Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review




Abstract
:1. Introduction
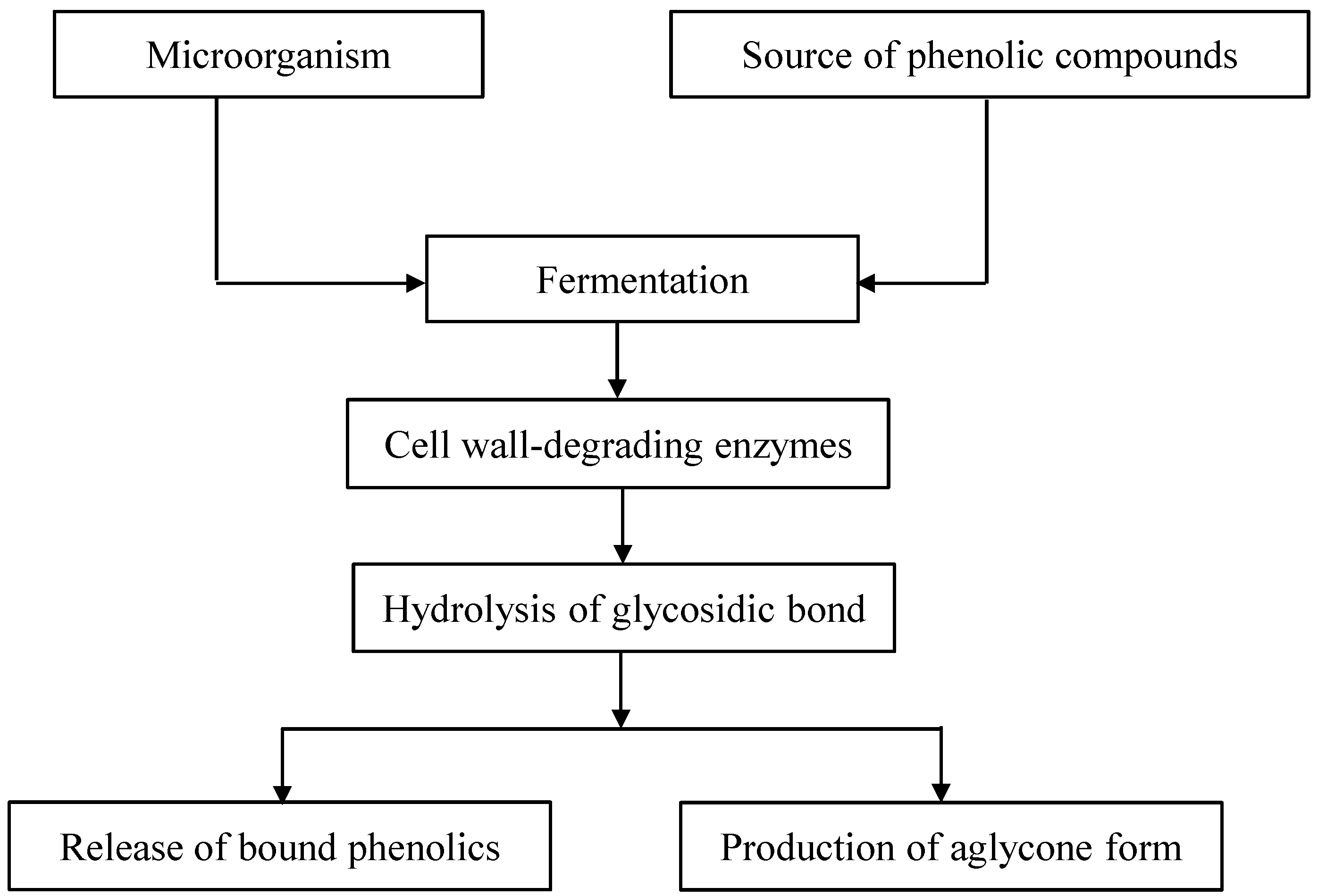
2. The Release of Phenolic Compounds from Plant Matrices by Fermentation Processes

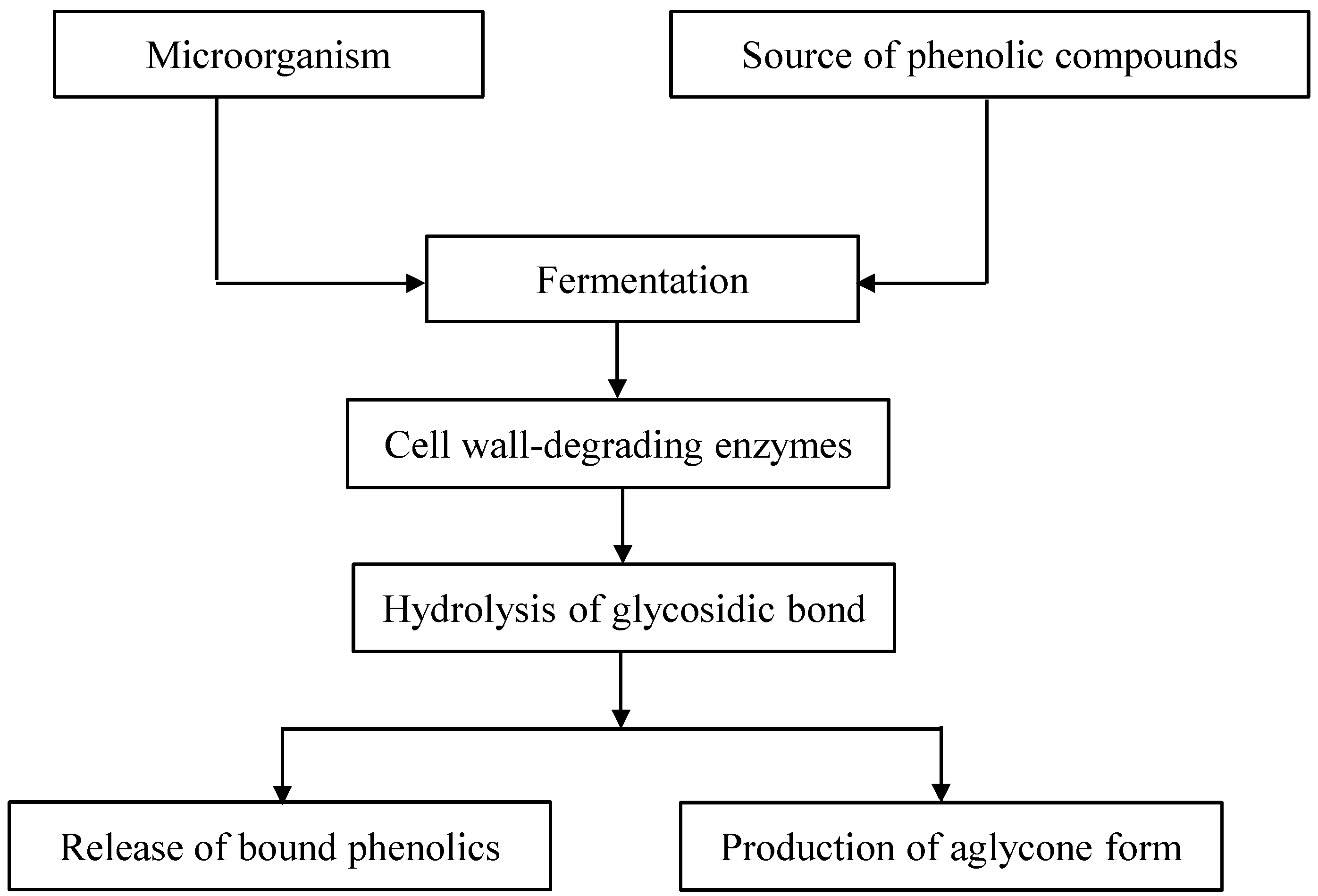{kind=link}
{kind=link}
| Microorganism | Source | Phenolic profile | Reference |
|---|---|---|---|
| Bacteria | |||
| Bacillus pumilus | Soybean | Gallic acid, catechin, epicatechin | [31] |
| Bacillus subtilis | Soybean | Chlorogenic acid, naringin | [32] |
| Bacillus subtilis | Cheonggukjang (soybean paste) | Daidzein, genistein | [33,34] |
| Lactobacillus acidophilus | Apple juice | Gallic acid | [35] |
| Lactobacillus johnsonii, Lactobacillus reuteri, Lactobacillus acidophilus | Whole grain barley, oat groat | Sinapic acid, caffeic acid, p-coumaric acid, ferulic acid | [36] |
| Lactobacillus plantarum | Cowpeas | Quercetin | [37] |
| Lactobacillus plantarum, Lactobacillus delbrueckii supsp. lactis | Soybean | Daidzein, genistein | [38] |
| Yeast | |||
| Saccharomyces cerevisiae | Wheat bran | Syringic acid, p-coumaric acid, ferulic acid | [39] |
| Fungi | |||
| Aspergillus oryzae, Monascus purpureus | Soybean | Daidzein, genistein | [40,41,42] |
| Aspergillus oryzae var. effuses, Aspergillus oryzae, Aspergillus niger | Oat (Avena sativa L.) | Chlorogenic acid, ferulic acid, p-coumaric acid, caffeic acid | [29] |
| Aspergillus oryzae | Green tea | Gallic acid, gallocatechin, epigallocatechin, epicatechin, 3-p-coumaroylquinic acid, kaempferol-rutinoside | [43] |
| Lentinus edodes | Cranberry pomace (Vaccinium acrocarpon) | Ellagic acid | [27,28] |
| Rhizopus oryzae | Rice bran | Gallic acid, ferulic acid, p-hydroxybenzoic acid, caffeic acid, chlorogenic acid, vanillin | [30] |
| Rhizopus oligosporus, Rhizopus oryzae | Black soybean | Daidzein, genistein | [44] |
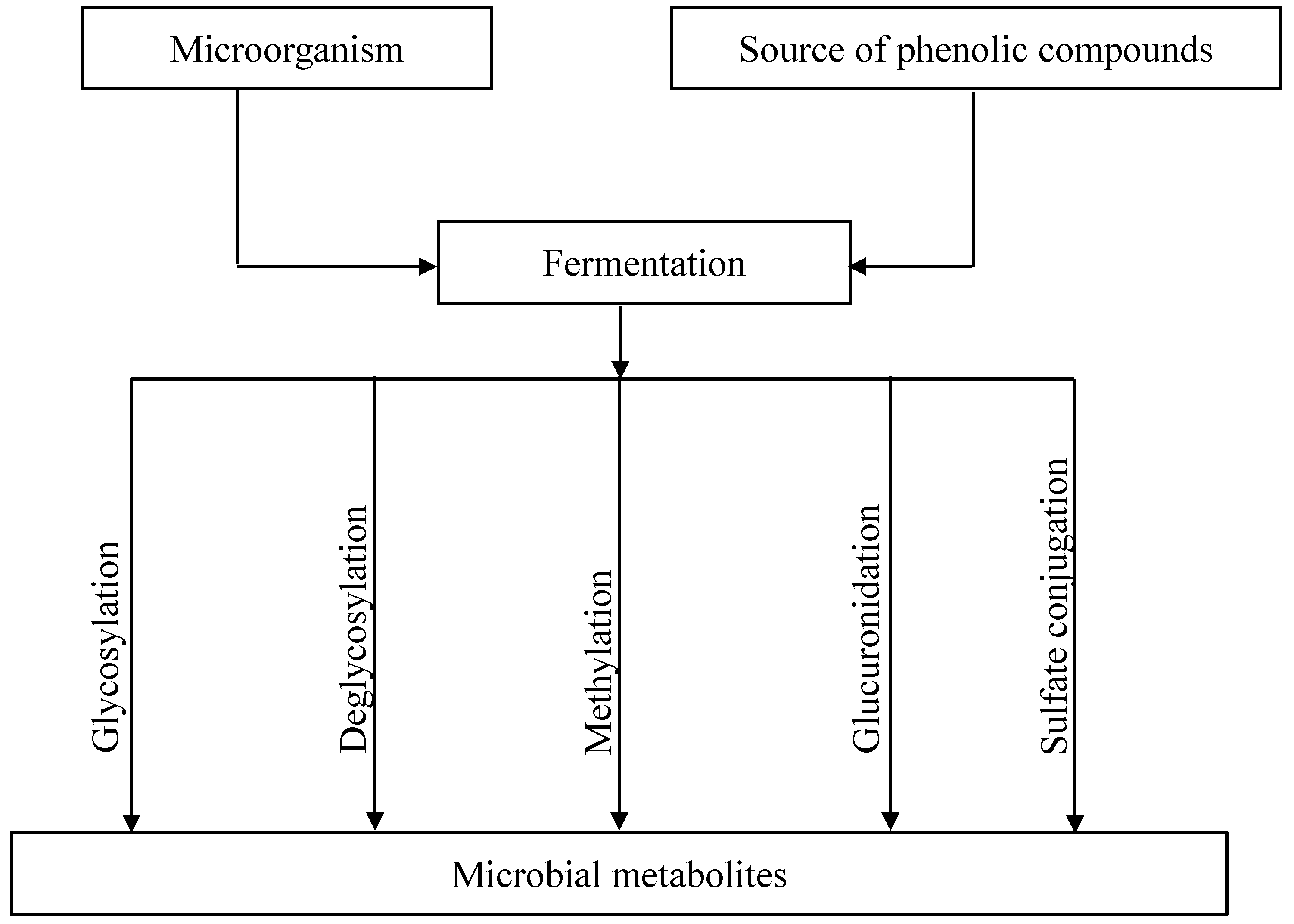
3. Microbial Metabolism of Flavonoids during Fermentation
| Microorganisms | Species | Enzymes | References |
|---|---|---|---|
| Bacteria | Lactobacillus lactis | Esterase, decarboxylase | [46] |
| Lactobacillus plantarum | β-Glucosidase, decarboxylase | [46] | |
| Lactobacillus rhamnosus | Cellulase, esterase, β-glucosidase | [46,47] | |
| Bacillus cereus | Cellulase, tannase | [46] | |
| Bacillus subtilis | Cellulase, β-glucanase | [46] | |
| Bacillus thuringiensis | Cellulase, tannase | [46] | |
| Fungi | Aspergillus awamori nakazawa | Xylanase, α-l-arabinofuranosidase, feruloyl esterase | [48,49] |
| Aspergillus niger | Cellulase, esterase, β-glucosidase, xylanase | [46,48,49,50] | |
| Aspergillus oryzae | Cellulase, β-glucosidase, xylanase, pectinase | [51] | |
| Lentinus edodes | Cellulase, β-glucosidase, xylanase, manganese peroxidase, laccase | [52,53] | |
| Penicillium brasilianum | Feruoylesterase | [54] | |
| Pleurotus ostreatus | Laccase, α-/β-glucosidase | [55,56] | |
| Rhizopus oligosporus | β-glucosidase, β-glucuronidase, xylanase | [57,58,59] | |
| Phanerochaete chrysosporium | β-Glucosidase, lignin peroxidases, manganese peroxidase, laccase | [60] | |
| Rhizopus oryzae | β-glucosidase, tannase, pectinase | [24] | |
| Yeast | Crytococcus flavus | β-glucosidase, β-glucanase, esterase, xylanase | [46] |
| Rhodotorula glutimis | β-Glucosidase | [46] | |
