Complete Proteome of a Quinolone-Resistant Salmonella Typhimurium Phage Type DT104B Clinical Strain

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serotype | Strain | Main Purpose | Main Findings | Ref. |
|---|---|---|---|---|
| Typhimurium and Typhi | LT2 and Ty2 | To perform a quantitative comparative proteomic analysis between S. Typhimurium and S. Typhi using SILAC coupled with LC-MS/MS. | Potential biomarker proteins with serovar-specific expression were identified. Flagella and chemotaxis genes were down-regulated in S. Typhi and proteins involved in metabolism and transport of carbohydrates and amino acids were differentially expressed. | [20] |
| Infantis | Soil isolate (from cattle manure) | To elucidate the global modulation of bacteria and plant protein expression after Salmonella internalization into lettuce. | Fifty proteins were differentially expressed between internalized and cutured S. Infantis. Internalized S. Infantis triggered the lettuce defense mechanisms. The bacteria might use ascorbate as a carbon source and require stress response proteins to cope with stresses incurred in plants. | [21] |
| Paratyphi A | YN07077, GZ9A0503, ZJ98053, ATCC 9150 | To perform a 2-DE comparative proteomics analysis for 4 epidemic strains with different geospatial and temporal characteristics in order to obtain their core and pan proteomes. | The proteomes of the four strains were highly conserved. Few strain-specific proteins were found and non-core proteins were found in similar categories as core proteins. Significant fluctuations in the abundance of some core proteins suggest a variation in protein expression in the different strains even when cultured in the same conditions. | [22] |
| Typhimurium | ATCC 14028 | To profile the intact proteome by single-dimension ultra-high-pressure liquid chromatography coupled with Velos-Orbitrap MS. | Identification of 563 proteins including 1665 proteoforms generated by PTMs. Report of a unique protein S-thiolation switch in response to infection-like conditions. | [23] |
| Typhimurium | ATCC 14028 | To observe changes in protein abundance or location between phagosome-mimicking and standard laboratory conditions. | The protein subcellular localization of over 1000 proteins was catalogued. New insights into dynamic protein localization and potential moonlighting. | [24] |
| Typhimurium | ST23 | To elucidate biocide tolerance mechanisms by comparing 2-D DIGE protein profiles of a triclosan sensitive strain and the isogenic tolerant mutant in the presence and absence of triclosan. | Triclosan exposure induced multiple changes in cellular metabolism, permeability, transport and also modifications involving mutations in the triclosan specific target FabI. Broader changes that may confer cross-resistance to antimicrobial agents were also observed. | [25] |
| Typhimurium | SL1344 | To analyze differentially expressed proteins between a wild-type strain and an opgGH mutant to elucidate proteomic pleiotropy under low osmolarity. | The opgGH mutant had decreased protein amounts, consistent with the genotype and the expected phenotypes, and revealed pleiotropic proteome effects likely to enable survival under low-nutrient and low-osmotic growth conditions. | [26] |
| Typhimurium, Typhi and Choleraesuis | LT2, ATCC 33458 and SC-B67 | To analyze the ability of pseudogenes to express normal protein sequences and to develop an experimental approach to detect recoding at the genomic scale using LC-MS/MS. | The majority of pseudogenes failed to express, validating the overall accuracy of in silico annotation. A few annotated pseudogenes translated normal peptides, suggesting that recoding may be common in bacterial species. | [27] |
| Gallinarum | 9R and WT (287/91 and 06Q110) | To compare the proteome and transcriptome of wild-type and live vaccine strains of S. Gallinarum by 2-DE MALDI-TOF MS and microarray analysis. | One protein relevant to virulence absent from 9R. Analysis revealed 42 virulence genes down-regulated in the 9R transcriptome. The attenuation of 9R may be associated with a combination of impaired virulence factors so reversion to virulence is probably not caused by single mutation. | [28] |
| Enteritidis, Typhimurium and Gallinarum | Human and chicken isolates; 9R | To examine protein profile variability among S. Enteritidis, S. Typhimurium and S. Gallinarum by a comparative 2-DE MALDI-TOF MS proteomic analysis. | A high level of variation between serotypes was observed and several serotype-specific factors were detected. Proteins related to virulence, such as β-lactamase, RfbH protein, and shikimate kinase were identified. | [29] |
| Typhimurium | - | To characterize the proteome and ionome of wild type and znuA mutant strains under Zn starvation or Zn-replete conditions to gain further insight into Zn influx regulation. | Several differentially regulated proteins were predicted to be metal-binding proteins; their over-expression in the znuA mutant strain strictly depends on Zn starvation and correlates with differences found at the ionome level. | [30] |
| Typhimurium | VNP20009 | To profile protein expression in the tumor-specific VNP strain under anaerobic and aerobic conditions, and to develop a hypoxia-inducible promoter system to confine expression of therapeutic genes within the tumor microenvironment. | The hypoxia-inducible adhE promoter was screened from the hypoxia-regulated endogenous proteins of Salmonella and proof-of-principle was provided that these promoter systems can be employed to target the hypoxic region of solid tumors and exert enhanced anticancer effects. | [31] |
| Typhimurium | ATCC 14028 | To identify effector proteins secreted under SPI-2-inducing growth conditions using LC-MS/MS. | Eight novel effectors and ~80% of the previously reported ATCC14028 repertoire were identified including novel secreted effectors and new pathways for Salmonella virulence factors. | [32] |
