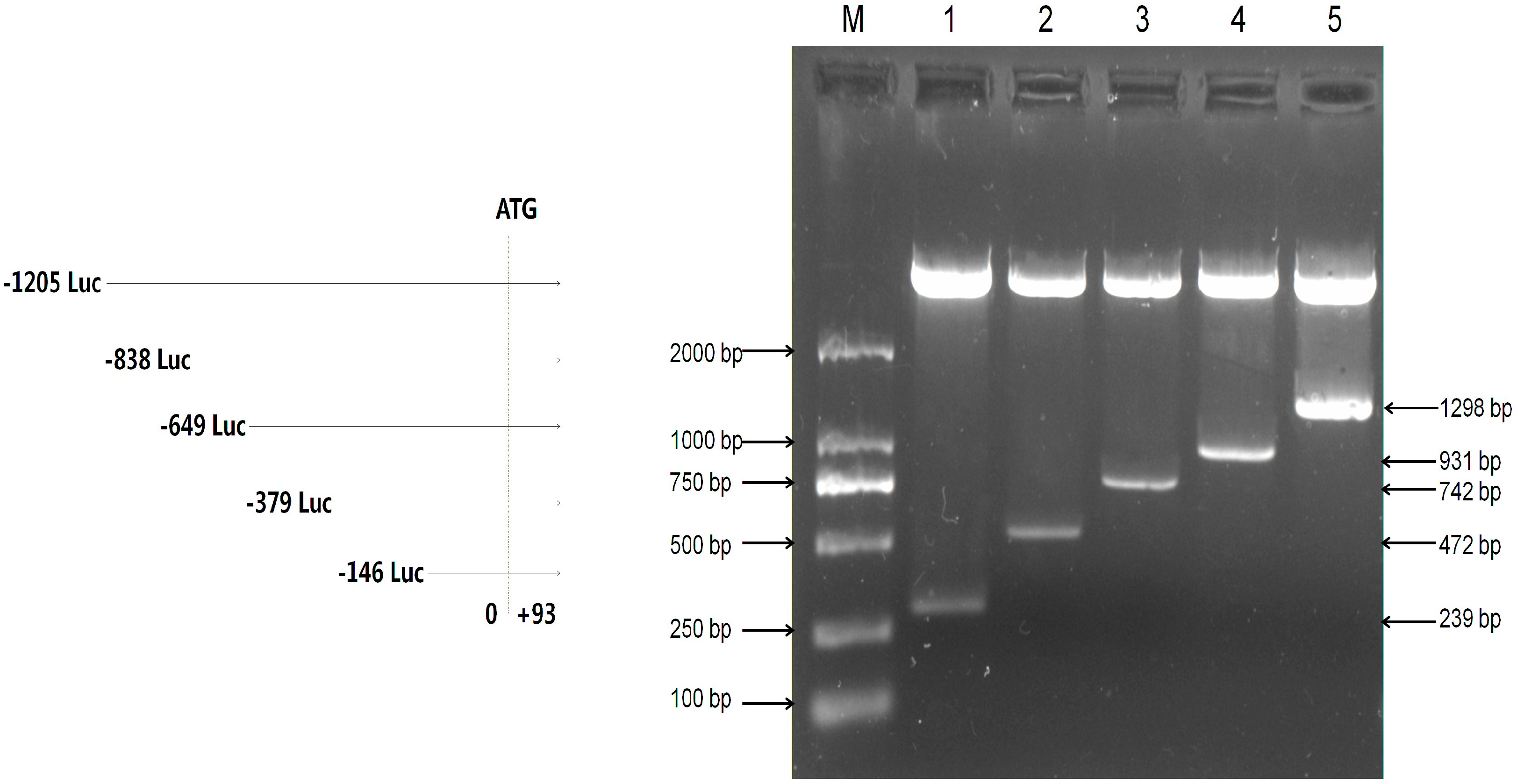
2.2. Assessment of the Activity of Mouse Omi/HtrA2 Promoter Luciferase Full-Length and Truncated Vectors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5’–3’) | Amplification Region (bp) | Length (bp) |
|---|---|---|---|
| F1 | AGAATTTCGAGGGCAGGGCTAA | −146~+93 | 239 |
| F2 | CGTGAGGTGGCGATTAAG | −379~+93 | 472 |
| F3 | TGGAAATAAGGACCCTGACG | −649~+93 | 742 |
| F4 | GTAGAGCAGTAGCGCGAGCA | −838~+93 | 931 |
| F5 | CTGGGAAGGCGGAGTCTTT | −1205~+93 | 1298 |
| R | GGCAAGCGGTCTAGGAGAA | – | – |
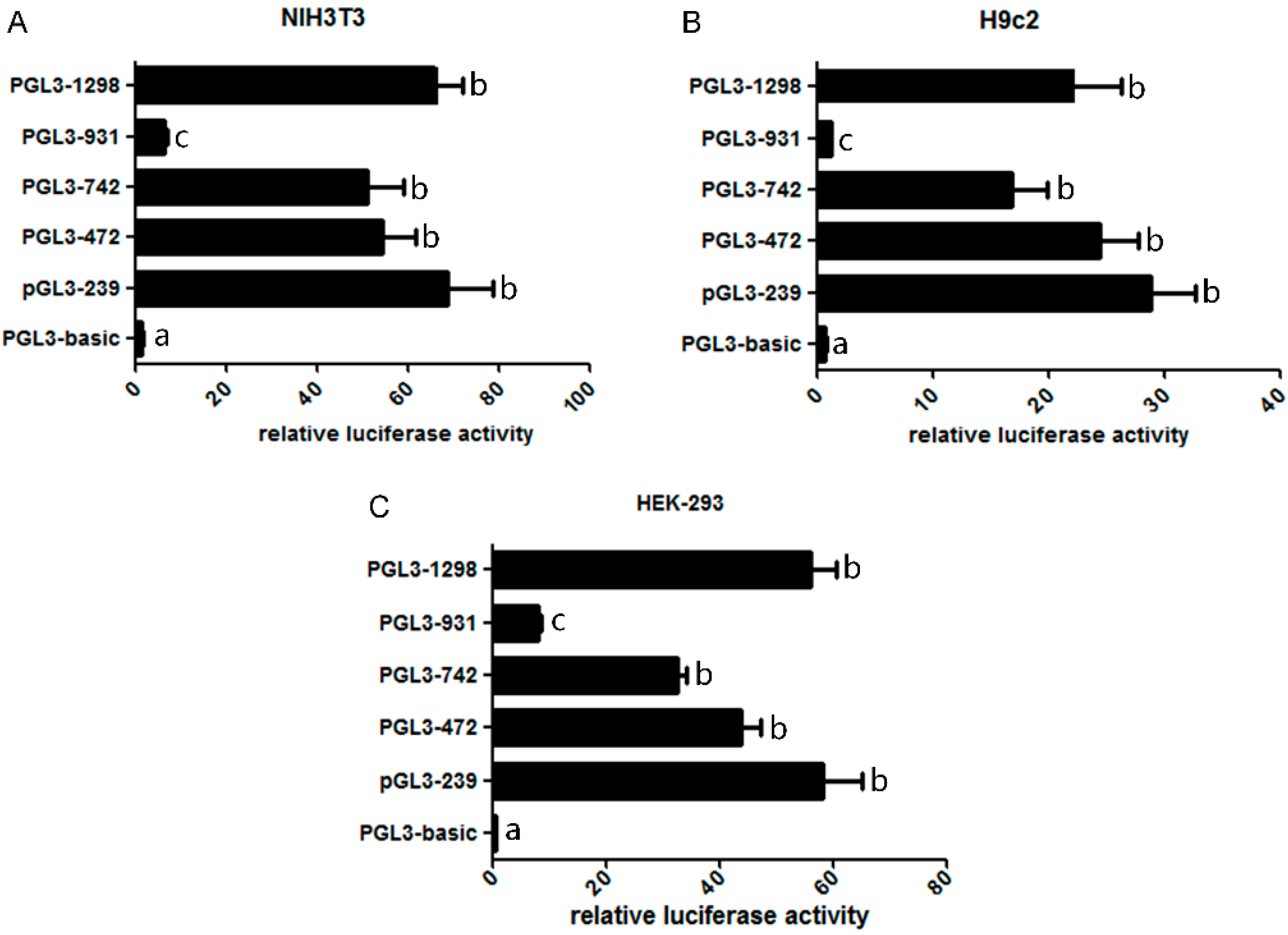
2.3. Bioinformatics Analysis of the Mouse Omi/HtrA2 Promoter
| Prediction Area | Start | End | Promoter Sequence | Score |
|---|---|---|---|---|
| promoter core area 1 | −1197 | −1148 | GAGGGCAGGGCTAAAAGTGGGCAGACAGGAAAGGAACTAGGGCACCCACT | 0.98 |
| promoter core area 2 | −359 | −310 | CCGGTGCGAGTCAAAGAGCCGCTCCGGCCCCGGAGCTGGGGGAGGTTCCA | 0.90 |
| CpG island | −709 | +37 | Observed/Expected ratio > 0.60 | Percent C + Percent G > 50.00 |
| Transcription Factors | Number | Sequence of Putative Binding Sites | Location |
|---|---|---|---|
| HSF1 | 1 | AAGGAACT | +83~+90 |
| 2 | GTGGAAGC | −46~−39 | |
| 3 | CTTCTTGCTTTTTCT | −172~−158 | |
| 4 | ATGGAAGG | −181~−174 | |
| 5 | GGGGAACA | −285~−278 | |
| 6 | AGGGAAGA | −805~−798 | |
| 7 | ATGGAACC | −868~−861 | |
| 8 | CACAGAAT | −1028~−1021 | |
| SP1 | 1 | GCCTCGCCCC | −722~−713 |
| 2 | CACCCGCCTAT | −999~−899 | |
| AP1 | 1 | CTATTGA | −944~−938 |
| p53 | 1 | CGTGCCC | +65~+71 |
| 2 | GCGGCCC | −54~−48 | |
| 3 | GGGCTAG | −84~−78 | |