| Sacharomyces cerevisiae | β-Glucosidase, feruoylesterase | [46,61] | |
| Wickerhamomyces anomalus | β-Glucosidase, esterase | [62] |
| Substrate | Production | Microorganism | Reference |
|---|---|---|---|
| Glycosylation | |||
| Quercetin | Isoquercetin (quercetin-3-glucoside) | Bacillus cereus | [63] |
| Catechin | Catechin 7-α-d-glucopyranoside | Bacillus stearothermophilus | [64] |
| Catechin 5-α-d-glucopyranoside | |||
| Luteolin | Luteolin-3'-O-α-d-glucopyranoside | Leuconostoc mesenteroides | [65] |
| Luteolin-4'-O-α-d-glucopyranoside | |||
| Catechin | Catechin-4'-β-d-fucopyranoside | Aspergillus niger | [64] |
| Kaempferol | Kaempferol 3-β-O-glucopyranoside | Cunninghamella blakesleeana | [66] |
| Kaempferol 4'-O-α-L-rhamnopyranoside | |||
| Kaempferol | Kaempferol 3-β-O-glucopyranoside | Cunninghamella echinulata | [67] |
| Flavonol | Flavonol 3-β-O-glucopyranoside | Cunninghamella echinulata | [67] |
| Quercetin | Quercetin 3-O-β-d-glucopyranoside | Cunninghamella elegans | [68] |
| Quercetin | Quercetin glycoside | Penicillium decumbens | [69] |
| Kaempferol | Kaempferol glycoside | Penicillium decumbens | [69] |
| Isorhamnetin | Isorhamnetin glycoside | Penicillium decumbens | [69] |
| Deglycosylation | |||
| Daidzin | Daidzein | Bacillus pumilus | [31] |
| Daidzin | Daidzein | Bacillus subtilis | [33] |
| Kaempferol-3-O-glucoside | Kaempferol | Bifidobacterium pseudocatenulatum | [70] |
| Naringin | Prunin | Clostridium stercorarium | [71] |
| Quercetin-glucoside | Quercetin | Lactobacillus plantarum | [72] |
| Ploridzin | Phloretin | Lactobacillus plantarum | [72] |
| Kaempferol-3-rutinoside | Kaempferol, kaempferol-3-glucoside | Aspergillus awamori | [17] |
| Rutin | Quercetin, quercetin-3-O-glucoside | Aspergillus awamori | [17] |
| Rutin | Quercetin, quercetin-3-O-glucoside | Aspergillus niger | [73] |
| Daidzin, glycitin, genistin | Daidzein, glycitein, genistein | Aspergillus oryzae | [15] |
| Narigin | Naringenin | Curvularia lunata | [74] |
| Ring cleavage | |||
| Quercetin | 2-Protocatechuoylphloroglucinol carboxylic acid | Aspergillus flavus | [2] |
| Flavanone | 2'-Hydroxydibenzoylmethane | Aspergillus niger | [2] |
| Quercetin | 2-Protocatechuoylphloroglucinol carboxylic acid | Asperillus niger | [75] |
| Flavanone | 2'-Hydroxychalcone; 2',4-hydroxydihydrochalcone | Gibberella fujikuroi | [76] |
| 2,4-Dihydroxychalcone | |||
| Flavanone | 2',3'',4''-Trihydroxydihydrochalcone | Penicillium chrysogenum | [2] |
| 2'-Hydroxydihydrochalcone | |||
| Methylation | |||
| Quercetin | 3'-O-methylquercetin | Beauveria sp. | [77] |
| Quercetin | Methylquercetin | Beauveria bassiana | [78] |
| Rutin | Methylrutin | Cunninghamella echinulata | [78] |
| Quercetin 3-O-β-d-glucopyranoside | Isorhamnetin 3-O-β-d-glucopyranoside | Cunninghamella elegans | [68] |
| 7-Hydroxyflavanone | 7-Methoxyflavanone | Penicillium chermesinum | [79] |
| 3',4'-Dihydroxy-7-methoxyflavanone | |||
| Glucuronidation | |||
| Quercetin | Quercetin glucuronide | Beauveria bassiana | [78] |
| Rutin | Rutin glucuronide | Cunninghamella echinulata | [78] |
| Quercetin | Quercetin-4'-O-β-d-glucuronide | Streptomyces sp. | [20] |
| Quercetin-3'-O-β-d-glucuronide | |||
| Quercetin-3-O-β-d-glucuronide | |||
