Antioxidant and Immunoregulatory Activity of Polysaccharides from Quinoa (Chenopodium quinoa Willd.)



Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction and Purification of Polysaccharide
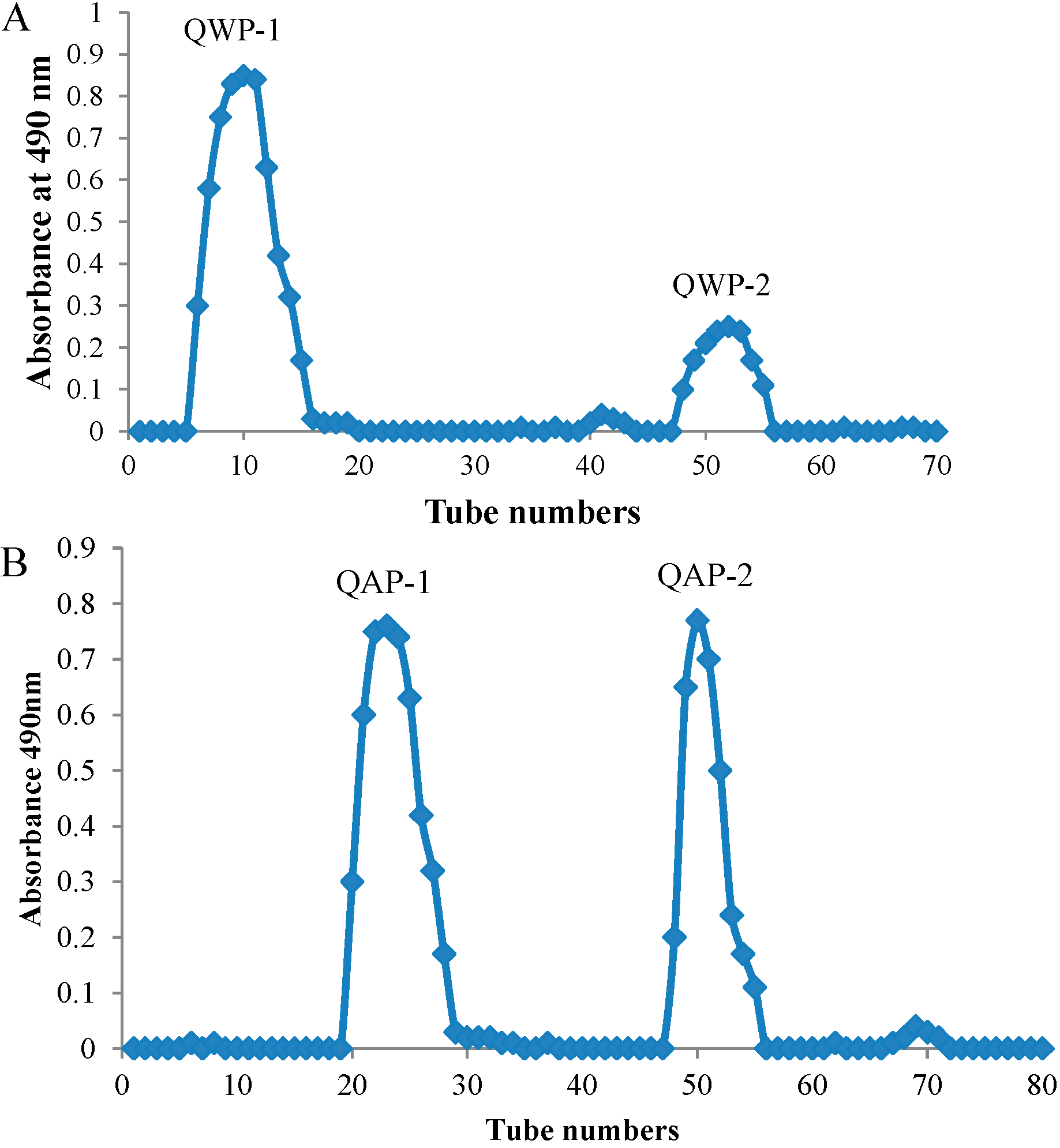
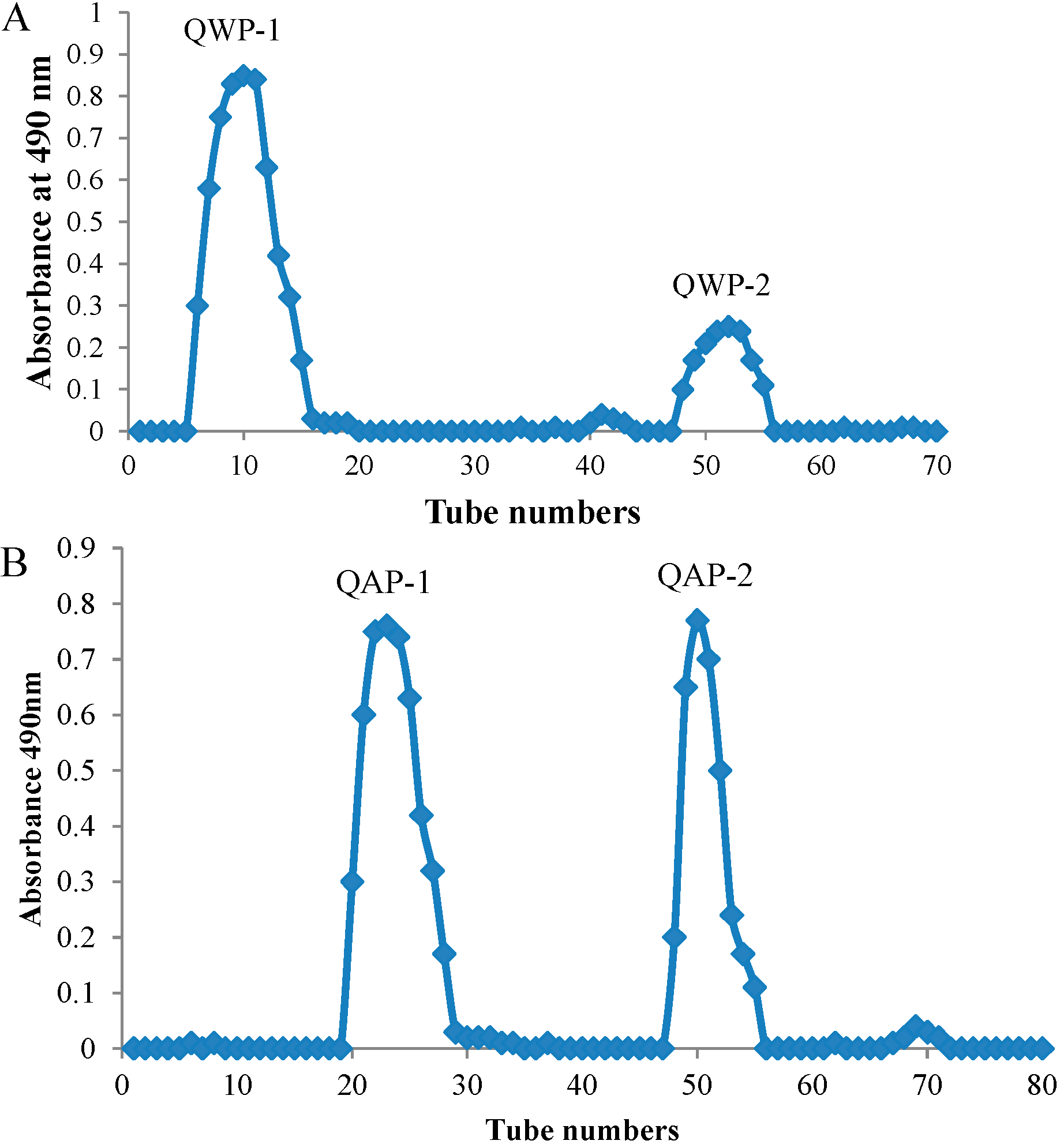
2.2. Molecular Weight and Monosaccharide Composition
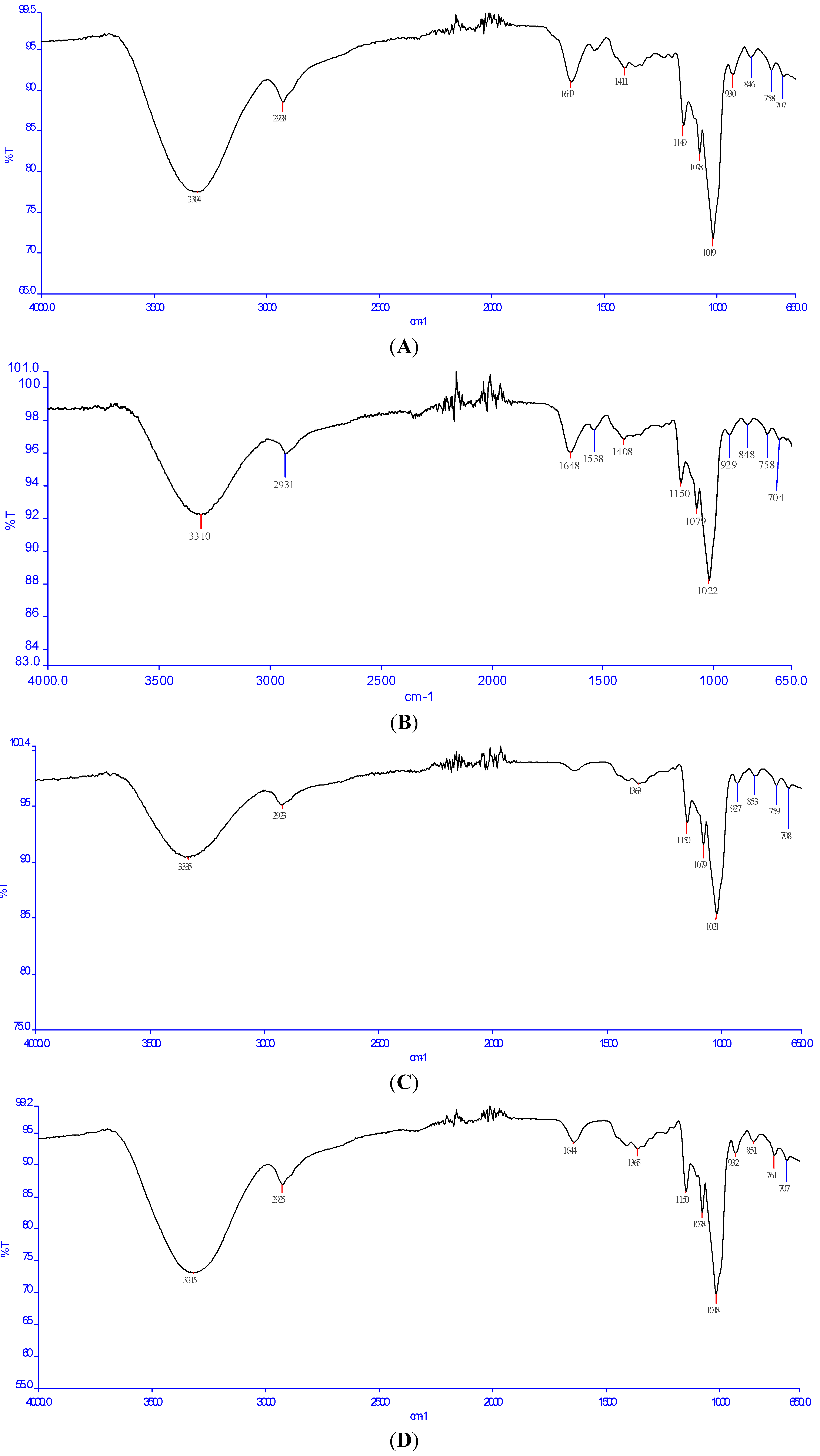
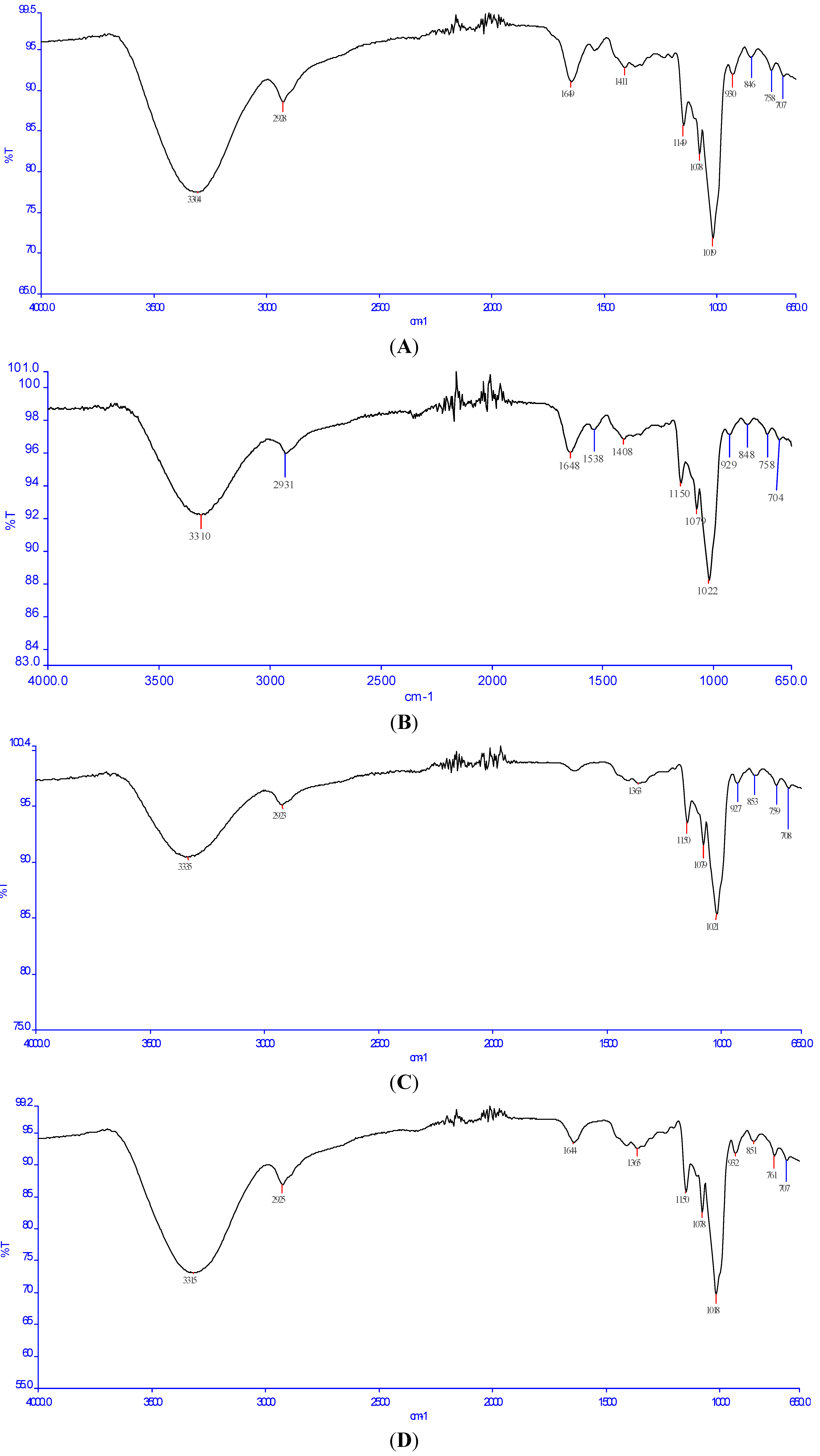
2.3. Fourier Transform Infrared (FT-IR) Analysis
2.4. Antioxidant Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | DPPH | ORAC |
|---|---|---|
| QWP | 4.76 ± 0.16 b | 14.62 ± 0.84 c |
| QAP | 4.24 ± 0.28 b | 14.66 ± 0.42 c |
| QWP-1 | 4.78 ± 1.03 b | 27.19 ± 1.74 b |
| QWP-2 | 4.94 ± 0.01 b | 40.90 ± 0.54 a |
| QAP-1 | 6.61 ± 0.93 a | 29.22 ± 2.33 b |
| QAP-2 | 5.78 ± 0.50 a | 24.86 ± 2.63 b |
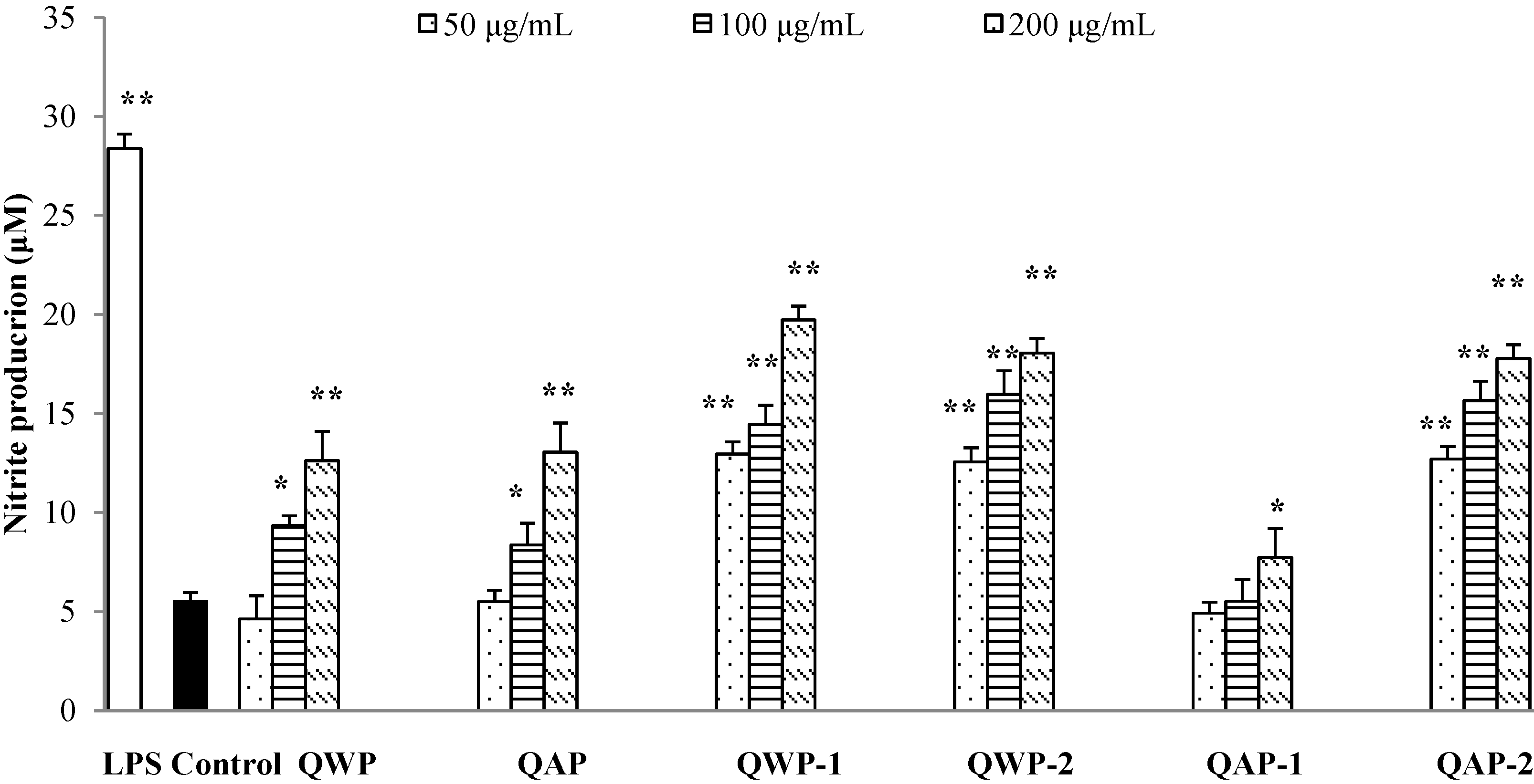
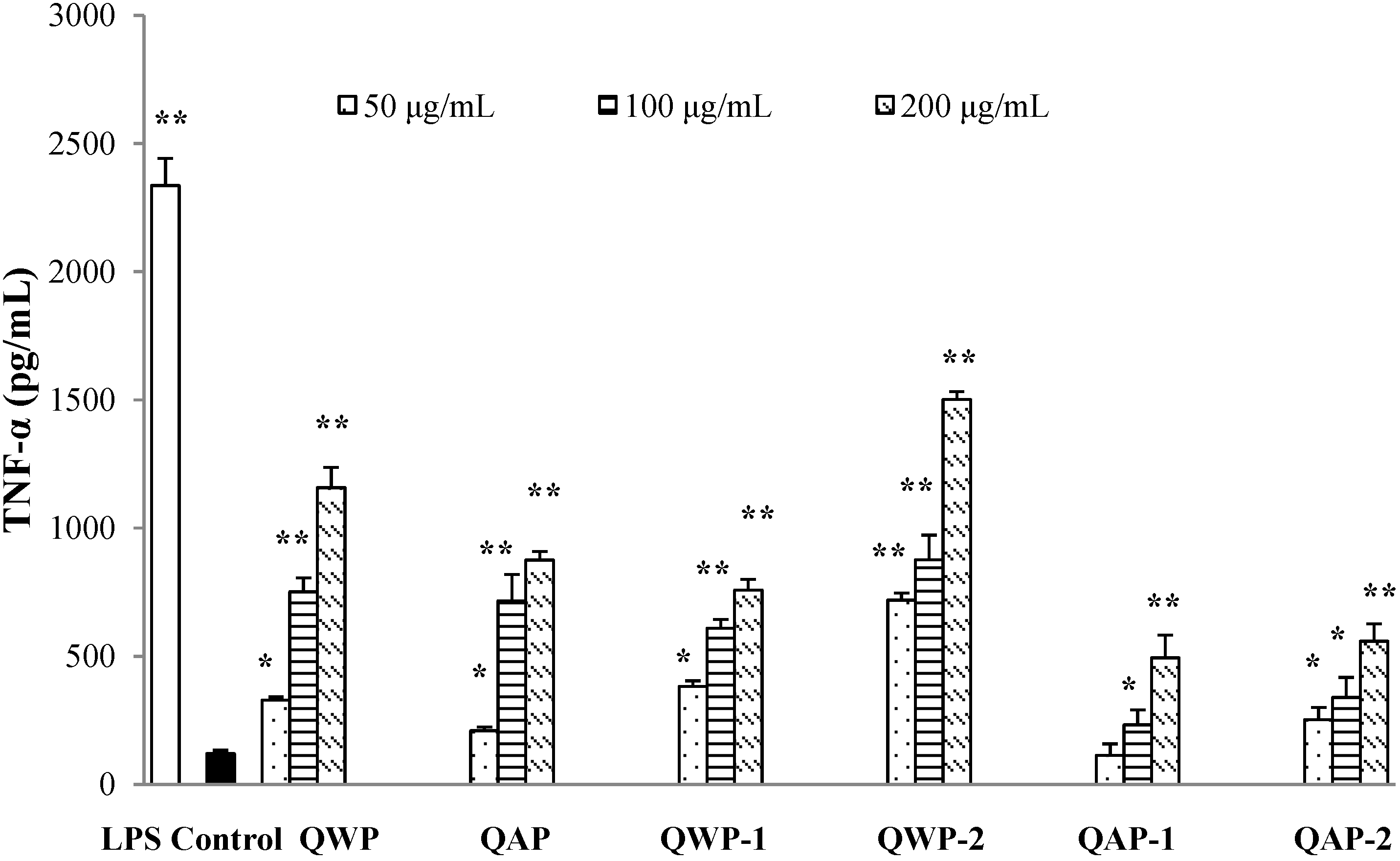
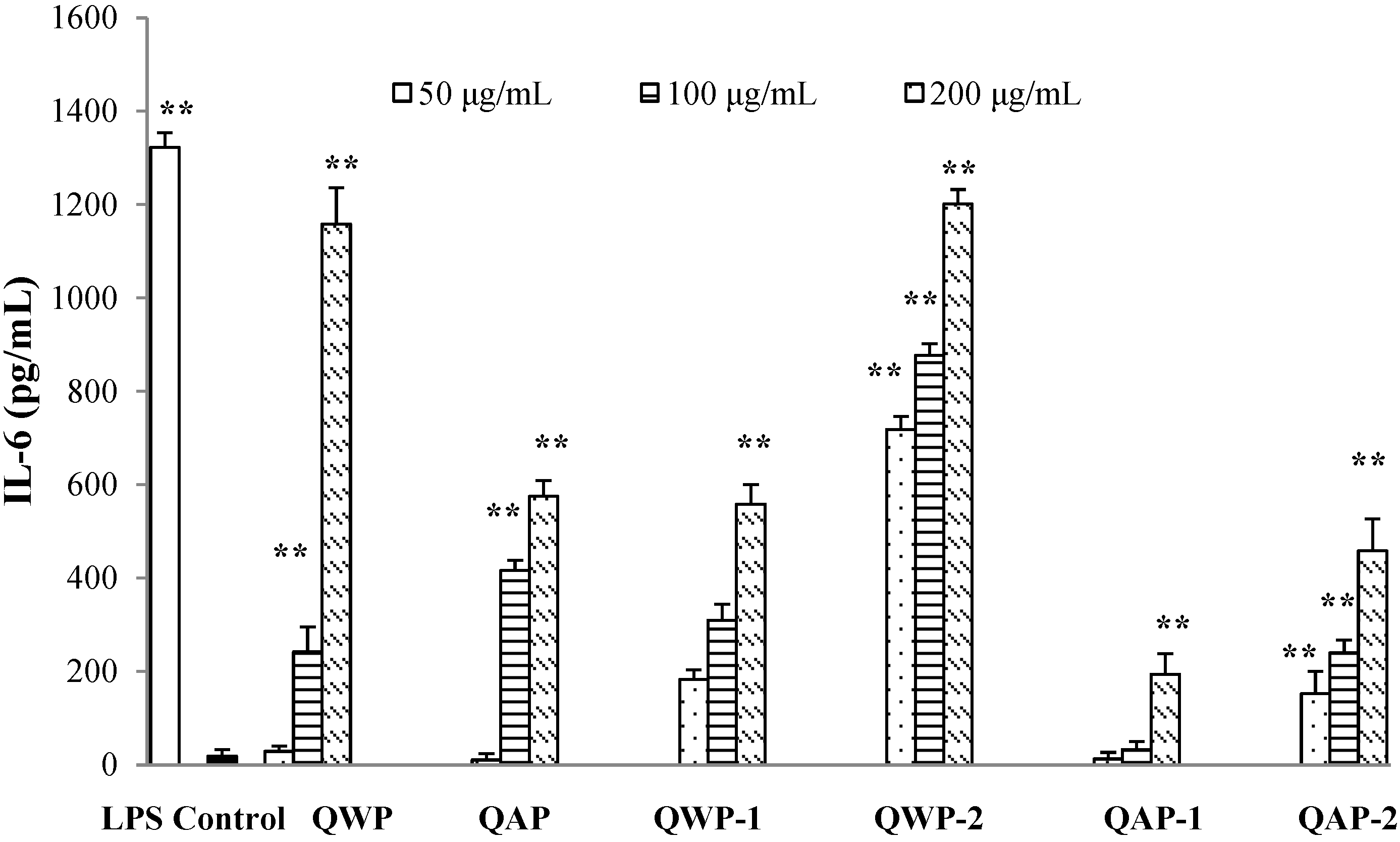
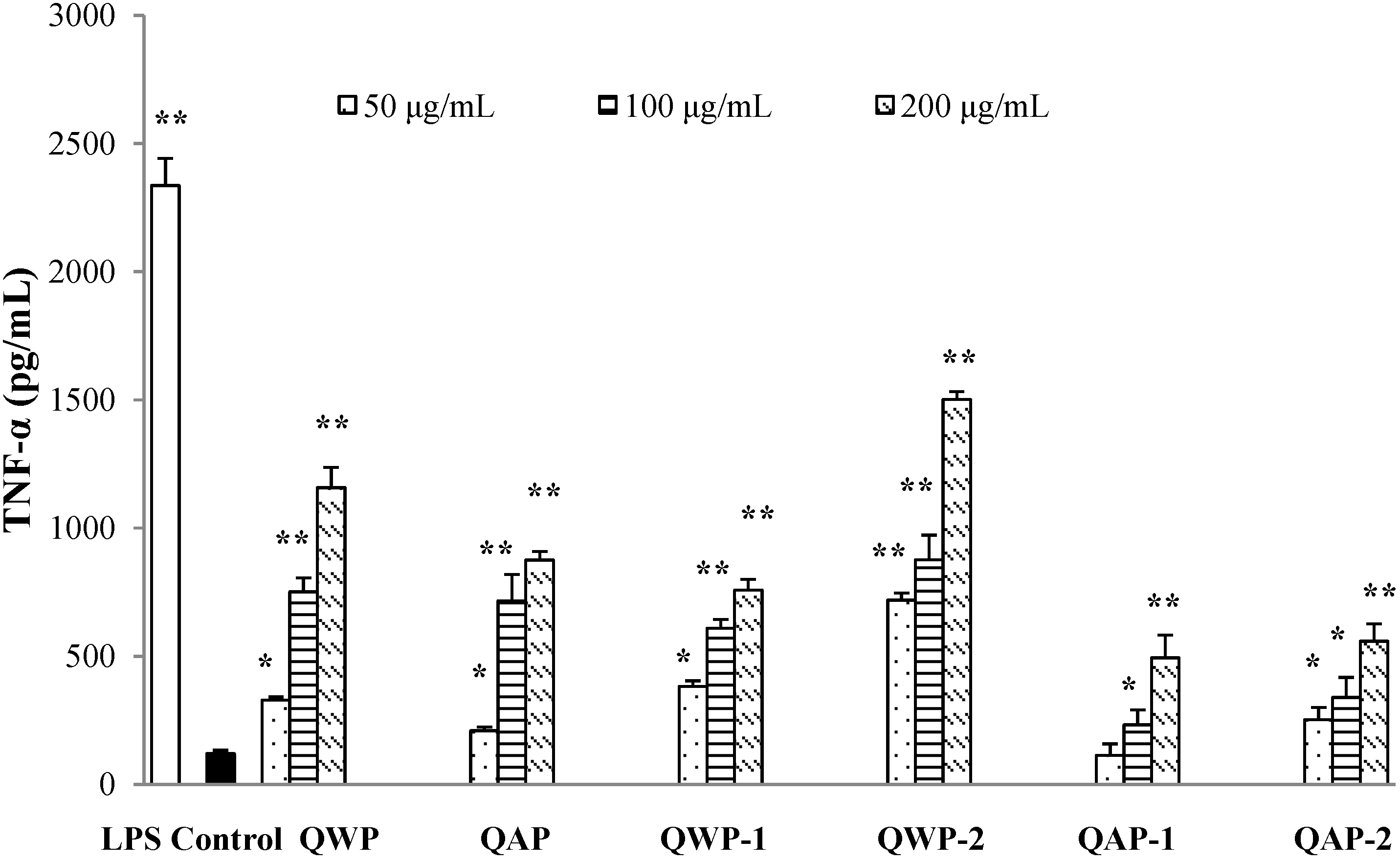
