Effects of Narrow Band UVB (311 nm) Irradiation on Epidermal Cells

{kind=link}
Abstract
:1. Introduction
2. Narrow Band UVB (311 nm) and Keratinocyte Proliferation
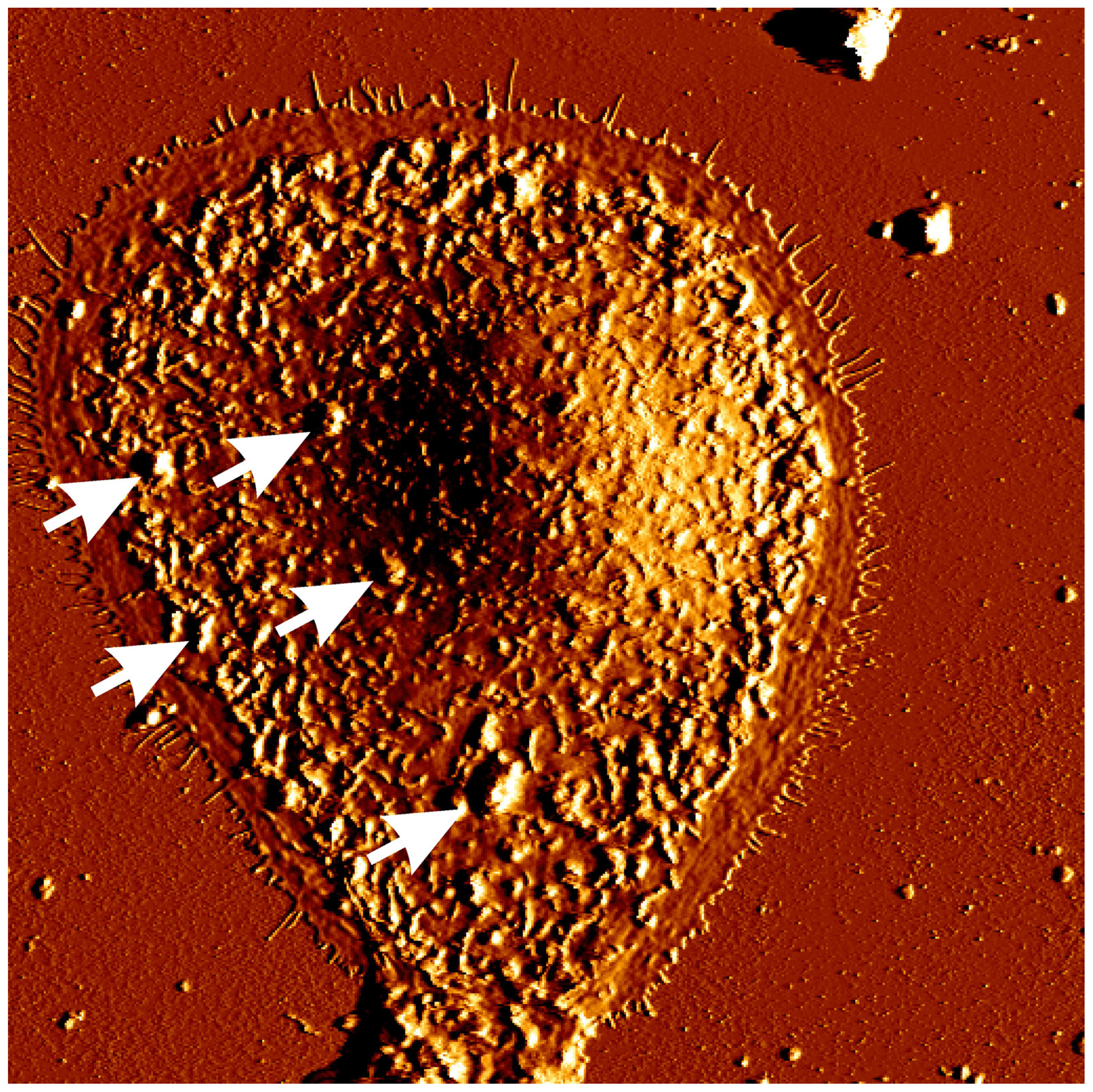
3. Narrow Band UVB (311 nm) and Apoptosis of Keratinocytes
4. Narrow Band UVB (311 nm) Action on Other Cell Types
5. Conclusions
Conflict of Interest
References
- Bissett, D.L.; Hannon, D.P.; Orr, T.V. Wavelength dependence of histological, physical, and visible changes in chronically UV-irradiated hairless mouse skin. Photochem. Photobiol 1989, 50, 763–769. [Google Scholar]
- Xu, Y.; Fisher, G.J. Ultraviolet (UV) light irradiation induced signal transduction in skin photoaging. J. Dermatol. Sci. Suppl 2005, 1, S1–S8. [Google Scholar]
- Weichenthal, M.; Schwarz, T. Phototherapy: How does UV work? Photodermatol. Photoimmunol. Photomed 2005, 21, 260–266. [Google Scholar]
- Canguilhem, B.; Pradines, A.; Baudouin, C.; Boby, C.; Lajoie-Mazenc, I.; Charveron, M.; Favre, G. RhoB protects human keratinocytes from UVB-induced apoptosis through epidermal growth factor receptor signaling. J. Biol. Chem 2005, 280, 43257–43263. [Google Scholar]
- D’Errico, M.; Teson, M.; Calcagnile, A.; Nardo, T.; de Luca, N.; Lazzari, C.; Soddu, S.; Zambruno, G.; Stefanini, M.; Dogliotti, E. Differential role of transcription-coupled repair in UVB-induced response of human fibroblasts and keratinocytes. Cancer Res 2005, 65, 432–438. [Google Scholar]
- Svobodova, A.; Vostalova, J. Solar radiation induced skin damage: Review of protective and preventive options. Int. J. Radiat. Biol 2010, 86, 999–1030. [Google Scholar]
- Lewis, D.A.; Hurwitz, S.A.; Spandau, D.F. UVB-induced apoptosis in normal human keratinocytes: Role of the erbB receptor family. Exp. Cell Res 2003, 284, 316–327. [Google Scholar]
- Enk, C.D.; Jacob-Hirsch, J.; Gal, H.; Verbovetski, I.; Amariglio, N.; Mevorach, D.; Ingber, A.; Givol, D.; Rechavi, G.; Hochberg, M. The UVB-induced gene expression profile of human epidermis in vivo is different from that of cultured keratinocytes. Oncogene 2006, 25, 2601–2614. [Google Scholar]
- Van Weelden, H.; Young, E.; van der Leun, J.C. Therapy of psoriasis: Comparison of photochemotherapy and several variants of phototherapy. Br. J. Dermatol 1980, 103, 1–9. [Google Scholar]
- Parrish, J.A.; Jaenicke, K.F. Action spectrum for phototherapy of psoriasis. J. Invest. Dermatol 1981, 76, 359–362. [Google Scholar]
- Bandow, G.D.; Koo, J.Y.M. Narrow-band ultraviolet B radiation: A review of the current literature. Int. J. Dermatol 2004, 43, 555–561. [Google Scholar]