| 4 | GGGCATC | −235~−229 | |
| 5 | CCAGCCCAGCCC | −520~−509 | |
| 6 | GGGCTGG | −816~−810 | |
| 7 | GCAGCCC | −1103~−1197 | |
| YY1 | 1 | GAAAAGACA | −867~−859 |
| 2 | AGGGTGACA | −843~−835 | |
| 3 | CTGGGGACA | −551~−543 | |
| 4 | TGTCGCCGC | −177~−169 |
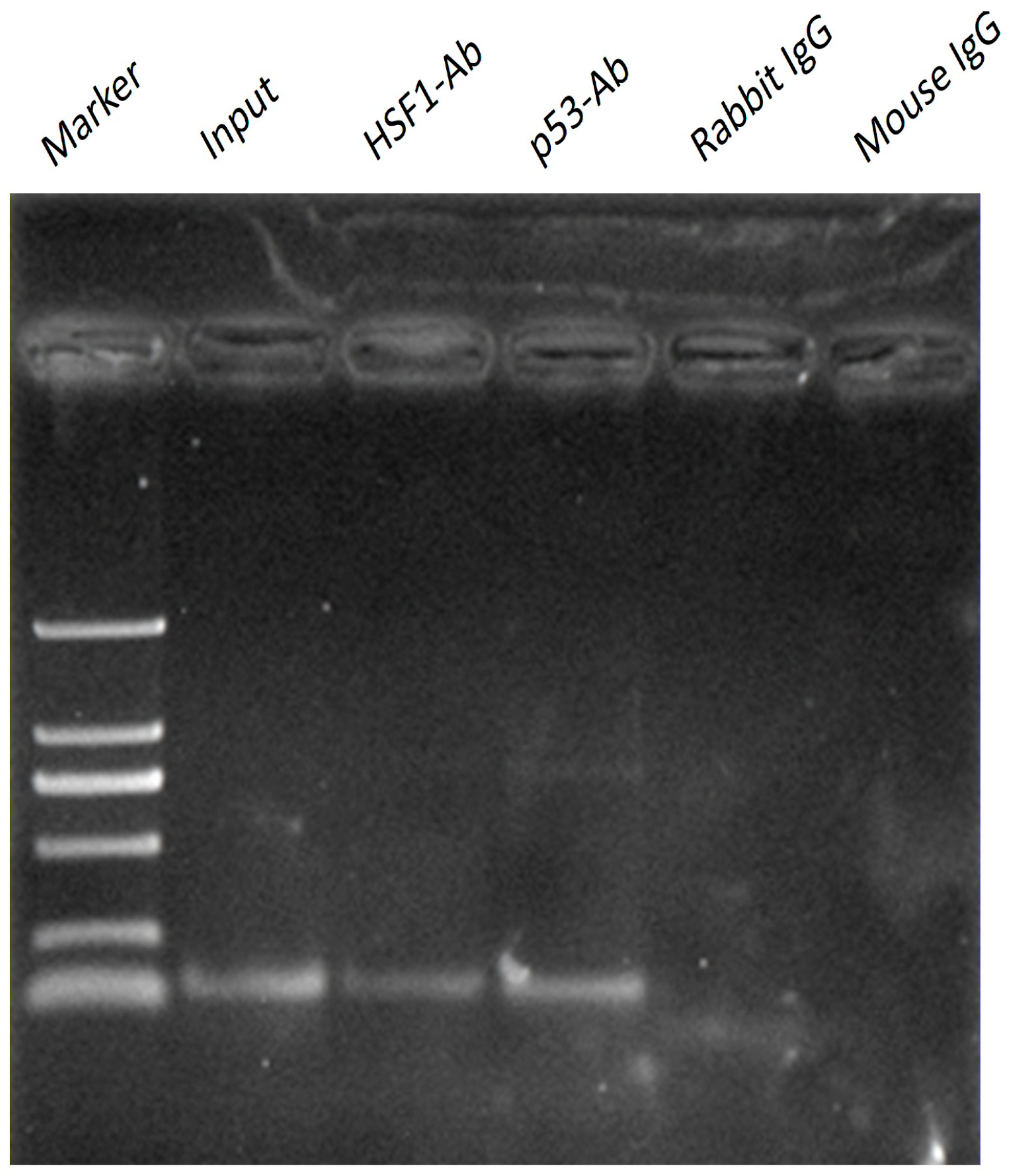
2.4. p53 and HSF1 Directly Binds to Omi/HtrA2 Promoter with ChIP Assay
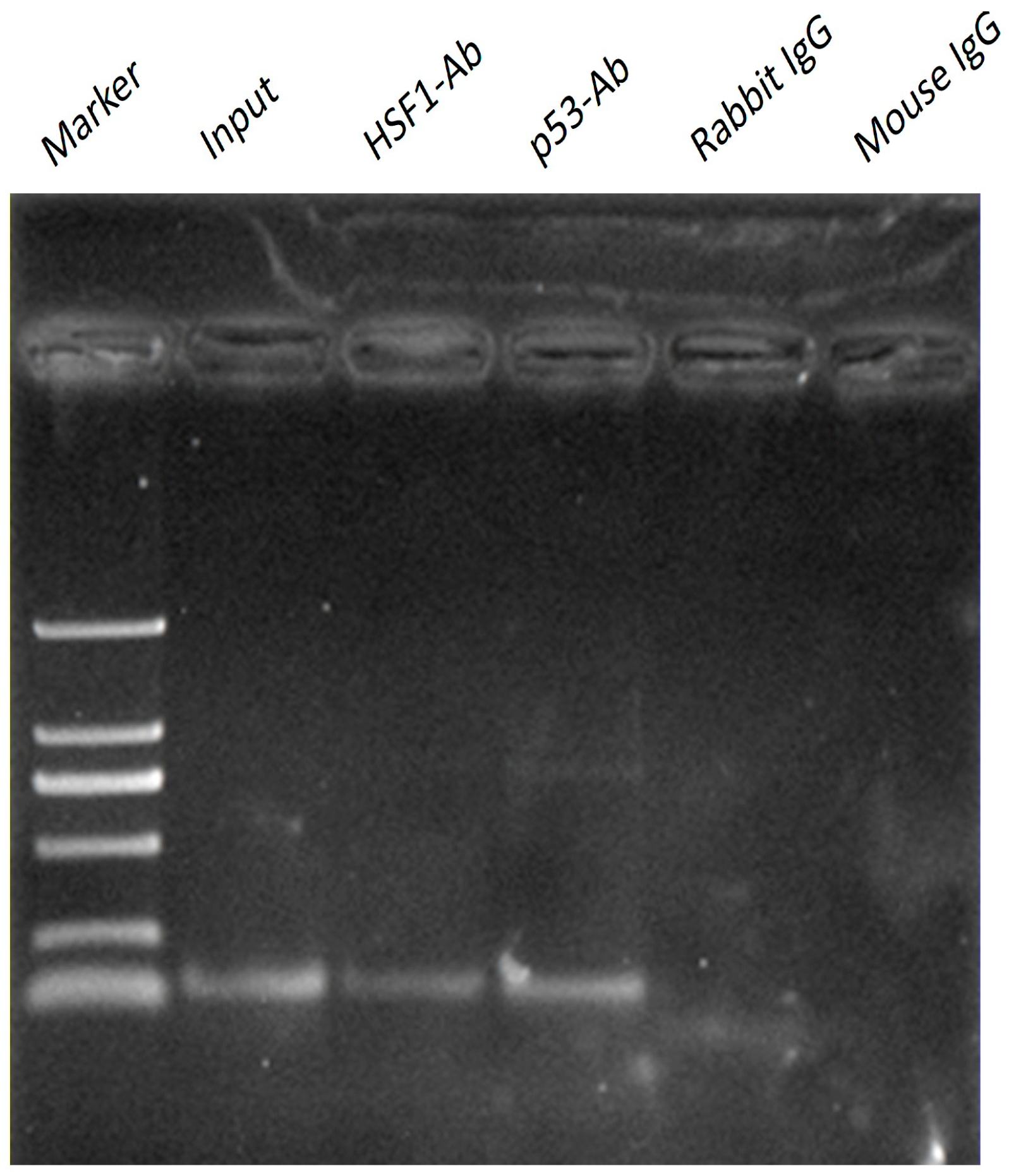
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Lines and Cell Culture
4.3. Isolation of Genomic DNA
4.4. Design and Preparation of Mouse Omi/HtrA2 Gene Promoter Constructs
4.5. Transfection and Luciferase Activity Assay
4.6. Bioinformatics Analysis
4.7. Western Blotting
4.8. RNA Preparation and Quantitative Reverse Transcription-Polymerase Chain Reaction (QRT-PCR) Analysis
4.9. ChIP Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bhattacharjee, A.; Richards, W.G.; Staunton, J.; Li, C.; Monti, S.; Vasa, P.; Ladd, C.; Beheshti, J.; Bueno, R.; Gillette, M.; et al. Classification of human lung carcinomas by mRNA expression profiling reveals distinct adenocarcinoma subclasses. Proc. Natl. Acad. Sci. USA 2001, 98, 13790–13795. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, Y.; Xu, G.; Peng, C.; Liu, E.; Li, Y.; Niu, J.; Li, C. Omi/HtrA2 pro-apoptotic marker differs in various hepatocellular carcinoma cell lines owing to ped/pea-15 expression level. Oncol. Rep. 2015, 33, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, X.; Peng, C.; Liu, E.; Li, Y.; Li, C.; Niu, J. The expression and clinical significance of Omi/HtrA2 