
Tudor-SN Regulates Milk Synthesis and Proliferation of Bovine Mammary Epithelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
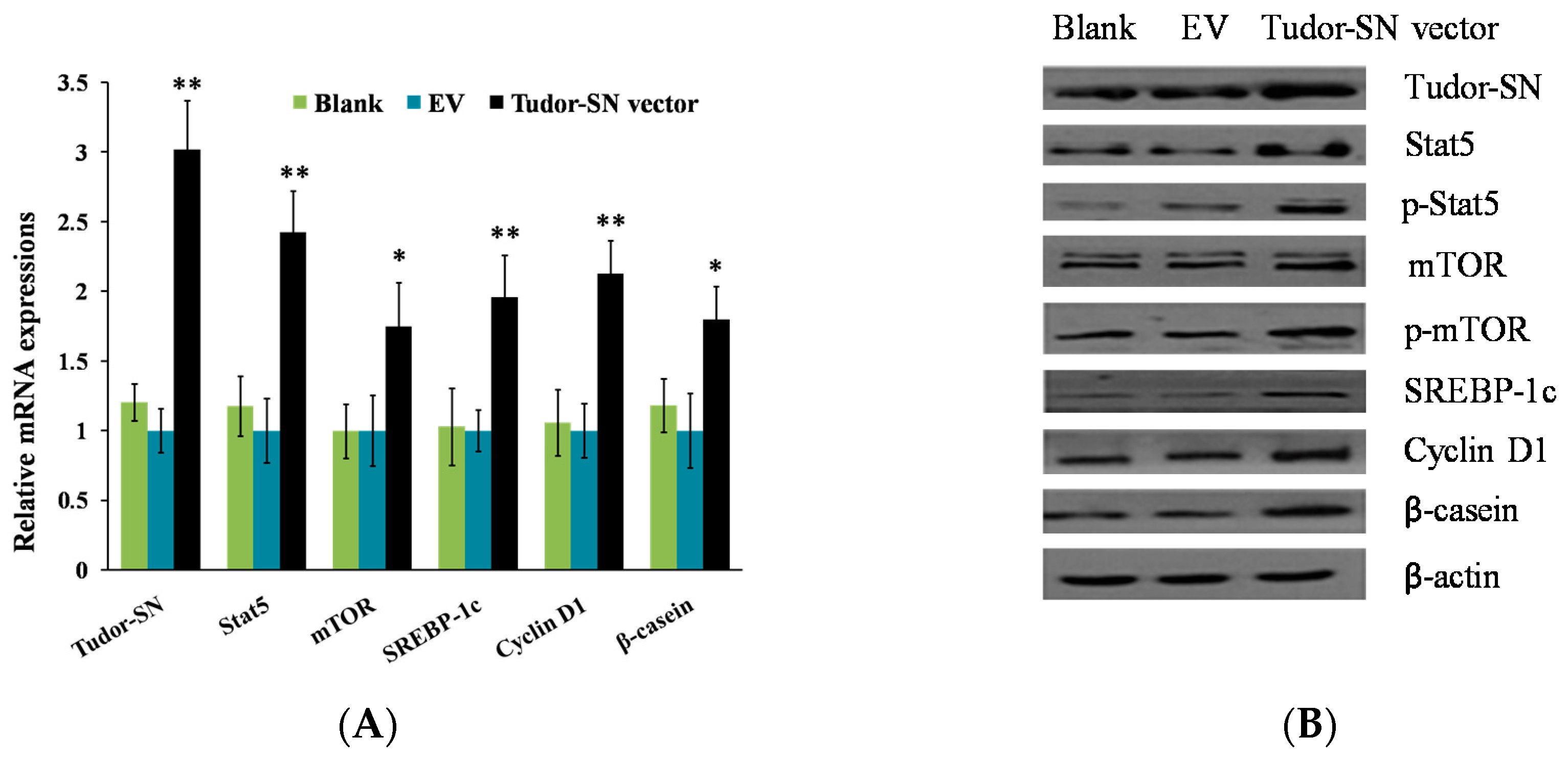
2.1. Overexpression of Tudor-SN Enhances Cell Signaling Pathways Leading to Milk Protein and Fat Synthesis and Proliferation of BMEC
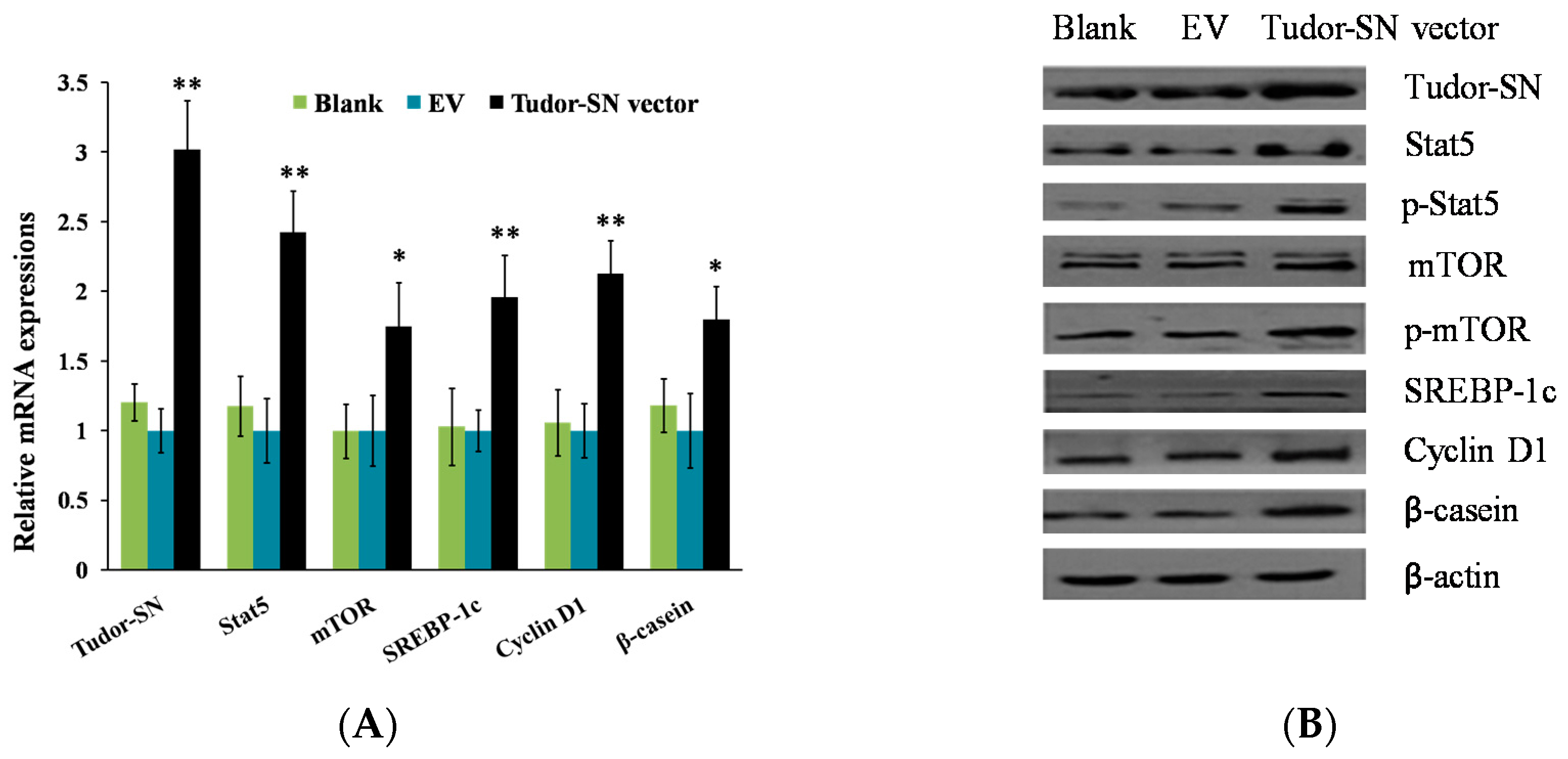
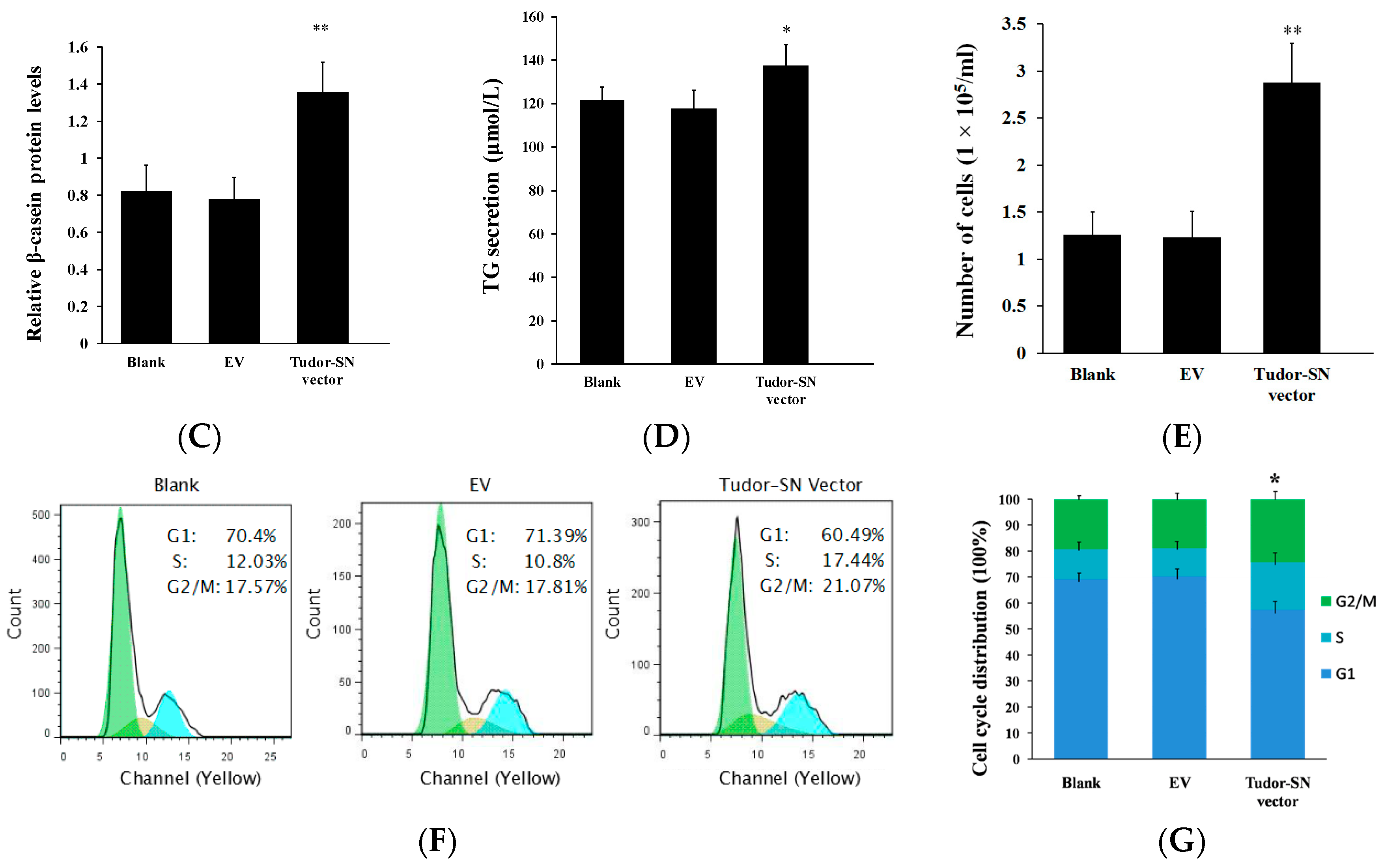
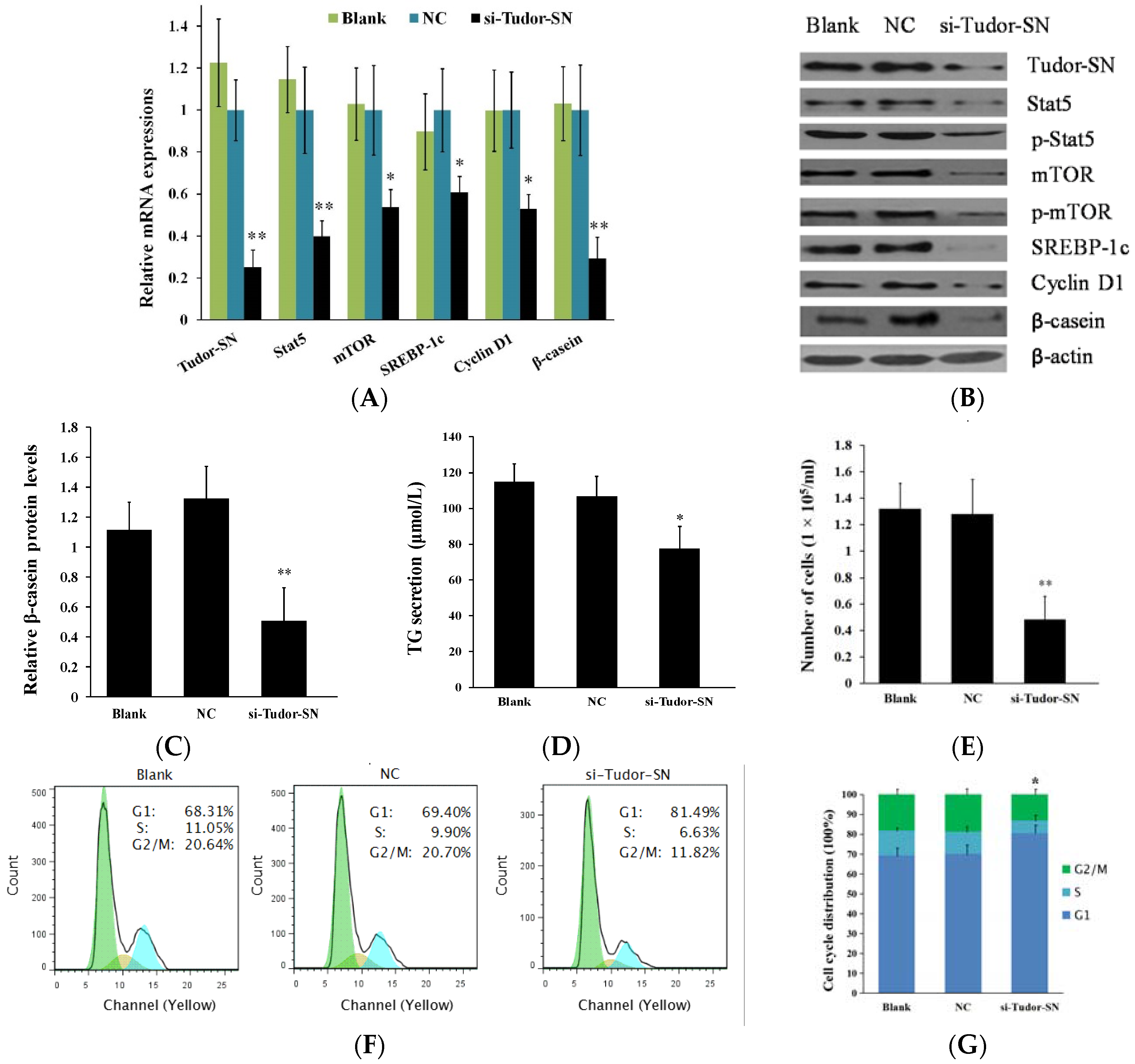
2.2. Gene Silencing of Tudor Staphylococcal Nuclease (Tudor-SN) Represses Cell Signaling Pathways Leading to Milk Protein and Fat Synthesis and Proliferation of Bovine Mammary Epithelial Cells (BMEC)
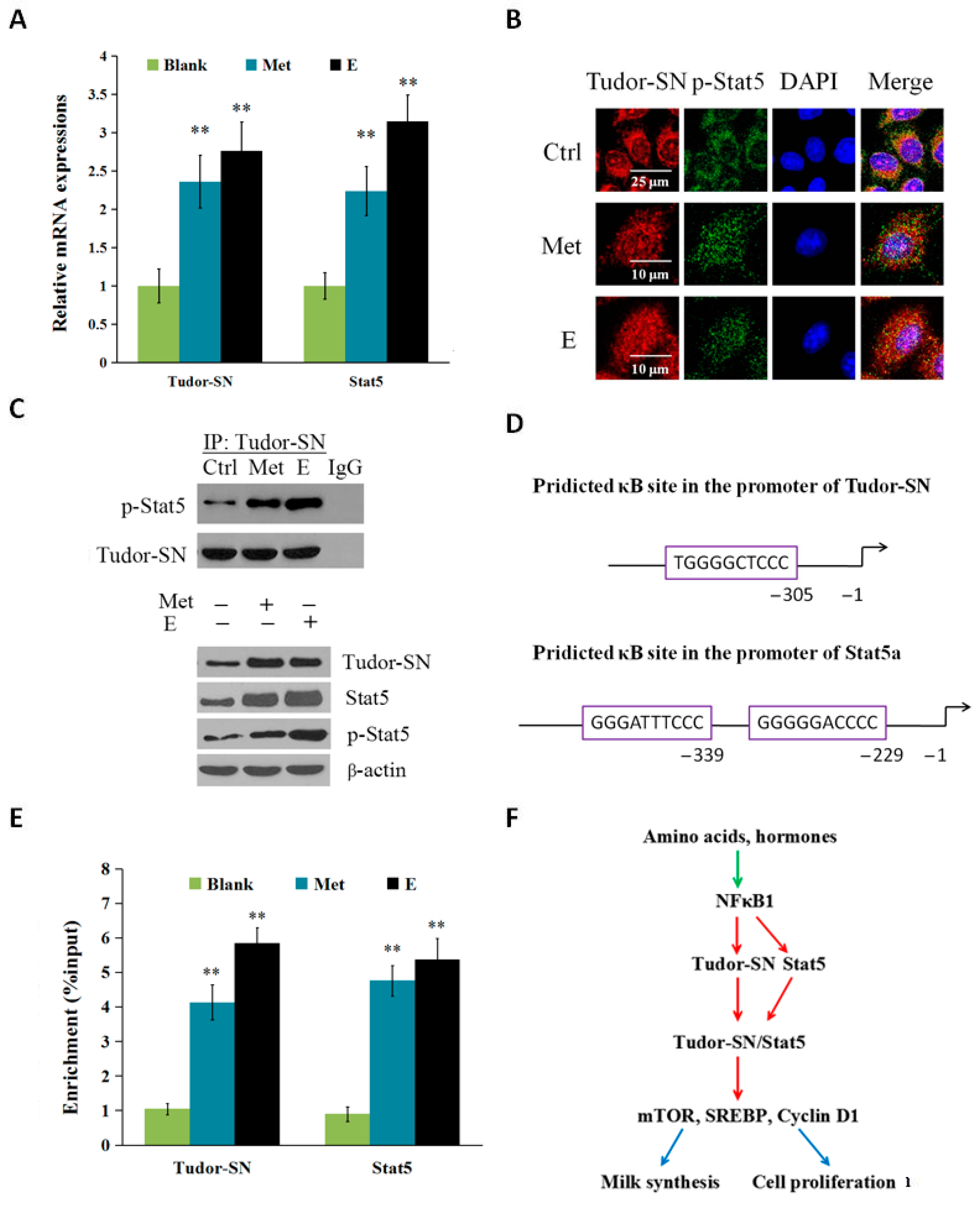
2.3. Met and Estrogen Upregulates the Expression of Tudor-SN and Interaction with p-Stat5 in the Nucleus
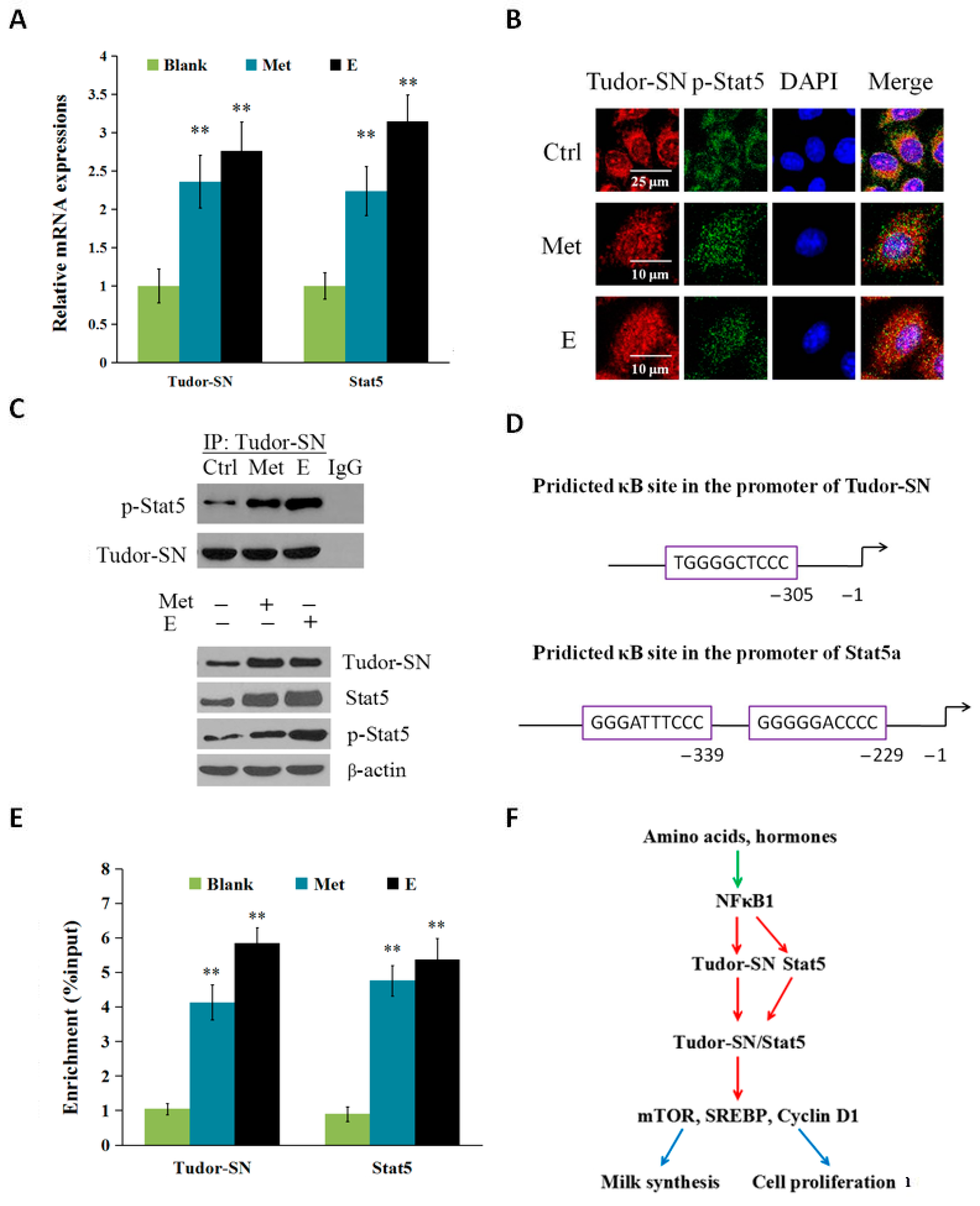
2.4. Discussion
3. Materials and Methods
3.1. Cell Preparation and Treatments
3.2. Cell Number Assay
3.3. Analysis of Cell Cycle Progression
3.4. Detection of Triglyceride Secretion
3.5. RNA Extraction and Real Time PCR
3.6. Tudor-SN Overexpression
3.7. siRNA and Transfection
3.8. Western Blotting Analysis
3.9. Immunofluorescence
3.10. Co-IP Assay
3.11. ChIP-PCR and ChIP-qPCR
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Callebaut, I.; Mornon, J.P. The human EBNA-2 coactivator p100: Multidomain organization and relationship to the staphylococcal nuclease fold and to the Tudor protein involved in Drosophila melanogaster development. Biochem. J. 1997, 321, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P. P100, a transcriptional coactivator, is a human homologue of staphylococcal nuclease. Protein