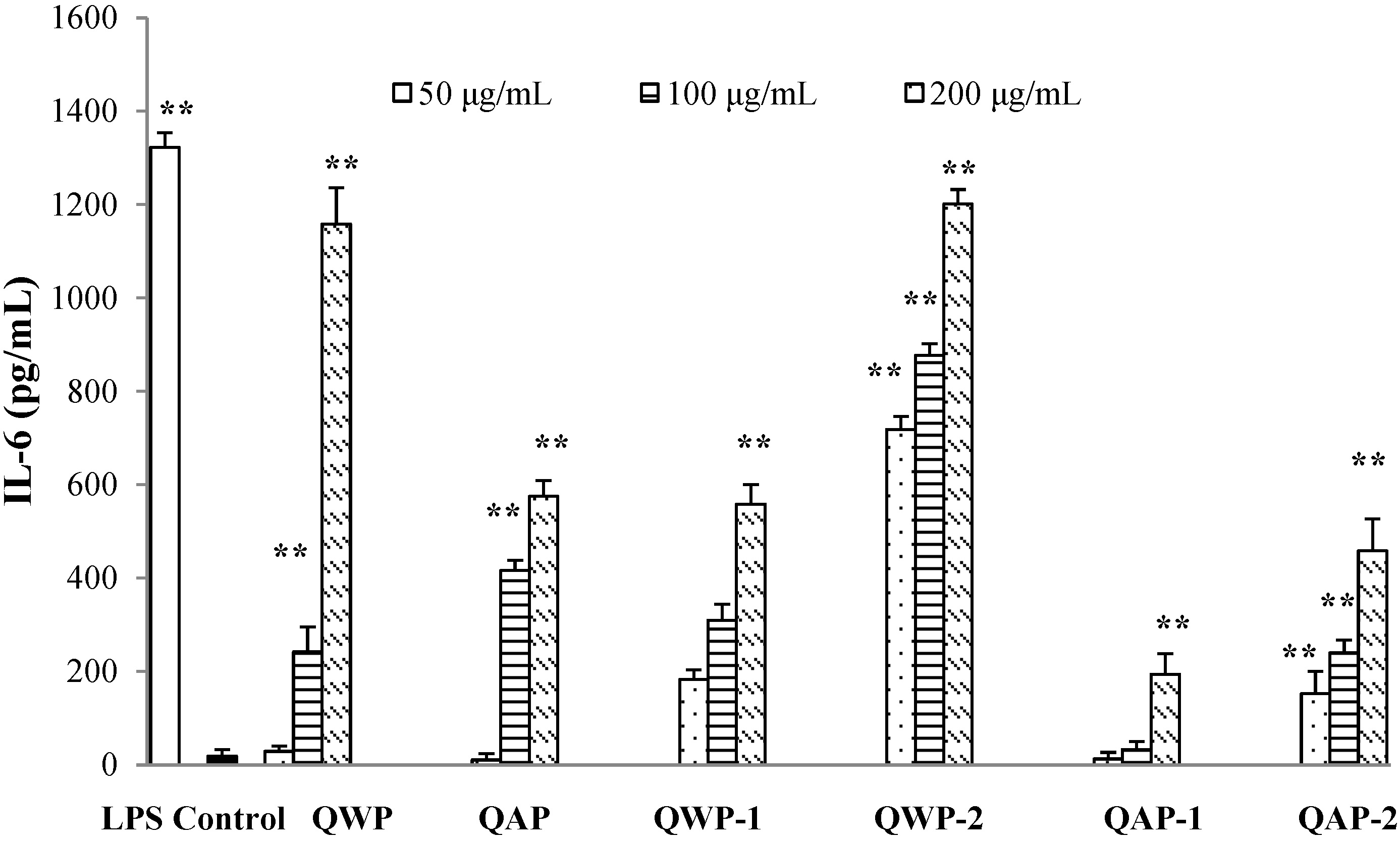
2.5. Immunomodulatory Activity Analysis
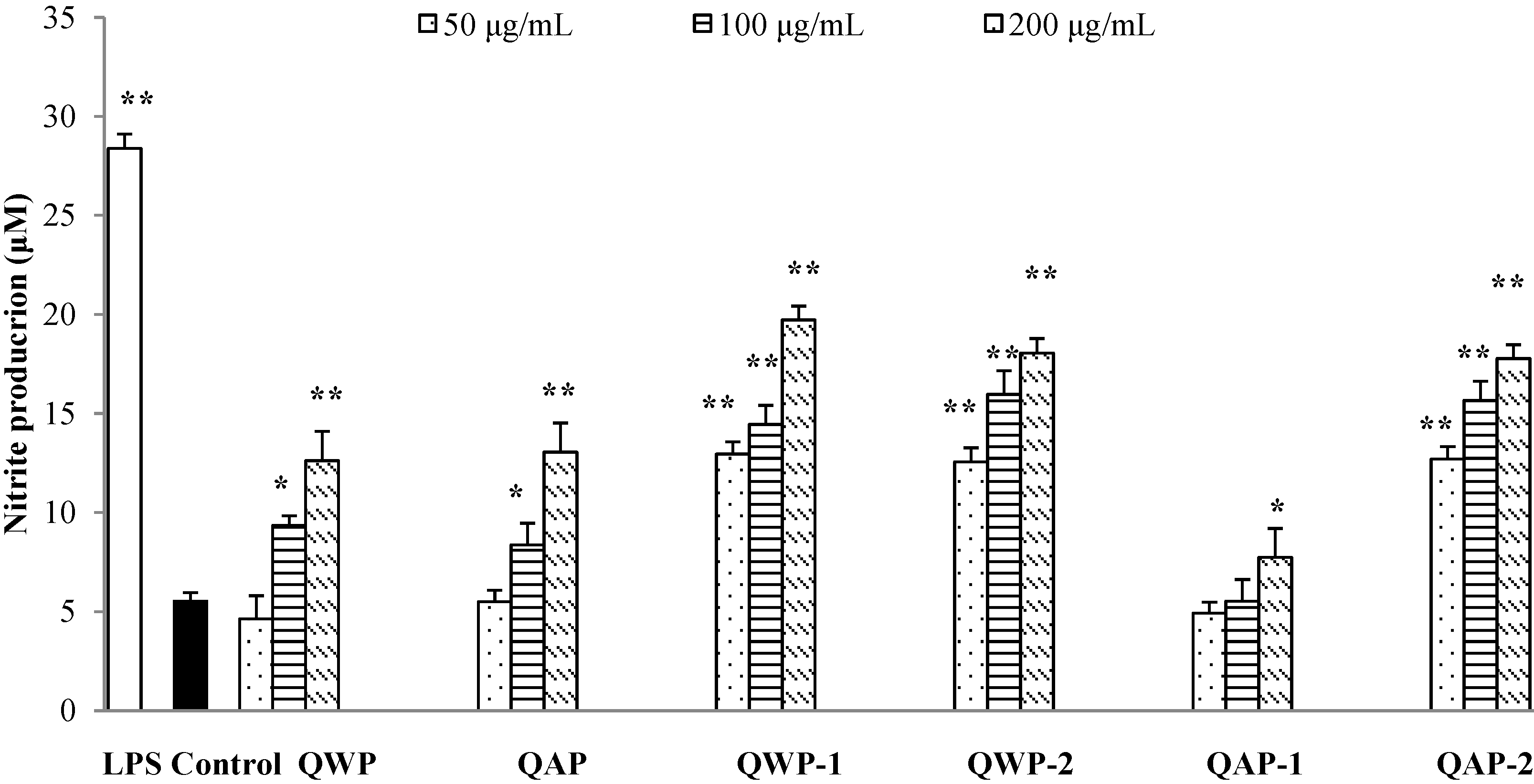
3. Materials and Methods
3.1. Materials and Regents
3.2. Extraction, Separation, and Purification of Polysaccharide
3.3. Analysis of Molecular Weight
3.4. Analysis of Monosaccharide Composition
3.5. Infrared Spectrum Analysis
3.6. Antioxidant Activities Evaluation
3.7. Assay for Immunomodulatory Activity
3.8. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Song, H.F.; Zhang, Q.B.; Zhang, Z.S.; Wang, J. In vitro antioxidant activity of polysaccharides extracted from Bryopsis plumosa. Carbohydr. Polym. 2010, 80, 1057–1061. [Google Scholar]
- Wang, M.; Guilbert, L.J.; Li, J.; Wu, Y.; Pang, P.; Basu, T.K; Shan, J.J. A proprietary extract from North American ginseng (Panax. quinquefolium) enhances IL-2 and IFN-γ productions in murine spleen cells induced by Con-A. Int. J. Immunopharmacol. 2004, 4, 311–315. [Google Scholar]
