
Novel Radiolytic Rotenone Derivative, Rotenoisin B with Potent Anti-Carcinogenic Activity in Hepatic Cancer Cells

 and
and
Abstract
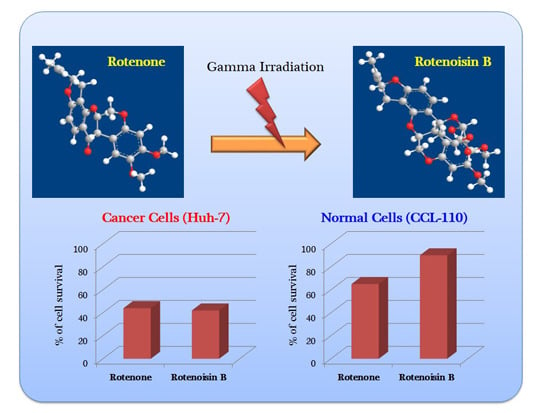
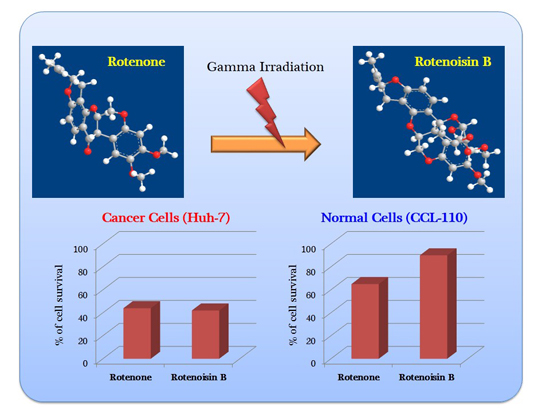
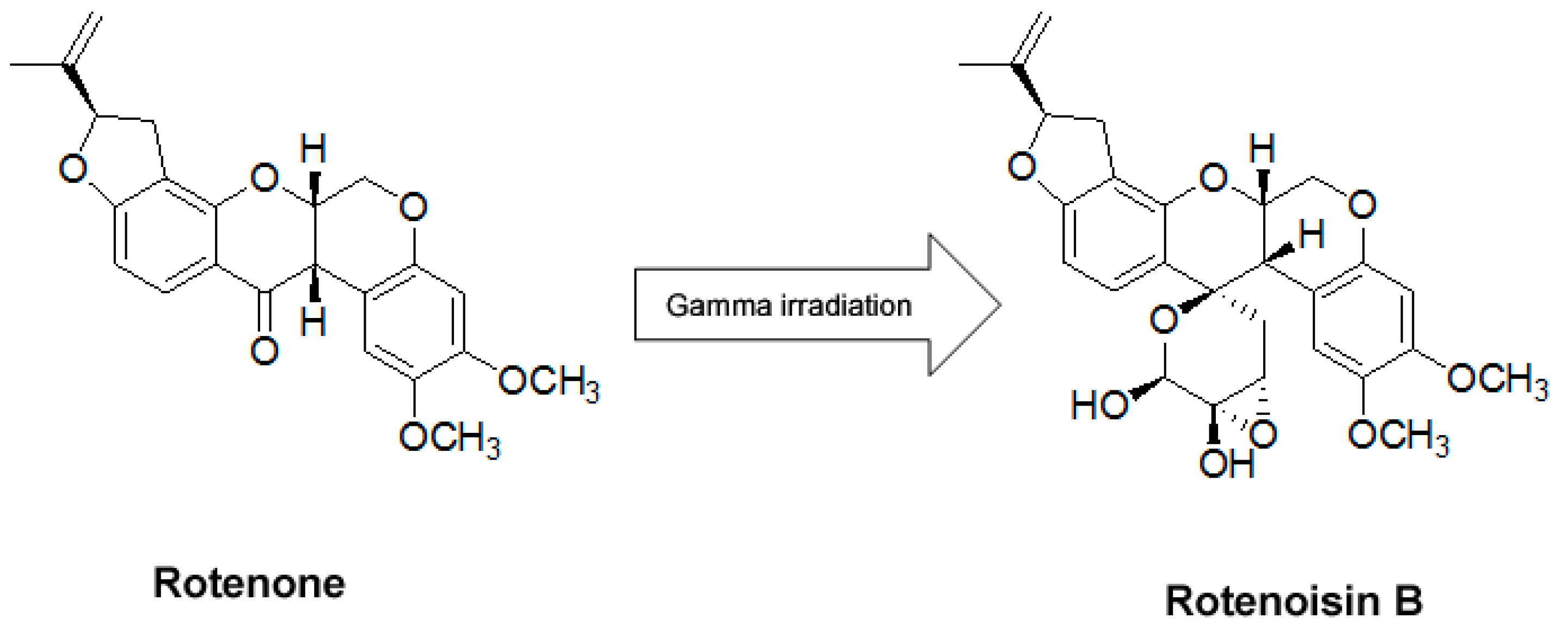
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Cytotoxicity Induced by Rotenoisin B Compared with Parent Rotenone

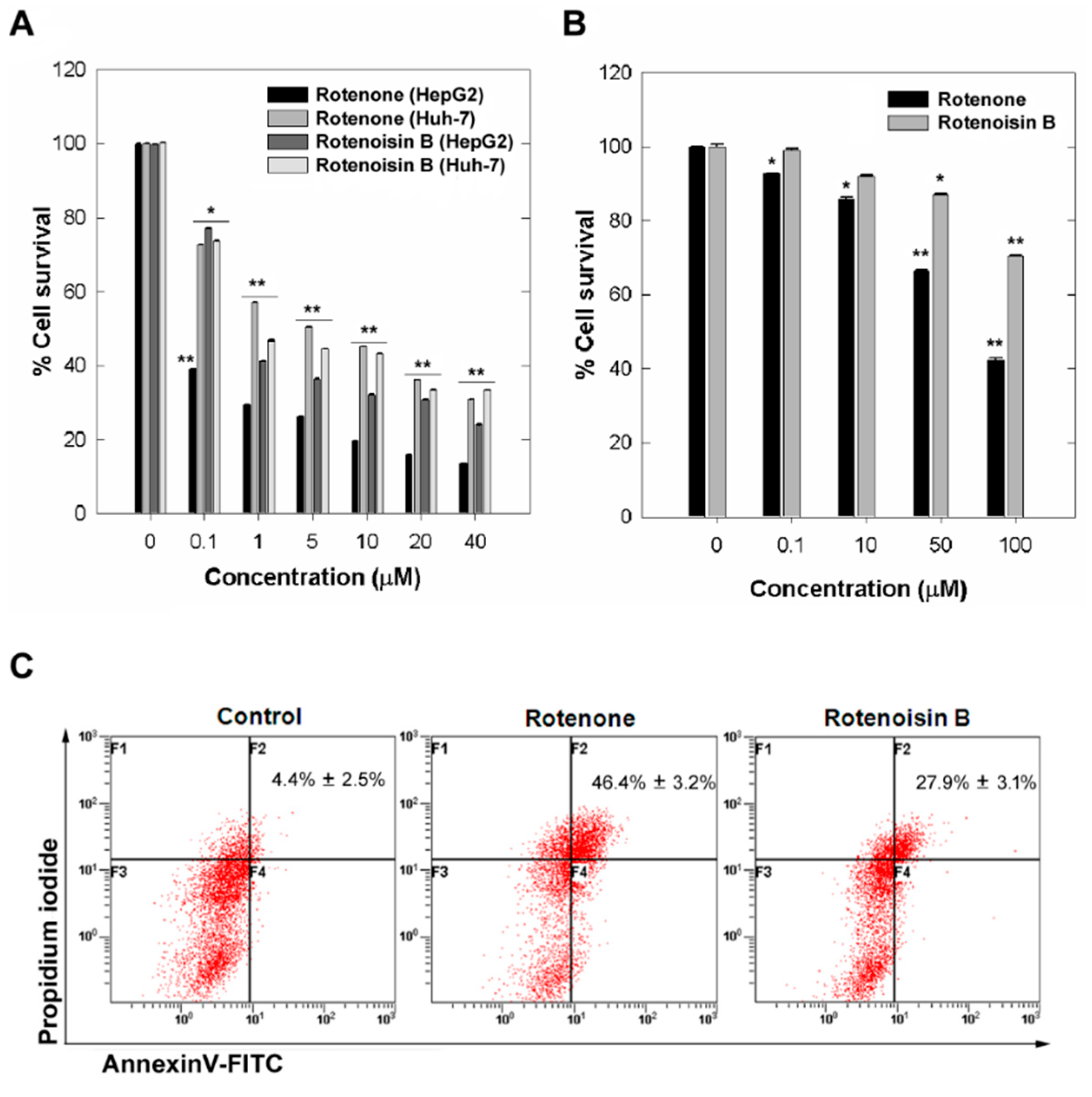
2.2. Effect of Rotenoisin B on Ψm, ROS and Signaling Pathways
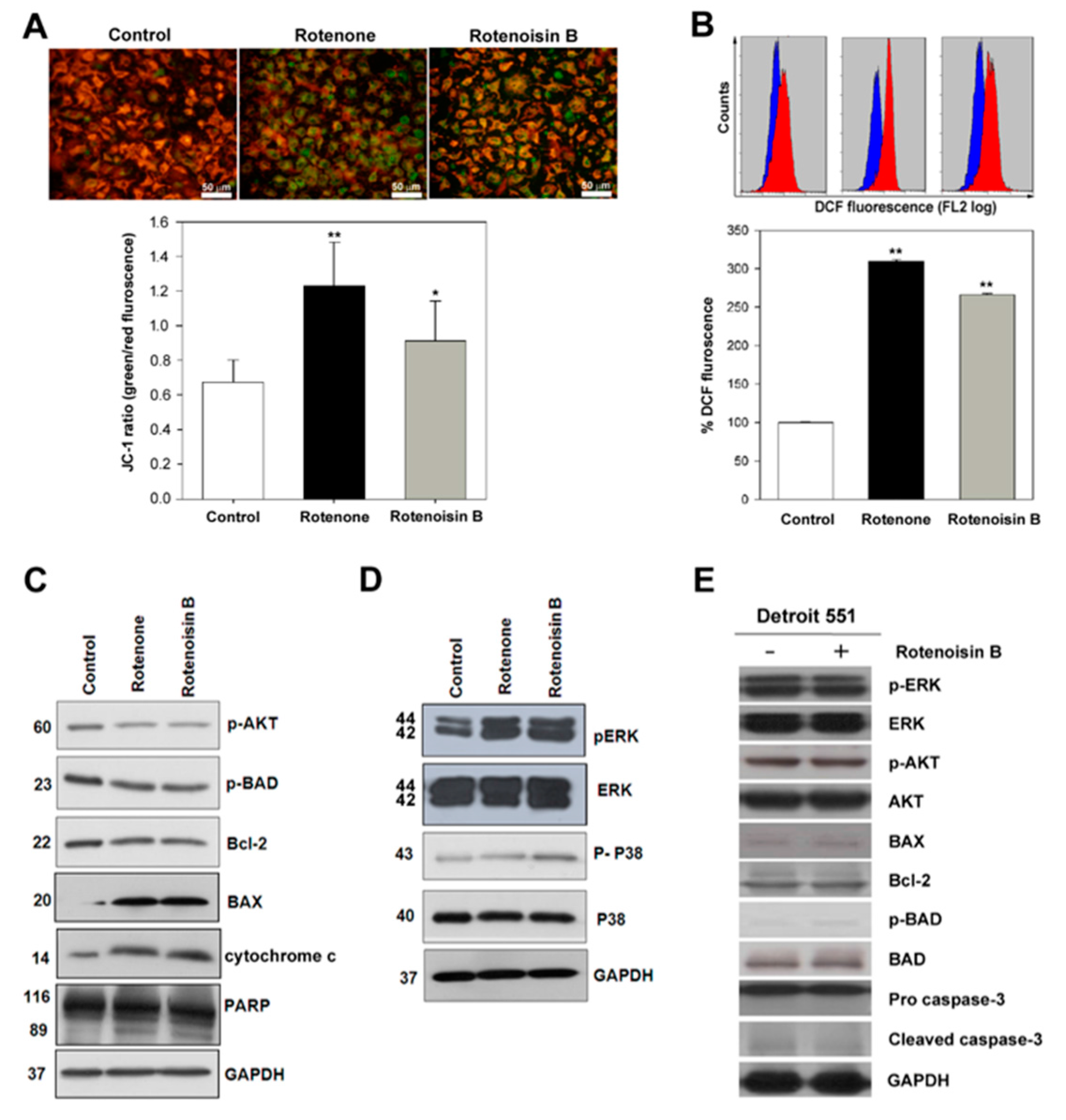
2.3. Rotenoisin B Effect on G2/M Cell Cycle Arrest
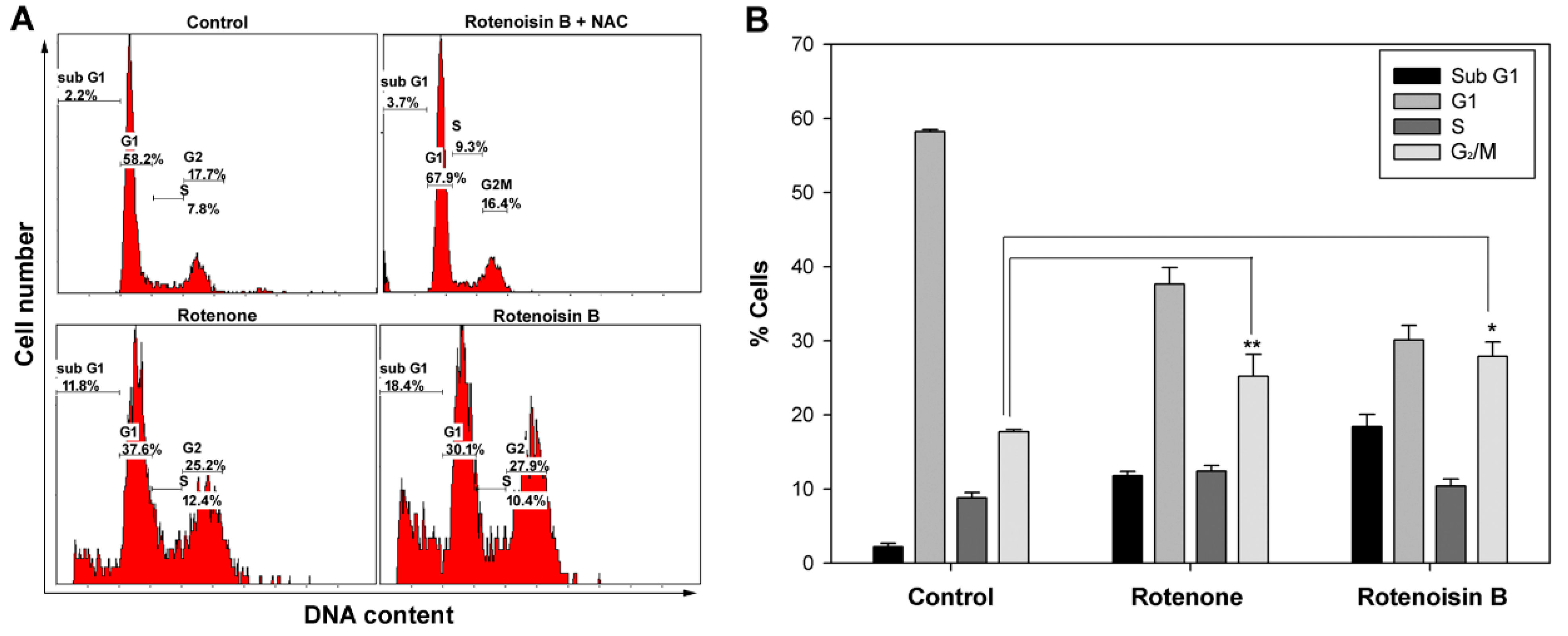
3. Materials and Methods
3.1. Reagent and Chemicals
3.2. Sample Preparation and Various Treatments of Stresses
3.3. Cell Culture and Viability Assay
3.4. Evaluation of Apoptosis by Annexin-V FITC/Propidium Iodide
3.5. Mitochondrial Membrane Potential (Ψm) Detection by Spectroscopy
3.6. Intracellular ROS Detection by H2DCFDA
3.7. Immuno Blot Experiment
3.8. Cell Cycle Analysis by Flow Cytometry