| Typhimurium | SL1344 | To identify post-transcriptional regulatory events by analyzing proteome changes after activation of the RcsCDB regulatory system. | Two new post-transcriptional regulatory processes were defined, inverse regulation by the metE and pckA genes and expression control of the small RNA FnrS by the RcsCDB system. | [33] |
| Typhimurium | MA6926 | To survey the proteomic changes in response to low Mg2+ concentrations or CAMP in a SILAC-based quantitative proteomic approach. | CAMP activates a portion of the PhoP/PhoQ regulatory network. Low Mg2+ concentrations up-regulate nearly all known and some previously unknown members of this network, and also proteins regulated by IHF and RpoS. | [34] |
| Typhi | CT18 | Characterization of anti-S. Typhi antibody responses in bacteremic Bangladeshi patients by immunoaffinity proteomics-based technology. | Identification of 57 proteins whose capture by affinity-purified antibody fractions from plasma of patients with S. Typhi bacteremia was significantly increased compared to the capture by the column without antibody. | [35] |
| Typhimurium | ATCC 14028 | Proteome profiling of wild-type and mutant strains with ProteinChip arrays coupled to SELDI-TOF. | Revelation of differential regulation of the σ-dependent yciGFE(katN) locus by YncC and H-NS in Salmonella and Escherichia coli K-12. | [36] |
| Enteritidis | chicken isolate (LK5) | Global 2-DE MALDI-TOF MS protein analysis of S. Enteritidis adapted or unadapted to propionate. | The stress-related proteins Dps and CpxR5 were up-regulated in propionate-adapted cultures and play an important role in propionate-induced acid resistance. | [37] |
| Enteritidis | clinical isolate (SE2472) | To develop a stable isotope labeling procedure coupled with MS analysis to carry out quantitative proteomic analysis of S. Enteritidis upon exposure to hydrogen peroxide. | Identification of 76 proteins with H2O2 modulated expression. SPI-1 effector SipC was overexpressed and was found to be highly expressed in the spleen at late stage of in vivo infection, suggesting a role of SipC in supporting survival and replication under oxidative stress and during systemic infection in vivo. | [38] |
| Typhimurium and Enteritidis | wild boar and wild rabbit isolates | To determine and compare the proteomes of S. Typhimurium and S. Enteritidis recovered from faecal samples from wild boars and rabbits. | Different serotypes had different SDS-PAGE profiles. Proteins related to antibiotic resistance, pathogenesis and virulence were identified in both strains. | [39] |
| Typhimurium | LT2 (ATCC 700720) | To elucidate the expression of OMPs of S. Typhimurium using a LPI™ Flow-Cell lipid-based protein immobilization technique. | The LPI™ technique provided wide coverage with 54 OMPs identified, enabling the incorporation of a multi-step protease workflow that allows the identification of more membrane proteins with higher confidence. | [40] |
| Thompson | MCV1 | To study the proteome changes of S. Thompson during stress adaptation to sublethal concentrations of thymol with 2-DE MALDI-TOF MS. | Several proteins from different functional classes were significantly up- or down-regulated showing that thymol plays a role in altering very different metabolic pathways. | [41] |
| Typhi and Typhimurium | Ty2, CT18, Ty800 and LT2 | Comparative proteomic analysis to study PhoP/Q-dependent protein expression differences between S. Typhi and S. Typhimurium. | Identification of 53 PhoP-regulated proteins in LT2 and 56 in S. Typhi, including 3 S. Typhi-unique proteins (CdtB, HlyE and STY1499). First protein expression profile of the live attenuated bacterial vaccine studied in humans Ty800. | [42] |
| Typhimurium | clinical isolate and NCTC 74 | To characterize proteins that are differentially expressed in the presence or absence of oxygen to reveal proteins that may allow the species to adapt and initiate infection in anaerobic conditions. | A drastic transformation in expression was observed with the shift to anaerobiosis. The responses of different isolates were not uniform and the high degree of change showed the potential limitation of using laboratory-grown strains to search for vaccine targets. | [43] |
| Typhimurium | DT104 (ATCC 700408) | To determine if protein profiling by GC-MS analysis of fatty acids with PCA and 2-DE can be used for rapid assessment and interpretation of the impact of SC-CO2 treatment. | SC-CO2 caused significant alterations in the fatty acid and protein profiles with 11 spots becoming more than 50% less intense. The low levels of the latter proteins may have negatively affected the survival of microbial cells. | [44] |
| Gallinarum and Enteritidis | JOL394 | To discover host specificity and/or pathogenicity proteins among different host-adapted serovars by 2-DE MALDI-TOF MS/QRT-PCR analysis of serovar Gallinarum in comparison with Enteritidis. | In S. Gallinarum 22 proteins were over-expressed comparing to S. Enteritidis. Proteins were identified that are related to virulence or have unknown functions that may be important in the host adaptation and/or pathogenicity of S. Gallinarum. | [45] |
| Typhimurium | ATCC 14028 | To investigate the macrophage response to infection by infecting RAW 264.7 macrophages and analyzing time course responses at the global proteomic level. | Identification of 1006 macrophage and 115 Salmonella proteins with high confidence. Most of the Salmonella proteins were observed in the late stage of infection, which is consistent with the fact that the bacterial cells proliferate inside RAW 264.7 macrophages. | [46] |
| Typhimurium | ATCC 14028 | To determine the impact of a low Mg2+/pH defined growth medium (MgM) on the proteome of S. Typhimurium by a comparative LC-MS/MS approach. | MgM shock-induced proteins usually induced by low O2. MgM dilution induced the T3SS proteins SsaQ and SseE and also the biotin biosynthesis proteins BioB and BioD that also increased after infection of RAW 264.7 macrophages. | [47] |
