1. Introduction
Half a century ago, Lerner
et al. [
1,
2] isolated from thousands of bovine pineal glands,
N-acetyl-5-methoxytryptamine or melatonin. Its name comes from the Greek
melano = black and
tonos = color, due to its lightening effect on frog skin. The main action of this molecule, at least in vertebrates, is as a signal of the photoperiod length, which allows organisms to better adapt to and also to anticipate periodic environmental changes [
3,
4,
5]. This indoleamine is present in a variety of organisms, from unicellular algae to mammals, and its production follows a circadian rhythm [
6,
7]. Melatonin has also been identified in crustaceans, although its production and functions are not as well understood as they are in vertebrates [
8,
9]. Maciel and colleagues [
10] reported that when the gills of the crab
Neohelice granulata were incubated with melatonin (20 × 10
−9 M), oxygen consumption decreased. Tilden
et al. [
11] observed a shift in the circadian rhythm of glucose and lactate in the hemolymph of melatonin-injected
Uca pugilator (3 × 10
−9 mol·animal
−1), with one peak in the middle of the light phase. In another study, Sainath and Reddy [
12] demonstrated a dose-dependent effect of melatonin on hyperglycemia in the crab
Oziotelphusa senex senex. These reports suggest that this indoleamine may have a role in metabolic regulation in crustaceans.
Since glucose is one of the main energy substrates for the general metabolic processes in crustaceans [
13], several studies have analyzed the circulating glucose levels in relation to various environmental stresses imposed on the animals, such as temperature [
14,
15,
16,
17,
18], salinity [
19,
20] and hypoxia [
21,
22,
23,
24,
25]. The general response to these stressors is hyperglycemia, evoked by the crustacean hyperglycemic hormone (CHH). This hormone is a peptide that has been identified mainly in the neuroendocrine system localized in the optic ganglia of eyestalk (x-organ/sinus gland complex), but also in other sites such as cerebral ganglia, ventral nerve cord, gill, hindgut and others (for review see [
26]). The CHH nucleotide gene sequence was determined in a variety of crustacean species, and its main action is the modulation of hemolymth glucose levels through carbohydrate mobilization [
26]. However, the control of synthesis and release of this peptide are not completely understood.
The estuarine crab
Neohelice granulata has a high tolerance to hypoxia and anoxia, conditions related to decreased dissolved-oxygen content in the water or to air exposure [
27,
28], common situations in salt marshes, their natural habitat [
29]. Specific metabolic adjustments, such as decreasing locomotor activity and oxygen consumption (VO
2), and increasing glucose and lactate levels in the hemolymph, allow this crab to tolerate these stressors [
21,
28,
30,
31,
32]. Maciel
et al. [
25] found that severe hypoxia (45 min at 0.7 mgO
2·L
−1) induced a sharp decrease of VO
2 and an increase of circulating lactate in both intact and eyestalkless crabs, but hyperglycemia only in intact ones, indicating a dependence on eyestalk CHH for this response. As some of the above-mentioned studies indicate that melatonin is involved in the metabolic regulation of crustaceans, the present study evaluated whether melatonin is capable of inducing metabolic changes similar to those that occur during hypoxia, and if these responses are eyestalk- and melatonin receptor-dependent.
2. Results
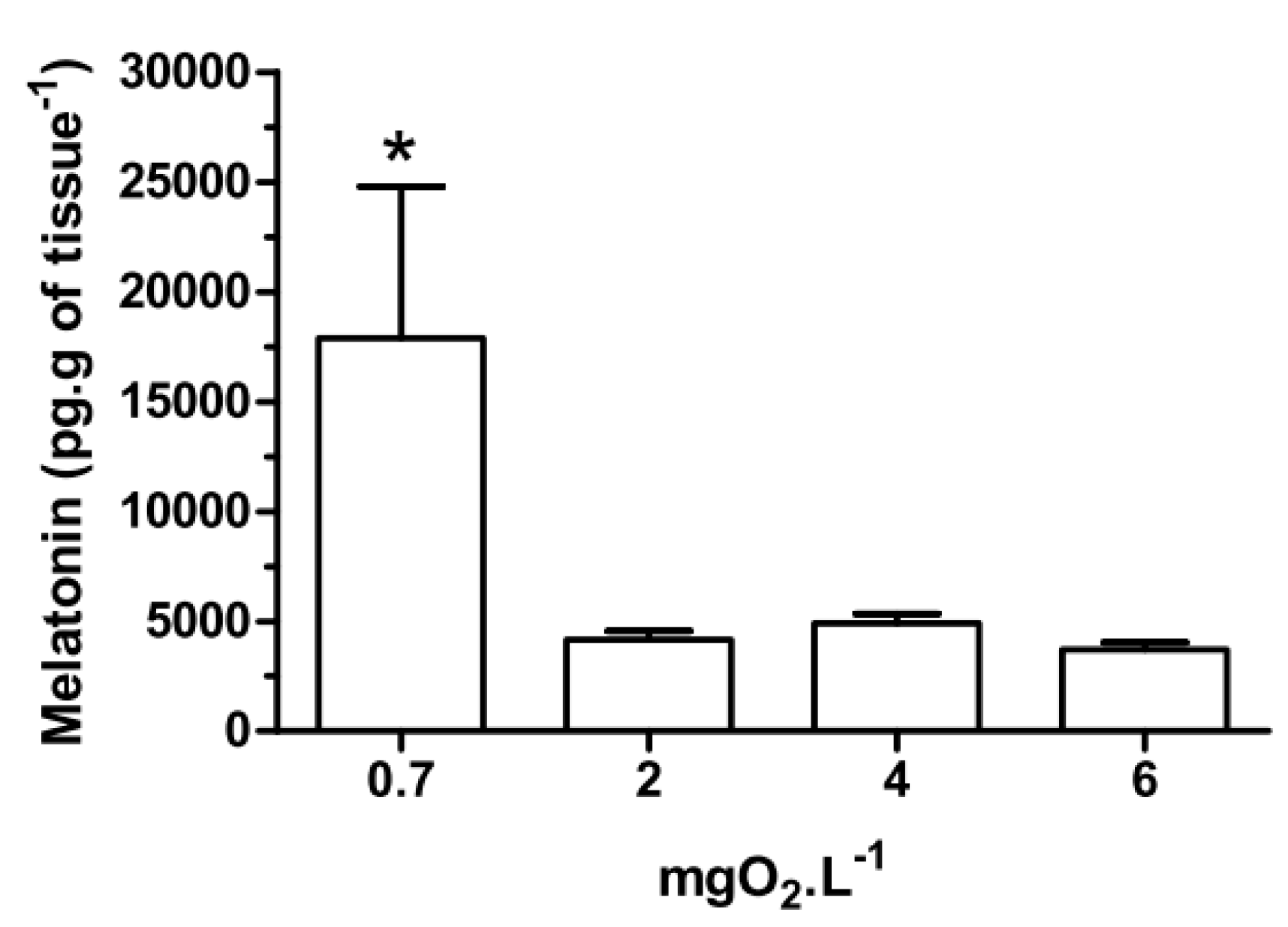
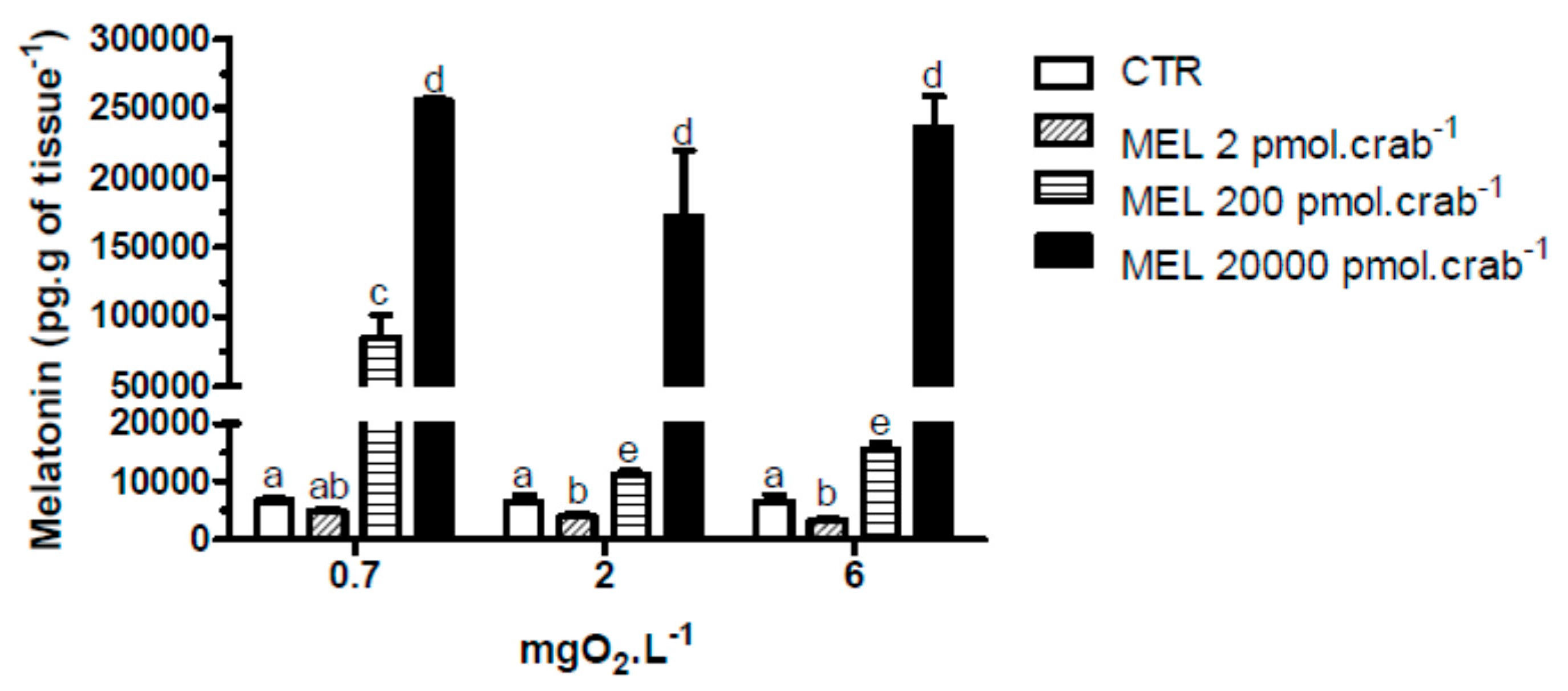
With respect to melatonin levels, eyestalk ganglia incubated for 45 min at 0.7 mgO
2·L
−1 (
Figure 1), showed a significant (
p < 0.05) increase (17,886 ± 6908 pg·g of tissue
−1) compared to 2, 4 and 6 mgO
2·L
−1 (4165 ± 397, 4907 ± 428 and 3709 ± 334 pg·g of tissue
−1, respectively). Interestingly, in the
in vivo experiment, crabs treated with 20,000 pmol·crab
−1 had a higher eyestalk melatonin content (
p < 0.05) in all three levels of oxygen incubation (255,206 ± 2129 pg·g of tissue
−1 for 0.7 mgO
2·L
−1; 171,409 ± 37,423 pg·g of tissue
−1 for 2 mgO
2·L
−1; and 235,900 ± 20,693 pg·g of tissue
−1 for 6 mgO
2·L
−1) compared to the other treatment groups. Crabs treated with 200 pmol·crab
−1 and exposed for 45 min at 0.7 mgO
2·L
−1 showed a higher melatonin content in the eyestalks (84,831 ± 16,511 pg·g of tissue
−1) compared with crabs exposed to 2 (11,273 ± 489 pg·g of tissue
−1) and 6 (15,429 ± 1258 pg·g of tissue
−1) mgO
2·L
−1 under the same treatment, and also compared to crabs treated with 2 pmol·crab
−1 and physiological saline (see
Figure 2).
