Interplay of Dynamic Transcription and Chromatin Remodeling: Lessons from Yeast

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Regulation of Transcriptional Activation
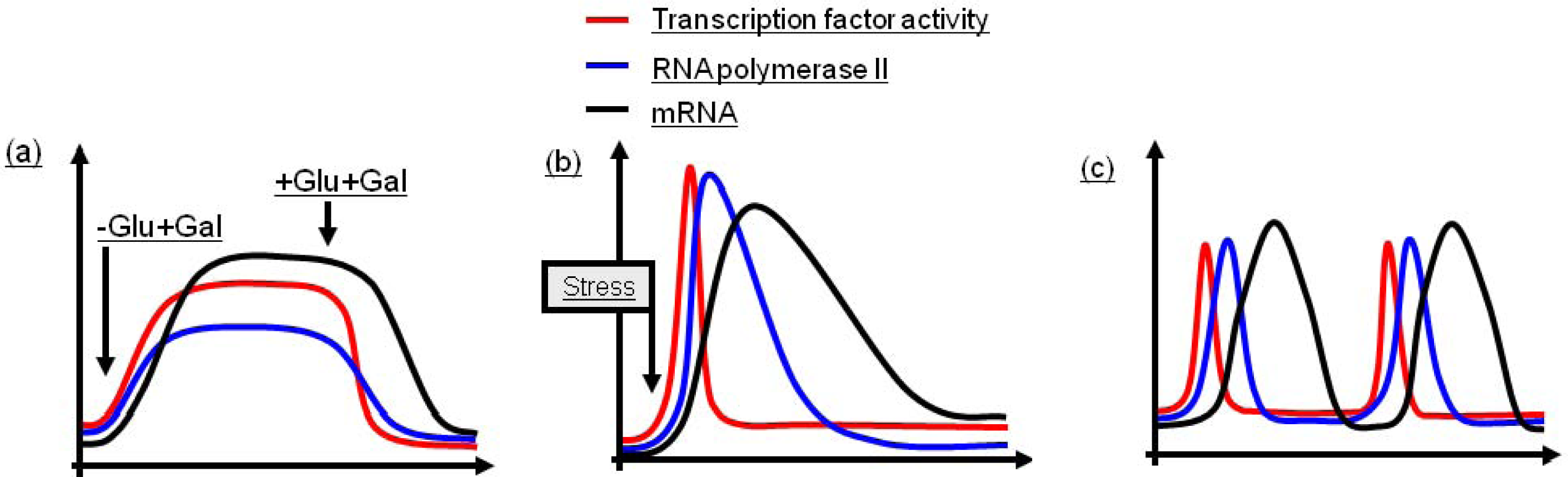
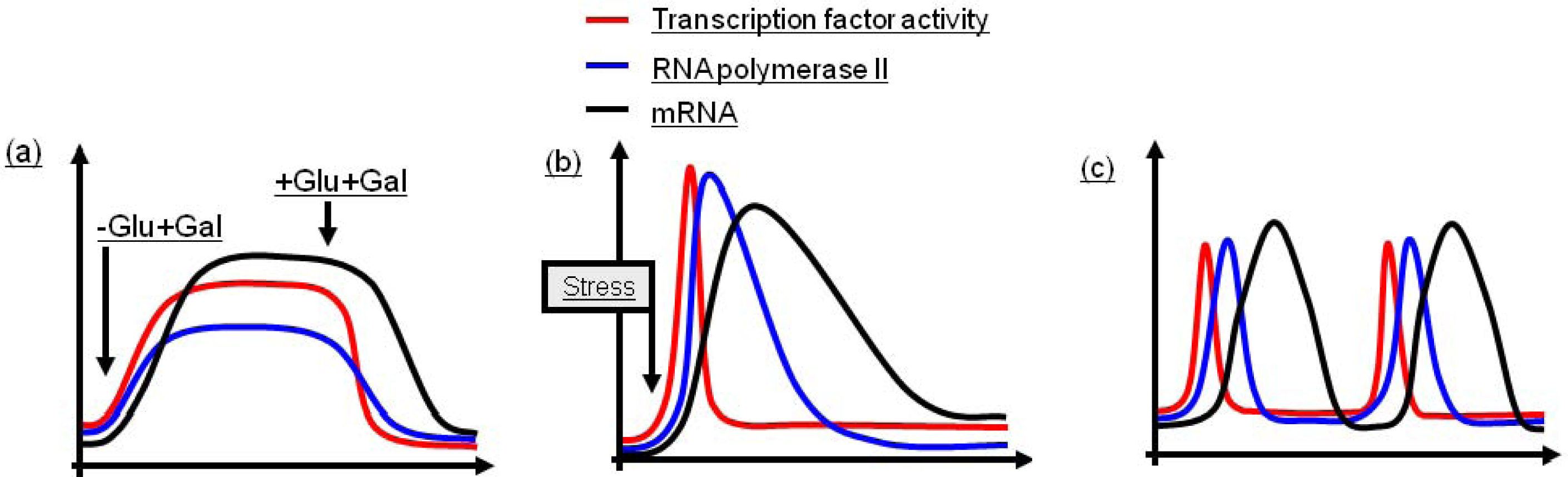
2.1. Regulation of Induced, Sustained Transcription
2.2. Regulation of Induced, Single Pulse Transcription