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- EI-Serag, H.B.; Rudolph, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Yamanoi, A.; Nazmy El Assal, O.; Kohno, H.; Nagasue, N. Adjuvant chemotherapy after resection of hepatocellular carcinoma causes deterioration of long-term prognosis in cirrhotic patients: Metaanalysis of three randomized controlled trials. Cancer 2001, 91, 2378–2385. [Google Scholar] [CrossRef]
- Chau, G.Y.; Lui, W.Y.; Tsay, S.H.; Chao, Y.; King, K.L.; Wu, C.W. Postresectional adjuvant intraportal chemotherapy in patients with hepatocellular carcinoma: A case-control study. Ann. Surg. Oncol. 2006, 13, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.E. Pesticides derived from plants and other organisms. In Handbook of Pesticide Toxicology; Hayes, W.J., Laws, E.R., Eds.; Academic Press: New York, NY, USA, 1991; Volume 2. [Google Scholar]
- Lee, J.; Huang, M.S.; Yang, I.C.; Lai, T.C.; Wang, J.L.; Pang, V.F.; Hsiao, M.; Kuo, M.Y. Essential roles of caspases and their upstream regulators in rotenone-induced apoptosis. Biochem. Biophys. Res. Commun. 2008, 371, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Abdel, A.E. The neuroprotective effects of purslane (Portulaca oleracea) rotenone-induced biochemical changes and apoptosis in brain of rats. CNS Neurol. Disord. 2013, 12, 830–841. [Google Scholar] [CrossRef]