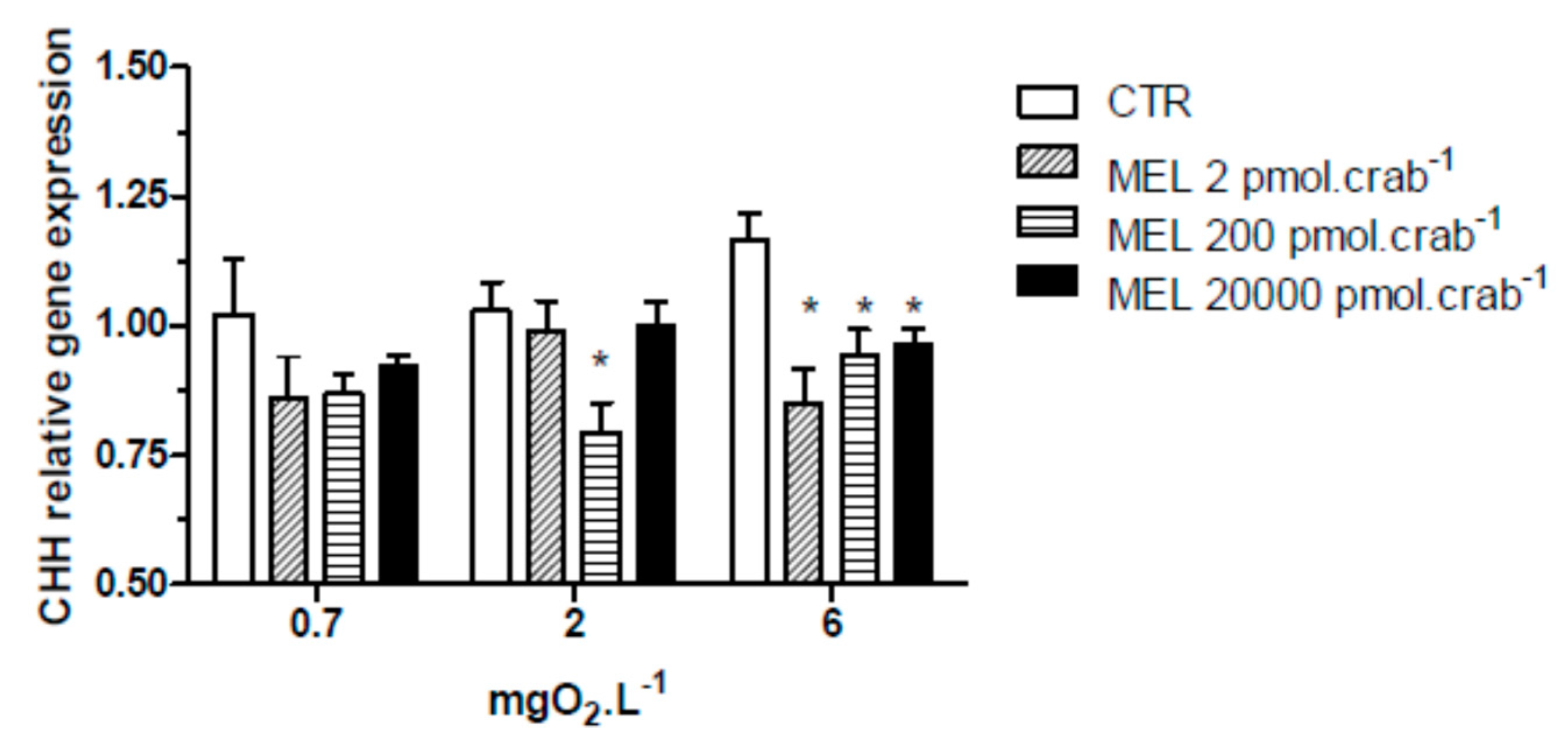
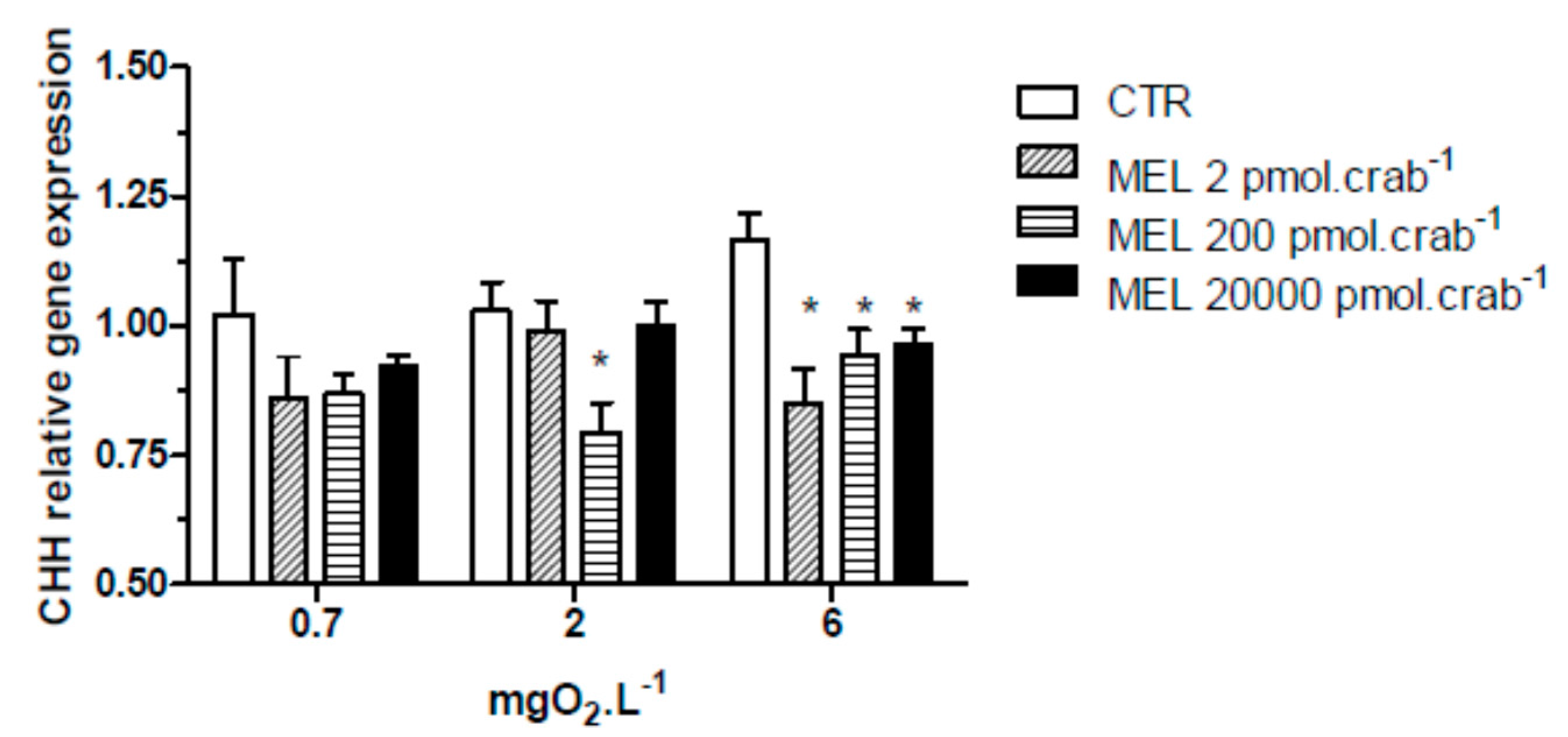
All three doses of melatonin tested were capable of significantly (
p < 0.05) decreasing CHH gene expression (
Figure 3) in the eyestalks of crabs exposed to 6 mgO
2·L
−1 (0.85 ± 0.07, 0.94 ± 0.04 and 0.96 ± 0.03 relative expression for 2, 200 and 20,000 pmol·crab
−1 of melatonin, respectively), compared to the control group (1.16 ± 0.06 relative expression). In crabs exposed to 2 mgO
2·L
−1, the dose of 200 pmol·crab
−1 of melatonin significantly (
p < 0.05) decreased (0.79 ± 0.03 relative expression) the CHH gene expression compared to the control group (1.03 ± 0.05 relative expression). No significant differences (
p > 0.05) were observed between the melatonin doses and the control group in crabs exposed to 0.7 mgO
2·L
−1.
Figure 1.
Melatonin content in the eyestalks of the crab Neohelice granulata in vitro. The eyestalks were incubated for 45 min at 0.7, 2, 4 or 6 mgO2·L−1. Each point represents the mean ± standard error (SE) (n = 5). * represents a significant difference (p < 0.05) from the other groups.
Figure 1.
Melatonin content in the eyestalks of the crab Neohelice granulata in vitro. The eyestalks were incubated for 45 min at 0.7, 2, 4 or 6 mgO2·L−1. Each point represents the mean ± standard error (SE) (n = 5). * represents a significant difference (p < 0.05) from the other groups.
Figure 2.
Melatonin content in the eyestalks of the crab Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (control group—CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). Different letters represent significant differences (p < 0.05) between the groups.
Figure 2.
Melatonin content in the eyestalks of the crab Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (control group—CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). Different letters represent significant differences (p < 0.05) between the groups.
Figure 3.
Crustacean Hyperglycemic Hormone (CHH) relative gene expression in the eyestalks of Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). * represents a significant difference (p < 0.05) from the CTR group. The values are normalized for β-actin gene expression.
Figure 3.
Crustacean Hyperglycemic Hormone (CHH) relative gene expression in the eyestalks of Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). * represents a significant difference (p < 0.05) from the CTR group. The values are normalized for β-actin gene expression.