- Yang, X.B.; Zhao, Y.; Lv, Y. In vivo macrophage activation and physicochemical property of the different polysaccharide fractions purified from Angelica sinensis. Carbohydr. Polym. 2008, 71, 372–379. [Google Scholar]
- Zhu, X.L.; Lin, Z.B. Modulation of cytokines production, granzyme B and perforin in murine CIK cells by Ganoderma lucidum polysaccharides. Carbohydr. Polym. 2006, 63, 188–197. [Google Scholar]
- Xiong, S.L.; Li, A.L.; Huang, N.; Lu, F.; Hou, D.B. Antioxidant and immunoregulatory activity of different polysaccharide fractions from tuber of Ophiopogon japonicus. Carbohydr. Polym. 2011, 86, 1273–1280. [Google Scholar]
- Wang, R.; Chen, P.; Jia, F.; Tang, J.; Ma, F. Optimization of polysaccharides from Panax. japonicus C.A. Meyer by RSM and its anti-oxidant activity. Int. J. Biol. Macromol. 2012, 50, 331–336. [Google Scholar]
- Delatorre-Herrera, J. Current use of quinoa in Chile. Food Rev. Int. 2003, 19, 155–165. [Google Scholar]
- Ando, H.; Chen, Y.; Tang, H.; Shimizu, M.; Watanabe, K.; Mitsunaga, T. Food components in fractions of quinoa seed. Food Sci. Technol. Res. 2002, 8, 80–84. [Google Scholar]
- Woldemichael, G.M.; Wink, M. Identification and biological activities of triterpenoid saponins from Chenopodium quinoa. J. Agric. Food Chem. 2001, 49, 2327–2332. [Google Scholar]
- Yao, Y.; Yang, X.; Shi, Z.; Ren, G. Anti-inflammatory activity of saponins from quinoa (Chenopodium quinoa Willd.) seeds in lipopolysaccharide-stimulated RAW 264.7 macrophages cells. J. Food Sci. 2014, 79, 1018–1023. [Google Scholar]
- Estrada, A.; Li, B.; Laarveld, B. Adjuvant action of Chenopodium quinoa saponins on the induction of antibody responses to intragastric and intranasal administered antigens in mice. Comp. Immunol. Microbiol. Infect. Dis. 1998, 21, 225–236. [Google Scholar]
- G’omez-Caravaca, A.M.; Segura-Carretero, A.; Fernandez-Gutierrez, A.; Caboni, M.F. Simultaneous determination of phenolic compounds and saponins in quinoa (Chenopodium. quinoa Willd.) by a liquid chromatography–diode array detection–electrospray ionization–time-of-flight mass spectrometry methodology. J. Agric. Food Chem. 2011, 59, 10815–10825. [Google Scholar]
