
Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Nutritional Composition

{kind=link}
{kind=link}
{kind=link}
| Parameter | Tropaeolum majus | Tagetes erecta | Spilanthes oleracea |
|---|---|---|---|
| Moisture (%) | 89.32 ± 0.16 a | 83.39 ± 0.17 b | 81.74 ± 0.13 c |
| Total carbohydrates (%) | 7.14 ± 0.87 c | 14.15 ± 1.24 a | 13.56 ± 0.79 b |
| TDF (%) | 4.51 ± 0.52 b | 9.20 ± 0.04 a | 10.11 ± 0.41 a |
| Protein (%) | 1.99 ± 0.06 b | 1.32 ± 0.01 b | 2.84 ± 0.11 a |
| Fat (%) | 0.33 ± 0.03 a | 0.32 ± 0.02 a | 0.41 ± 0.03 a |
| Ash (%) | 0.63 ± 0.01 c | 0.80 ± 0.05 b | 1.44 ± 0.02 a |
| Energy (kcal/100 g) | 21.44 ± 0.89 b | 28.02 ± 1.1 a | 28.84 ± 1.20 a |
| Mineral | Tropaeolum majus | Tagetes erecta | Spilanthes oleracea |
|---|---|---|---|
| Ca (mg/100 g) | 0.055 ± 0.007 a | 0.110 ± 0.042 a | 0.105 ± 0.035 a |
| Cu (mg/100 g) | 0.472 ± 0.020 a | 0.104 ± 0.025 b | 0.165 ± 0.057 b |
| Fe (mg/100 g) | 0.551 ± 0.074 a | 1.026 ± 0.052 a | 1.500 ± 0.540 a |
| K (mg/100 g) | 0.225 ± 0.007 a | 0.215 ± 0.007 a | 0.355 ± 0.007 b |
| Mg (mg/100 g) | 0.035 ± 0.007 a | 0.060 ± 0.00 a | 0.06 ± 0.028 a |
| Mn (mg/100 g) | 0.397 ± 0.026 a | 0.303 ± 0.027 a | 0.555 ± 0.239 a |
| Na (mg/100g) | 0.010 ± 0.00 a | 0.015 ± 0.007 a | 0.010 ± 0.00 a |
| P (mg/100 g) | 0.050 ± 0.000 a | 0.065 ± 0.007 a | 0.080 ± 0.020 a |
| S (mg/100 g) | 0.040 ± 0.000 a | 0.045 ± 0.007 a | 0.060 ± 0.014 a |
| Sr (mg/100 g) | 0.388 ± 0.002 a | 1.017 ± 0.470 a | 0.897 ± 0.328 a |
| Zn (mg/100 g) | 0.660 ± 0.064 a | 0.568 ± 0.093 a | 0.543 ± 0.144 a |
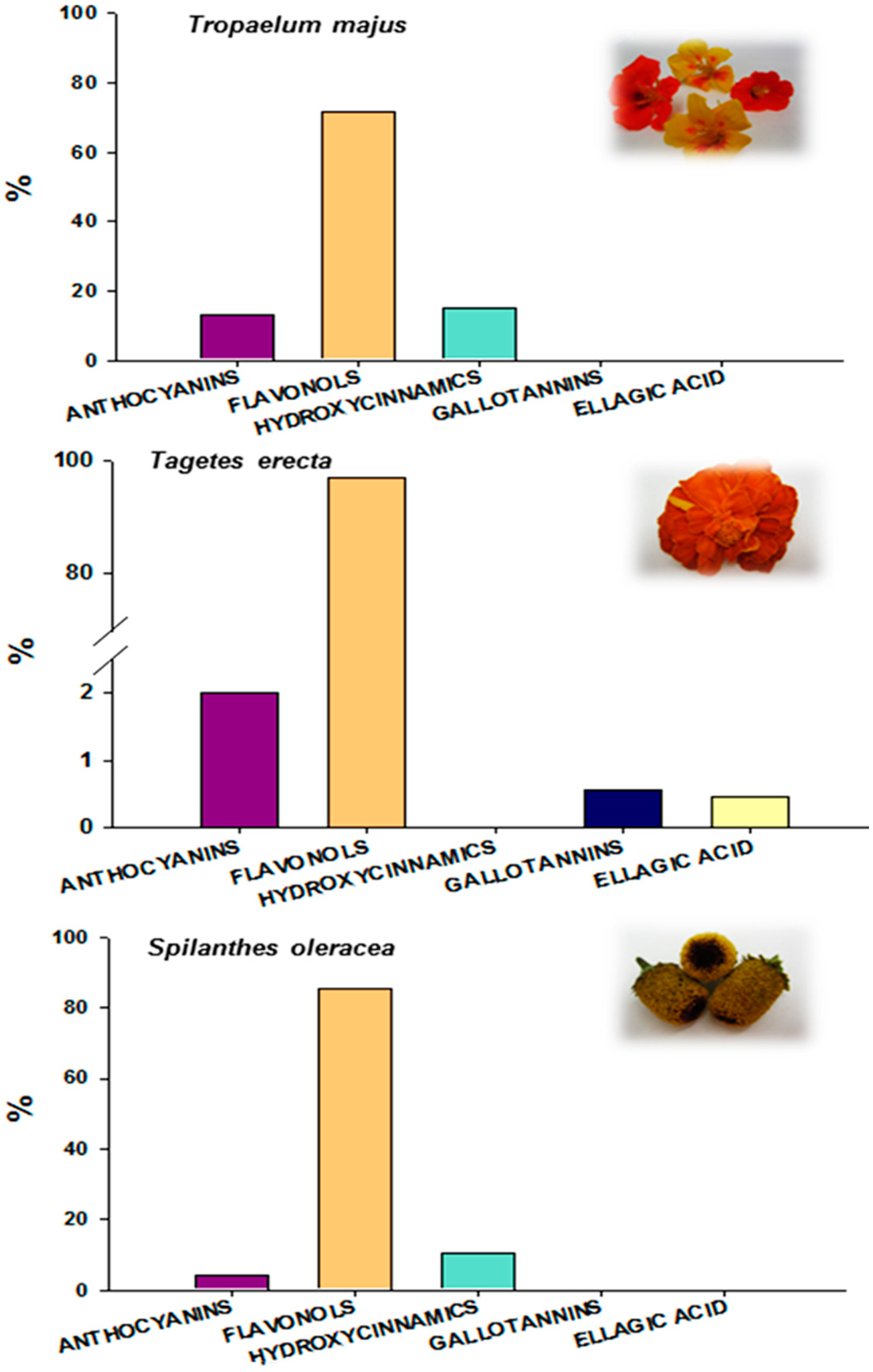
2.2. Total Penolic Compounds and Their Identification by HPLC-DAD-MSn
| Parameters | Tropaeolum majus | Tagetes erecta | Spilanthes oleracea |
|---|---|---|---|
| TPC (mg GAE/g) | 12.95 ± 2.21 b | 26.63 ± 4.22 a | 6.64 ± 0.45 c |
| ORAC (µmol TE/g) | 47.84 ± 0.80 b | 266.11 ± 55.9 a | 10.82 ± 0.53 c |
| TEAC (µmol TE/g) | 9.51 ± 0.10 a | 66.22 ± 1.10 b | 5.52 ± 0.13 c |
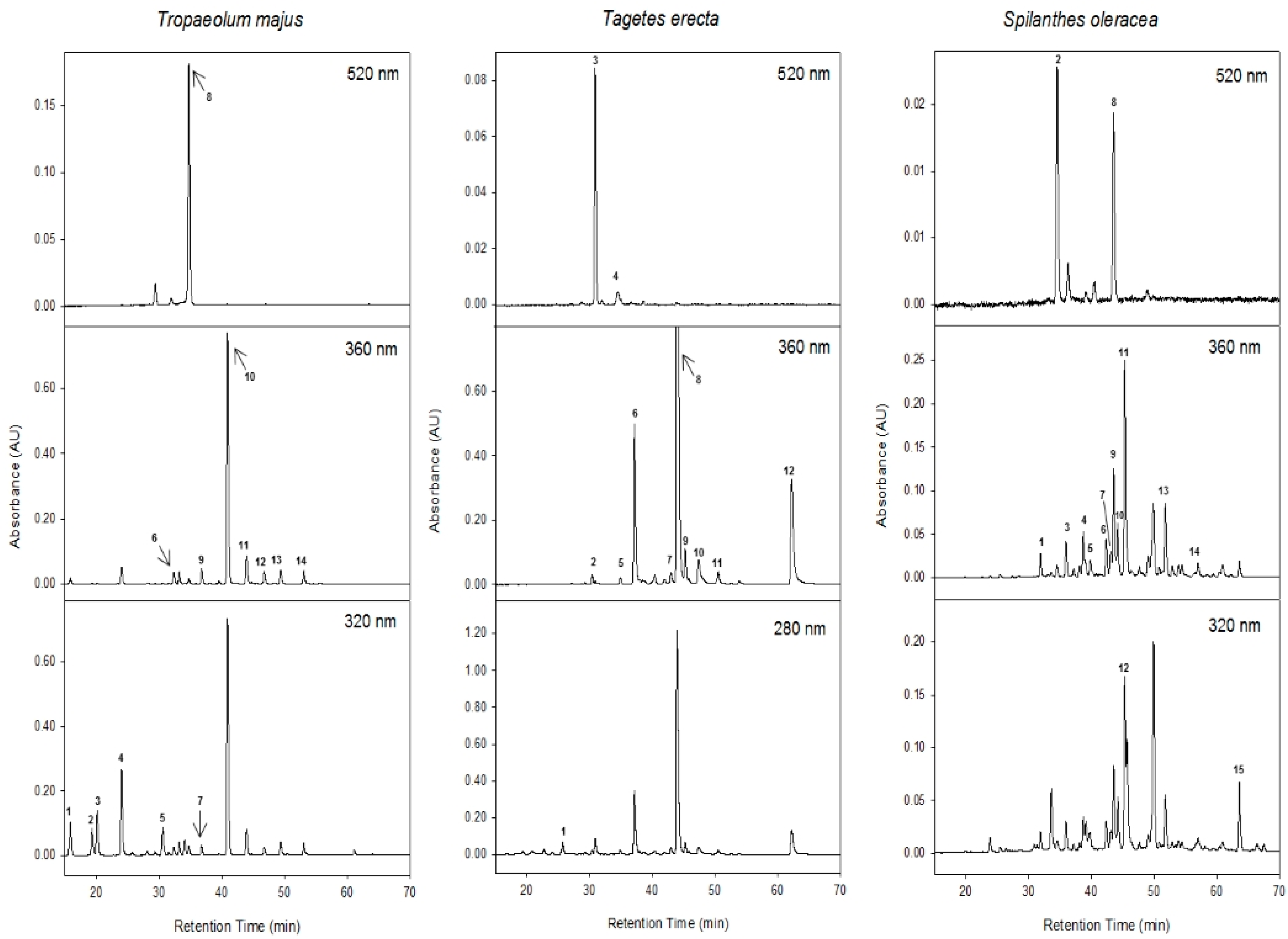
| Sample | Peak | Rt (min) | Tentative Identification | λmax (nm) | [M-H]− (m/z) | MS2 (m/z) |
|---|---|---|---|---|---|---|
| Tropaeolum majus | 1 | 15.8 | 3-O-caffeoylquinic acid | 325 | 353 | 191, 179 |
| 2 | 19.3 | cis-3-O-p-coumaroylquinic acid | 305 | 337 | 163, 173 | |
| 3 | 20.1 | trans-3-O-p-coumaroylquinic acid | 310 | 337 | 163, 173 | |