Sci. 1997, 6, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Drapkin, R.; Yalamanchili, R.; Mosialos, G.; Kieff, E. The Epstein-Barr virus nuclear protein 2 acidic domain forms a complex with a novel cellular coactivator that can interact with TFIIE. Mol. Cell. Biol. 1995, 15, 4735–4744. [Google Scholar] [CrossRef] [PubMed]
- Dash, A.B.; Orrico, F.C.; Ness, S.A. The EVES motif mediates both intermolecular and intramolecular regulation of c-Myb. Genes Dev. 1996, 10, 1858–1869. [Google Scholar] [CrossRef] [PubMed]
- Leverson, J.D.; Koskinen, P.J.; Orrico, F.C.; Rainio, E.M.; Jalkanen, K.J.; Dash, A.B.; Eisenman, R.N.; Ness, S.A. Pim-1 kinase and p100 cooperate to enhance c-Myb activity. Mol. Cell 1998, 2, 417–425. [Google Scholar] [CrossRef]
- Paukku, K.; Yang, J.; Silvennoinen, O. Tudor and nuclease-like domains containing protein p100 function as coactivators for signal transducer and activator of transcription 5. Mol. Endocrinol. 2003, 17, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Välineva, T.; Yang, J.; Palovuori, R.; Silvennoinen, O. The transcriptional co-activator protein p100 recruits histone acetyltransferase activity to Stat6 and mediates interaction between the CREB-binding protein and Stat6. J. Biol. Chem. 2005, 280, 14989–14996. [Google Scholar] [CrossRef] [PubMed]
- Saarikettu, J.; Ovod, V.; Vuoksio, M.; Grönholm, J.; Yang, J.; Silvennoinen, O. Monoclonal antibodies against human Tudor-SN. Hybridoma 2010, 29, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, M.K.; Lee, R.S.; Hawkins, S.; Wheeler, T.T. The p100 EBNA-2 coactivator: A highly conserved protein found in a range of