| Typhimurium | SL1344 | To investigate the role of AI-2/LuxS by a comparative 2D-DIGE analysis of wild type and a luxS mutant strain. | A few proteins were differentially expressed but further analysis of the LuxS protein revealed a PTM and a potential translocation across the cytoplasmic membrane. | [48] |
| Typhimurium | SL1344 | To investigate the combined effect of low oxygen tension and high osmolarity on the proteome of S. Typhimurium compared to standard laboratory conditions by 2-D DIGE. | Under in vivo-like conditions anaerobic fumarate respiration and the utilization of 1,2-propanediol are up-regulated and an arginine deiminase pathway is expressed for l-arginine catabolism. Proteins involved in quorum sensing and virulence are also differentially expressed. | [49] |
| Typhimurium | SL1344 | To determine and compare the proteomes of three triclosan resistant mutants to identify proteins involved in triclosan resistance. | Proteins involved in pyruvate or fatty acid production were differentially expressed in all mutants. Triclosan resistance is multifactorial and several resistance mechanisms act in synergy to achieve high-level resistance. | [50] |
| Typhimurium | ATCC 13311 | Characterization of the OMP-immunoreactive fractions in Salmonella induced reactive arthritis by SDS-PAGE and MALDI-TOF MS. | Identification of 10 low molecular weight OMPs which are T-cell immunoreactive in patients with Salmonella induced reactive arthritis/undifferentiated spondyloarthropathy. | [51] |
| Typhimurium | 01-45, R200 and 6B7 | To compare OMP profiles between a yjeH mutant with reduced resistance to ceftriaxone and the resistant parental strain, by 2-DE MALDI-TOF MS/MS. | yjeH gene inactivation resulted in a 4-fold reduction in ceftriaxone resistance and in an underexpression of STM1530, STM3031, MopA, and NuoB, but overexpression of OmpD. Expression of the S. Typhimurium yjeH gene also confers ceftriaxone resistance in E. coli. | [52] |
| Typhimurium | CS022 | To compare a proteome defined by shotgun proteomics directly on an LTQ-FT and by proteome pre-fractionation on an LCQ-DUO. | Shotgun proteomic analyses on the LCQ-DUO adequately characterized a PhoP constitutive strain if proteome pre-fractionation steps and gas-phase fractionation were included. | [53] |
| Typhimurium | STM14028 | To identify key proteins linked to macrophage colonization by LC-MS analysis of protein abundance in Salmonella cells isolated from RAW264.7 macrophages, with or without functional Nramp1, at various time points of infection. | After infection 39 proteins were strongly induced, 6 of which are modulated by Nramp1, including STM3117. Deletion of the STM3117 gene caused a dramatic reduction in the ability to colonize macrophages, demonstrating that STM3117 is an important virulence factor that promotes replication inside macrophages. | [54] |
| Typhimurium | SL1344 | To investigate the physiological response of S. Typhimurium to fluoroquinolone antibiotics by 2-DE and 2D-LC-MS. | Several proteins were over or underexpressed. An increase in AcrAB/TolC was associated with resistance while F1F0-ATP synthase and Imp increased in response to fluoroquinolones. | [55] |
| Typhimurium | ATCC 14028 and LT2 | To analyze the S. Typhimurium proteome under laboratory and infection-like conditions through a LC-MS-based “bottom-up” proteomic approach. | A comprehensive view of protein abundances as they vary with respect to time, environment, and genotype. Results support earlier observations that pdu gene expression contributes to S. Typhimurium pathogenesis. | [56] |
| Typhimurium and Pullorum | NCTC 74, 4 clinical isolates, A01, C01; NCTC 10704, B52 | To compare the expression patterns of host restricted S. Pullorum and host generalist S. Typhimurium isolates with a combined 2-DE LC-MS/MS proteomic approach. | Isolates varied greatly and, in some cases, more between the same serotype than between different serotypes. New serotype-specific proteins were identified, including intermediates in sulphate utilization and cysteine synthesis. | [57] |
| Typhi | clinical isolate (5866) | Analysis of the pleiotropic effects of a deficiency in the periplasmic disulfide-bond oxidoreductase DsbA using 2-DE MALDI-TOF MS. | In total, 25 spots were exclusive to the wild-type strain, 10 to the dsbA-null mutant, and 21 were common to both. DsbA, glucose-1-phosphatase, flagellin and the AI-2 autoinducer-producing LuxS were absent in the dsbA-null mutant. | [58] |
| Typhimurium | SL1344 | Proteome characterization by 2D-HPLC MS to provide a platform for subsequent proteomic studies of low level multiple antibiotic resistance. | A total of 34 OMPs were detected and 20 proteins previously associated with the mar locus in E. coli were also identified including the key MAR effectors AcrA, TolC and OmpF. | [59] |
| Typhimurium | UK1 (WT) and RJ1827 | To compare changes in gene expression caused by fis mutation through a 2-DE MS proteomic approach in order to elucidate the role of Fis in Salmonella virulence. | Identification of 11 proteins upregulated and 7 downregulated by Fis, involved in translation, sugar metabolism, flagellar synthesis, and virulence. Changes in SPI expression suggest that gene regulation in SPI-2 and in SPI-1 is affected by Fis. | [60] |
| Typhimurium | ATCC 14028 | To identify low-level expressed proteins by expressing several SPI2 T3SS putative proteins as recombinant products followed by by 2-DE MALDI-MS detection. | Recombinant expression is a complementary tool to analyze low abundant or membrane proteins. Pre-fractionation and pulse labeling allowed the identification of growth phase regulated SPI2 proteins that might not be detected otherwise. | [61] |