| Quercetin-7-O-β-d-glucuronide | |||
| Rutin | Quercetin-4'-O-β-d-glucuronide | Streptomyces sp. | [20] |
| Quercetin-3-O-β-d-glucuronide | |||
| Naringenin | Quercetin-7-O-β-d-glucuronide | ||
| Naringenin-7-O-β-d-glucuronide | Streptomyces sp. | [20] | |
| Naringenin-4'-O-β-d-glucuronide | |||
| Sulfate conjugation | |||
| Kaempferol | Kaempferol-4'-sulfate | Cunninghamella blakesleeana | [66] |
| Rutin | Rutin sulfate | Cunninghamella echinulata | [78] |
| Hesperitin | Hesperetin-7-sulfate | Mucor ramannianus | [80] |
| 5-Hydroxyflavone | 5,4'-Dihydroxyflavone-4'-sulfate | Streptomyces fulvissimus | [2] |
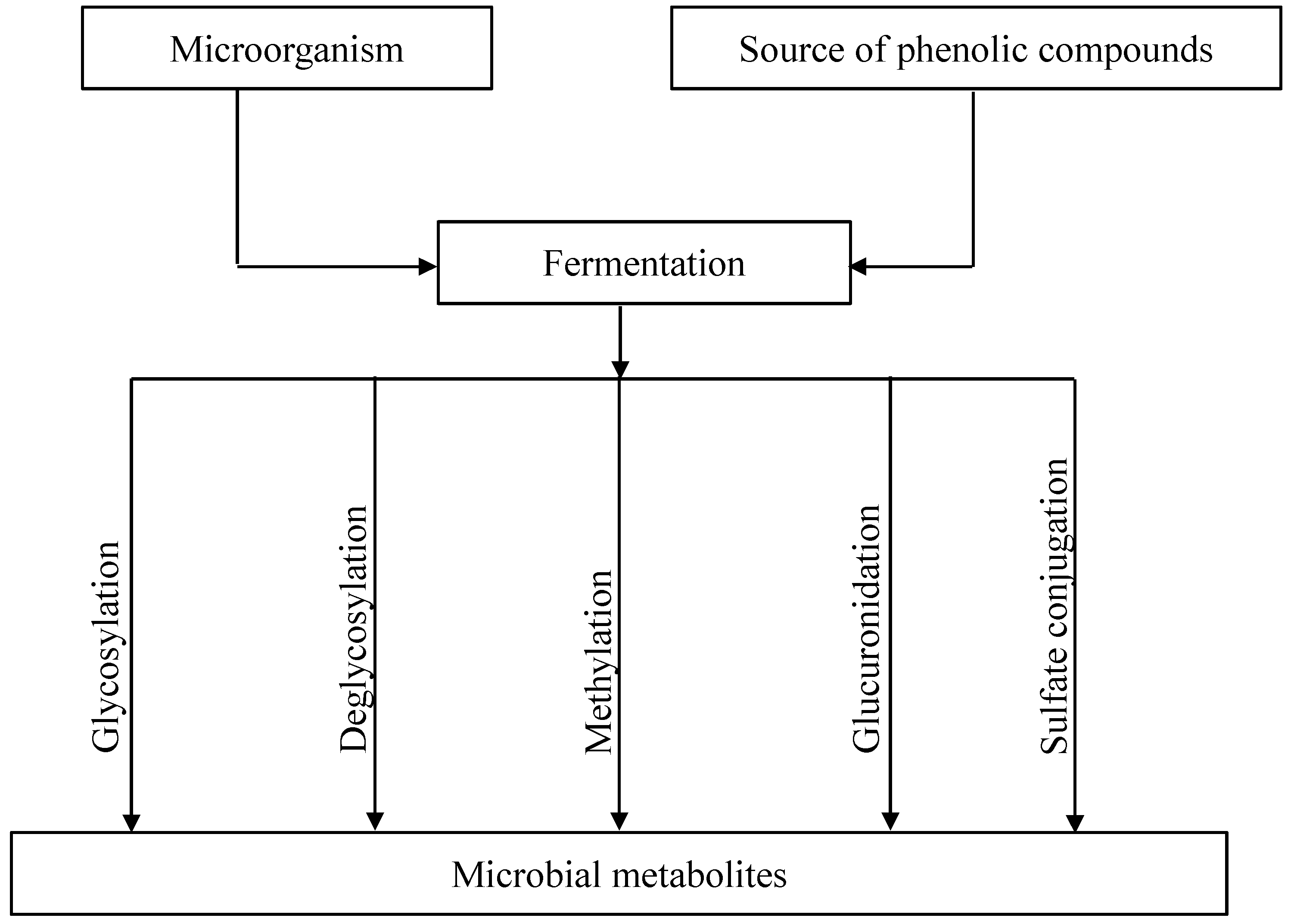
3.1. Glycosylation and Deglycosylation of Flavonoids
3.1.1. Glycosylation of Flavonoids
3.1.2. Deglycosylation of Flavonoids
3.2. Ring Cleavage of Flavonoids
3.3. Methylation of Flavonoids
3.4. Glucuronidation of Flavonoids
3.5. Sulfate Conjugation of Flavonoids
4. Perspectives
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Celep, G.S.; Rastmanesh, R.; Marotta, F. Chapter 43—Microbial metabolism of polyphenols and health. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 577–589. [Google Scholar]
- Das, S.; Rosazza, J.P.N. Microbial and enzymatic transformations of flavonoids. J. Nat. Prod. 2006, 69, 499–508. [Google Scholar]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation: A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar]
- Wijngaard, H.; Hossain, M.B.; Rai, D.K.; Brunton, N. Techniques to extract bioactive compounds from food by-products of plant origin. Food Res. Int. 2012, 46, 505–513. [Google Scholar]
- Muñiz-Márquez, D.B.; Martínez-Ávila, G.C.; Wong-Paz, J.E.; Belmares-Cerda, R.; Rodríguez-Herrera, R.; Aguilar, C.N. Ultrasound-assisted extraction of phenolic compounds from Laurus nobilis L. and their antioxidant activity. Ultrason. Sonochem. 2013, 20, 1149–1154. [Google Scholar]
- Švarc-Gajić, J.; Stojanović, Z.; Segura Carretero, A.; Arráez Román, D.; Borrás, I.; Vasiljević, I. Development of a microwave-assisted extraction for the analysis of phenolic compounds from Rosmarinus officinalis. J. Food Eng. 2013, 119, 525–532. [Google Scholar]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods: A review. Food Chem. 2014, 152, 46–55. [Google Scholar]
- Puri, M.; Sharma, D.; Barrow, C.J. Enzyme-assisted extraction of bioactives from plants. Trends Biotechnol. 2012, 30, 37–44. [Google Scholar]
- Rodriguez-Mateos, A.; Heiss, C.; Borges, G.; Crozier, A. Berry (poly)phenols and cardiovascular health. J. Agric. Food Chem. 2014, 62, 3842–3851. [Google Scholar]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar]
- Ranilla, L.G.; Kwon, Y.-I.; Apostolidis, E.; Shetty, K. Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Bioresour. Technol. 2010, 101, 4676–4689. [Google Scholar]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar]
- Chen, H.; Sang, S. Biotransformation of tea polyphenols by gut microbiota. J. Funct. Foods 2014, 7, 26–42. [Google Scholar]
- Francisco, J.P.-C.; Malen, M.-C.; Àngels, F.; Cristina, C.; Margarida, C. The effects of cocoa on the immune system. Front. Pharmacol. 2013. [Google Scholar] [CrossRef]
- Lee, S.-H.; Seo, M.-H.; Oh, D.-K. Deglycosylation of isoflavones in isoflavone-rich soy germ flour by Aspergillus oryzae KACC 40247. J. Agric. Food Chem. 2013, 61, 12101–12110. [Google Scholar]