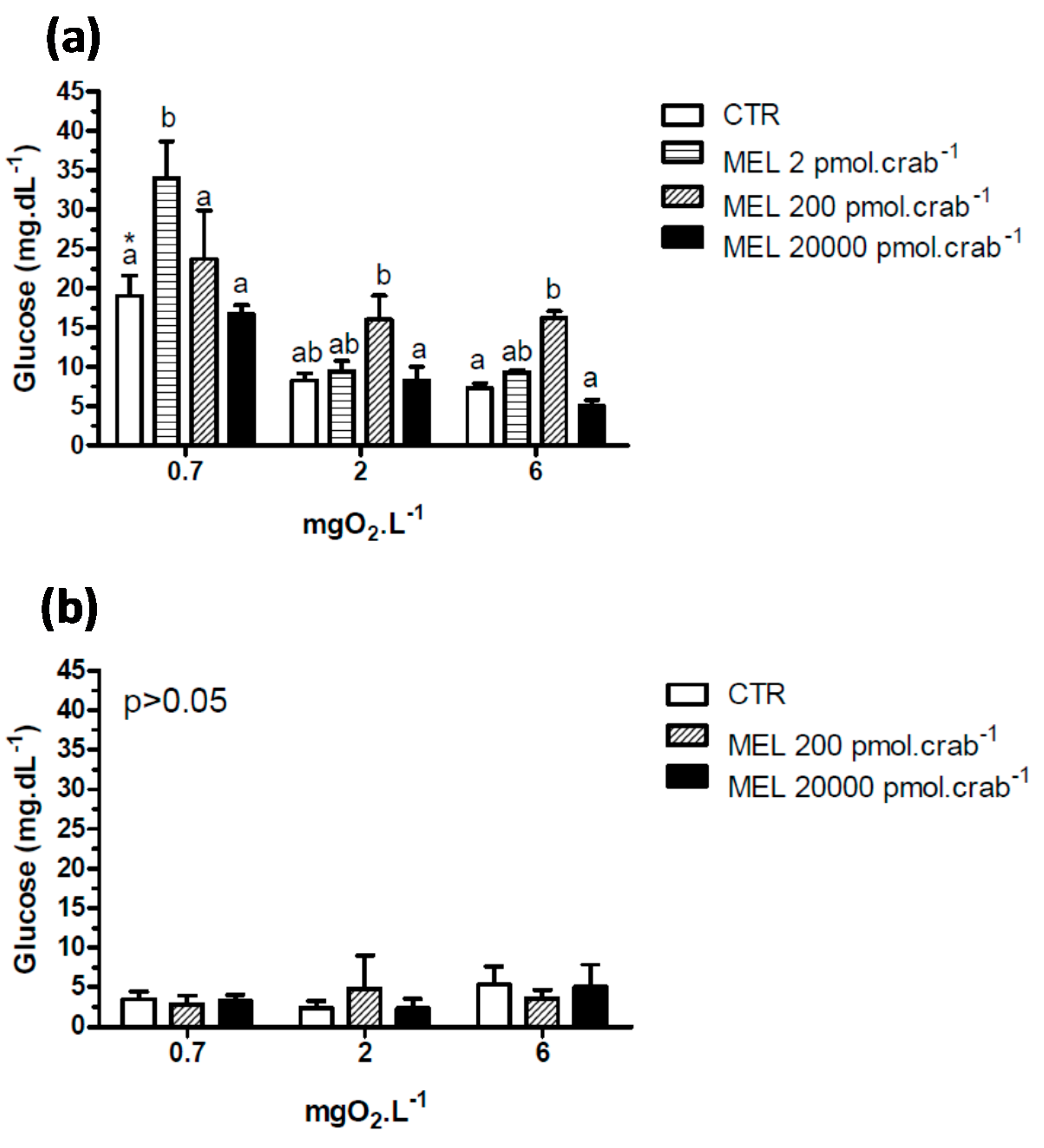
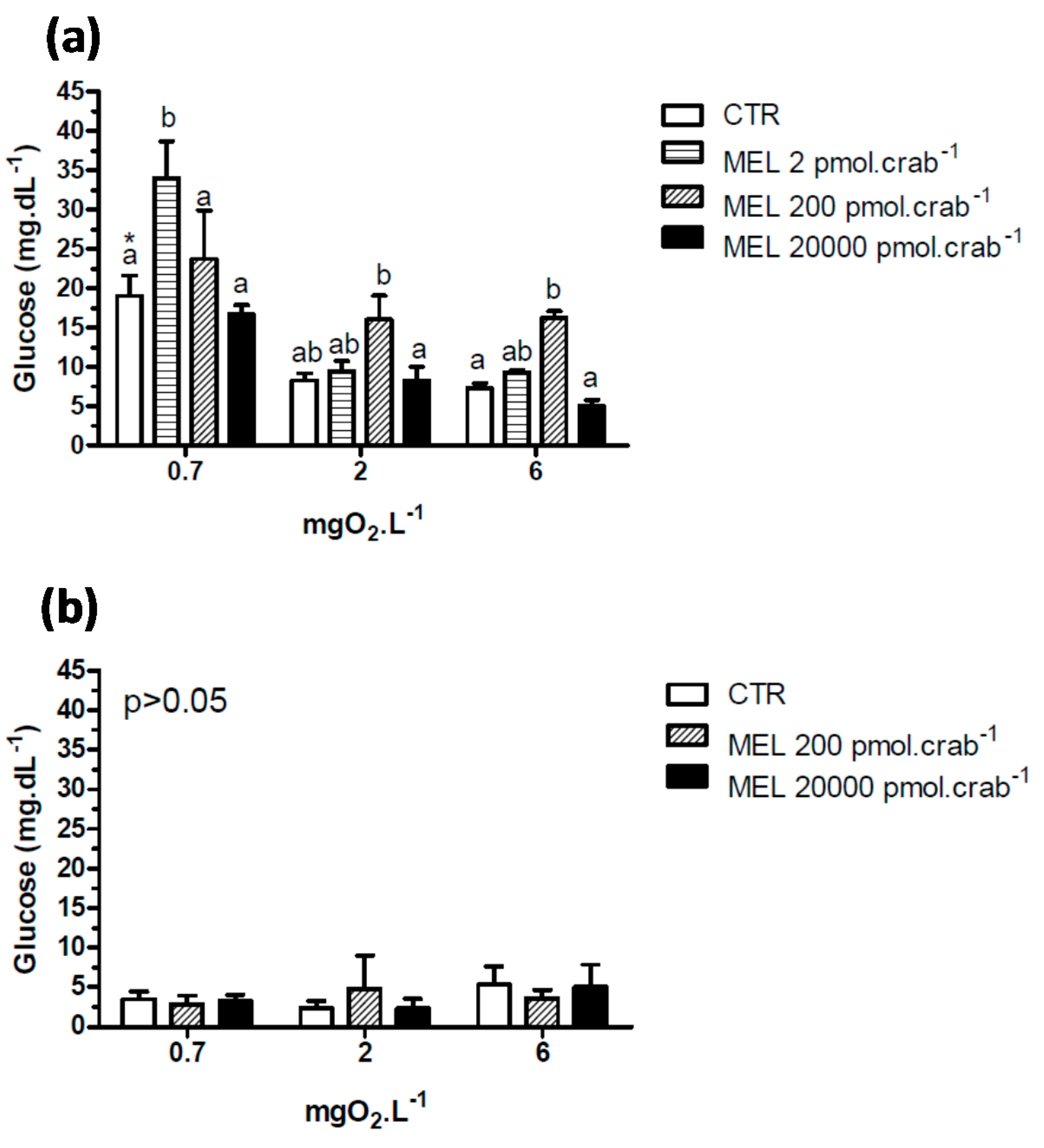
A significant hyperglycemia (
p < 0.05) was observed in intact crabs (
Figure 4a) exposed to 6 mgO
2·L
−1 and injected with 200 pmol·crab
−1 of melatonin (16.2 ± 0.87 mg·dL
−1) compared to the control group (7.3 ± 0.7 mg·dL
−1). This effect was not significant (
p > 0.05) in crabs exposed to 2 mgO
2·L
−1. In crabs exposed to 0.7 mgO
2·L
−1, a significant (
p < 0.05) hyperglycemia was observed in the control group (19 ± 2.6 mg·dL
−1) compared to the same group exposed to 6 (7.3 ± 0.7 mg·dL
−1) and 2 (8.2 ± 0.9 mg·dL
−1) mgO
2·L
−1. Interestingly, glucose levels increased significantly (
p < 0.05) in crabs exposed to 0.7 and injected with 2 pmol·crab
−1 of melatonin (33.9 ± 4.7 mg·dL
−1). Eyestalkless crabs (
Figure 4b) showed no significant differences (
p > 0.05) in all dissolved-oxygen concentrations and melatonin doses injected.
Figure 4.
Circulating glucose concentration in (a) intact and (b) eyestalkless Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 4–7). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. Different letters represent significant differences (p < 0.05) between the groups.
Figure 4.
Circulating glucose concentration in (a) intact and (b) eyestalkless Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 4–7). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. Different letters represent significant differences (p < 0.05) between the groups.
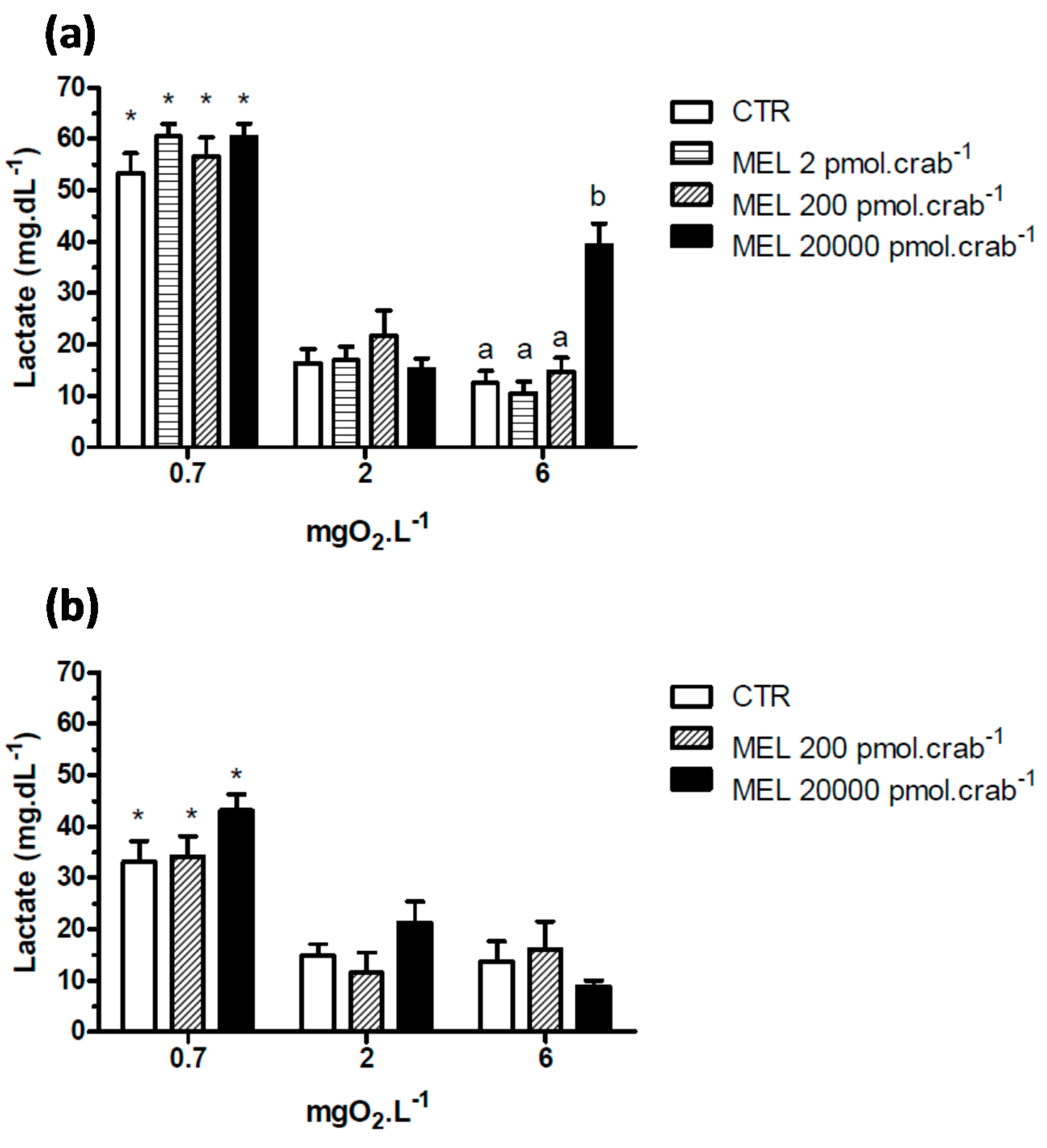
Intact crabs exposed to 6 mgO
2·L
−1 and injected with 20,000 pmol·crab
−1 of melatonin showed a significant (
p < 0.05) increase in lactate (
Figure 5a) concentration (39.3 ± 4.2 mg·dL
−1) compared to the control group (12.5 ± 2.1 mg·dL
−1). No significant effects (
p > 0.05) were observed in crabs exposed to 2 mgO
2·L
−1 and injected with the doses of melatonin. However, when the crabs were exposed to 0.7 mgO
2·L
−1, all groups showed a significant increase (
p < 0.05) in lactate concentration compared to the other dissolved-oxygen concentrations. In eyestalkless crabs (
Figure 5b) only the exposure to 0.7 mgO
2·L
−1 significantly increased (
p < 0.05) lactate levels in all treatment groups.