| Typhimurium | SL1344 | To identify acid-regulated elements of the flagellar system and to study how they are regulated by low pH. | Flagella-mediated cell motility is co-regulated by low pH via the PhoPQ signal transduction system. | [62] |
| Typhimurium | SL1344 | To test the feasibility of proteome determination by identifying 53 randomly sequenced cell envelope proteins by N-terminal sequencing of spots from 2D gels. | The existence of previously hypothetical proteins predicted from genomic sequencing projects was confirmed, and approximately 20% of the proteins had no matches in sequence databases. | [63] |
| Typhimurium | SL1344 | To present a 2D reference map for proteins of the cell envelope fraction of S. Typhimurium SL1344. | In total 49 proteins were identified by microsequencing and assigned to a 2D reference map. Of these, 10 proteins seem to be new and others closely match putative proteins or proteins found in other bacteria but not previously reported in salmonellae. | [64] |
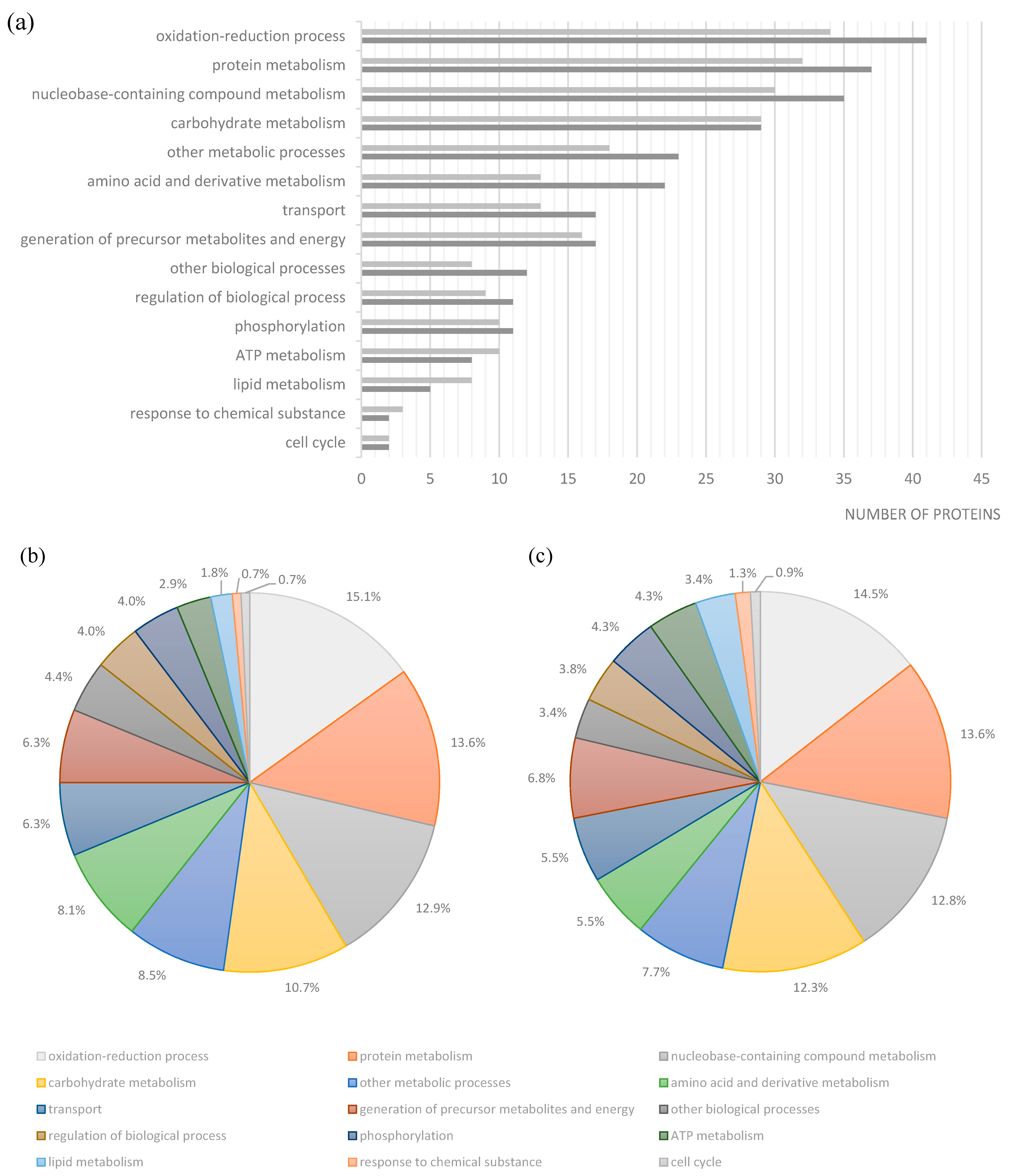
2. Results and Discussion


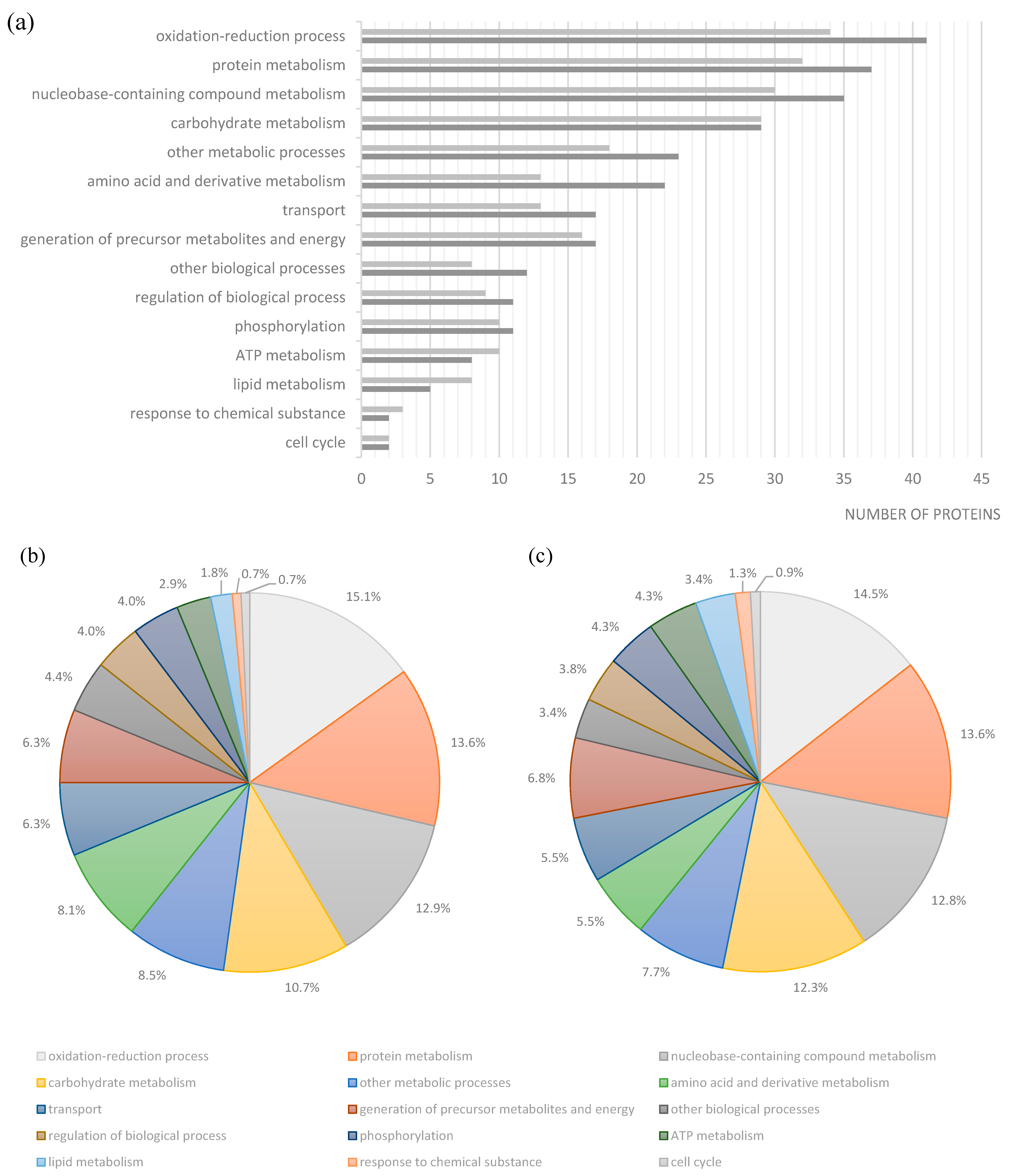
| Strain | Spots | Protein | Gene | Biological Process |
|---|---|---|---|---|
| Se20 | 24/98 | Flagellin | fljB | ciliary or bacterial-type flagellar motility |
| 75 | aminoglycoside 6'-N-acetyltransferase type Ib-cr, AAC(6')-Ib-cr4 | aac(6')-Ib-cr4 | metabolic process | |
| 99 | ethanolamine ammonia-lyase heavy subunit | eutB | cellular amino acid metabolic process | |
| 106 | ATP-dependent protease | hslU | ATP catabolic process, proteolysis, response to stress, protein unfolding | |
| 134 | universal stress protein E | uspE | response to stress | |
| 142/143 | aminoglycoside resistance protein A | strA | response to antibiotic | |
| 148 | Chain E, Alkyl Hydroperoxide Reductase C (Substrate-Ready Conformation) | ahpC | response to oxidative stress, oxidation-reduction process | |
| 182 | 5'-nucleotidase | ushA | dephosphorylation, nucleotide catabolic process | |
| SL1344 | 205/341 | arginine deiminase | arcA | protein citrullination |
| 215 | ornithine carbamoyltransferase | arcB | ornithine metabolic process | |
| 225 | fumarate reductase iron-sulfur subunit | frdB | tricarboxylic acid cycle | |
| 227 | carbamate kinase | arcC | arginine metabolic process | |
| 237/238/287 | glycerol-3-phosphate dehydrogenase | glpD | glycerol-3-phosphate metabolic process | |
| 240 | inosine 5'-monophosphate dehydrogenase | guaB | purine nucleotide biosynthetic process | |
| 259 | NADH dehydrogenase subunit G | nuoG | ATP synthesis coupled electron transport | |
| 296 | molecular chaperone DnaJ | dnaJ | response to stress | |
| 332 | Hydrogenase | - | - | |
| 344 | Phosphoglucomutase | pgm | carbohydrate metabolic process | |
| 346 | oligopeptidase A | prlC | proteolysis | |
| 378 | exonuclease III | xth | DNA catabolic process, exonucleolytic | |
| 396 | serine endoprotease | htrA | proteolysis | |
| 406 | cell invasion protein SipA | sipA | pathogenesis |
3. Experimental Section
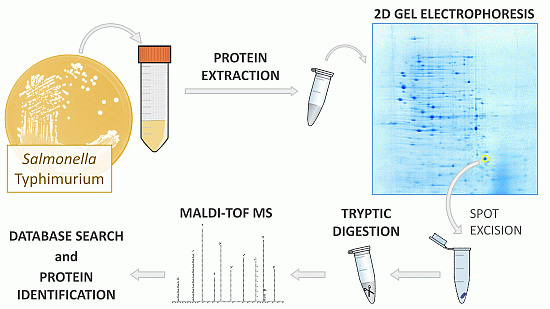
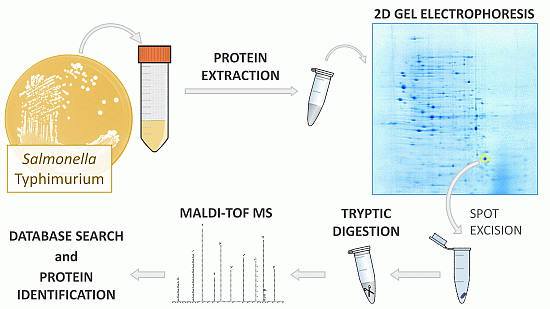
3.1. Bacterial Strains and Growth Conditions
3.2. Protein Extraction
3.3. Two-Dimensional Gel Electrophoresis
3.4. Tryptic Digestion of In-Gel Proteins
3.5. Peptide Mass Fingerprinting
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pacheco, R.; Correia, S.; Poeta, P.; Pinto, L.; Igrejas, G. The role of proteomics in elucidating multiple antibiotic resistance in Salmonella and in novel antibacterial discovery. In Salmonella—Distribution, Adaptation, Control Measures and Molecular Technologies; Annous, B.A., Gurtler, J.B., Eds.; InTech: Rijeka, Croatia, 2012; pp. 187–220. [Google Scholar]
- Su, L.H.; Wu, T.L.; Chiu, C.H. Development of carbapenem resistance during therapy for non-typhoid Salmonella infection. Clin. Microbiol. Infect. 2012, 18, E91–E94. [Google Scholar] [CrossRef]
- Mather, A.E.; Reid, S.W.; Maskell, D.J.; Parkhill, J.; Fookes, M.C.; Harris, S.R.; Brown, D.J.; Coia, J.E.; Mulvey, M.R.; Gilmour, M.W.; et al. Distinguishable epidemics of multidrug-resistant Salmonella Typhimurium DT104 in different hosts. Science 2013, 341, 1514–1517. [Google Scholar] [CrossRef]
- Marathe, S.A.; Kumar, R.; Ajitkumar, P.; Nagaraja, V.; Chakravortty, D. Curcumin reduces the antimicrobial activity of ciprofloxacin against Salmonella Typhimurium and Salmonella Typhi. J. Antimicrob. Chemother. 2013, 68, 139–152. [Google Scholar] [CrossRef]
- Bumann, D. Pathogen proteomes during infection: A basis for infection research and novel control strategies. J. Proteomics 2010, 73, 2267–2276. [Google Scholar] [CrossRef]
- Humphrey, S.; Clark, L.F.; Humphrey, T.J.; Jepson, M.A. Enhanced recovery of Salmonella Typhimurium DT104 from exposure to stress at low temperature. Microbiology 2011, 157, 1103–1114. [Google Scholar] [CrossRef]
- Fux, C.A.; Shirtliff, M.; Stoodley, P.; Costerton, J.W. Can laboratory reference strains mirror “real-world” pathogenesis? Trends Microbiol. 2005, 13, 58–63. [Google Scholar] [CrossRef]
- Threlfall, E.J. Epidemic Salmonella Typhimurium DT 104—A truly international multiresistant clone. J. Antimicrob. Chemother. 2000, 46, 7–10. [Google Scholar] [CrossRef]
- Mendoza Mdel, C.; Herrero, A.; Rodicio, M.R. Evolutionary engineering in Salmonella: Emergence of hybrid virulence-resistance plasmids in non-typhoid serotypes. Enferm. Infect. Microbiol. Clin. 2009, 27, 37–43. [Google Scholar] [CrossRef]
- Giraud, E.; Baucheron, S.; Virlogeux-Payant, I.; Nishino, K.; Cloeckaert, A. Effects of natural mutations in the ramRA locus on invasiveness of epidemic fluoroquinolone-resistant Salmonella enterica serovar Typhimurium isolates. J. Infect. Dis. 2013, 207, 794–802. [Google Scholar] [CrossRef]
- Research Topic in Antimicrobials, Resistance and Chemotherapy. Available online: http://www.frontiersin.org/antimicrobials,_resistance_and_chemotherapy/researchtopics/proteomics_of_antimicrobial_re/1620 (accessed on 28 March 2014).