| 4 | 24.0 | 5-O-caffeoylquinic acid | 325 | 353 | 191, 179 | |
| 5 | 30.6 | trans-5-O-p-coumaroylquinic acid | 311 | 337 | 191, 163 | |
| 6 | 32.3 | Myricetin-3-O-sophoroside | 354 | 641 | 317, 461, 479 | |
| 7 | 33.2 | cis-5-O-p-coumaroylquinic acid | 305 | 337 | 191, 163 | |
| 8 | 34.7 | Pelargonidin-3-O-sophoroside | 502 | 595 * | 271, 415 | |
| 9 | 36.8 | Quercetin-3-O-sophoroside | 353 | 625 | 301, 445 | |
| 10 | 40.9 | Kaempferol-3-O-sophoroside | 347 | 609 | 285, 429 | |
| 11 | 43.9 | Kaempferol-3-O-acetyl-sophoroside | 348 | 651 | 471, 285, 489 | |
| 12 | 46.8 | Quercetin-O-acetylhexoxide | 355 | 505 | 301, 463 | |
| 13 | 49.4 | Kaempferol-3-O-hexoside | 348 | 447 | 285 | |
| 14 | 53.1 | Kaempferol-O-acetylhexoxide | 348 | 489 | 285 | |
| Tagetes erecta | 1 | 25.7 | Gallotannin | 277 | 797 | 645, 627, 493 |
| 2 | 30.4 | Laricitrin-di-hexoside | 356 | 655 | 493, 331 | |
| 3 | 30.8 | Cyanidin-di-hexoside | 518 | 611 * | 449, 287 | |
| 4 | 34.9 | Delphinidin-3-O-hexoside | 516 | 465 * | 303 | |
| 5 | 35.7 | Myricetin-hexoside | 357 | 479 | 317 | |
| 6 | 37.1 | Laricitrin-hexoside | 358 | 493 | 331 | |
| 7 | 42.9 | Ellagic acid | 365 | 301 | 257, 229 | |
| 8 | 43.9 | Laricitrin-hexoside | 354 | 493 | 331 | |
| 9 | 45.2 | Laricitrin-hexoside | 352 | 493 | 331 | |
| 10 | 47.3 | Myricetin | 360 | 317 | 299, 271, 167 | |
| 11 | 50.5 | Isorhamnetin-3-O-hexoside | 354 | 477 | 315 | |
| 12 | 62.2 | Laricitrin | 364 | 331 | 316 | |
| Spilanthes oleracea | 1 | 33.6 | Quercetin-deoxyhexoside-di-hexoside | 353 | 771 | 625, 446, 301 |
| 2 | 34.5 | Cyanidin-3-O-glucoside | 517 | 449 * | 287 | |
| 3 | 35.9 | Quercetin-dihexoside | 354 | 625 | 301, 463 | |
| 4 | 38.1 | Quercetin-rhamnosyl-hexoside | 356 | 609 | 447, 463, 301 | |
| 5 | 38.7 | Quercetin-rhamnosyl-rutinoside | 355 | 755 | 609, 301 | |
| 6 | 42.3 | Quercetin-3-O-rutinoside | 356 | 609 | 301 | |
| 7 | 43.1 | Quercetin-3-O-glucoside | 353 | 463 | 301 | |
| 8 | 43.5 | Delphinidina-3-O-glucuronide | 517 | 479 * | 303 | |
| 9 | 43.6 | Quercetin-3-O-glucuronide | 354 | 477 | 301 | |
| 10 | 44.2 | Quercetin-acetylhexose-deoxyhexoside | 352 | 651 | 609, 505, 447, 301 | |
| 11 | 45.3 | Quercetin-acetyl dihexoside | 354 | 667 | 625, 301 | |
| 12 | 49.9 | dicaffeoylquinic acid | 330 | 515 | 353 | |