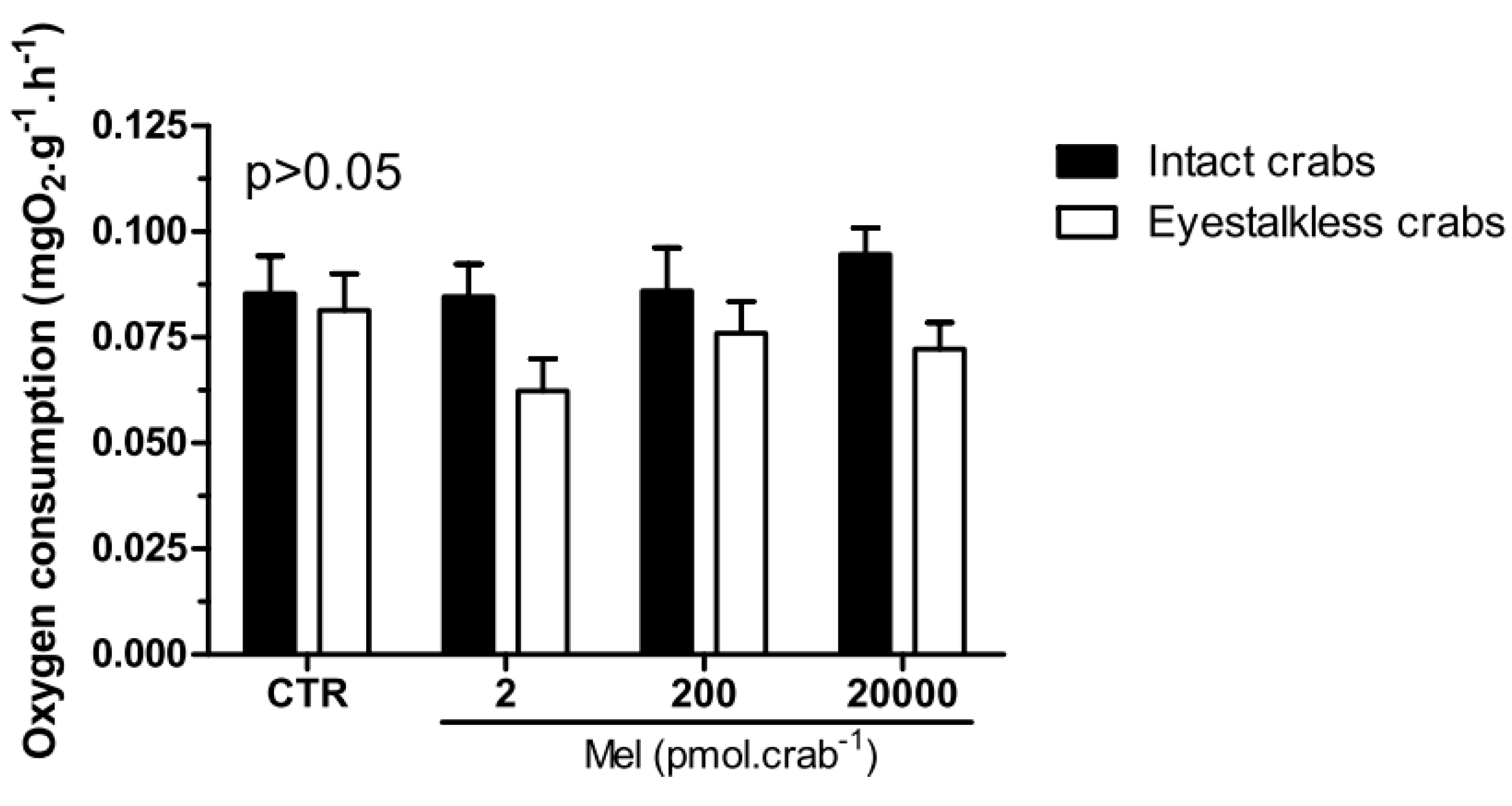
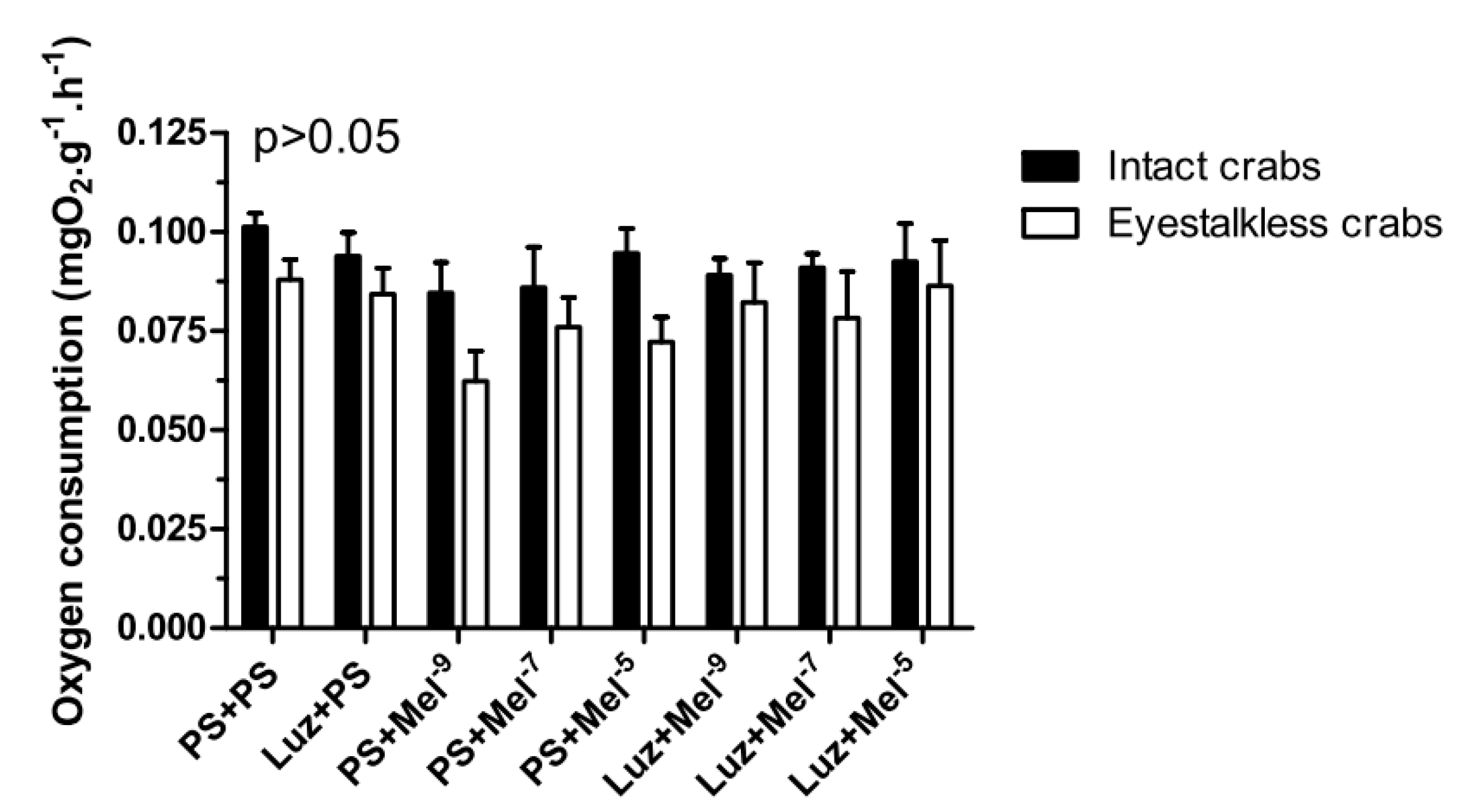
Intact and eyestalkless (
Figure 6) crabs exposed to 6 mgO
2·L
−1 and treated with melatonin showed no significant differences (
p > 0.05) in VO
2.