- Berneburg, M.; Röcken, M.; Benedix, F. Phototherapy with narrowband vs. broadband UVB. Acta Derm. Venereol 2005, 85, 98–108. [Google Scholar]
- Kunisada, M.; Kumimoto, H.; Ishizaki, K.; Sakumi, K.; Nakabeppu, Y.; Nishigori, C. Narrow-band UVB induces more carcinogenic skin tumors than broad-band UVB through the formation of cyclobutane pyrimidine dimer. J. Invest. Dermatol 2007, 127, 2865–2871. [Google Scholar]
- Hearn, R.M.; Kerr, A.C.; Rahim, K.F.; Ferguson, J.; Dawe, R.S. Incidence of skin cancers in 3867 patients treated with narrow-band ultraviolet B phototherapy. Br. J. Dermatol 2008, 159, 931–935. [Google Scholar]
- Dawe, R.S.; Cameron, H.; Yule, S.; Man, I.; Ibbotson, S.H.; Ferguson, J. UV-B phototherapy clears psoriasis through local effects. Arch. Dermatol 2002, 138, 1071–1076. [Google Scholar]
- Macve, J.C.; Norval, M. The effects of UV waveband and cis-urocanic acid on tumour outgrowth in mice. Photochem. Photobiol. Sci 2002, 1, 1006–1011. [Google Scholar]
- Gibbs, N.K. Narrowband UV-B phototherapy clears psoriasis through a combination of local and systemic effects. Arch. Dermatol 2003, 139, 665–666. [Google Scholar]
- Yule, S.; Dawe, R.S.; Cameron, H.; Ferguson, J.; Ibbotson, S.H. Does narrow-band ultraviolet B phototherapy work in atopic dermatitis through a local or a systemic effect? Photodermatol. Photoimmunol. Photomed 2005, 21, 333–335. [Google Scholar]
- Schindl, A.; Klosner, G.; Hönigsmann, H.; Jori, G.; Calzavara-Pinton, P.C.; Trautinger, F. Flow cytometric quantification of UV-induced cell death in a human squamous cell carcinoma-derived cell line: Dose and kinetic studies. J. Photochem. Photobiol. B 1998, 44, 97–106. [Google Scholar]
- Aufiero, B.M.; Talwar, H.; Young, C.; Krishnan, M.; Hatfield, J.S.; Lee, H.K.; Wong, H.K.; Hamzavi, I.; Murakawa, G.J. Narrow-band UVB induces apoptosis in human keratinocytes. J. Photochem. Photobiol. B 2006, 82, 132–139. [Google Scholar]
- Luo, S.; Peng, Z.; Zheng, Y.; Zhang, L.; Feng, Y.; Wang, G. Synergistic effects of acitretin and narrow-band UVB on inducing the expression of heparin-binding epidermal-growth-factor-like growth factor in normal human keratinocytes. Arch. Dermatol. Res 2007, 299, 409–413. [Google Scholar]
- Reich, A.; Lehmann, B.; Meurer, M.; Muller, D.J. Structural alterations provoked by narrow-band ultraviolet B in immortalized keratinocytes: Assessment by atomic force microscopy. Exp. Dermatol 2007, 16, 1007–1015. [Google Scholar]