- Greenamyre, J.T.; Betarbet, R.; Sherer, T.B. The rotenone model of Parkinson’s disease: Genes, environment and mitochondria. Parkinsonism Relat. Disord. 2003, 9, S59–S64. [Google Scholar] [CrossRef]
- Murillo, G.; Salti, G.I.; Kosmeder, J.W.; Pezzuto, J.M.; Mehta, R.G. Deguelin inhibits the growth of colon cancer cells through the induction of apoptosis and cell cycle arrest. Eur. J. Cancer 2002, 38, 2446–2454. [Google Scholar] [CrossRef]
- Lee, H.Y.; Oh, S.H.; Woo, J.K.; Kim, W.Y.; van Pelt, C.S.; Price, R.E.; Cody, D.; Tran, H.; Pezzuto, J.M.; Moriarty, R.M.; et al. Chemopreventive effects of deguelin, a novel Akt inhibitor, on tobacco induced lung tumorigenesis. J. Nat. Cancer Inst. 2005, 16, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.T.; Huang, H.C.; Lin, J.K. Rotenone Induces Apoptosis in MCF-7 human breast cancer cell-mediated ROS through JNK and p38 signaling. Mol. Carcinog 2010, 49, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Klaunig, J.E. Role of the mitochondrial membrane permeability transition (MPT) in rotenone-induced apoptosis in liver cells. Toxicol. Sci. 2000, 53, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Kim, J.K.; Ito, H.; Jo, C. Enhancement of pancreatic lipase inhibitory activity of curcumin by radiolytic transformation. Bioorg. Med. Chem. Lett. 2011, 21, 1512–1514. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Chung, B.Y.; Lee, S.S.; Bai, H.W.; Cho, J.Y.; Jo, C.; Kim, T.H. Radiolytic transformation of rotenone with potential anti-adipogenic activity. Bioorg. Med. Chem. Lett. 2013, 23, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [PubMed]
- Radad, K.; Rausch, W.D.; Gille, G. Rotenone induces cell death in primary dopaminergic culture by increasing ROS production and inhibiting mitochondrial respiration. Neurochem. Int. 2006, 49, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Yang, U.-J.; Ko, S.; Shim, S.-M. Vitamin C from standardized water spinach extract on inhibition of cytotoxicity and oxidative stress induced by heavy metals in HepG2 cells. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 161–166. [Google Scholar]