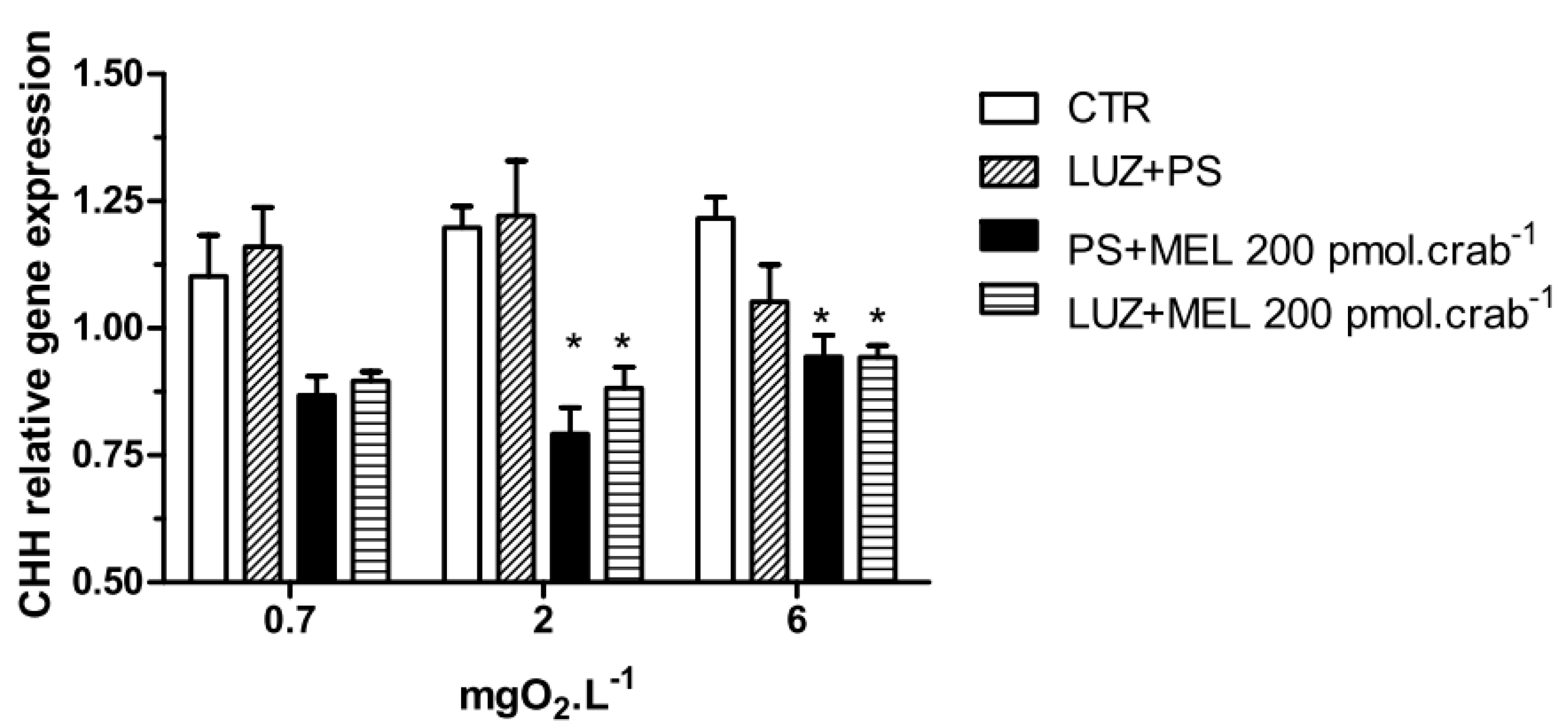
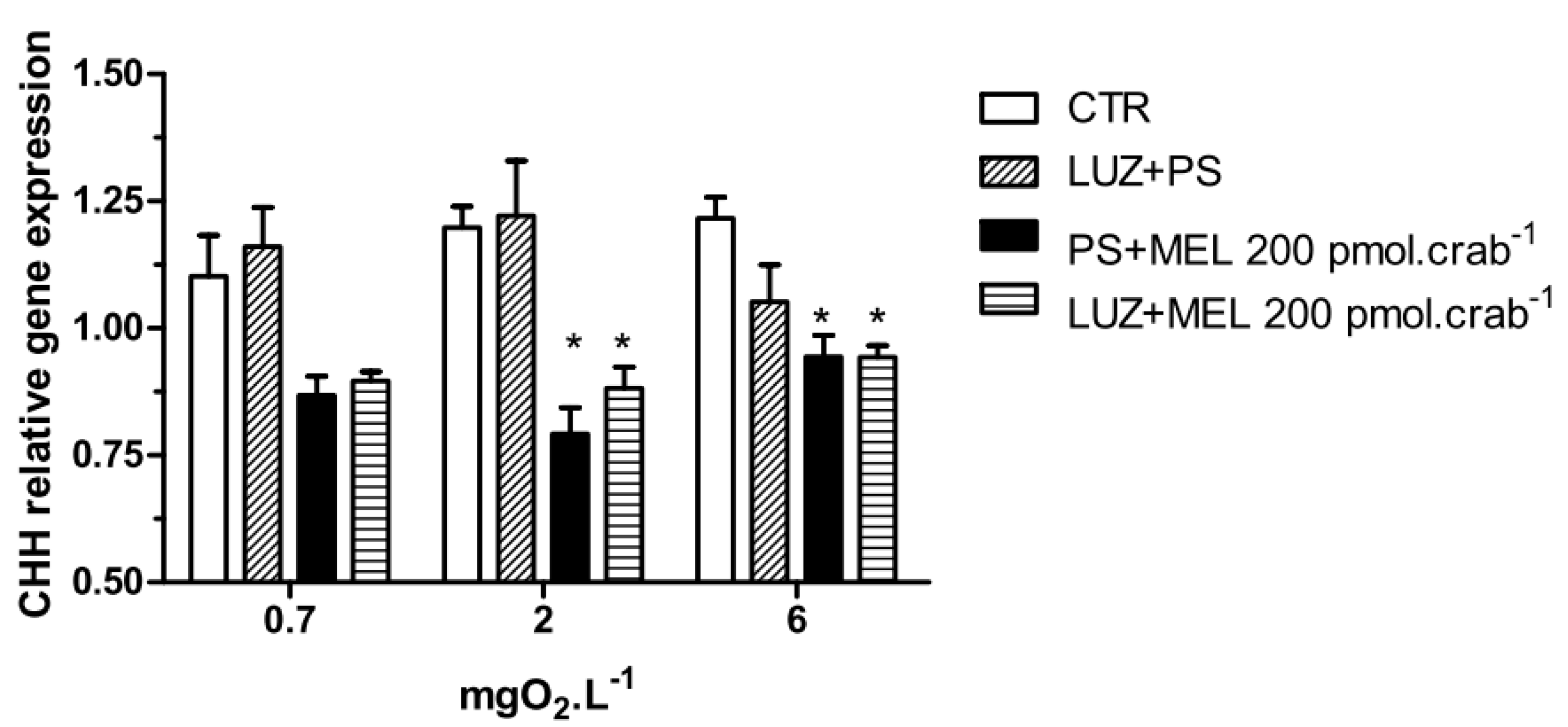
Melatonin (200 pmol·crab
−1) significantly decreased (
p < 0.05) the CHH gene expression in crabs exposed to 6 (0.94 ± 0.02 relative expression) and 2 (0.8 ± 0.05 relative expression) mgO
2·L
−1 independently of whether they were injected with luzindole (200 nmol·crab
−1), compared to the control group (1.21 ± 0.04 and 1.19 ± 0.04 relative expression, respectively) (
Figure 7). No significant differences were observed in animals exposed to 0.7 mgO
2·L
−1.
Figure 5.
Circulating lactate in (a) intact and (b) eyestalkless Neohelice granulata exposed for 45 min to three different dissolved oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 4–7). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
Figure 5.
Circulating lactate in (a) intact and (b) eyestalkless Neohelice granulata exposed for 45 min to three different dissolved oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 4–7). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
Figure 6.
Oxygen consumption (VO2) of intact (black bars) and eyestalkless (open bars) Neohelice granulata exposed for 45 min only to 6 mgO2·L−1 and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–6). No significant differences (p > 0.05) were observed.
Figure 6.
Oxygen consumption (VO2) of intact (black bars) and eyestalkless (open bars) Neohelice granulata exposed for 45 min only to 6 mgO2·L−1 and injected with physiological saline (CTR) or melatonin (2, 200 or 20,000 pmol·crab−1). Each point represents the mean ± SE (n = 3–6). No significant differences (p > 0.05) were observed.
Figure 7.