- Cordeiro, L.M.C.; Reinhardt, V.D.; Baggio, C.H.; Werner, M.F.D.; Burci, L.M.; Sassaki, G.L.; Iacomini, M. Arabinan and arabinan-rich pectic polysaccharides from quinoa (Chenopodium. quinoa) seeds, Structure and gastroprotective activity. Food Chem. 2012, 130, 937. [Google Scholar]
- Tong, H.; Liu, X.; Tian, D.; Sun, X. Purification, chemical characterization and radical scavenging activities of alkali-extracted polysaccharide fractions isolated from the fruit bodies of Tricholoma matsutake. World J. Microbiol. Biotechnol. 2013, 29, 775–780. [Google Scholar]
- Wu, H.; Min, T.; Li, X.; Li, L.; Lai, F.; Tang, Y.; Yang, X. Physicochemical properties and antioxidant activities of acidic polysaccharides from wampee seeds. Int. J. Biol. Macromol. 2013, 59, 90–95. [Google Scholar]
- Gnanasambanda, R.; Proctor, A. Determination of pectin degree of esterification by diffuse reflectance Fourier transform infrared spectroscopy—Quantitative evaluation of uronic acid and acetamidodeoxyhexose moieties. Food Chem. 2000, 68, 327–332. [Google Scholar]
- Kacurakova, M.; Capek, P.; Sasinkova, V.; Wellner, N.; Ebringerova, A. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar]
- Dini, I.; Tenore, G.C.; Dini, A. Saponins in Ipomoea batatas tubers, isolation, characterization, quantification and antioxidant properties. Food Chem. 2009, 113, 411–419. [Google Scholar]
- Beretta, G.; Granata, P.; Ferrero, M.; Orioli, M.; Analytica, F.R. Standardization of antioxidant properties of honey by a combination of spectrophotometric/fluorimetric assays and chemometrics. Anal. Chim. Acta 2005, 533, 185–191. [Google Scholar]
- Methacanon, P.; Madla, S.; Kirtikara, K.; Prasitsil, M. Tructural elucidation of bioactive fungi-derived polymers. Carbohydr. Polym. 2005, 60, 199–203. [Google Scholar]