- Braune, A.; Blaut, M. Intestinal bacterium Eubacterium cellulosolvens deglycosylates flavonoid C- and O-glucosides. Appl. Environ. Microbiol. 2012, 78, 8151–8153. [Google Scholar]
- Lin, S.; Zhu, Q.; Wen, L.; Yang, B.; Jiang, G.; Gao, H.; Chen, F.; Jiang, Y. Production of quercetin, kaempferol and their glycosidic derivatives from the aqueous-organic extracted residue of litchi pericarp with Aspergillus awamori. Food Chem. 2014, 145, 220–227. [Google Scholar]
- Ibrahim, A.R.; Abul-Hajj, Y.J. Aromatic hydroxylation and sulfation of 5-hydroxyflavone by Streptomyces fulvissimus. Appl. Environ. Microbiol. 1989, 55, 3140–3142. [Google Scholar]
- Mikell, J.R.; Khan, I.A. Bioconversion of 7-hydroxyflavanone: Isolation, vharacterization and bioactivity evaluation of twenty-one phase I and phase II microbial metabolites. Chem. Pharm. Bull. 2012, 60, 1139–1145. [Google Scholar]
- Marvalin, C.; Azerad, R. Microbial glucuronidation of polyphenols. J. Mol. Catal. B: Enzym. 2011, 73, 43–52. [Google Scholar]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar]
- Pereira, D.; Valentão, P.; Pereira, J.; Andrade, P. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar]
- Peres-Gregorio, M.R.; Regueiro, J.; Simal-Gandara, J.; Rodriguez, A.S.; Almeida, D.P.F. Increasing the added value of onions as a source of antioxidant flavonoids: A critical review. Crit. Rev. Food Sci. 2014, 54, 1050–1062. [Google Scholar]
- Yadav, G.; Singh, A.; Bhattacharya, P.; Yuvraj, J.; Banerjee, R. Comparative analysis of solid-state bioprocessing and enzymatic treatment of finger millet for mobilization of bound phenolics. Bioprocess Biosyst. Eng. 2013, 36, 1563–1569. [Google Scholar]
- Pinelo, M.; Arnous, A.; Meyer, A.S. Upgrading of grape skins: Significance of plant cell-wall structural components and extraction techniques for phenol release. Trends Food Sci. Technol. 2006, 17, 579–590. [Google Scholar]
- Cerda, A.; Martínez, M.E.; Soto, C.; Poirrier, P.; Perez-Correa, J.R.; Vergara-Salinas, J.R.; Zúñiga, M.E. The enhancement of antioxidant compounds extracted from Thymus vulgaris using enzymes and the effect of extracting solvent. Food Chem. 2013, 139, 138–143. [Google Scholar]
- Vattem, D.A.; Shetty, K. Ellagic acid production and phenolic antioxidant activity in cranberry pomace (Vaccinium macrocarpon) mediated by Lentinus edodes using a solid-state system. Process Biochem. 2003, 39, 367–379. [Google Scholar]
- Vattem, D.A.; Lin, Y.T.; Labbe, R.G.; Shetty, K. Phenolic antioxidant mobilization in cranberry pomace by solid-state bioprocessing using food grade fungus Lentinus edodes and effect on antimicrobial activity against select food borne pathogens. Innov. Food Sci. Emerg. Technol. 2004, 5, 81–91. [Google Scholar]
- Cai, S.; Wang, O.; Wu, W.; Zhu, S.; Zhou, F.; Ji, B.; Gao, F.; Zhang, D.; Liu, J.; Cheng, Q. Comparative study of the effects of solid-state fermentation with three filamentous fungi on the total phenolics content (TPC), flavonoids, and antioxidant activities of subfractions from oats (Avena sativa L.). J. Agric. Food. Chem. 2011, 60, 507–513. [Google Scholar]