- Fernandez-Reyes, M.; Rodriguez-Falcon, M.; Chiva, C.; Pachon, J.; Andreu, D.; Rivas, L. The cost of resistance to colistin in Acinetobacter baumannii: A proteomic perspective. Proteomics 2009, 9, 1632–1645. [Google Scholar] [CrossRef]
- Becker, D.; Selbach, M.; Rollenhagen, C.; Ballmaier, M.; Meyer, T.F.; Mann, M.; Bumann, D. Robust Salmonella metabolism limits possibilities for new antimicrobials. Nature 2006, 440, 303–307. [Google Scholar] [CrossRef]
- Lima, T.B.; Pinto, M.F.; Ribeiro, S.M.; de Lima, L.A.; Viana, J.C.; Gomes Junior, N.; Candido Ede, S.; Dias, S.C.; Franco, O.L. Bacterial resistance mechanism: What proteomics can elucidate. FASEB J. 2013, 27, 1291–1303. [Google Scholar] [CrossRef]
- Kleinert, P.; Kuster, T.; Arnold, D.; Jaeken, J.; Heizmann, C.W.; Troxler, H. Effect of glycosylation on the protein pattern in 2-d-gel electrophoresis. Proteomics 2007, 7, 15–22. [Google Scholar] [CrossRef]
- Lee, A.Y.; Park, S.G.; Jang, M.; Cho, S.; Myung, P.K.; Kim, Y.R.; Rhee, J.H.; Lee, D.H.; Park, B.C. Proteomic analysis of pathogenic bacterium Vibrio vulnificus. Proteomics 2006, 6, 1283–1289. [Google Scholar] [CrossRef]
- Vranakis, I.; Goniotakis, I.; Psaroulaki, A.; Sandalakis, V.; Tselentis, Y.; Gevaert, K.; Tsiotis, G. Proteome studies of bacterial antibiotic resistance mechanisms. J. Proteomics 2014, 97, 88–99. [Google Scholar] [CrossRef]
- De Toro, M.; Rojo-Bezares, B.; Vinue, L.; Undabeitia, E.; Torres, C.; Saenz, Y. In vivo selection of aac(6')-Ib-cr and mutations in the gyrA gene in a clinical qnrS1-positive Salmonella enterica serovar Typhimurium DT104B strain recovered after fluoroquinolone treatment. J. Antimicrob. Chemother. 2010, 65, 1945–1949. [Google Scholar] [CrossRef]
- Hoiseth, S.K.; Stocker, B.A. Aromatic-dependent Salmonella Typhimurium are non-virulent and effective as live vaccines. Nature 1981, 291, 238–239. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, K.Y.; Huo, Y. Proteomic comparison between Salmonella Typhimurium and Salmonella Typhi. J. Microbiol. 2014, 52, 71–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Nandakumar, R.; Bartelt-Hunt, S.L.; Snow, D.D.; Hodges, L.; Li, X. Quantitative proteomic analysis of the Salmonella-lettuce interaction. Microb. Biotechnol. 2014. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, D.; Pang, B.; Zhang, Q.; Zhou, H.; Zhang, L.; Zhang, J.; Kan, B. The core proteome and pan proteome of Salmonella Paratyphi A epidemic strains. PLoS One 2014, 9, e89197. [Google Scholar]
- Ansong, C.; Wu, S.; Meng, D.; Liu, X.; Brewer, H.M.; Deatherage Kaiser, B.L.; Nakayasu, E.S.; Cort, J.R.; Pevzner, P.; Smith, R.D.; et al. Top-down proteomics reveals a unique protein S-thiolation switch in Salmonella Typhimurium in response to infection-like conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 10153–10158. [Google Scholar] [CrossRef]
- Brown, R.N.; Sanford, J.A.; Park, J.H.; Deatherage, B.L.; Champion, B.L.; Smith, R.D.; Heffron, F.; Adkins, J.N. A Comprehensive Subcellular Proteomic Survey of Salmonella Grown under Phagosome-Mimicking versus Standard Laboratory Conditions. Int. J. Proteomics 2012, 2012, 123076. [Google Scholar]
- Condell, O.; Sheridan, A.; Power, K.A.; Bonilla-Santiago, R.; Sergeant, K.; Renaut, J.; Burgess, C.; Fanning, S.; Nally, J.E. Comparative proteomic analysis of Salmonella tolerance to the biocide active agent triclosan. J. Proteomics 2012, 75, 4505–4519. [Google Scholar] [CrossRef]
- Cooper, B.; Chen, R.; Garrett, W.M.; Murphy, C.; Chang, C.; Tucker, M.L.; Bhagwat, A.A. Proteomic pleiotropy of OpgGH, an operon necessary for efficient growth of Salmonella enterica serovar typhimurium under low-osmotic conditions. J. Proteome Res. 2012, 11, 1720–1727. [Google Scholar] [CrossRef]
- Feng, Y.; Chien, K.Y.; Chen, H.L.; Chiu, C.H. Pseudogene recoding revealed from proteomic analysis of Salmonella serovars. J. Proteome Res. 2012, 11, 1715–1719. [Google Scholar] [CrossRef]
- Kang, M.S.; Kwon, Y.K.; Kim, H.R.; Oh, J.Y.; Kim, M.J.; An, B.K.; Shin, E.G.; Kwon, J.H.; Park, C.K. Comparative proteome and transcriptome analyses of wild-type and live vaccine strains of Salmonella enterica serovar Gallinarum. Vaccine 2012, 30, 6368–6375. [Google Scholar]
- Sun, J.S.; Hahn, T.W. Comparative proteomic analysis of Salmonella enterica serovars Enteritidis, Typhimurium and Gallinarum. J. Vet. Med. Sci./Jpn. Soc. Vet. Sci. 2012, 74, 285–291. [Google Scholar]
- Ciavardelli, D.; Ammendola, S.; Ronci, M.; Consalvo, A.; Marzano, V.; Lipoma, M.; Sacchetta, P.; Federici, G.; di Ilio, C.; Battistoni, A.; et al. Phenotypic profile linked to inhibition of the major Zn influx system in Salmonella enterica: Proteomics and ionomics investigations. Mol. Biosyst. 2011, 7, 608–619. [Google Scholar] [CrossRef]
- Chen, J.; Wei, D.; Zhuang, H.; Qiao, Y.; Tang, B.; Zhang, X.; Wei, J.; Fang, S.; Chen, G.; Du, P.; et al. Proteomic screening of anaerobically regulated promoters from Salmonella and its antitumor applications. Mol. Cell. Proteomics: MCP 2011, 10, M111 009399. [Google Scholar] [CrossRef]
- Niemann, G.S.; Brown, R.N.; Gustin, J.K.; Stufkens, A.; Shaikh-Kidwai, A.S.; Li, J.; McDermott, J.E.; Brewer, H.M.; Schepmoes, A.; Smith, R.D.; et al. Discovery of novel secreted virulence factors from Salmonella enterica serovar Typhimurium by proteomic analysis of culture supernatants. Infect. Immun. 2011, 79, 33–43. [Google Scholar] [CrossRef]
- Paradela, A.; Mariscotti, J.F.; Navajas, R.; Ramos-Fernandez, A.; Albar, J.P.; Garcia-del Portillo, F. Inverse regulation in the metabolic genes pckA and metE revealed by proteomic analysis of the Salmonella RcsCDB regulon. J. Proteome Res. 2011, 10, 3386–3398. [Google Scholar] [CrossRef]
- Yu, J.L.; Guo, L. Quantitative proteomic analysis of Salmonella enterica serovar Typhimurium under PhoP/PhoQ activation conditions. J. Proteome Res. 2011, 10, 2992–3002. [Google Scholar] [CrossRef]