- Azike, C.G.; Charpentier, P.A.; Hou, J.; Pei, H.; Lui, E.M. The Yin and Yang actions of North American ginseng root in modulating the immune function of macrophages. Chin. Med. 2011, 6, 21–32. [Google Scholar]
- Zhang, Y.J.; Zhang, L.X.; Yang, J.F.; Liang, Z.Y. Structure analysis of water-soluble polysaccharide CPPS3 isolated from Codonopsis pilosula. Fitoterapia 2010, 81, 157–161. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Yu, X.; Yang, X.; Cui, B.; Wang, L.; Ren, G.X. Antioxidant and immunoregulatory activity of alkali-extractable polysaccharides from North American ginseng. Int. J. Biol. Macromol. 2014, 65, 351–361. [Google Scholar]
- Yao, Y.; Ren, G.X. Effect of thermal treatment on phenolic composition and antioxidant activities of two celery cultivars. LWT-Food Sci. Technol. 2011, 44, 181–185. [Google Scholar]
- Yao, Y.; Yang, X.S.; Tian, J.; Liu, C.Y.; Cheng, X.Z.; Ren, G.X. Antioxidant and antidiabetic activities of black mung bean (Vigna. radiata L.). J. Agric. Food Chem. 2013, 61, 8104–8109. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.; Shi, Z.; Ren, G. Antioxidant and Immunoregulatory Activity of Polysaccharides from Quinoa (Chenopodium quinoa Willd.). Int. J. Mol. Sci. 2014, 15, 19307-19318. https://doi.org/10.3390/ijms151019307
Yao Y, Shi Z, Ren G. Antioxidant and Immunoregulatory Activity of Polysaccharides from Quinoa (Chenopodium quinoa Willd.). International Journal of Molecular Sciences. 2014; 15(10):19307-19318. https://doi.org/10.3390/ijms151019307
Chicago/Turabian StyleYao, Yang, Zhenxing Shi, and Guixing Ren. 2014. "Antioxidant and Immunoregulatory Activity of Polysaccharides from Quinoa (Chenopodium quinoa Willd.)" International Journal of Molecular Sciences 15, no. 10: 19307-19318. https://doi.org/10.3390/ijms151019307
APA StyleYao, Y., Shi, Z., & Ren, G. (2014). Antioxidant and Immunoregulatory Activity of Polysaccharides from Quinoa (Chenopodium quinoa Willd.). International Journal of Molecular Sciences, 15(10), 19307-19318. https://doi.org/10.3390/ijms151019307