| 13 | 51.8 | Quercetin-acetyl hexoside | 356 | 505 | 463, 301 | |
| 14 | 57.0 | Quercetin-diacetyl hexoside | 353 | 547 | 505, 463, 301 | |
| 15 | 63.6 | Caffeoylquinic acid dihexose derivative | 326 | 677 | 515, 353 |
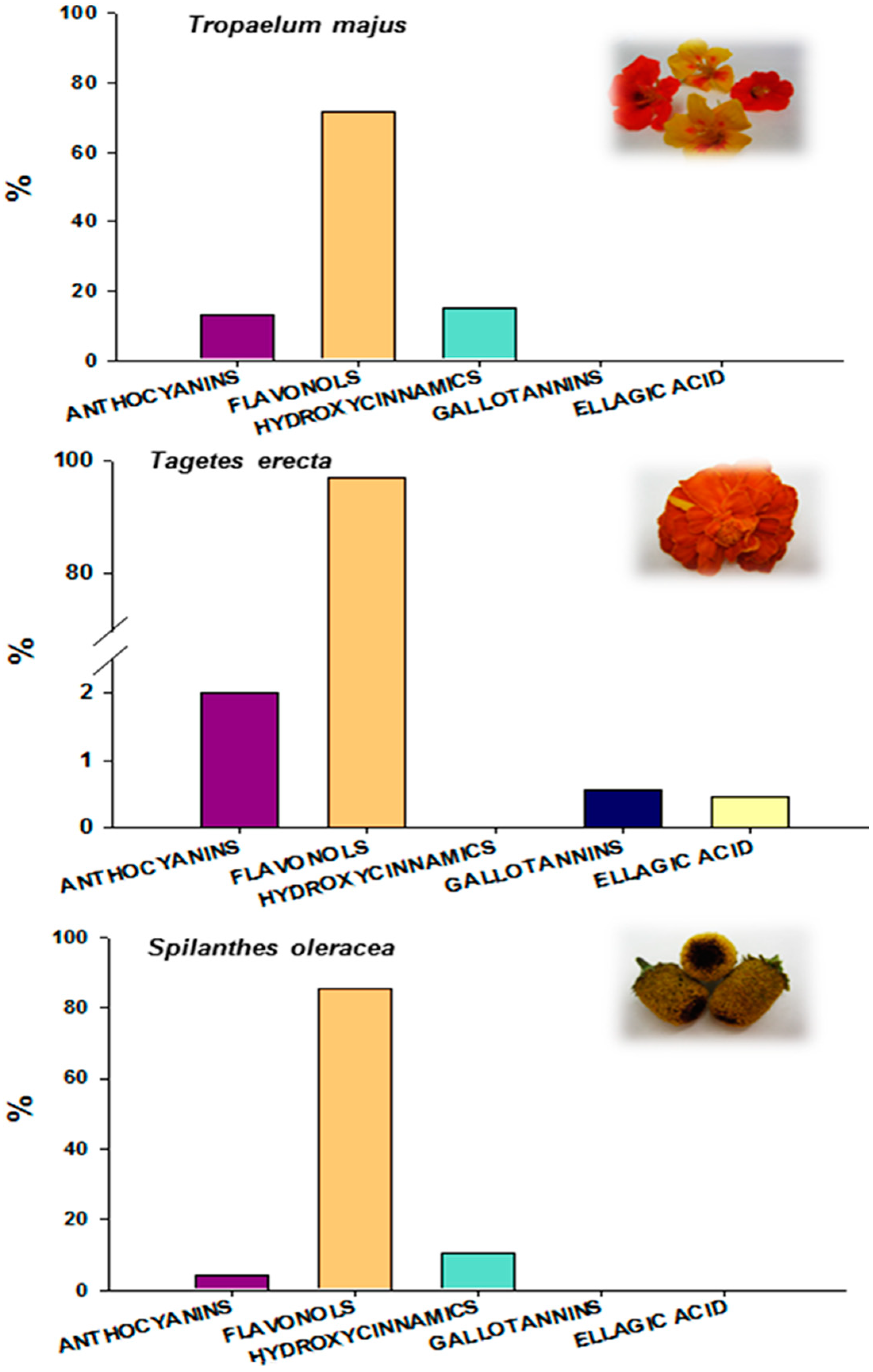
2.3. Antioxidant Capacity
| Parameters | rORAC | p | rTEAC | p |
|---|---|---|---|---|
| TPC | 0.865 | <0.05 | 0.931 | <0.05 |
| Total anthocyanins | −0.182 | ns | −0.259 | ns |
| Total flavonols | 0.976 | <0.01 | 0.987 | <0.01 |
| Total hydroxycinnamic acids | −0.596 | ns | −0.666 | ns |
| Hydrolysable tannins | 0.971 | <0.01 | 0.998 | <0.01 |
| Ellagic acid | 0.971 | <0.01 | 0.998 | <0.01 |
| Σ Individual phenolics | 0.953 | <0.01 | 0.952 | <0.01 |
3. Experimental Section
3.1. Samples
3.2. Standards and Reagents
3.3. Proximate and Mineral Composition
3.4. Preparation of the Flower Extracts
3.5. Total Phenolic Compounds
3.6. Hydrophilic Antioxidant Capacity Assays
3.6.1. TEAC Assay
3.6.2. ORAC assay
3.7. Analysis of Phenolic Compounds by HPLC-DAD and HPLC-DAD-ESI-MSn
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kaisoon, O.; Siriamornpun, S.; Weerapreeyakul, N.; Meeso, N. Phenolic compounds and antioxidant activities of edible flowers from Thailand. J. Funct. Foods 2011, 3, 88–99. [Google Scholar] [CrossRef]
- Mlcek, J.; Rop, O. Fresh edible flowers of ornamental plants are a new source of nutraceutical foods. Trends Food Sci. Technol. 2011, 22, 561–569. [Google Scholar] [CrossRef]
- Lara-Cortés, E.; Osorio-Díaz, P.; Jiménez-Aparicio, A.; Bautista-Baños, S. Contenido nutricional, propiedades funcionales y conservación de flores comestibles. Arch. Latinoam. Nutr. 2013, 63, 197–208. [Google Scholar] [PubMed]
- Alasalvar, C.; Pelvan, E.; Özdemir, K.S.; Kocadagh, T.; Mogol, B.A.; Pash, A.A.; Özcan, N.; Özçelik, B.; Gökmen, V. Compositional, nutritional, and functional characteristics of instant teas produced from low- and high-quality black teas. J. Agric. Food Chem. 2013, 61, 7529–7536. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, A.; López-García, S.; Basurto-Peña, F. Content of nutrients and antinutrients in edible flowers of wild plants in Mexico. Plant Foods Hum. Nutr. 2007, 62, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Nnam, N.M.; Onyeke, N.G. Chemical composition of two varieties of sorrel (Hibiscus sabdariffa L.) calyces and the drinks made from them. Plant Food Hum. Nutr. 2003, 58, 1–7. [Google Scholar]