- Charles, R.C.; Sheikh, A.; Krastins, B.; Harris, J.B.; Bhuiyan, M.S.; LaRocque, R.C.; Logvinenko, T.; Sarracino, D.A.; Kudva, I.T.; Eisenstein, J.; et al. Characterization of anti-Salmonella enterica serotype Typhi antibody responses in bacteremic Bangladeshi patients by an immunoaffinity proteomics-based technology. Clin. Vaccine Immunol.: CVI 2010, 17, 1188–1195. [Google Scholar] [CrossRef]
- Beraud, M.; Kolb, A.; Monteil, V.; D’Alayer, J.; Norel, F. A proteomic analysis reveals differential regulation of the σS-dependent yciGFE(katN) locus by YncC and H-NS in Salmonella and Escherichia coli K-12. Mol. Cell. Proteomics: MCP 2010, 9, 2601–2616. [Google Scholar] [CrossRef]
- Calhoun, L.N.; Liyanage, R.; Lay, J.O., Jr.; Kwon, Y.M. Proteomic analysis of Salmonella enterica serovar Enteritidis following propionate adaptation. BMC Microbiol. 2010, 10, 249. [Google Scholar] [CrossRef]
- Kim, K.; Yang, E.; Vu, G.P.; Gong, H.; Su, J.; Liu, F.; Lu, S. Mass spectrometry-based quantitative proteomic analysis of Salmonella enterica serovar Enteritidis protein expression upon exposure to hydrogen peroxide. BMC Microbiol. 2010, 10, 166. [Google Scholar] [CrossRef]
- Pinto, L.; Poeta, P.; Vieira, S.; Caleja, C.; Radhouani, H.; Carvalho, C.; Vieira-Pinto, M.; Themudo, P.; Torres, C.; Vitorino, R.; et al. Genomic and proteomic evaluation of antibiotic resistance in Salmonella strains. J. Proteomics 2010, 73, 1535–1541. [Google Scholar] [CrossRef]
- Chooneea, D.; Karlsson, R.; Encheva, V.; Arnold, C.; Appleton, H.; Shah, H. Elucidation of the outer membrane proteome of Salmonella enterica serovar Typhimurium utilising a lipid-based protein immobilization technique. BMC Microbiol. 2010, 10, 44. [Google Scholar] [CrossRef]
- Di Pasqua, R.; Mamone, G.; Ferranti, P.; Ercolini, D.; Mauriello, G. Changes in the proteome of Salmonella enterica serovar Thompson as stress adaptation to sublethal concentrations of thymol. Proteomics 2010, 10, 1040–1049. [Google Scholar]
- Charles, R.C.; Harris, J.B.; Chase, M.R.; Lebrun, L.M.; Sheikh, A.; LaRocque, R.C.; Logvinenko, T.; Rollins, S.M.; Tarique, A.; Hohmann, E.L.; et al. Comparative proteomic analysis of the PhoP regulon in Salmonella enterica serovar Typhi versus Typhimurium. PLoS One 2009, 4, e6994. [Google Scholar]
- Encheva, V.; Shah, H.N.; Gharbia, S.E. Proteomic analysis of the adaptive response of Salmonella enterica serovar Typhimurium to growth under anaerobic conditions. Microbiology 2009, 155, 2429–2441. [Google Scholar] [CrossRef]
- Kim, S.R.; Kim, H.T.; Park, H.J.; Kim, S.; Choi, H.J.; Hwang, G.S.; Yi, J.H.; Ryu do, H.; Kim, K.H. Fatty acid profiling and proteomic analysis of Salmonella enterica serotype Typhimurium inactivated with supercritical carbon dioxide. Int. J. Food Microbiol. 2009, 134, 190–195. [Google Scholar] [CrossRef]
- Osman, K.M.; Ali, M.M.; Radwan, M.I.; Kim, H.K.; Han, J. Comparative proteomic analysis on Salmonella Gallinarum and Salmonella Enteritidis exploring proteins that may incorporate host adaptation in poultry. J. Proteomics 2009, 72, 815–821. [Google Scholar] [CrossRef]
- Shi, L.; Chowdhury, S.M.; Smallwood, H.S.; Yoon, H.; Mottaz-Brewer, H.M.; Norbeck, A.D.; McDermott, J.E.; Clauss, T.R.; Heffron, F.; Smith, R.D.; et al. Proteomic investigation of the time course responses of RAW 264.7 macrophages to infection with Salmonella enterica. Infect. Immun. 2009, 77, 3227–3333. [Google Scholar]
- Shi, L.; Ansong, C.; Smallwood, H.; Rommereim, L.; McDermott, J.E.; Brewer, H.M.; Norbeck, A.D.; Taylor, R.C.; Gustin, J.K.; Heffron, F.; et al. Proteome of Salmonella enterica serotype Typhimurium grown in a Low Mg/pH medium. J. Proteomics Bioinform. 2009, 2, 388–397. [Google Scholar] [CrossRef]
- Kint, G.; Sonck, K.A.; Schoofs, G.; de Coster, D.; Vanderleyden, J.; de Keersmaecker, S.C. 2D proteome analysis initiates new insights on the Salmonella Typhimurium LuxS protein. BMC Microbiol. 2009, 9, 198. [Google Scholar] [CrossRef]
- Sonck, K.A.; Kint, G.; Schoofs, G.; Vander Wauven, C.; Vanderleyden, J.; de Keersmaecker, S.C. The proteome of Salmonella Typhimurium grown under in vivo-mimicking conditions. Proteomics 2009, 9, 565–579. [Google Scholar] [CrossRef]
- Webber, M.A.; Coldham, N.G.; Woodward, M.J.; Piddock, L.J. Proteomic analysis of triclosan resistance in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2008, 62, 92–97. [Google Scholar] [CrossRef]
- Singh, R.; Shasany, A.K.; Aggarwal, A.; Sinha, S.; Sisodia, B.S.; Khanuja, S.P.; Misra, R. Low molecular weight proteins of outer membrane of Salmonella Typhimurium are immunogenic in Salmonella induced reactive arthritis revealed by proteomics. Clin. Exp. Immunol. 2007, 148, 486–493. [Google Scholar] [CrossRef]
- Hu, W.S.; Lin, Y.H.; Shih, C.C. A proteomic approach to study Salmonella enterica serovar Typhimurium putative transporter YjeH associated with ceftriaxone resistance. Biochem. Biophys. Res. Commun. 2007, 361, 694–699. [Google Scholar] [CrossRef]
- Nunn, B.L.; Shaffer, S.A.; Scherl, A.; Gallis, B.; Wu, M.; Miller, S.I.; Goodlett, D.R. Comparison of a Salmonella Typhimurium proteome defined by shotgun proteomics directly on an LTQ-FT and by proteome pre-fractionation on an LCQ-DUO. Brief. Funct. Genomics Proteomics 2006, 5, 154–168. [Google Scholar] [CrossRef]
- Shi, L.; Adkins, J.N.; Coleman, J.R.; Schepmoes, A.A.; Dohnkova, A.; Mottaz, H.M.; Norbeck, A.D.; Purvine, S.O.; Manes, N.P.; Smallwood, H.S.; et al. Proteomic analysis of Salmonella enterica serovar Typhimurium isolated from RAW 264.7 macrophages: Identification of a novel protein that contributes to the replication of serovar Typhimurium inside macrophages. J. Biol. Chem. 2006, 281, 29131–29140. [Google Scholar] [CrossRef]
- Coldham, N.G.; Randall, L.P.; Piddock, L.J.; Woodward, M.J. Effect of fluoroquinolone exposure on the proteome of Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2006, 58, 1145–1153. [Google Scholar] [CrossRef]