exocrine and endocrine cells and tissues in cattle. Biochim. Biophys. Acta 2005, 1681, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.W.; Winter, S.; Rackwitz, H.R.; Heid, H.W. Nuclear coactivator protein p100 is present in endoplasmic reticulum and lipid droplets of milk secreting cells. Biochim. Biophys. Acta 2000, 1523, 84–90. [Google Scholar] [CrossRef]
- Fresnedo, O.; de Heredia, M.L.; Martínez, M.J.; Cristóbal, S.; Rejas, M.T.; Cuezva, J.M.; Ochoa, B. Immunolocalization of a novel cholesteryl ester hydrolase in the endoplasmic reticulum of murine and human hepatocytes. Hepatology 2001, 33, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arcos, I.; Rueda, Y.; González-Kother, P.; Palacios, L.; Ochoa, B.; Fresnedo, O. Association of SND1 protein to low density lipid droplets in liver steatosis. J. Physiol. Biochem. 2010, 66, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, N.; Rajasekaran, D.; Srivastava, J.; Gredler, R.; Akiel, M.A.; Robertson, C.L.; Emdad, L.; Fisher, P.B.; Sarkar, D. Role of the staphylococcal nuclease and Tudor domain containing 1 in oncogenesis (review). Int. J. Oncol. 2015, 46, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Armengol, S.; Arretxe, E.; Rodríguez, L.; Ochoa, B.; Chico, Y.; Martínez, M.J. NF-κB, Sp1 and NF-Y as transcriptional regulators of human SND1 gene. Biochimie 2013, 95, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Song, H.; Guo, B.; Zhang, Y.; Zheng, Y.; Lin, C.; Wu, Y.; Guan, G.; Sha, R.; Zhou, Q.; et al. MiR-361-5p inhibits colorectal and gastric cancer growth and metastasis by targeting staphylococcal nuclease domain containing-1. Oncotarget 2015, 6, 17404–17416. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Zhao, F.Q. Current major advances in the regulation of milk protein gene expression. Crit. Rev. Eukaryot. Gene Expr. 2014, 24, 357–378. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.J.; Burdon, T.G. Prolactin signal transduction mechanisms in the mammary gland: The role of the Jak/Stat pathway. Rev. Reprod. 1996, 1, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Imada, K.; Leonard, W.J. The Jak-Stat pathway. Mol. Immunol. 2000, 37, 1–11. [Google Scholar] [CrossRef]
- Swiatek-Machado, K.; Kaminska, B. Stat signaling in glioma cells. Adv. Exp. Med. Biol. 2013, 986, 189–208. [Google Scholar] [PubMed]
- Morales, J.K.; Falanga, Y.T.; Depcrynski, A.; Fernando, J.; Ryan, J.J. Mast cell homeostasis and the Jak-Stat pathway. Genes Immun. 2010, 11, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.J. The growth hormone receptor. Growth Horm. IGF Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, F.; Si, Y.; Huang, Y.L.; Yu, C.P.; Luo, C.C.; Zhang, N.; Li, Q.Z.; Gao, X.J. Negative feedback-loop regulation of GSK3β phosphorylation and mTOR/S6K1 signaling pathway independent of AKT to regulate milk synthesis and cell proliferation of dairy cow mammary epithelial cells. Molecules 2014, 19, 9435–9452. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Luo, C.; Qu, B.; Khudhair, N.; Gu, X.; Zang, Y.; Wang, C.; Zhang, N.; Li, Q.; Gao, X. Molecular network including eIF1AX, RPS7, and 14-3-3γ regulates protein translation and cell proliferation in bovine mammary epithelial cells. Arch. Biochem. Biophys. 