in hepatocellular carcinoma. Hepatogastroenterology 2013, 60, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Ginos, M.A.; Page, G.P.; Michalowicz, B.S.; Patel, K.J.; Volker, S.E.; Pambuccian, S.E.; Ondrey, F.G.; Adams, G.L.; Gaffney, P.M. Identification of a gene expression signature associated with recurrent disease in squamous cell carcinoma of the head and neck. Cancer Res. 2004, 64, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wan, M.; Cao, X.; Ren, Y. Expression of AIF and Omi/HtrA2 in small lymphocytic lymphoma and diffuse large B-cell lymphoma. Arch. Pathol. Lab. Med. 2011, 135, 903–908. [Google Scholar] [PubMed]
- Zurawa-Janicka, D.; Kobiela, J.; Galczynska, N.; Stefaniak, T.; Lipinska, B.; Lachinski, A.; Skorko-Glonek, J.; Narkiewicz, J.; Proczko-Markuszewska, M.; Sledzinski, Z. Changes in expression of human serine protease HtrA1, HtrA2 and HtrA3 genes in benign and malignant thyroid tumors. Oncol. Rep. 2012, 28, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Mao, H.; Shen, L.; Zhao, Z.; Liu, R.; Liu, P. miR-519d represses ovarian cancer cell proliferation and enhances cisplatin-mediated cytotoxicity in vitro by targeting XIAP. Onco Targets Ther. 2014, 7, 587–597. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, X.; Huang, L.; Zhao, Y.; Tan, W. Downregulation of miR-130a contributes to cisplatin resistance in ovarian cancer cells by targeting X-linked inhibitor of apoptosis (XIAP) directly. Acta Biochim. Biophys. Sin. (Shanghai) 2013, 45, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Narkiewicz, J.; Klasa-Mazurkiewicz, D.