- Rop, O.; Mlcek, J.; Jurikova, T.; Neugebauerova, J. Edible flowers—A new promising source of mineral elements in human nutrition. Molecules 2012, 17, 6672–6683. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Alamb, M.S.; Jabba, Z.; Javed, K.; Athar, M. Evaluation of antioxidant activity of Cassia siamea flowers. J. Ethnopharmacol. 2006, 108, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Youwei, Z.; Jinlian, Z.; Yonghong, P. A comparative study on the free radical scavenging activities of some fresh flowers in southern China. LWT Food Sci. Technol. 2008, 41, 1586–1591. [Google Scholar] [CrossRef]
- Song, L.; Wang, X.; Zheng, X.; Huang, D. Polyphenolic antioxidant profiles of yellow camellia. Food Chem. 2011, 129, 351–357. [Google Scholar] [CrossRef]
- Issa, A.Y.; Volate, S.R.; Wargovich, M.J. The role of phytochemicals in inhibition of cancer and inflammation: new directions and perspectives. J. Food Compos. Anal. 2006, 19, 405–419. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Visioli, F. Polyphenols and health: moving beyond antioxidants. J. Berry Res. 2012, 2, 63–71. [Google Scholar]
- Niizu, P.Y.; Rodriguez-Amaya, D.B. Flowers and leaves of Tropaeolum majus L. as rich sources of lutein. J. Food Sci. 2005, 70, 605–609. [Google Scholar] [CrossRef]
- Pintão, A.M.; Pais, M.S.; Coley, H.; Kelland, L.R.; Judson, I.R. In vitro and in vivo antitumor activity ob benzyl isothiocyanate: A natural product from Tropaeolum majus. Planta Medica 1995, 61, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Martínez, R.; Díaz, B.; Vásquez, L.; Compagnone, R.S.; Tillet, S.; Canelon, D.J.; Torrico, F.; Suárez, A.I. Chemical composition of essential oils and toxicological evaluation of Tagetes erecta and Tagetes patula from Venezuela. J. Essent. Oil Res. 2009, 12, 476–481. [Google Scholar]
- Hellinger, R.; Koehbach, J.; Fedchuk, H.; Sauer, B.; Huber, R.; Gruber, C.W.; Gründemann, C. Immunosuppressive activity of an aqueous Viola tricolor herbal extract. J. Ethnopharmacol. 2014, 151, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Goos, K.H.; Albercht, U.; Schneider, B. Eficacy and safety profile of a herbal drug containing nasturtium herb and horseradish root in acute sinusitis, acute bronchitis ad acute urinary tract infection in comparison with other treatments in the daily practice/results of a prospective cohort study. Arzneimittelforschung 2006, 56, 249–257. [Google Scholar] [PubMed]