Crustacean Hyperglycemic Hormone (CHH) relative gene expression in the eyestalks of Neohelice granulata exposed for 45 min to three different dissolved oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). * represents a significant difference (p < 0.05) from the CTR. The values are normalized for β-actin gene expression.
Figure 7.
Crustacean Hyperglycemic Hormone (CHH) relative gene expression in the eyestalks of Neohelice granulata exposed for 45 min to three different dissolved oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). * represents a significant difference (p < 0.05) from the CTR. The values are normalized for β-actin gene expression.
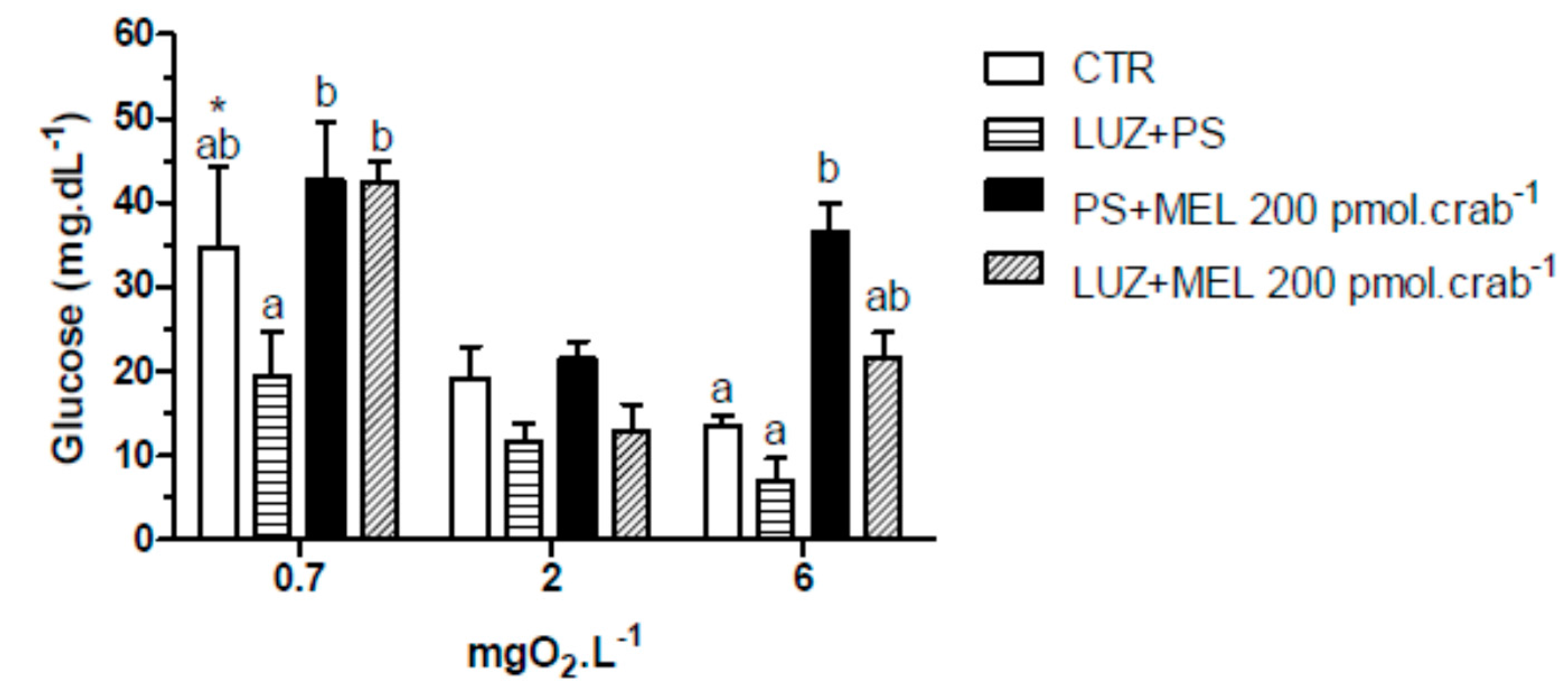
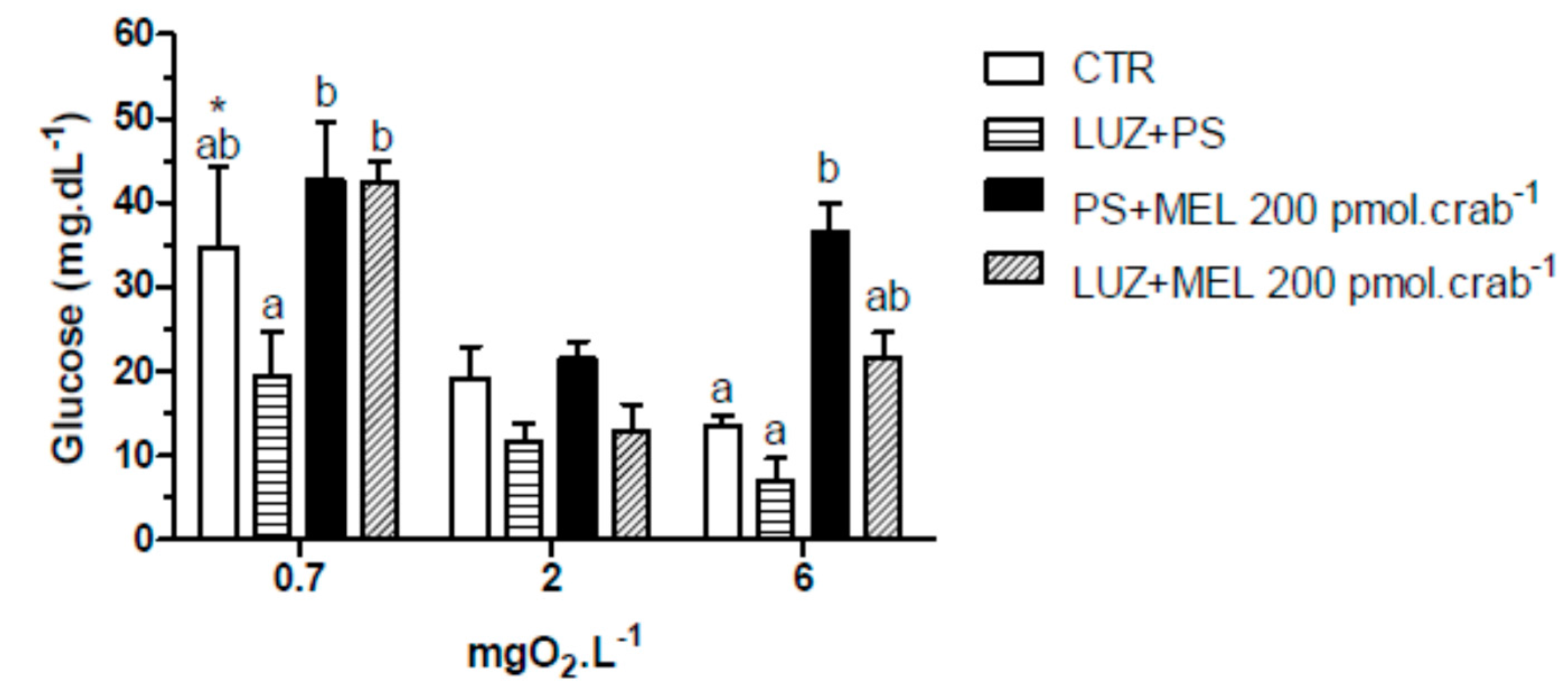
Again, injections of melatonin (200 pmol·crab
−1) significantly increased (
p < 0.05) glucose levels in intact crabs (
Figure 8) exposed to 6 mgO
2·L
−1 (36.4 ± 3.5 mg·dL
−1); but crabs that were also injected with luzindole (200 nmol·crab
−1) did not differ (
p > 0.05) from the control group (21.5 ± 3.5 and 3.4 ± 1.3 mg·dL
−1, respectively). No significant effects (
p > 0.05) of melatonin or luzindole were observed in crabs exposed to 2 mgO
2·L
−1. In 0.7 mgO
2·L
−1 a general increase of glucose levels was observed in the treatment groups (34.7 ± 9.5 mg·dL
−1, CTR as reference;
p < 0.05 compared to CTR of 6 mgO
2·L
−1).
Figure 8.
Circulating glucose concentration in intact Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 4–6). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
Figure 8.
Circulating glucose concentration in intact Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 4–6). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
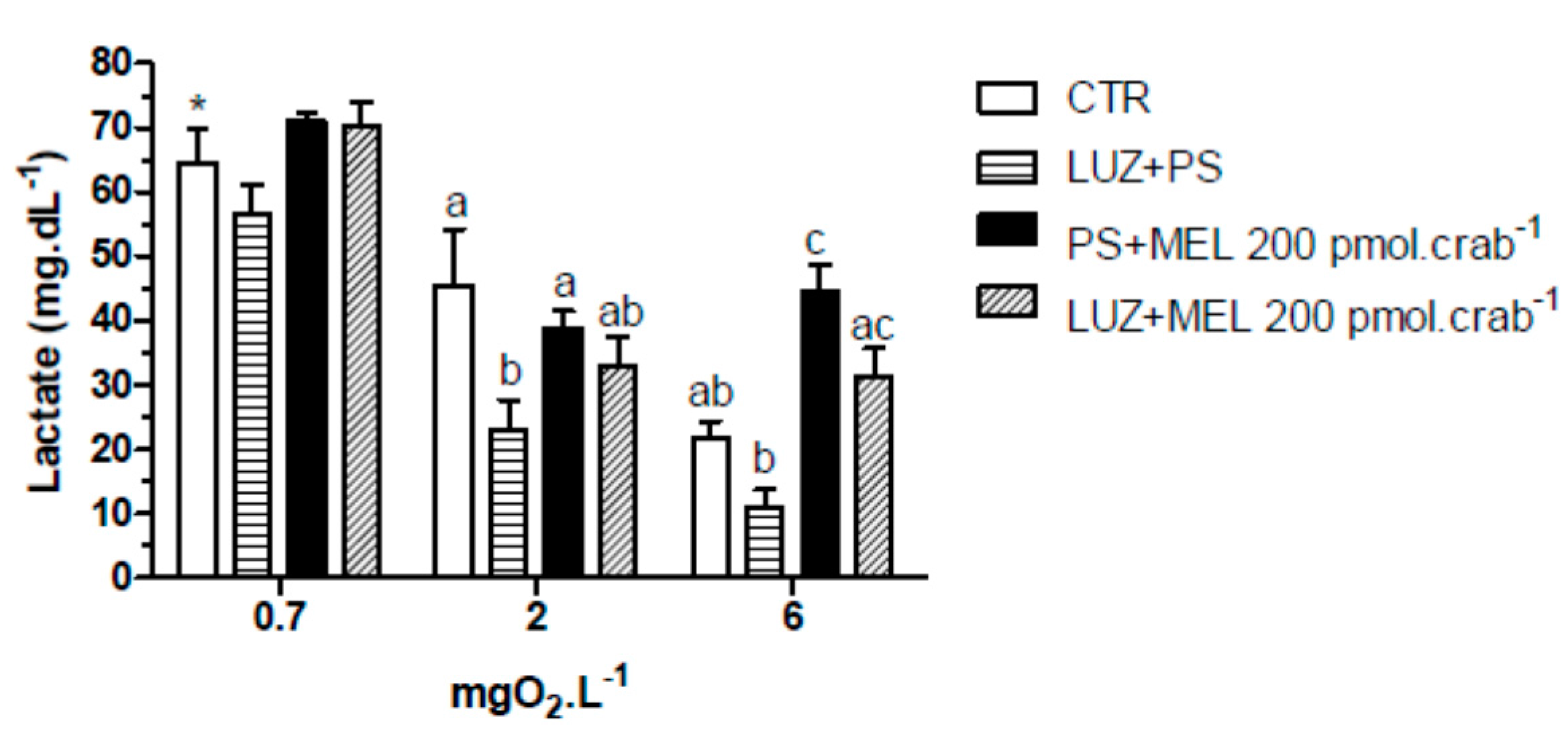
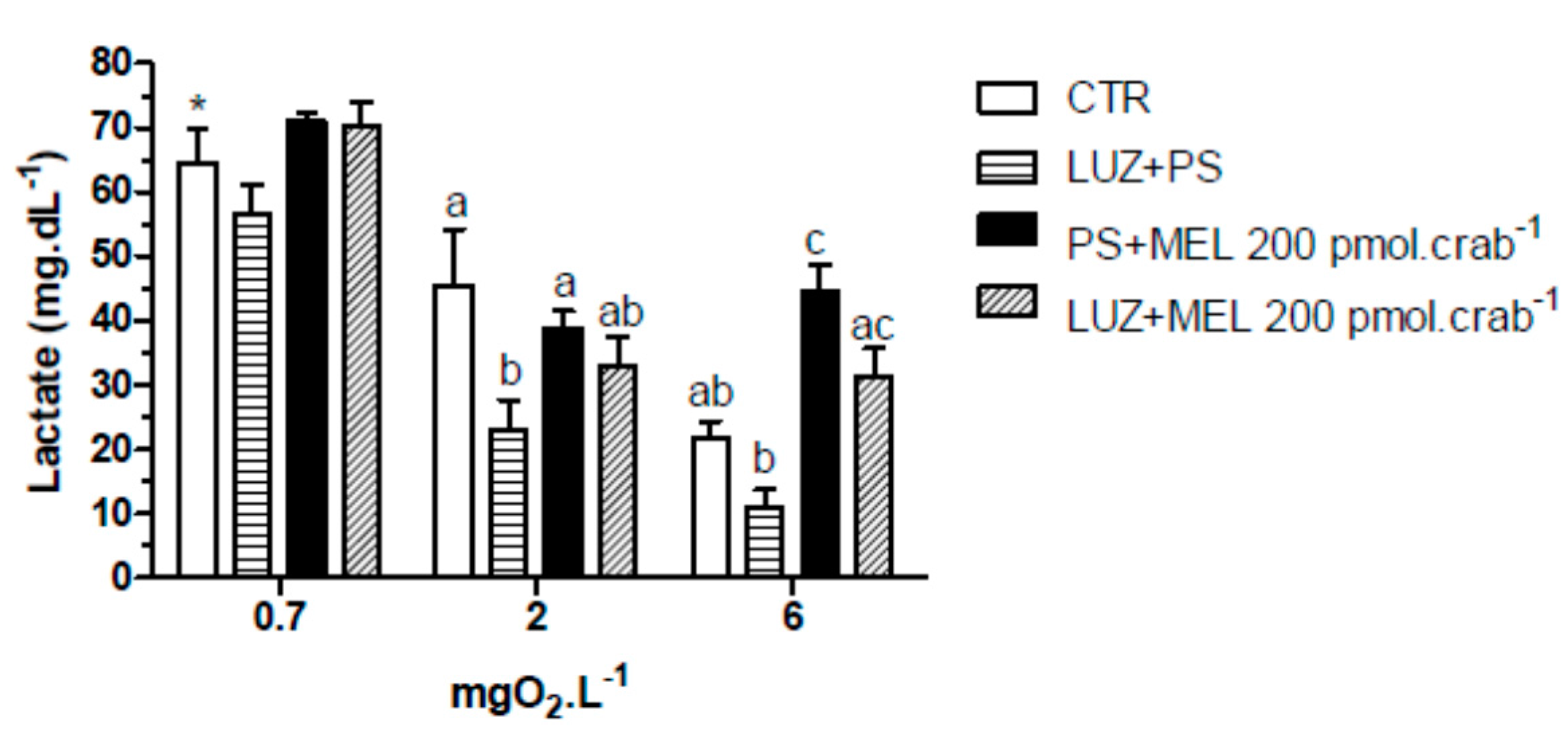
Figure 9.