- Schmidt, C.G.; Gonçalves, L.M.; Prietto, L.; Hackbart, H.S.; Furlong, E.B. Antioxidant activity and enzyme inhibition of phenolic acids from fermented rice bran with fungus Rizhopus oryzae. Food Chem. 2014, 146, 371–377. [Google Scholar]
- Cho, K.M.; Hong, S.Y.; Math, R.K.; Lee, J.H.; Kambiranda, D.M.; Kim, J.M.; Islam, S.M.A.; Yun, M.G.; Cho, J.J.; Lim, W.J.; et al. Biotransformation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chem. 2009, 114, 413–419. [Google Scholar]
- Chung, I.-M.; Seo, S.-H.; Ahn, J.-K.; Kim, S.-H. Effect of processing, fermentation, and aging treatment to content and profile of phenolic compounds in soybean seed, soy curd and soy paste. Food Chem. 2011, 127, 960–967. [Google Scholar]
- Cho, K.M.; Lee, J.H.; Yun, H.D.; Ahn, B.Y.; Kim, H.; Seo, W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Comp. Anal. 2011, 24, 402–410. [Google Scholar]
- Shin, E.-C.; Lee, J.; Hwang, C.; Lee, B.; Kim, H.; Ko, J.; Baek, I.-Y.; Shin, J.; Nam, S.; Seo, W.; et al. Enhancement of total phenolic and isoflavone-aglycone contents and antioxidant activities during Cheonggukjang fermentation of brown soybeans by the potential probiotic Bacillus subtilis CSY191. Food Sci. Biotechnol. 2014, 23, 531–538. [Google Scholar]
- Ankolekar, C.; Johnson, K.; Pinto, M.; Johnson, D.; Labbe, R.G.; Greene, D.; Shetty, K. Fermentation of whole apple juice using lactobacillus acidophilus for potential dietary management of hyperglycemia, hypertension, and modulation of beneficial bacterial responses. J. Food Biochem. 2012, 36, 718–738. [Google Scholar]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved bioavailability of dietary phenolic acids in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L): Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agric. 2005, 85, 297–304. [Google Scholar]
- Pyo, Y.-H.; Lee, T.-C.; Lee, Y.-C. Effect of lactic acid fermentation on enrichment of antioxidant properties and bioactive isoflavones in soybean. J. Food Sci. 2005, 70, S215–S220. [Google Scholar]
- Moore, J.; Cheng, Z.; Hao, J.; Guo, G.; Liu, J.-G.; Lin, C.; Yu, L. Effects of solid-state yeast treatment on the antioxidant properties and protein and fiber compositions of common hard wheat bran. J. Agric. Food Chem. 2007, 55, 10173–10182. [Google Scholar]
- Da Silva, L.H.; Celeghini, R.M.S.; Chang, Y.K. Effect of the fermentation of whole soybean flour on the conversion of isoflavones from glycosides to aglycones. Food Chem. 2011, 128, 640–644. [Google Scholar]
- Handa, C.L.; Couto, U.R.; Vicensoti, A.H.; Georgetti, S.R.; Ida, E.I. Optimisation of soy flour fermentation parameters to produce β-glucosidase for bioconversion into aglycones. Food Chem. 2014, 152, 56–65. [Google Scholar]
- Hwan Nam, D.; Jung Kim, H.; Sun Lim, J.; Heon Kim, K.; Park, C.-S.; Hwan Kim, J.; Lim, J.; Young Kwon, D.; Kim, I.-H.; Kim, J.-S. Simultaneous enhancement of free isoflavone content and antioxidant potential of soybean by fermentation with Aspergillus oryzae. J. Food Sci. 2011, 76, H194–H200. [Google Scholar]
- Kim, M.J.; John, K.M.M.; Choi, J.N.; Lee, S.; Kim, A.J.; Kim, Y.M.; Lee, C.H. Changes in secondary metabolites of green tea during fermentation by Aspergillus oryzae and its effect on antioxidant potential. Food Res. Int. 2013, 53, 670–677. [Google Scholar]