- De Medeiros, J.M.R.; Macedo, M.; Contancia, J.P.; Nguyen, C.; Cunningham, G.; Miles, D.H. Antithrombin activity of medical plants of the Azores. J. Ethnopharmacol. 2000, 72, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Arquímedes, G.J.; Marcos, A.B.; Emerson, L.B.L.; María, E.A.S.; Cândida, A.L.K.; María, C.A.M. Natriuretic and diuretic effect of Tropaeolum majus (Tropaeolaceae) in rats. J. Ethnopharmacol. 2009, 122, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Arquímedes, G.J.; Francielly, M.G.; Emerson, L.B.L.; Sandra, C.; María, E.A.S.; Marcos, J.S.; José, E.S.S.; M, C.A.M.; Cândida, A.L.K. Antihypertensive effects of isoquercitrin and extracts from Tropaeolum majus L.: Evidence for the inhibition of angiotensin converting enzyme. J. Ethnopharmacol. 2011, 134, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Wongsawatkul, O.; Prachayasittikul, S.; Isarankura-Na-Ayudhya, C.; Satayavivad, J.; Ruchirawat, S.; Prachayasittiku, V. Vasorelaxant and antioxidant activities of Spilanthes acmella Murr. Int. J. Mol. Sci. 2008, 9, 2724–2744. [Google Scholar] [CrossRef] [PubMed]
- Savadi, R.V.; Yadav, R.; Yadav, N. Study on immunomodulatory activity of ethanolic extract of Spilanthes acmella Murr. leaves. Ind. J. Nat. Prod. Res. 2010, 1, 204–207. [Google Scholar]
- Ratnasooriya, W.D.; Pieris, K.P.; Samaratunga, U.; Jayakody, J.R. Diuretic activity of Spilanthes acmella flowers in rat. J. Ethnopharmacol. 2004, 91, 185–205. [Google Scholar] [CrossRef]
- Vasudevan, P.; Kashyap, S.; Sharma, S. Tagetes: A multipurpose plant. Bioresour. Technol. 1997, 62, 29–35. [Google Scholar] [CrossRef]
- Bashir, S.; Gilani, A.H. Studies on the antioxidant and analgesic activities of Aztec marigold (Tagetes erecta) flowers. Phytother. Res. 2008, 22, 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- Khomdram, S.D.; Salam, J.S.; Singh, P.K. Estimation of nutritive indices in eight Lamiaceae plants of Manipur. Am. J. Food Technol. 2011, 6, 924–931. [Google Scholar] [CrossRef]
- Kruger, M.; Sayed, N.; Langenhoven, M.; Holing, F. Composition of South African Foods: Vegetables and Fruit; Medical Research Council: Cape Town, South Africa, 1998. [Google Scholar]