2014, 564, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Lollivier, V.; Dessauge, F.; Bruckmaier, R.M.; Ollier, S.; Boutinaud, M. New developments on the galactopoietic role of prolactin in dairy ruminants. Domest. Anim. Endocrinol. 2012, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Brisken, C.; O’Malley, B. Hormone action in the mammary gland. Cold Spring Harb. Perspect. Biol. 2010, 2, a003178. [Google Scholar] [CrossRef] [PubMed]
- Sting, J. Estrogen and progesterone in normal mammary gland development and in cancer. Horm. Cancer 2011, 2, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Arretxe, E.; Armengol, S.; Mula, S.; Chico, Y.; Ochoa, B.; Martínez, M.J. Profiling of promoter occupancy by the SND1 transcriptional coactivator identifies downstream glycerolipid metabolic genes involved in TNFα response in human hepatoma cells. Nucleic Acids Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.M.; Gao, X.J.; Li, Q.Z.; Huang, J.G.; Liu, R.; Li, H.M. Comparative phosphoproteomics analysis of the effects of l-methionine on dairy cow mammary epithelial cells. Can. J. Anim. Sci. 2012, 92, 433–442. [Google Scholar] [CrossRef]
- Huang, Y.L.; Zhao, F.; Luo, C.C.; Zhang, X.; Si, Y.; Sun, Z.; Zhang, L.; Li, Q.Z.; Gao, X.J. SOCS3-mediated blockade reveals major contribution of Jak2/Stat5 signaling pathway to lactation and proliferation of dairy cow mammary epithelial cells in vitro. Molecules 2013, 18, 12987–13002. [Google Scholar] [CrossRef] [PubMed]