- Adkins, J.N.; Mottaz, H.M.; Norbeck, A.D.; Gustin, J.K.; Rue, J.; Clauss, T.R.; Purvine, S.O.; Rodland, K.D.; Heffron, F.; Smith, R.D. Analysis of the Salmonella Typhimurium proteome through environmental response toward infectious conditions. Mol. Cell. Proteomics: MCP 2006, 5, 1450–1461. [Google Scholar] [CrossRef]
- Encheva, V.; Wait, R.; Gharbia, S.E.; Begum, S.; Shah, H.N. Proteome analysis of serovars Typhimurium and Pullorum of Salmonella enterica subspecies I. BMC Microbiol. 2005, 5, 42. [Google Scholar] [CrossRef]
- Agudo, D.; Mendoza, M.T.; Castanares, C.; Nombela, C.; Rotger, R. A proteomic approach to study Salmonella typhi periplasmic proteins altered by a lack of the DsbA thiol: Disulfide isomerase. Proteomics 2004, 4, 355–363. [Google Scholar] [CrossRef]
- Coldham, N.G.; Woodward, M.J. Characterization of the Salmonella Typhimurium proteome by semi-automated two dimensional HPLC-mass spectrometry: Detection of proteins implicated in multiple antibiotic resistance. J. Proteome Res. 2004, 3, 595–603. [Google Scholar] [CrossRef]
- Yoon, H.; Lim, S.; Heu, S.; Choi, S.; Ryu, S. Proteome analysis of Salmonella enterica serovar Typhimurium fis mutant. FEMS Microbiol. Lett. 2003, 226, 391–396. [Google Scholar] [CrossRef]
- Deiwick, J.; Rappl, C.; Stender, S.; Jungblut, P.R.; Hensel, M. Proteomic approaches to Salmonella Pathogenicity Island 2 encoded proteins and the SsrAB regulon. Proteomics 2002, 2, 792–799. [Google Scholar] [CrossRef]
- Adams, P.; Fowler, R.; Kinsella, N.; Howell, G.; Farris, M.; Coote, P.; O’Connor, C.D. Proteomic detection of PhoPQ- and acid-mediated repression of Salmonella motility. Proteomics 2001, 1, 597–607. [Google Scholar] [CrossRef]
- O’Connor, C.D.; Farris, M.; Fowler, R.; Qi, S.Y. The proteome of Salmonella enterica serovar Typhimurium: Current progress on its determination and some applications. Electrophoresis 1997, 18, 1483–1490. [Google Scholar] [CrossRef]
- Qi, S.Y.; Moir, A.; O’Connor, C.D. Proteome of Salmonella Typhimurium SL1344: Identification of novel abundant cell envelope proteins and assignment to a two-dimensional reference map. J. Bacteriol. 1996, 178, 5032–5038. [Google Scholar]
- Hogg, G.; Dimovski, K.; Hiley, L.; Holt, K.E. Draft Genome Sequences for Ten Salmonella enterica Serovar Typhimurium Phage Type 135 Variants. Genome Announc. 2013, 1. [Google Scholar] [CrossRef]
- De Toro, M.; Rodriguez, I.; Rojo-Bezares, B.; Helmuth, R.; Torres, C.; Guerra, B.; Saenz, Y. pMdT1, a small ColE1-like plasmid mobilizing a new variant of the aac(6')-Ib-cr gene in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2013, 68, 1277–1280. [Google Scholar] [CrossRef]
- Rankin, J.D.; Taylor, R.J. The estimation of doses of Salmonella Typhimurium suitable for the experimental production of disease in calves. Vet. Rec. 1966, 78, 706–777. [Google Scholar]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; Macpherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, 2177–2189. [Google Scholar]
- Supek, F.; Bosnjak, M.; Skunca, N.; Smuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS One 2011, 6, e21800. [Google Scholar]
- Poirel, L.; Cattoir, V.; Nordmann, P. Plasmid-Mediated Quinolone Resistance; Interactions between Human, Animal, and Environmental Ecologies. Front. Microbiol. 2012, 3, 24. [Google Scholar]
- Cattoir, V.; Nordmann, P. Plasmid-mediated quinolone resistance in gram-negative bacterial species: An update. Curr. Med. Chem. 2009, 16, 1028–1046. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Zheng, C.; Chitsaz, F.; Derbyshire, M.K.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Lanczycki, C.J.; et al. CDD: Conserved domains and protein three-dimensional structure. Nucleic Acids Res. 2013, 41, D348–D352. [Google Scholar] [CrossRef]
- Righetti, P.G. Real and imaginary artefacts in proteome analysis via two-dimensional maps. J. Chromatogr. B, Anal. Technol. Biomed. Life Sci. 2006, 841, 14–22. [Google Scholar] [CrossRef]
- Skold, O. Sulfonamide resistance: Mechanisms and trends. Drug Resist. Updates 2000, 3, 155–160. [Google Scholar] [CrossRef]
- Kroger, C.; Dillon, S.C.; Cameron, A.D.; Papenfort, K.; Sivasankaran, S.K.; Hokamp, K.; Chao, Y.; Sittka, A.; Hebrard, M.; Handler, K.; et al. The transcriptional landscape and small RNAs of Salmonella enterica serovar Typhimurium. Proc. Natl. Acad. Sci. USA 2012, 109, E1277–E1286. [Google Scholar] [CrossRef]
- Lal, A.; Vela, J.; Yee, J.; Yuan, E. Rescuing the ompA Deletion Mutant Escherichia coli JW0940 by Reintroducing ompA in the TOPO Cloning Vector pBAD. J. Exp. Microbiol. Immunol. 2012, 16, 108–112. [Google Scholar]
- Kleinschmidt, J.H. Folding kinetics of the outer membrane proteins OmpA and FomA into phospholipid bilayers. Chem. Phys. Lipids 2006, 141, 30–47. [Google Scholar]
- Mahendran, K.R.; Kreir, M.; Weingart, H.; Fertig, N.; Winterhalter, M. Permeation of antibiotics through Escherichia coli OmpF and OmpC porins: Screening for influx on a single-molecule level. J. Biomol. Screen. 2010, 15, 302–307. [Google Scholar] [CrossRef]
- Paterson, G.K.; Northen, H.; Cone, D.B.; Willers, C.; Peters, S.E.; Maskell, D.J. Deletion of tolA in Salmonella Typhimurium generates an attenuated strain with vaccine potential. Microbiology 2009, 155, 220–228. [Google Scholar] [CrossRef]
- Nirudodhi, S.; Parsonage, D.; Karplus, P.A.; Poole, L.B.; Maier, C.S. Conformational studies of the robust 2-Cys peroxiredoxin Salmonella Typhimurium AhpC by solution phase hydrogen/deuterium (H/D) exchange monitored by electrospray ionization mass spectrometry. Int. J. Mass Spectrom. 2011, 302, 93–100. [Google Scholar]
- Guerrero, P.; Collao, B.; Alvarez, R.; Salinas, H.; Morales, E.H.; Calderon, I.L.; Saavedra, C.P.; Gil, F. Salmonella enterica serovar Typhimurium BaeSR two-component system positively regulates sodA in response to ciprofloxacin. Microbiology 2013, 159, 2049–2057. [Google Scholar] [CrossRef]
- Bakshi, C.S.; Malik, M.; Regan, K.; Melendez, J.A.; Metzger, D.W.; Pavlov, V.M.; Sellati, T.J. Superoxide dismutase B gene (sodB)-deficient mutants of Francisella tularensis demonstrate hypersensitivity to oxidative stress and attenuated virulence. J. Bacteriol. 2006, 188, 6443–6448. [Google Scholar] [CrossRef]