- Luo, S.; Zheng, Y.; Peng, Z.; Jiang, J.; Gondokaryono, S.; Wang, G.; Ikeda, S. Effects of narrow-band ultraviolet B and tazarotene therapy on keratinocyte proliferation and TIG3 expression. J. Dermatol 2008, 35, 651–657. [Google Scholar]
- Reich, A.; Meurer, M.; Viehweg, A.; Muller, D.J. Narrow-band UVB-induced externalization of selected nuclear antigens in keratinocytes: Implications for lupus erythematosus pathogenesis. Photochem. Photobiol 2009, 85, 1–7. [Google Scholar]
- Reich, A.; Schwudke, D.; Meurer, M.; Lehmann, B.; Shevchenko, A. Lipidome of narrow-band ultraviolet B irradiated keratinocytes shows apoptotic hallmarks. Exp. Dermatol 2010, 19, E103–E110. [Google Scholar]
- Stoll, S.W.; Rittie, L.; Johnson, J.L.; Elder, J.T. Heparin-binding EGF-like growth factor promotes epithelial-mesenchymal transition in human keratinocytes. J. Invest. Dermatol 2012, 132, 2148–2157. [Google Scholar]
- Rácz, E.; Kurek, D.; Kant, M.; Baerveldt, E.M.; Florencia, E.; Mourits, S.; de Ridder, D.; Laman, J.D.; van der Fits, L.; Prens, E.P. GATA3 expression is decreased in psoriasis and during epidermal regeneration; induction by narrow-band UVB and IL-4. PLoS One 2011, 6, e19806. [Google Scholar]
- Yamamoto, M.; Ko, L.J.; Leonard, M.W.; Beug, H.; Orkin, S.H.; Engel, J.D. Activity and tissue-specific expression of the transcription factor NF-E1 multigene family. Genes Dev 1990, 4, 1650–1662. [Google Scholar]
- De Guzman Strong, C.; Wertz, P.W.; Wang, C.; Yang, F.; Meltzer, P.S.; Andl, T.; Millar, S.E.; Ho, I.C.; Pai, S.Y.; Segre, J.A. Lipid defect underlies selective skin barrier impairment of an epidermal-specific deletion of Gata-3. J. Cell Biol 2006, 175, 661–670. [Google Scholar]
- Son do, N.; Li, L.; Katsuyama, H.; Komatsu, N.; Saito, M.; Tanii, H.; Saijoh, K. Abundant expression of Kallikrein 1 gene in human keratinocytes was mediated by GATA3. Gene 2009, 436, 121–127. [Google Scholar]
- Nagui, N.A.; Abdel Hay, R.M.; Rashed, L.A. Effect of narrow band ultraviolet B on survivin in psoriatic skin lesions. Eur. J. Dermatol 2011, 21, 866–869. [Google Scholar]
- Gloor, M.; Scherotzke, A. Age dependence of ultraviolet light-induced erythema following narrow-band UVB exposure. Photodermatol. Photoimmunol. Photomed 2002, 18, 121–126. [Google Scholar]
- Tjioe, M.; Smiths, T.; van de Kerkhof, P.C.M.; Geritsen, M.J.P. The differential effect of broad band vs. narrow-band UVB with respect to photodamage and cutaneous inflammation. Exp. Dermatol 2003, 12, 729–733. [Google Scholar]