Circulating lactate concentration in intact Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 4–6). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
Figure 9.
Circulating lactate concentration in intact Neohelice granulata exposed for 45 min to three different dissolved-oxygen concentrations (0.7, 2 or 6 mgO2·L−1) and injected with physiological saline only (CTR), luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 4–6). Different letters represent significant differences (p < 0.05) between the groups with the same dissolved oxygen concentration. * represents a significant difference (p < 0.05) from the CTR group at 6 mgO2·L−1.
Similar effects were observed in the lactate measurements (
Figure 9). Melatonin (200 pmol·crab
−1) significantly increased (
p < 0.05) lactate concentration in crabs exposed to 6 mgO
2·L
−1 (44.5 ± 4.2 mg·dL
−1), but crabs also injected with luzindole (200 nmol·crab
−1) (31.5 ± 4.8 mg·dL
−1) did not differ (
p > 0.05) from the control group (21.8 ± 2.9 mg·dL
−1). At 2 mgO
2·L
−1 luzindole significantly decreased (
p < 0.05) lactate levels (23.1 ± 5.2 mg·dL
−1) compared to the control group (45.5 ± 6.1 mg·dL
−1) but did not significantly affect the melatonin group. Crabs exposed to 0.7 mgO
2·L
−1 showed a general lactemia in all treatment groups (64.5 ± 6.1 mg·dL
−1, CTR as reference;
p < 0.05 compared to CTR of 6 mgO
2·L
−1).
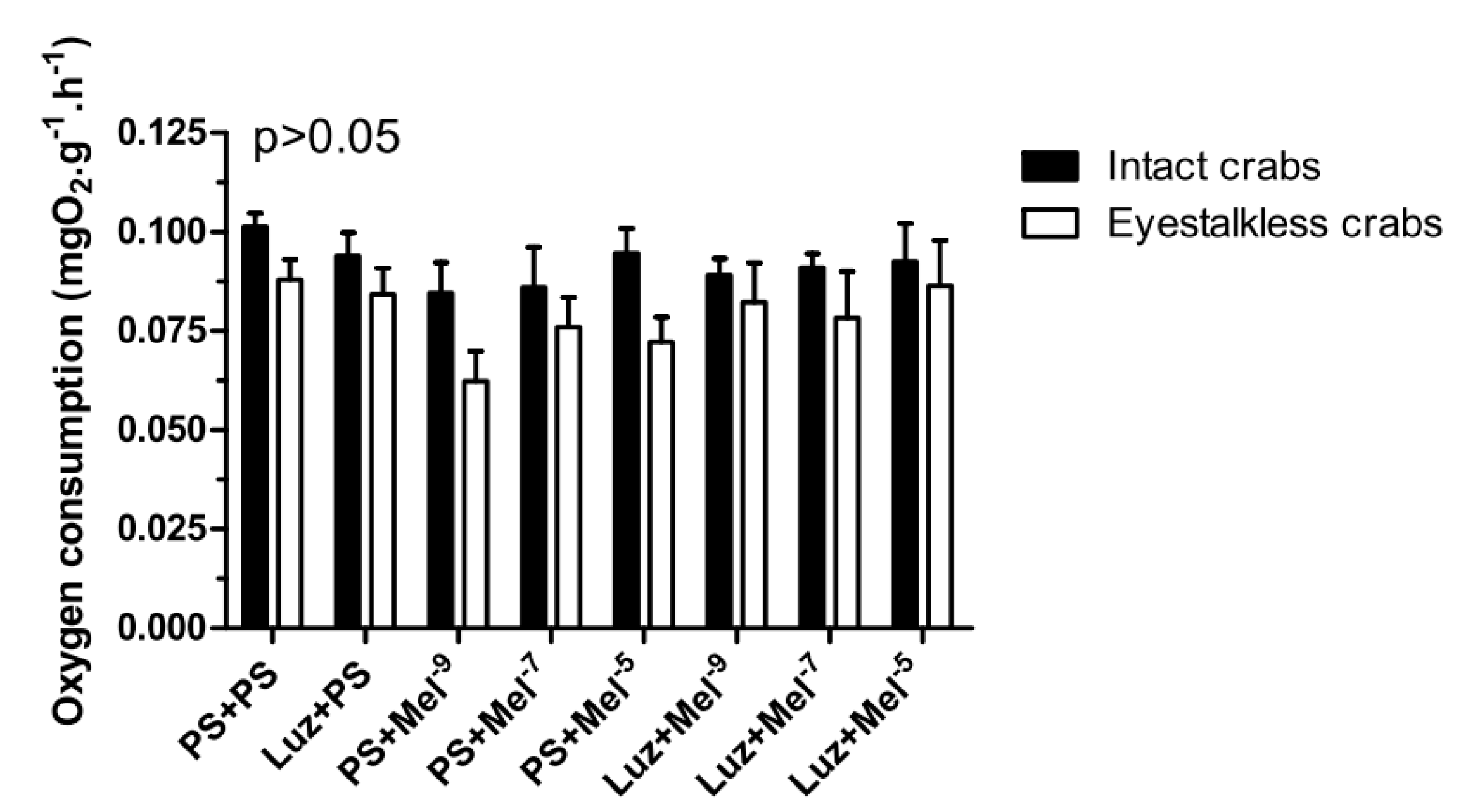
In the VO
2 measurements, no significant differences (
p > 0.05) were observed between the treatment groups in intact and eyestalkless (
Figure 10) crabs exposed to 6 mgO
2·L
−1.
Figure 10.
Oxygen consumption (VO2) of intact (black bars) and eyestalkless (open bars) Neohelice granulata exposed for 45 min only to 6 mgO2·L−1 and injected with physiological saline only (CTR) luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). No significant differences (p > 0.05) were observed.
Figure 10.
Oxygen consumption (VO2) of intact (black bars) and eyestalkless (open bars) Neohelice granulata exposed for 45 min only to 6 mgO2·L−1 and injected with physiological saline only (CTR) luzindole (200 nmol·crab−1) plus physiological saline, physiological saline plus melatonin (200 pmol·crab−1), or luzindole (200 nmol·crab−1) plus melatonin (200 pmol·crab−1). Each point represents the mean ± SE (n = 3–4). No significant differences (p > 0.05) were observed.
3. Discussion
As mentioned above, melatonin is ubiquitous in living organisms. In crustaceans, melatonin has been identified in a dozen species [
8,
9], but its function is unclear. Recent studies have shown an association of this molecule with metabolism in crabs. Maciel and co-workers [
10] found that gills of
N. granulata decreased their VO
2 when incubated for 120 min with 20 × 10
−9 M of melatonin, and also decreased gill VO
2 in eyestalkless crabs injected with melatonin after 390 min. In contrast, when an equivalent dose of melatonin was injected in the same species, no effect was observed in muscle VO
2 [
33]. In the present study, melatonin did not cause any change in overall VO
2 in both intact and eyestalkless crabs. Taken together, these data suggest that this indoleamine effect may be tissue-specific for depression of aerobic metabolism, but as muscle tissue is the most important, the overall VO
2 in this crab is not significantly affected by melatonin, at least in the dosages tested.
In addition to aerobic metabolism, melatonin seems to have a role in glucose/lactate effects in crustaceans. In eyestalkless
Uca pugilator, injections of melatonin (3 × 10
−9 mol·animal
−1) caused an increase in glucose and lactate levels, indicating an independence of this effect of CHH from the eyestalk [
11]. In this case, melatonin could act as a secretagogue of other CHH-releasing tissue and/or act directly in the breakdown of glycogen to glucose through glycogen phosphorylase in the tissues. Sainath and Reddy [
12] observed an increase of phosphorylase activity in hepatopancreas and muscle of intact and eyestalkless crab
Oziotelphusa senex senex when injected with melatonin (10
−7 mol·crab
−1), and also hyperglycemia in a dose-dependent manner (10
−10–10
−5 mol·crab
−1), which supports the hypothesis that melatonin-induced hyperglycemia is not through CHH from the eyestalk, at least in this species. Here, however, a hyperglycemia was observed in intact but not in eyestalkless
N. granulata injected with 200 pmol·crab
−1, indicating a dependence on CHH from the eyestalks. In addition, this effect seems to occur through melatonin receptors, since with the administration of luzindole (200 nmol·crab
−1), glucose levels did not significantly differ from the control group.
The effect of melatonin on hyperglycemia in N. granulata is affected by hypoxia. In severe hypoxia (0.7 mg·L−1), the lowest melatonin dose (2 pmol·crab−1) caused hyperglycemia, but 200 pmol·crab−1 did not. The hypoxic condition seems to increase sensitization to melatonin effects, suggesting that melatonin in low doses could mobilize carbohydrates.
Melatonin (20,000 pmol·crab
−1) also increased lactate levels in
N. granulata, but only when the crabs were exposed to 6 mgO
2·L
−1. In severe hypoxia, the lactate concentration increased significantly, independently of whether the crabs were treated with melatonin or not. In this species, some studies have found that lactate is the main metabolite of anaerobic metabolism. In addition, the lactate level increases when the crab is exposed to hypoxic or anoxic situations [
23,
25,
32]. However, this effect is not totally eyestalk-dependent, since eyestalkless crabs showed a lower but still significant increase in lactate levels.
Interestingly, melatonin decreased CHH gene expression when
N. granulata was exposed for 45 min to 6 (2, 200 and 20,000 pmol·crab
−1) and 2 (200 pmol·crab
−1) mgO
2·L
−1, but its effect seems to occur through another pathway than the melatonin receptor. This downregulation of the CHH gene expression by melatonin could be an indirect effect of this indoleamine and a negative feedback response to the increase in circulating glucose levels. Chung and Zmora [
34] also observed an inhibition of CHH expression in the blue crab
Callinectes sapidus, but in response to 1 h of hypoxia conditions. Again, a negative feedback could be regulating this gene in response to the hyperglycemia seen in
C. sapidus under hypoxic conditions [
34]. However, this effect was not observed in
N. granulata [
25], perhaps because this species is more tolerant to hypoxia or anoxia than
C. sapidus, and 45 min was not sufficient to induce downregulation in eyestalk CHH expression.
In addition to the various functions previously described for melatonin, this molecule has the ability, at least in mammals, to influence insulin secretion, and metabolism of lipids and glucose [
35,
36,
37,
38,
39]. Additionally, some studies have observed that this indoleamine may modulate the release of neurohormones in insects, such as the prothoracicotropic hormone in
Periplaneta americana [
40] and the adipokinetic precursor-related peptide in
Locusta migratoria [
41]. The only similar study on crustaceans was conducted by Tilden
et al. [
42], who observed an increase in synaptic transmission at the neuromuscular junction when melatonin was injected into
Procambarus clarkii. In the present study, melatonin induced hyperglycemia, increased circulating lactate, and inhibited CHH gene expression. This indoleamine could be stimulating CHH release, and as a consequence, the higher levels of glucose could inhibit CHH gene expression; or perhaps the CHH peptide regulates its transcription.