- Kim, B.I.; Joo, Y.H.; Pak, P.J.; Kim, J.-S.; Chung, N. Different shapes of Al2O3 particles induce differential cytotoxicity via a mechanism involving lysosomal destabilization and reactive oxygen species generation. Korean Soc. Appl. Biol. Chem. 2015, 58, 433–442. [Google Scholar] [CrossRef]
- Li, N.; Ragheb, K.; Lawler, G.; Sturgis, J.; Rajwa, B.; Melendez, J.A.; Robinson, J.P. Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production. J. Biol. Chem. 2003, 278, 8516–8525. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.S.; Hornung, B.; Lecane, P.; Jones, D.P.; Knox, S.J. Rotenone-induced G2/M cell cycle arrest and apoptosis in a human B lymphoma cell line PW. Biochem. Biophys. Res. Commun. 2001, 289, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, X.; Chen, R.; Zhang, H. Radio sensitization by inhibiting complex I activity in human hepatoma HepG2 cells to X-ray radiation. J. Radiat. Res. 2012, 53, 257–263. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badaboina, S.; Bai, H.-W.; Na, Y.H.; Park, C.-H.; Kim, T.H.; Lee, T.-H.; Chung, B.Y. Novel Radiolytic Rotenone Derivative, Rotenoisin B with Potent Anti-Carcinogenic Activity in Hepatic Cancer Cells. Int. J. Mol. Sci. 2015, 16, 16806-16815. https://doi.org/10.3390/ijms160816806