- Weatherhead, S.C.; Farr, P.M.; Jamieson, D.; Hallinan, J.S.; Lloyd, J.J.; Wipat, A.; Reynolds, N.J. Keratinocyte apoptosis in epidermal remodeling and clearance of psoriasisinduced by UV radiation. J. Invest. Dermatol 2011, 131, 1916–1926. [Google Scholar]
- Weatherhead, S.C.; Farr, P.M.; Reynolds, N.J. Spectral effects of UV on psoriasis. Photochem. Photobiol. Sci 2013, 12, 47–53. [Google Scholar]
- Cotter, T.G.; Lennon, S.V.; Glynn, J.M.; Green, D.R. Microfilament-disrupting agents prevent the formation of apoptotic bodies in tumor cells undergoing apoptosis. Cancer Res 1992, 52, 997–1005. [Google Scholar]
- Malorni, W.; Donelli, G.; Straface, E.; Santini, M.T.; Paradisi, S.; Giacomoni, P.U. Both UVB and UVB induce cytoskeleton-dependent surface blebbing in epidermoid cells. J. Photochem. Photobiol. B 1994, 26, 265–270. [Google Scholar]
- Malorni, W.; Rivabene, R.; Straface, E.; Rainaldi, G.; Monti, D.; Salvioli, S.; Cossarizza, A.; Franceschi, C. 3-Amiono-benzamide protects cells from UB-V-induced apoptosis by acting on cytoskeleton and substrate adhesion. Biochem. Biophys. Res. Commun 1995, 207, 715–724. [Google Scholar]
- Casciola-Rosen, L.A.; Anhalt, G.; Rosen, A. Auto-antigens targeted in systemic lupus erythematosus are clustered in two populations of surface structures on apoptotic keratinocytes. J. Exp. Med 1994, 179, 1317–1330. [Google Scholar]
- Mathias, S.; Pena, L.A.; Kolesnick, R.N. Signal transduction of stress via ceramide. Biochem. J 1998, 335, 465–480. [Google Scholar]
- Verdonck, L.F.; van Heugten, H.G. Ether lipids are effective cytotoxic drugs against multidrug-resistant acute leukemia cells and can act by the inductionof apoptosis. Leuk. Res 1997, 21, 37–43. [Google Scholar]
- Cabaner, C.; Gajate, C.; Macho, A.; Munoz, E.; Modolell, M.; Mollinedo, F. Induction of apoptosis in human mitogen-activated peripheral blood T-lymphocytes by the ether phospholipid ET-18-OCH3: Involvement of the Fas receptor ligand system. Br. J. Pharmacol 1999, 127, 813–825. [Google Scholar]
- Finstad, H.S.; Dyrendal, H.; Myhrstad, M.C.; Heimli, H.; Drevon, C.A. Uptake and activation of eicosapentaenoic acid are related to accumulation of triacylglycerolin Ramos cells dying from apoptosis. J. Lipid Res 2000, 41, 554–563. [Google Scholar]
- Al-Saffar, N.M.; Titley, J.C.; Robertson, D.; Clarke, P.A.; Jackson, L.E.; Leach, M.O.; Ronen, S.M. Apoptosis is associated with triacylglycerol accumulation in Jurkat T-cells. Br. J. Cancer 2002, 86, 963–970. [Google Scholar]
- Listenberger, L.L.; Han, X.; Lewis, S.E.; Cases, S.; Farese, R.V., Jr; Ory, D.S.; Schaffer, JE. Triglyceride accumulation protects against fatty acid-induced lipotoxicity. Proc. Natl. Acad. Sci. USA 2003, 100, 3077–3082. [Google Scholar]