Melatonin membrane receptors have been identified in vertebrates, and are classified as MT1, MT2 and MT3 [
43,
44]. MT1 and MT2 are G protein-coupled receptors, and MT3 is an enzyme belonging to the family of quinone reductases, although its signaling pathway is still unknown. In invertebrates, melatonin receptors are little understood. Richter
et al. [
40] found that luzindole (10 nM) inhibited the prothoracicotropic hormone from the brain of
P. americana when incubated with melatonin (10 nM). Concerning crustaceans, Mendoza-Vargas [
45] observed an inhibition of the receptor potential amplitude of retinular photoreceptors by DH97 (1 nM), a MT
2 receptor-selective antagonist, whereas melatonin (100 nM) increased the receptor potential amplitude in the crayfish
P. clarkii. Here, luzindole (200 nmol·crab
−1) inhibited the increase in glucose and lactate caused by melatonin (200 pmol·crab
−1), but did not affect the CHH gene expression in
N. granulata. Thus, melatonin may have different modes of action in the regulation of carbohydrate metabolism, since the melatonin response was seen at transcriptional and physiological levels.
Eyestalks incubated for 45 min at 2, 4 or 6 mgO
2·L
−1 showed similar levels of melatonin, as previously reported [
8]. However, at 0.7 mgO
2·L
−1 the melatonin content increased (
Figure 1). This response was seen only
in vitro, not in
in vivo experiments, suggesting that melatonin may be a signaling molecule in this tissue only under conditions of severe hypoxia. Interestingly, injection of melatonin increased the melatonin content in the eyestalks of crabs exposed to different dissolved-oxygen concentrations, in a dose-dependent manner (see
Figure 2). But, in animals injected with 200 pmol·crab
−1 and exposed to 0.7 mgO
2·L
−1, the melatonin content in the eyestalks was higher than in the other groups. Again, these results reinforce the suggestion that melatonin might be involved in metabolic regulation in this crab when it undergoes hypoxic conditions.
In conclusion, melatonin seems to induce hyperglycemia and anaerobic metabolism in N. granulata, since glucose and lactate are augmented, but these responses are eyestalk- and melatonin receptor-dependent. Also, melatonin down regulates CHH gene expression, but not by a known melatonin receptor mechanism.
4. Material and Methods
4.1. Animals
Adult male crabs of N. granulata (mean weight ± confidence interval: 11.2 ± 1.1 g) were collected in salt marshes near the city of Rio Grande in southern Brazil. In the laboratory, the crabs were acclimated under constant conditions of salinity (20), temperature (20 °C) and photoperiod (12L:12D) for at least 7 days. During this period, the crabs were fed regularly with ground beef.
4.2. Reagents
Melatonin (MEL) and luzindole (LUZ) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Stock solutions were prepared with ethanol and dimethyl sulfoxide (DMSO), respectively. Working solutions were prepared with crustacean physiological solution (PS), which contained: 1 × 10−2 M MgCl2, 355 × 10−3 M NaCl, 16.6 × 10−3 M CaCl2, 5 × 10−3 M H3BO3, 1 × 10−2 M KHCO3 and 8 × 10−3 M sodium citrate (Na3C6H5O7·2H2O); pH adjusted to 7.6.
4.3. RNA Isolation and cDNA Synthesis
The eyestalk ganglia were collected, dissected, and used for total RNA extraction. Total RNA was isolated by the Trizol Reagent Solution™ method, following the manufacturer’s instructions (Applied Biosystems, Grand Island, NY, USA). Genomic DNA was degraded with DNase I (Invitrogen) and total RNA was quantified by a Qubit™ fluorometer using the Quant-iT™ RNA assay kit (Invitrogen, Waltham, MA, USA), and calibration was carried out using a two-point standard curve. First-strand cDNA was synthesized from approximately 1 μg of total RNA and used as a template with the SuperScript III enzyme kit (Invitrogen) according to the manufacturer’s suggested protocol.
4.4. Experimental Design and Measurements of CHH Gene Expression, VO2, Glucose and Lactate for Different O2 Concentrations and Melatonin Dose Effects
Twelve aquaria (3 L each) were purged with nitrogen gas to reduce the level of dissolved oxygen to 6 mgO2·L−1 (normoxia), 2 mgO2·L−1 (low hypoxia) or 0.7 mgO2·L−1 (severe hypoxia) (4 aquaria each). After that, the crabs (total of 192 animals) were injected with 100 μL of PS (control) or MEL (20 × 10−9, 20 × 10−7 or 20 × 10−5 M) and exposed for 45 min in one of the three oxygen concentrations, prior to eyestalk collection for CHH gene expression (n = 4, pool of 4 crabs) and hemolymph sampling (n = 5). An additional experiment was carried out with eyestalkless crabs (total of 45 animals) under the same conditions as for hemolymph sampling; animals had their eyestalks ablated (the eyestalks were cut off with scissors and then cauterized) 24 h prior to the beginning of the experiment. The eyestalk dissection and total RNA and cDNA synthesis were performed as described above. Quantitative analysis of CHH mRNA expression was carried out for 10 min at 95 °C and 40 cycles of 15 s at 95 °C and 1 min at 60 °C by real-time RT-PCR (7300 Real-Time System, Applied Biosystems), using SYBR GREEN PCR Master Mix™ (Applied Biosystems), CHH-specific primers (sense 5'-CGCACACGTGTTCCAGTTTG-3', and anti-sense 5'-AGATGCCCTTGCAGGAACG-3'; GenBank accession number ADM26761), Ultra Pure Distilled Water (Invitrogen) and 1.0 μL cDNA (dilution 1:19). Β-actin expression (sense 5'-CCAGATCATGTTTGAGGTGTTCA-3' and anti-sense 5'-GGGACAGCACAGCCTGGAT-3' primers; GenBank accession number AY074923) was used as an internal control to normalize the data.
Glucose and lactate were measured using the GOD-PAD monoreagent kit (Kovalent, Rio de Janeiro, Brazil) and lactate kit (Kovalent), respectively. Glucose and lactate are expressed in mg per dL of hemolymph.
For VO2 measurement, 9 aquaria (3 L each) were purged with nitrogen gas to reduce the level of dissolved oxygen to 6, 2 or 0.7 mgO2·L−1 (3 aquaria each). Thirty-six intact (n = 4) and 36 eyestalkless (n = 4) crabs were injected with 100 μL of PS (control) or MEL (20 × 10−9 or 20 × 10−7 M) and exposed to the above oxygen concentrations in the respirometer chamber with saline water (salinity at 20) for 15 min prior to the beginning of the experiment. Oxygen concentration was measured at time zero and after 30 min using a portable oxymeter. Oxygen consumption was expressed in milligrams of oxygen per gram of body weight per hour. The eyestalks were ablated as described above.
4.5. Experimental Design and Measurements of CHH Gene Expression, VO2, Glucose and Lactate for Different O2 Concentrations and Melatonin and Luzindole Dose Effects
Twelve aquaria (3 L each) were purged with nitrogen gas to reduce the level of dissolved oxygen to 6, 2 or 0.7 mgO2·L−1 (4 aquaria each). Next, the crabs (total of 192 animals) received two injections of 100 μL with a 15-min interval between them. One group received 2 injections of PS (control), the second group was injected first with LUZ (10−4 M) and then with PS, the third group was injected with PS and then with MEL (20 × 10−7 M), and the fourth group was injected first with LUZ (10−4 M) and then with MEL (20 × 10−7 M). Next, the crabs were exposed for 45 min to each oxygen concentration prior to eyestalk collection for CHH gene expression (n = 4, pool of 4 crabs) and hemolymph sampling (n = 5). An additional experiment was carried out with eyestalkless crabs (total of 45 animals) under the same conditions as for hemolymph sampling. The eyestalk dissection and analyses of total RNA, cDNA synthesis and CHH gene expression were performed as described above, as were the glucose and lactate measurements.
For VO2 measurement, 12 aquaria (3 L each) were purged with nitrogen gas to reduce the level of dissolved oxygen to 6 mgO2·L−1. Thirty-six intact (n = 4) and 36 eyestalkless (n = 4) crabs received two injections of 100 μL with a 15-min interval between them. The treatments were the same as described above: PS plus PS, LUZ (10−4 M) plus PS, PS plus MEL (20 × 10−7 M), or LUZ (10−4 M) plus MEL (20 × 10−7 M). After that, the animals were exposed to the above oxygen concentration in the respirometer chamber with saline water (salinity at 20) for 15 min prior to the beginning of the experiment. Oxygen concentration was measured at time zero and after 30 min, using a portable oxymeter. Oxygen consumption was expressed in milligrams of oxygen per gram of body weight per hour. The eyestalks were ablated as described above.
4.6. Experimental Design and Measurements of Melatonin Eyestalk Ganglia in Vitro and in Vivo
In in vitro experiments, 10 eyestalks (n = 5) were incubated for 45 min at 0.7, 2, 4 or 6 mgO2·L−1. The eyestalks were then frozen at −80 °C for further analyses.
The frozen optic lobes were homogenized in 0.5 mL ethanol, centrifuged for 10 min (1000×
g), and the supernatants were collected and lyophilized. After the samples were diluted with phosphate-buffered saline (PBS) and Radioimmunoassay (RIA) buffer (400 µL/sample), they were sonicated for 4–5 s in an ice bath and centrifuged for 5 min (10,000×
g). The supernatants were directly analyzed for melatonin (2 × 100 µL) as described previously, using radio-iodinated melatonin as the tracer [
46,
47].
In in vivo experiments, 12 aquaria (3 L each) were purged with nitrogen gas to reduce the level of dissolved oxygen to 6, 2 or 0.7 mgO2·L−1 (4 aquaria each). Forty-eight (n = 4) crabs were injected with 100 μL of PS (control) or MEL (20 × 10−12, 20 × 10−9 or 20 × 10−7 M) and exposed to the above oxygen concentrations in the respirometer chamber with saline water (salinity at 20) for 15 min prior to the beginning of the experiment. Oxygen concentration was measured at time zero and after 30 min, using a portable oxymeter. The eyestalks were then frozen at −80 °C for further melatonin quantification as described above.
4.7. Statistical Analysis
In the CHH gene expression, the REST
© software (Munich, Germany) was applied in the relative quantification [
48], with pairwise comparisons. For VO
2, glucose and lactate, a two-way ANOVA (Analysis of Variance) was carried out followed by an
a posteriori comparison (Newman–Keuls test, α = 0.05). ANOVA assumptions (normality and homogeneity of variance) were verified prior to ANOVA analysis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}