- Peterson, T.R.; Sengupta, S.S.; Harris, T.E.; Carmack, A.E.; Kang, S.A.; Balderas, E.; Guertin, D.A.; Madden, K.L.; Carpenter, A.E.; Finck, B.N.; et al. mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 2011, 146, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Bakan, I.; Laplante, M. Connecting mTORC1 signaling to SREBP-1 activation. Curr. Opin. Lipidol. 2012, 23, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Corl, B.A. Transcriptional regulation of lipid synthesis in bovine mammary epithelial cells by sterol regulatory element binding protein-1. J. Dairy Sci. 2012, 95, 3743–3755. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhao, F.; Wei, C.; Liang, M.; Zhang, N.; Wang, C.; Li, Q.Z.; Gao, X.J. Function of SREBP1 in the milk fat synthesis of dairy cow mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 16998–17013. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinillos, A.; Ferrari, A.C. mTOR signaling pathway and mTOR inhibitors in cancer therapy. Hematol. Oncol. Clin. N. Am. 2012, 26, 483–505. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, P.J. mTOR in renal cell cancer: Modulator of tumor biology and therapeutic target. Expert Rev. Mol. Diagn. 2009, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.S.; Lee, A.; Li, J.; Liyanage, H.; Yang, Y.; Guo, L.; Asmann, Y.W.; Li, P.W.; Erickson-Johnson, M.; Sakai, Y.; et al. Common oncogene mutations and novel SND1-BRAF transcript fusion in lung adenocarcinoma from never smokers. Sci. Rep. 2015, 5, 9755. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhang, C.; Tecle, A.; Fu, X.; He, J.; Song, J.; Zhang, W.; Sun, X.; Ren, Y.; Silvennoinen, O.; et al. Tudor staphylococcal nuclease (Tudor-SN), a novel regulator facilitating G1/S phase transition, acting as a co-activator of E2F-1 in cell cycle regulation. J. Biol. Chem. 2015, 290, 7208–7220. [Google Scholar] [CrossRef] [PubMed]