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Changes in mRNA and protein levels of human HtrA1, HtrA2 and HtrA3 in ovarian cancer. Clin. Biochem. 2008, 41, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xing, H.; Gao, Q.; Chen, G.; Lu, Y.; Wang, S.; Ma, D. Regulation of Omi/HtrA2 by X-linked inhibitor of apoptosis protein in chemoresistance in human ovarian cancer cells. Gynecol. Oncol. 2005, 97, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Kim, E.M.; Lee, J.A.; Rhim, H.; Hwang, O. Matrix metalloproteinase-3 is activated by Omi/HtrA2 in dopaminergic cells: relevance to Parkinson’s disease. Neurochem. Int. 2012, 60, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.R.; Gao, E.; Hu, A.; Tao, L.; Qu, Y.; Most, P.; Koch, W.J.; Christopher, T.A.; Lopez, B.L.; Alnemri, E.S.; et al. Role of Omi/HtrA2 in apoptotic cell death after myocardial ischemia and reperfusion. Circulation 2005, 111, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, J.; Liu, J.; Tian, J.; Wu, Y.; Wang, X.; Quan, L.; Xu, H.; Wang, W.; Liu, H. Variations in the protein level of Omi/HtrA2 in the heart of aged rats may contribute to the increased susceptibility of cardiomyocytes to ischemia/reperfusion injury and cell death: Omi/HtrA2 and aged heart injury. Age (Dordr) 2013, 35, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Takahashi-Niki, K.; Akagi, T.; Hashikawa, T.; Takahashi, R. Mitochondrial protease Omi/HtrA2 enhances caspase activation through multiple pathways. Cell Death Differ. 2004, 11, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Fernandes-Alnemri, T.; Alnemri, E.S. A novel role for the mitochondrial HtrA2/Omi protease in aging. Autophagy 2013, 9, 420–421. [Google Scholar] [CrossRef] [PubMed]