- Martins de Lima, T.; Cury-Boaventura, M.F.; Giannocco, G.; Nunes, M.T.; Curi, R. Comparative toxicity of fatty acids on a macrophage cell line (J774). Clin. Sci 2006, 111, 307–317. [Google Scholar]
- Cnop, M.; Hannaert, J.C.; Hoorens, A.; Eizirik, D.L.; Pipeleers, D.G. Inverse relationshipbetween cytotoxicity of free fatty acids in pancreatic islet cells and cellulartriglyceride accumulation. Diabetes 2001, 50, 1771–1777. [Google Scholar]
- Healy, D.A.; Watson, R.W.; Newsholem, P. Polyunsaturated and monounsaturated fatty acids increase neutral lipid accumulation, caspase activation and apoptosis in a neutrophil-like, differentiated HL-60 cell line. Clin. Sci 2003, 104, 171–179. [Google Scholar]
- Williams, S.N.O.; Anthony, M.L.; Brindle, K.M. Induction of apoptosis in twomammalian cell lines results in increased levels of fructose-1,6-bisphosphateand CDP-choline as determined by 31P MRS. Magn. Reson. Med 1998, 40, 411–420. [Google Scholar]
- Anthony, M.L.; Zhao, M.; Brindle, K.M. Inhibition of phosphatidylcholine biosynthesis following induction of apoptosis in HL-60 cells. J. Biol. Chem 1999, 274, 19686–19692. [Google Scholar]
- Iturralde, M.; Gamen, S.; Pardo, J.; Bosque, A.; Piñeiro, A.; Alava, M.A.; Naval, J.; Anel, A. Saturated free fatty acid release and intracellular ceramide generation during apoptosis induction are closely related processes. Biochim. Biophys. Acta 2003, 1634, 40–51. [Google Scholar]
- Aronis, A.; Madar, Z.; Tirosh, O. Mechanism of underlying oxidative stress-mediated lipotoxicity: Exposure of J744.2 macrophages to triacylglycerols facilitates mitochondrial reactive oxygen species production and cellular necrosis. Free Radical. Biol. Med 2005, 38, 1221–1230. [Google Scholar]
- Wefers, H.; Melnik, B.C.; Flur, M.; Bluhm, C.; Lehmann, P.; Plewig, G. Influence of UV irradiation on the composition of human stratum corneum lipids. J. Invest. Dermatol 1991, 96, 959–962. [Google Scholar]
- Ozawa, M.; Ferenczi, K.; Kikuchi, T.; Cardinale, I.; Austin, L.M.; Coven, T.R.; Burack, L.H.; Krueger, J.G. 312-Nanometer ultraviolet B light (narrow-band UVB) induces apoptosis of T cells within psoriaticlesions. J. Exp. Med 1999, 189, 711–718. [Google Scholar]
- Sigmundsdottir, H.; Johnston, A.; Gudjonsson, J.E.; Valdimarsson, H. Narrowband-UVB irradiation decreases the productionof pro-inflammatory cytokines by stimulated T cells. Arch. Dermatol. Res 2005, 297, 39–42. [Google Scholar]