- Kannan, N.; Eaves, C.J. Tipping the balance: MTDH-SND1 curbs oncogene-induced apoptosis and promotes tumorigenesis. Cell Stem Cell 2014, 15, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Wang, C.; Hu, X.; Pang, B.; Yang, Z.; He, Z.; He, M.; Wei, L.; Chu, M. Correlated overexpression of metadherin and SND1 in glioma cells. Biol. Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Fu, X.; Song, J.; Zhang, Y.; Cui, X.; Su, C.; Ge, L.; Shao, J.; Xin, L.; Saarikettu, J.; et al. Poly(A)+ mRNA-binding protein Tudor-SN regulates stress granules aggregation dynamics. FEBS J. 2015, 282, 874–890. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Shi, X.; Fu, X.; Ge, L.; Zhang, Y.; Su, C.; Yang, X.; Silvennoinen, O.; Yao, Z.; He, J.; et al. Human Tudor staphylococcal nuclease (Tudor-SN) protein modulates the kinetics of AGTR1-3’UTR granule formation. FEBS Lett. 2014, 588, 2154–2161. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Yue, W.; Wang, Y. The nuclear factor κB (NF-κB) activation is required for phagocytosis of staphylococcus aureus by RAW 264.7 cells. Exp. Cell Res. 2014, 327, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Odqvist, L.; Montes-Moreno, S.; Sánchez-Pacheco, R.E.; Young, K.H.; Martín-Sánchez, E.; Cereceda, L.; Sánchez-Verde, L.; Pajares, R.; Mollejo, M.; Fresno, M.F.; et al. NFκB expression is a feature of both activated B-cell-like and germinal center B-cell-like subtypes of diffuse large B-cell lymphoma. Mod. Pathol. 