- Goo, H.G.; Rhim, H.; Kang, S. HtrA2/Omi influences the stability of LON protease 1 and prohibitin, proteins involved in mitochondrial homeostasis. Exp. Cell Res. 2014, 328, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Goo, H.G.; Jung, M.K.; Han, S.S.; Rhim, H.; Kang, S. HtrA2/Omi deficiency causes damage and mutation of mitochondrial DNA. Biochim. Biophys. Acta 2013, 1833, 1866–1875. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.M.; Datta, P.; Srinivasula, S.M.; Ji, W.; Gupta, S.; Zhang, Z.; Davies, E.; Hajnoczky, G.; Saunders, T.L.; van Keuren, M.L.; et al. Loss of Omi mitochondrial protease activity causes the neuromuscular disorder of mnd2 mutant mice. Nature 2003, 425, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Cilenti, L.; Ambivero, C.T.; Ward, N.; Alnemri, E.S.; Germain, D.; Zervos, A.S. Inactivation of Omi/HtrA2 protease leads to the deregulation of mitochondrial Mulan E3 ubiquitin ligase and increased mitophagy. Biochim. Biophys. Acta 2014, 1843, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, J.; Lu, X.; Shen, C.; Zeng, J.; Chen, L.; Pei, Z. Melatonin protects against rotenone-induced cell injury via inhibition of Omi and Bax-mediated autophagy in Hela cells. J. Pineal Res. 2012, 52, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Poulose, N.; Raju, R. Aging and injury: Alterations in cellular energetics and organ function. Aging Dis. 2014, 5, 101–108. [Google Scholar] [PubMed]