- Mosha, T.C.; Gaga, H.E. Nutritive value and effect of blanching on the trypsin and chymotrypsin inhibitor activities of selected leafy vegetables. Plant Food Hum. Nutr. 1999, 54, 271–283. [Google Scholar] [CrossRef]
- Odhav, B.; Beekrum, S.; Akula, Us.; Baijnath, H. Preliminary assessment of nutritional value of traditional leafy vegetables in KwaZulu-Natal, South Africa. J. Food Compos. Anal. 2007, 20, 430–435. [Google Scholar] [CrossRef]
- Martins, D.; Barros, L.; Carvalho, A.M.; Ferreira, C.F.R. Nutritional and in vitro antioxidant properties of edible wild greens in Iberian Peninsula traditional diet. Food Chem. 2011, 125, 488–494. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in food and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Garzón, G.A.; Wrolstad, R.E. Major anthocyanins and antioxidant activity of nasturtium flowers (Tropaelum majus). Food Chem. 2009, 114, 44–49. [Google Scholar] [CrossRef]
- Khoudja, N.K.; Boulekbache-Makhlouf, L.; Madani, K. Antioxidant capacity of crude extracts and their solvent fractions of selected Algerian Lamiaceae. Ind. Crop. Prod. 2014, 52, 177–182. [Google Scholar] [CrossRef]
- Kaisoon, O.; Konczak, I.; Siriamornpun, S. Potential health enhancing properties of edible flowers from Thailand. Food Res. Int. 2012, 46, 563–571. [Google Scholar] [CrossRef]
- Mullen, W.; Yokota, T.; Lean, M.E. J.; Crozier, A. Analysis of ellagitannins and conjugates of ellagic acid and quercetin in raspberry fruits by LC-MSn. Phytochemistry 2003, 64, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cuyckens, F.; van den Heuvel, H.; Apers, S.; Pieters, L.; Steenkamp, V.; Stewart, M.J.; Luyckx, V.A.; Claeys, M. Structural characterization of chromone C-glucosides in a toxic herbal remedy. Rapid Commun. Mass. Spectrom. 2003, 17, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six idomers of dicaffeoylquinic acid by LC-MSn. J. Agric. Food Chem. 2005, 53, 3821–3232. [Google Scholar] [CrossRef] [PubMed]
- Regos, I.; Urbanella, A.; Treutter, D. Identification and quantification of phenolic compounds from the forage legume sainfoin (Onobrychis viciifolia). J. Agric. Food Chem. 2009, 57, 5843–5852. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.I.; Barros, L.; Alves, R.C.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C. F.R. Nutritional composition, antioxidant activity and phenolic compounds of wild Taraxacum sect. Ruderalia Food Res. Int. 2014, 56, 266–271. [Google Scholar] [CrossRef]