- Troxell, B.; Fink, R.C.; Porwollik, S.; McClelland, M.; Hassan, H.M. The Fur regulon in anaerobically grown Salmonella enterica sv. Typhimurium: Identification of new Fur targets. BMC Microbiol. 2011, 11, 236. [Google Scholar] [CrossRef]
- Yang, X.; Thornburg, T.; Suo, Z.; Jun, S.; Robison, A.; Li, J.; Lim, T.; Cao, L.; Hoyt, T.; Avci, R.; et al. Flagella overexpression attenuates Salmonella pathogenesis. PLoS One 2012, 7, e46828. [Google Scholar]
- Garsin, D.A. Ethanolamine utilization in bacterial pathogens: Roles and regulation. Nat. Rev. Microbiol. 2010, 8, 290–295. [Google Scholar] [CrossRef]
- Thiennimitr, P.; Winter, S.E.; Winter, M.G.; Xavier, M.N.; Tolstikov, V.; Huseby, D.L.; Sterzenbach, T.; Tsolis, R.M.; Roth, J.R.; Baumler, A.J. Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 17480–17485. [Google Scholar]
- Rittmann, D.; Sorger-Herrmann, U.; Wendisch, V.F. Phosphate starvation-inducible gene ushA encodes a 5' nucleotidase required for growth of Corynebacterium glutamicum on media with nucleotides as the phosphorus source. Appl. Environ. Microbiol. 2005, 71, 4339–4444. [Google Scholar]
- Choi, Y.; Choi, J.; Groisman, E.A.; Kang, D.H.; Shin, D.; Ryu, S. Expression of STM4467-encoded arginine deiminase controlled by the STM4463 regulator contributes to Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2012, 80, 4291–4297. [Google Scholar] [CrossRef]
- Cohen-Ben-Lulu, G.N.; Francis, N.R.; Shimoni, E.; Noy, D.; Davidov, Y.; Prasad, K.; Sagi, Y.; Cecchini, G.; Johnstone, R.M.; Eisenbach, M. The bacterial flagellar switch complex is getting more complex. EMBO J. 2008, 27, 1134–1144. [Google Scholar] [CrossRef]
- Liechti, G.; Goldberg, J.B. Helicobacter pylori relies primarily on the purine salvage pathway for purine nucleotide biosynthesis. J. Bacteriol. 2012, 194, 839–854. [Google Scholar] [CrossRef]
- Suvarnapunya, A.E.; Lagasse, H.A.; Stein, M.A. The role of DNA base excision repair in the pathogenesis of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2003, 48, 549–559. [Google Scholar] [CrossRef]
- Turner, A.K.; Barber, L.Z.; Wigley, P.; Muhammad, S.; Jones, M.A.; Lovell, M.A.; Hulme, S.; Barrow, P.A. Contribution of proton-translocating proteins to the virulence of Salmonella enterica serovars Typhimurium, Gallinarum, and Dublin in chickens and mice. Infect. Immun. 2003, 71, 3392–3401. [Google Scholar] [CrossRef]
- Maier, R.J.; Olczak, A.; Maier, S.; Soni, S.; Gunn, J. Respiratory hydrogen use by Salmonella enterica serovar Typhimurium is essential for virulence. Infect. Immun. 2004, 72, 6294–6299. [Google Scholar] [CrossRef]
- Paterson, G.K.; Cone, D.B.; Peters, S.E.; Maskell, D.J. The enzyme phosphoglucomutase (Pgm) is required by Salmonella enterica serovar Typhimurium for O-antigen production, resistance to antimicrobial peptides and in vivo fitness. Microbiology 2009, 155, 3403–3410. [Google Scholar] [CrossRef]
- Calhoun, L.N.; Kim, J.N.; Ren, Y.; Song, J.J.; Kwon, Y.M. The DNA-binding protein dps functions as a global regulator in starved Salmonella enterica serovar enteritidis during starvation. Int. J. Microbiol. Res. 2011, 3, 136–147. [Google Scholar]
- Raffatellu, M.; Wilson, R.P.; Chessa, D.; Andrews-Polymenis, H.; Tran, Q.T.; Lawhon, S.; Khare, S.; Adams, L.G.; Baumler, A.J. SipA, SopA, SopB, SopD, and SopE2 contribute to Salmonella enterica serotype Typhimurium invasion of epithelial cells. Infect. Immun. 2005, 73, 146–154. [Google Scholar] [CrossRef]
- Slattery, A.; Victorsen, A.H.; Brown, A.; Hillman, K.; Phillips, G.J. Isolation of highly persistent mutants of Salmonella enterica serovar Typhimurium reveals a new toxin-antitoxin module. J. Bacteriol. 2013, 195, 647–657. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar]
- Görg, A.; Klaus, A.; Lück, C.; Weiland, F.; Weiss, W. Two-Dimensional Electrophoresis with Immobilized pH Gradients for Proteome Analysis: A laboratory manual; Technical University of Munich: Freising-Weihenstephan, Germany, 2007; pp. 1–119. [Google Scholar]
- Gorg, A.; Obermaier, C.; Boguth, G.; Harder, A.; Scheibe, B.; Wildgruber, R.; Weiss, W. The current state of two-dimensional electrophoresis with immobilized pH gradients. Electrophoresis 2000, 21, 1037–1053. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Gorg, A.; Weiss, W.; Dunn, M.J. Current two-dimensional electrophoresis technology for proteomics. Proteomics 2004, 4, 3665–3685. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Correia, S.; Nunes-Miranda, J.D.; Pinto, L.; Santos, H.M.; De Toro, M.; Sáenz, Y.; Torres, C.; Capelo, J.L.; Poeta, P.; Igrejas, G. Complete Proteome of a Quinolone-Resistant Salmonella Typhimurium Phage Type DT104B Clinical Strain. Int. J. Mol. Sci. 2014, 15, 14191-14219. https://doi.org/10.3390/ijms150814191
Correia S, Nunes-Miranda JD, Pinto L, Santos HM, De Toro M, Sáenz Y, Torres C, Capelo JL, Poeta P, Igrejas G. Complete Proteome of a Quinolone-Resistant Salmonella Typhimurium Phage Type DT104B Clinical Strain. International Journal of Molecular Sciences. 2014; 15(8):14191-14219. https://doi.org/10.3390/ijms150814191
Chicago/Turabian StyleCorreia, Susana, Júlio D. Nunes-Miranda, Luís Pinto, Hugo M. Santos, María De Toro, Yolanda Sáenz, Carmen Torres, José Luis Capelo, Patrícia Poeta, and Gilberto Igrejas. 2014. "Complete Proteome of a Quinolone-Resistant Salmonella Typhimurium Phage Type DT104B Clinical Strain" International Journal of Molecular Sciences 15, no. 8: 14191-14219. https://doi.org/10.3390/ijms150814191
APA StyleCorreia, S., Nunes-Miranda, J. D., Pinto, L., Santos, H. M., De Toro, M., Sáenz, Y., Torres, C., Capelo, J. L., Poeta, P., & Igrejas, G. (2014). Complete Proteome of a Quinolone-Resistant Salmonella Typhimurium Phage Type DT104B Clinical Strain. International Journal of Molecular Sciences, 15(8), 14191-14219. https://doi.org/10.3390/ijms150814191