2014, 27, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lin, Y.; Bian, Y.; Liu, L.; Shao, L.; Lin, L.; Qu, B.; Zhao, F.; Gao, X.; Li, Q. Leucyl-tRNA synthetase regulates lactation and cell proliferation via mTOR signaling in dairy cow mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 5952–5969. [Google Scholar] [CrossRef] [PubMed]
- Kampa-Schittenhelm, K.M.; Heinrich, M.C.; Akmut, F.; Rasp, K.H.; Illing, B.; Döhner, H.; Döhner, K.; Schittenhelm, M.M. Cell cycle-dependent activity of the novel dual PI3K-MTORC1/2 inhibitor VP-BGT226 in acute leukemia. Mol. Cancer 2013, 12, 46. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ao, J.; Wei, C.; Si, Y.; Luo, C.; Lv, W.; Lin, Y.; Cui, Y.; Gao, X. Tudor-SN Regulates Milk Synthesis and Proliferation of Bovine Mammary Epithelial Cells. Int. J. Mol. Sci. 2015, 16, 29936-29947. https://doi.org/10.3390/ijms161226212
Ao J, Wei C, Si Y, Luo C, Lv W, Lin Y, Cui Y, Gao X. Tudor-SN Regulates Milk Synthesis and Proliferation of Bovine Mammary Epithelial Cells. International Journal of Molecular Sciences. 2015; 16(12):29936-29947. https://doi.org/10.3390/ijms161226212
Chicago/Turabian StyleAo, Jinxia, Chengjie Wei, Yu Si, Chaochao Luo, Wei Lv, Ye Lin, Yingjun Cui, and Xuejun Gao. 2015. "Tudor-SN Regulates Milk Synthesis and Proliferation of Bovine Mammary Epithelial Cells" International Journal of Molecular Sciences 16, no. 12: 29936-29947. https://doi.org/10.3390/ijms161226212
APA StyleAo, J., Wei, C., Si, Y., Luo, C., Lv, W., Lin, Y., Cui, Y., & Gao, X. (2015). Tudor-SN Regulates Milk Synthesis and Proliferation of Bovine Mammary Epithelial Cells. International Journal of Molecular Sciences, 16(12), 29936-29947. https://doi.org/10.3390/ijms161226212