- Ferreres, F.; Llorach, R.; Gil, A. Characterization of the interglycosidic linkage in di-, tri-, tetra- and pentaglycosylated flavonoids and differentiation of positional isomers by liquid chromatography/electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2004, 39, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. Comparative biochemistry of the flavonoids. VII. Correlations between flavonoid pigmentation and systematics in the family Primulaceae. Phytochemistry 1968, 7, 1215–1230. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef] [PubMed]
- Määttä, K.R.; Kamal-Eldin, A.; Törrönen, R. High-Performance liquid chromatography (HPLC) analysis of phenolic compounds in berries with diode array and electrospray ionization mass spectrometric (MS) detection: Ribes species. J. Agric. Food Chem. 2003, 51, 6736–6744. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; García-Conesa, M.T.; Espín, J.C.; Tomás-Barberán, F.A. Ellagitannins, ellagic acid and vascular health. Mol. Aspects Med. 2010, 31, 513–539. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Dueñas, D.; Carvalho, A.M.; Ferreira, I.C.F.R.; Santos-Buelga, C. Characterization of phenolic compounds in flowers of wild medicinal plants from Northeastern Portugal. Food Chem. Toxicol. 2012, 50, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.W.; Rautenbach, F.; Mabusela, M.T.; Valentine, A.J.; Marnewick, J.L. Comparative antioxidant-capacity and content of leaves, bulbs, roots, flowers and fruit of Gethyllis multifolia L. Bolus and G. villosa Thunb. species. S. Afr. J. Bot. 2011, 77, 711–717. [Google Scholar] [CrossRef]
- Tai, Z.; Cai, L.; Dai, L.; Dong, L.; Wang, M.; Yang, Y.; Cao, Q.; Ding, Z. Antioxidant activity and chemical constituents of edible flower of Sophora viciifolia. Food Chem. 2011, 126, 1648–1654. [Google Scholar] [CrossRef] [PubMed]