- Cheng, K.-C.; Wu, J.-Y.; Lin, J.-T.; Liu, W.-H. Enhancements of isoflavone aglycones, total phenolic content, and antioxidant activity of black soybean by solid-state fermentation with Rhizopus spp. Eur. Food Res. Technol. 2013, 236, 1107–1113. [Google Scholar]
- Perez-Gregorio, M.R.; Regueiro, J.; Alonso-Gonzalez, E.; Pastrana-Castro, L.M.; Simal-Gandara, J. Influence of alcoholic fermentation process on antioxidant activity and phenolic levels from mulberies (Morus nigra L.). LWT-Food Sci. Technol. 2011, 44, 1793–1801. [Google Scholar]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar]
- Marazza, J.A.; Garro, M.S.; Savoy de Giori, G. Aglycone production by Lactobacillus rhamnosus CRL981 during soymilk fermentation. Food Microbiol. 2009, 26, 333–339. [Google Scholar]
- Georgetti, S.R.; Vicentini, F.T.M.C.; Yokoyama, C.Y.; Borin, M.F.; Spadaro, A.C.C.; Fonseca, M.J.V. Enhanced in vitro and in vivo antioxidant activity and mobilization of free phenolic compounds of soybean flour fermented with different β-glucosidase-producing fungi. J. Appl. Microbiol. 2009, 106, 459–466. [Google Scholar]
- Bhanja, T.; Kumari, A.; Banerjee, R. Enrichment of phenolics and free radical scavenging property of wheat koji prepared with two filamentous fungi. Bioresour. Technol. 2009, 100, 2861–2866. [Google Scholar]
- Liu, C.; Sun, Z.-T.; Du, J.-H.; Wang, J. Response surface optimization of fermentation conditions for producing xylanase by Aspergillus niger SL-05. J. Ind. Microbiol. Biotechnol. 2008, 35, 703–711. [Google Scholar]
- Wu, Z.; Song, L.; Huang, D. Food grade fungal stress on germinating peanut seeds induced phytoalexins and enhanced polyphenolic antioxidants. J. Agric. Food Chem. 2011, 59, 5993–6003. [Google Scholar]
- Zheng, Z.; Shetty, K. Solid-state bioconversion of phenolics from cranberry pomace and role of Lentinus edodes β-glucosidase. J. Agric. Food Chem. 2000, 48, 895–900. [Google Scholar]
- Hatvani, N.; Mécs, I. Production of laccase and manganese peroxidase by Lentinus edodes on malt-containing by-product of the brewing process. Process Biochem. 2001, 37, 491–496. [Google Scholar]
- Panagiotou, G.; Olavarria, R.; Olsson, L. Penicillium brasilianum as an enzyme factory; the essential role of feruloyl esterases for the hydrolysis of the plant cell wall. J. Biotechnol. 2007, 130, 219–228. [Google Scholar]
- Żuchowski, J.; Pecio, Ł.; Jaszek, M.; Stochmal, A. Solid-state fermentation of rapeseed meal with the white-rot fungi Trametes versicolor and Pleurotus ostreatus. Appl. Biochem. Biotechnol. 2013, 171, 2075–2081. [Google Scholar]
- Tlecuitl-Beristain, S.; Sánchez, C.; Loera, O.; Robson, G.D.; Díaz-Godínez, G. Laccases of Pleurotus ostreatus observed at different phases of its growth in submerged fermentation: Production of a novel laccase isoform. Mycol. Res. 2008, 112, 1080–1084. [Google Scholar]
- Correia, R.T.P.; McCue, P.; MagalhÃEs, M.M.A.; MacÊDo, G.R.; Shetty, K. Phenolic antioxidant enrichment of soy flour-supplemented guava waste by Rhizopus oligosporus-mediated solid-state bioprocessing. J. Food Biochem. 2004, 28, 404–418. [Google Scholar]
- Vattem, D.A.; Lin, Y.T.; Labbe, R.G.; Shetty, K. Antimicrobial activity against select food-borne pathogens by phenolic antioxidants enriched in cranberry pomace by solid-state bioprocessing using the food grade fungus Rhizopus oligosporus. Process Biochem. 2004, 39, 1939–1946. [Google Scholar]