- Erkin, G.; Uğur, Y.; Gürer, C.K.; Aşan, E.; Korkusuz, P.; Sahin, S.; Kölemen, F. Effect of PUVA, narrow-band UVB and cyclosporin on inflammatory cells of the psoriatic plaque. J. Cutan. Pathol 2007, 34, 213–219. [Google Scholar]
- El-Ghorr, A.A.; Norval, M. Biological effects of narrow-band (311 nm TL01) UVB irradiation: A review. J. Photochem. Photobiol. B 1997, 38, 99–106. [Google Scholar]
- Walters, I.B.; Burack, L.H.; Coven, T.R.; Gilleaudeau, P.; Krueger, J.G. Suberythemogenic narrow-band UVB is markedly more effective than conventional UVB in treatment of psoriasis vulgaris. J. Am. Acad. Dermatol 1999, 40, 893–900. [Google Scholar]
- Norval, M.; Gibbs, N.K.; Gilmour, J. The role of urocanic acid in UV-induced immunosuppression: Recent advances (1992–1994). Photochem. Photobiol 1995, 62, 209–217. [Google Scholar]
- El-Zawahry, B.M.; Bassiouny, D.A.; Sobhi, R.M.; Abdel-Aziz, E.; Zaki, N.S.; Habib, D.F.; Shahin, D.M. A comparative study on efficacy of UVA1 vs. narrow-band UVB phototherapy in the treatment of vitiligo. Photodermatol. Photoimmunol. Photomed 2012, 28, 84–90. [Google Scholar]
- Wu, C.S.; Lan, C.C.; Yu, H.S. Narrow-band UVB irradiation stimulates the migration and functional development of vitiligo-IgG antibodies-treated pigment cells. J. Eur. Acad. Dermatol. Venereol 2012, 26, 456–464. [Google Scholar]
- Wu, C.S.; Yu, C.L.; Wu, C.S.; Lan, C.C.; Yu, H.S. Narrow-band ultraviolet-B stimulates proliferation and migration of cultured melanocytes. Exp. Dermatol 2004, 13, 755–763. [Google Scholar]
- Dong, D.; Jiang, M.; Xu, X.; Guan, M.; Wu, J.; Chen, Q.; Xiang, L. The effects of NB-UVB on the hair follicle-derived neural crest stem cells differentiating into melanocyte lineage in vitro. J. Dermatol. Sci 2012, 66, 20–28. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Reich, A.; Mędrek, K. Effects of Narrow Band UVB (311 nm) Irradiation on Epidermal Cells. Int. J. Mol. Sci. 2013, 14, 8456-8466. https://doi.org/10.3390/ijms14048456
Reich A, Mędrek K. Effects of Narrow Band UVB (311 nm) Irradiation on Epidermal Cells. International Journal of Molecular Sciences. 2013; 14(4):8456-8466. https://doi.org/10.3390/ijms14048456
Chicago/Turabian StyleReich, Adam, and Karolina Mędrek. 2013. "Effects of Narrow Band UVB (311 nm) Irradiation on Epidermal Cells" International Journal of Molecular Sciences 14, no. 4: 8456-8466. https://doi.org/10.3390/ijms14048456
APA StyleReich, A., & Mędrek, K. (2013). Effects of Narrow Band UVB (311 nm) Irradiation on Epidermal Cells. International Journal of Molecular Sciences, 14(4), 8456-8466. https://doi.org/10.3390/ijms14048456