- Hensen, S.M.; Heldens, L.; van Enckevort, C.M.; van Genesen, S.T.; Pruijn, G.J.; Lubsen, N.H. Heat shock factor 1 is inactivated by amino acid deprivation. Cell Stress Chaperones 2012, 17, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Kern, A.; Ackermann, B.; Clement, A.M.; Duerk, H.; Behl, C. HSF1-controlled and age-associated chaperone capacity in neurons and muscle cells of C. elegans. PLoS ONE 2010, 5, e8568. [Google Scholar] [CrossRef] [PubMed]
- Taimor, G.; Rakow, A.; Piper, H.M. Transcription activator protein 1 (AP-1) mediates NO-induced apoptosis of adult cardiomyocytes. FASEB J. 2001, 15, 2518–2520. [Google Scholar] [CrossRef] [PubMed]
- Schlieper, A.; Anwar, M.; Heger, J.; Piper, H.M.; Euler, G. Repression of anti-apoptotic genes via AP-1 as a mechanism of apoptosis induction in ventricular cardiomyocytes. Pflugers Arch. 2007, 454, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Gaur, P.; Prasad, S. Alterations in the Sp1 binding and Fmr-1 gene expression in the cortex of the brain during maturation and aging of mouse. Mol. Biol. Rep. 2014, 41, 6855–6863. [Google Scholar] [CrossRef] [PubMed]
- Sawai, M.; Ishikawa, Y.; Ota, A.; Sakurai, H. The proto-oncogene JUN is a target of the heat shock transcription factor HSF1. FEBS J. 2013, 280, 6672–6680. [Google Scholar] [CrossRef] [PubMed]