- McCue, P.; Horii, A.; Shetty, K. Solid-state bioconversion of phenolic antioxidants from defatted soybean powders by Rhizopus oligosporus: Role of carbohydrate-cleaving enzymes. J. Food Biochem. 2003, 27, 501–514. [Google Scholar]
- Ajila, C.M.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Solid-state fermentation of apple pomace using Phanerocheate chrysosporium—Liberation and extraction of phenolic antioxidants. Food Chem. 2011, 126, 1071–1080. [Google Scholar]
- Coghe, S.; Benoot, K.; Delvaux, F.; Vanderhaegen, B.; Delvaux, F.R. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 602–608. [Google Scholar]
- Restuccia, C.; Muccilli, S.; Palmeri, R.; Randazzo, C.L.; Caggia, C.; Spagna, G. An alkaline β-glucosidase isolated from an olive brine strain of Wickerhamomyces anomalus. FEMS Yeast Res. 2011, 11, 487–493. [Google Scholar]
- Rao, K.V.; Weisner, N.T. Microbial transformation of suercetin by Bacillus cereus. Appl. Environ. Microbiol. 1981, 42, 450–452. [Google Scholar]
- Gao, C.; Mayon, P.; MacManus, D.A.; Vulfson, E.N. Novel enzymatic approach to the synthesis of flavonoid glycosides and their esters. Biotechnol. Bioeng. 2000, 71, 235–243. [Google Scholar]
- Bertrand, A.; Morel, S.; Lefoulon, F.; Rolland, Y.; Monsan, P.; Remaud-Simeon, M. Leuconostoc mesenteroides glucansucrase synthesis of flavonoid glucosides by acceptor reactions in aqueous-organic solvents. Carbohydr. Res. 2006, 341, 855–863. [Google Scholar]
- Ibrahim, A.; Khalifa, S.I.; Khafagi, I.; Youssef, D.T.; Khan, S.; Mesbah, M.; Khan, I. Microbial metabolism of biologically active secondary metabolites from Nerium oleander L. Chem. Pharm. Bull. 2008, 56, 1253–1258. [Google Scholar]
- Miyakoshi, S.; Azami, S.; Kuzuyama, T. Microbial glucosylation of flavonols by Cunninghamella echinulata. J. Biosci. Bioeng. 2010, 110, 320–321. [Google Scholar]
- Zi, J.; Valiente, J.; Zeng, J.; Zhan, J. Metabolism of quercetin by Cunninghamella elegans ATCC 9245. J. Biosci. Bioeng. 2011, 112, 360–362. [Google Scholar]
- Chen, S.; Xing, X.-H.; Huang, J.-J.; Xu, M.-S. Enzyme-assisted extraction of flavonoids from Ginkgo biloba leaves: Improvement effect of flavonol transglycosylation catalyzed by Penicillium decumbens cellulase. Enzym. Microb. Technol. 2011, 48, 100–105. [Google Scholar]
- Di Gioia, D.; Strahsburger, E.; Lopez de Lacey, A.M.; Bregola, V.; Marotti, I.; Aloisio, I.; Biavati, B.; Dinelli, G. Flavonoid bioconversion in Bifidobacterium pseudocatenulatum B7003: A potential probiotic strain for functional food development. J. Funct. Food 2014, 7, 671–679. [Google Scholar]
- Kaur, A.; Singh, S.; Singh, R.S.; Schwarz, W.H.; Puri, M. Hydrolysis of citrus peel naringin by recombinant α-l-rhamnosidase from Clostridium stercorarium. J. Chem. Technol. Biotechnol. 2010, 85, 1419–1422. [Google Scholar]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar]
- You, H.J.; Ahn, H.J.; Ji, G.E. Transformation of rutin to antiproliferative quercetin-3-glucoside by Aspergillus niger. J. Agric. Food Chem. 2010, 58, 10886–10892. [Google Scholar]
- Liu, J.-Y.; Yu, H.-S.; Feng, B.; Kang, L.-P.; Pang, X.; Xiong, C.-Q.; Zhao, Y.; Li, C.-M.; Zhang, Y.; Ma, B.-P. Selective hydrolysis of flavonoid glycosides by Curvularia lunata. Chin. J. Nat. Med. 2013, 11, 684–689. [Google Scholar]