Badaboina S, Bai H-W, Na YH, Park C-H, Kim TH, Lee T-H, Chung BY. Novel Radiolytic Rotenone Derivative, Rotenoisin B with Potent Anti-Carcinogenic Activity in Hepatic Cancer Cells. International Journal of Molecular Sciences. 2015; 16(8):16806-16815. https://doi.org/10.3390/ijms160816806
Chicago/Turabian StyleBadaboina, Srilatha, Hyoung-Woo Bai, Yun Hee Na, Chul-Hong Park, Tae Hoon Kim, Tae-Hoon Lee, and Byung Yeoup Chung. 2015. "Novel Radiolytic Rotenone Derivative, Rotenoisin B with Potent Anti-Carcinogenic Activity in Hepatic Cancer Cells" International Journal of Molecular Sciences 16, no. 8: 16806-16815. https://doi.org/10.3390/ijms160816806
APA StyleBadaboina, S., Bai, H.-W., Na, Y. H., Park, C.-H., Kim, T. H., Lee, T.-H., & Chung, B. Y. (2015). Novel Radiolytic Rotenone Derivative, Rotenoisin B with Potent Anti-Carcinogenic Activity in Hepatic Cancer Cells. International Journal of Molecular Sciences, 16(8), 16806-16815. https://doi.org/10.3390/ijms160816806