- Tun, C.; Guo, W.; Nguyen, H.; Yun, B.; Libby, R.T.; Morrison, R.S.; Garden, G.A. Activation of the extrinsic caspase pathway in cultured cortical neurons requires p53-mediated down-regulation of the X-linked inhibitor of apoptosis protein to induce apoptosis. J. Neurochem. 2007, 102, 1206–1219. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, S.; Hou, Y.Y.; Guo, A.K.; Hirata, H.; Nakajima, W.; Yip, A.K.; Yu, C.H.; Harada, I.; Chiam, K.H.; Sawada, Y.; et al. p53-Mediated activation of the mitochondrial protease Omi/HtrA2 prevents cell invasion. J. Cell Biol. 2014, 204, 1191–1207. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feng, Y.; Xu, L.; Chen, Y.; Zhang, Y.; Su, D.; Ren, G.; Lu, J.; Huang, B. YY1 restrained cell senescence through repressing the transcription of p16. Biochim. Biophys. Acta 2008, 1783, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Liu, X.; Wu, Y.; Wang, W.; Ma, X.; Liu, H. Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter. Int. J. Mol. Sci. 2016, 17, 119. https://doi.org/10.3390/ijms17010119
Liu D, Liu X, Wu Y, Wang W, Ma X, Liu H. Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter. International Journal of Molecular Sciences. 2016; 17(1):119. https://doi.org/10.3390/ijms17010119
Chicago/Turabian StyleLiu, Dan, Xin Liu, Ye Wu, Wen Wang, Xinliang Ma, and Huirong Liu. 2016. "Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter" International Journal of Molecular Sciences 17, no. 1: 119. https://doi.org/10.3390/ijms17010119
APA StyleLiu, D., Liu, X., Wu, Y., Wang, W., Ma, X., & Liu, H. (2016). Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter. International Journal of Molecular Sciences, 17(1), 119. https://doi.org/10.3390/ijms17010119