- Hund, H.-K.; Breuer, J.; Lingens, F.; Hüttermann, J.; Kappl, R.; Fetzner, S. Flavonol 2,4-dioxygenase from Aspergillus niger DSM 821, a type 2 CuII-containing glycoprotein. Eur. J. Biochem. 1999, 263, 871–878. [Google Scholar]
- Udupa, S.R.; Banerji, A.; Chadha, M.S. Microbiological transformations of flavonoids—II: Transformations of (±) flavanone. Tetrahedron 1969, 25, 5415–5419. [Google Scholar]
- De, M.B.; Costa, E.M.; Pimenta, F.C.; Luz, W.C.; de Oliveira, V. Selection of filamentous fungi of the Beauveria genus able to metabolize quercetin like mammalian cells. Braz. J. Microbiol. 2008, 39, 405–408. [Google Scholar]
- Araújo, K.C.F.; de M. B. Costa, E.M.; Pazini, F.; Valadares, M.C.; de Oliveira, V. Bioconversion of quercetin and rutin and the cytotoxicity activities of the transformed products. Food Chem. Toxicol. 2013, 51, 93–96. [Google Scholar]
- Edyta, K.-S.; Tomasz, J. Microbial transformations of 7-hydroxyflavanone. Sci. World J. 2012, 254929:1–254929:8. [Google Scholar]
- Herath, W.; Khan, I.A. Microbial metabolism. Part 13: Metabolites of hesperetin. Bioorg. Med. Chem. Lett. 2011, 21, 5784–5786. [Google Scholar]
- Ma, B.; Zeng, J.; Shao, L.; Zhan, J. Efficient bioconversion of quercetin into a novel glycoside by Streptomyces rimosus subsp. rimosus ATCC 10970. J. Biosci. Bioeng. 2013, 115, 24–26. [Google Scholar]
- Slana, M.; Žigon, D.; Makovec, T.; Lenasi, H. The response of filamentous fungus Rhizopus nigricans to flavonoids. J. Basic Microbiol. 2011, 51, 433–441. [Google Scholar]
- Park, J.S.; Rho, H.S.; Kim, D.H.; Chang, I.S. Enzymatic preparation of kaempferol from green tea seed and its antioxidant activity. J. Agric. Food Chem. 2006, 54, 2951–2956. [Google Scholar]
- Turner, C.; Turner, P.; Jacobson, G.; Almgren, K.; Waldeback, M.; Sjoberg, P.; Karlsson, E.N.; Markides, K.E. Subcritical water extraction and [small beta]-glucosidase-catalyzed hydrolysis of quercetin glycosides in onion waste. Green Chem. 2006, 8, 949–959. [Google Scholar]
- Yadav, V.; Yadav, P.K.; Yadav, S.; Yadav, K.D.S. α-l-Rhamnosidase: A review. Process Biochem. 2010, 45, 1226–1235. [Google Scholar]
- Avila, M.; Jaquet, M.; Moine, D.; Requena, T.; Pelaez, C.; Arigoni, F.; Jankovic, I. Physiological and biochemical characterization of the two α-l-rhamnosidases of Lactobacillus plantarum NCC245. Microbiology 2009, 155, 2739–2749. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. Int. J. Mol. Sci. 2014, 15, 19369-19388. https://doi.org/10.3390/ijms151119369
Huynh NT, Van Camp J, Smagghe G, Raes K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. International Journal of Molecular Sciences. 2014; 15(11):19369-19388. https://doi.org/10.3390/ijms151119369
Chicago/Turabian StyleHuynh, Nguyen Thai, John Van Camp, Guy Smagghe, and Katleen Raes. 2014. "Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review" International Journal of Molecular Sciences 15, no. 11: 19369-19388. https://doi.org/10.3390/ijms151119369
APA StyleHuynh, N. T., Van Camp, J., Smagghe, G., & Raes, K. (2014). Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. International Journal of Molecular Sciences, 15(11), 19369-19388. https://doi.org/10.3390/ijms151119369