- Takashima, M.; Horie, M.; Shichiri, M.; Hagihara, Y.; Yoshida, Y.; Niki, E. Assessment of antioxidant capacity for scavenging free radicals in vitro: A rational basic and practical application. Free Radic. Biol. Med. 2012, 52, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- García-Valverde, V.; Navarro-González, I.; García-Alonso, J.; Periago, M.J. Antioxidant bioactive compounds in selected industrial processing and fresh consumption tomato cultivars. J. Food Bioprocess. Technol. 2013, 6, 391–402. [Google Scholar] [CrossRef]
- Borges, G.; Mullen, W.; Crozier, A. Comparison of the polyphenolic composition and antioxidant activity of European commercial fruit juices. Food Funct. 2010, 1, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef] [PubMed]
- AOAC (Official methods of analysis). Gaithrsburg, MD: Association of Official Analytical Chemists, 15th ed.; Board: Washington, DC, USA, 1990. [Google Scholar]
- Prosky, P.; Asp, N.G.; Scheweizer, T.F.; Devries, J.W.; Furda, I. Determination of insoluble, soluble, and total dietary fiber in foods, food products: Interlaboratory study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [PubMed]
- Singleton, V.L.; Rossi, A.J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Miller, N.J.; Sampson, J.; Candelas, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for hydrophilic and lipophilic antioxidant capacity (oxygen radical absorbance capacity (ORACFL)) of plasma and other biological and food samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-González, I.; González-Barrio, R.; García-Valverde, V.; Bautista-Ortín, A.B.; Periago, M.J. Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn. Int. J. Mol. Sci. 2015, 16, 805-822. https://doi.org/10.3390/ijms16010805
Navarro-González I, González-Barrio R, García-Valverde V, Bautista-Ortín AB, Periago MJ. Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn. International Journal of Molecular Sciences. 2015; 16(1):805-822. https://doi.org/10.3390/ijms16010805
Chicago/Turabian StyleNavarro-González, Inmaculada, Rocío González-Barrio, Verónica García-Valverde, Ana Belén Bautista-Ortín, and María Jesús Periago. 2015. "Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn" International Journal of Molecular Sciences 16, no. 1: 805-822. https://doi.org/10.3390/ijms16010805
APA StyleNavarro-González, I., González-Barrio, R., García-Valverde, V., Bautista-Ortín, A. B., & Periago, M. J. (2015). Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn. International Journal of Molecular Sciences, 16(1), 805-822. https://doi.org/10.3390/ijms16010805