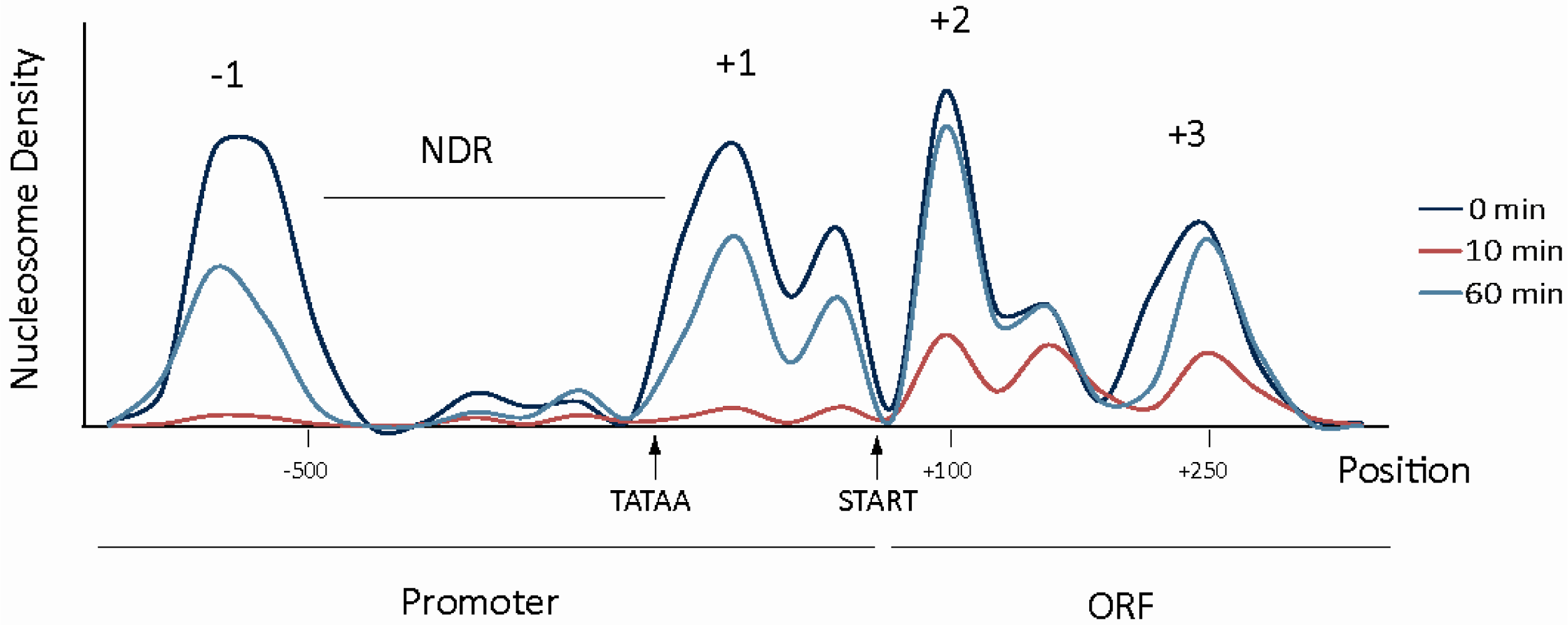
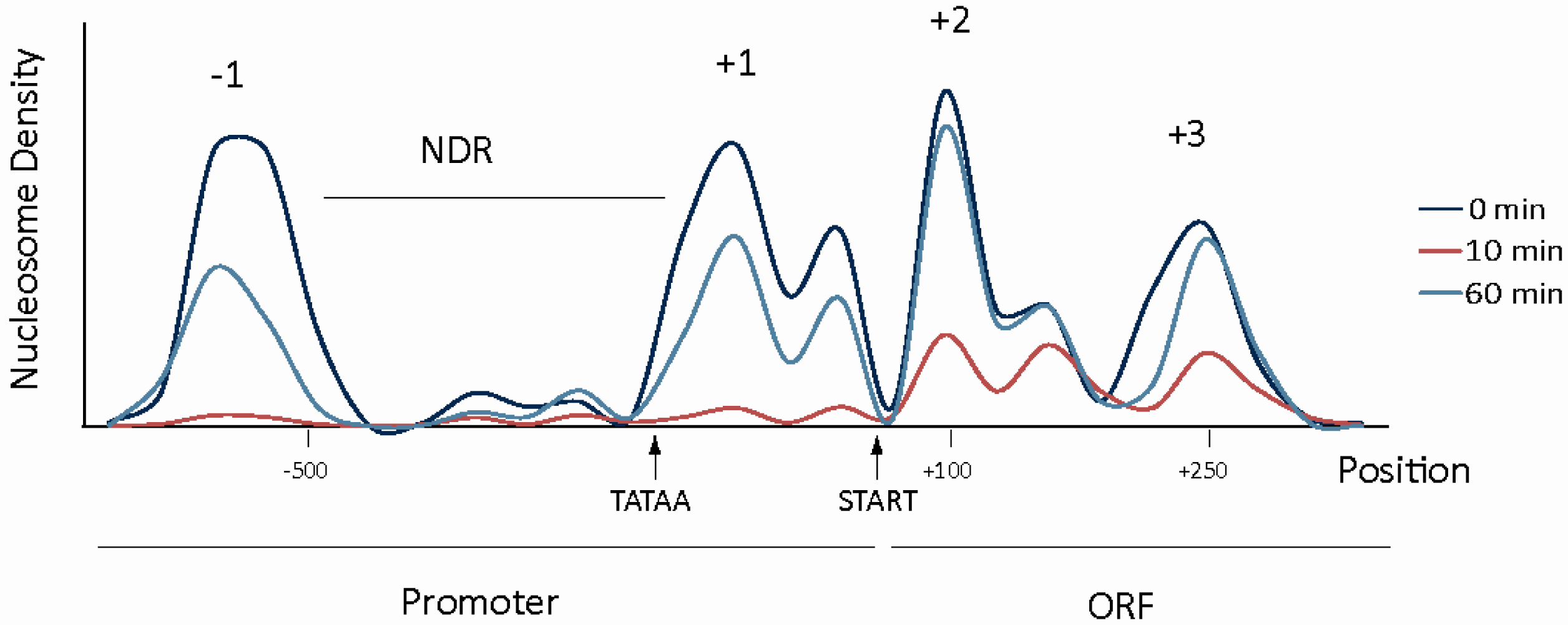
3. Repressive Role of Chromatin
4. Conclusions
Acknowledgments
References
- Luger, K; Mader, AW; Richmond, RK; Sargent, DF; Richmond, TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar]
- Yuan, GC; Liu, YJ; Dion, MF; Slack, MD; Wu, LF; Altschuler, SJ; Rando, OJ. Genome-scale identification of nucleosome positions in S. cerevisiae. Science 2005, 309, 626–630. [Google Scholar]
- Bai, L; Ondracka, A; Cross, FR. Multiple sequence-specific factors generate the nucleosome-depleted region on CLN2 promoter. Mol. Cell 2011, 42, 465–476. [Google Scholar]
- Jiang, C; Pugh, BF. Nucleosome positioning and gene regulation: Advances through genomics. Nat. Rev. Genet 2009, 10, 161–172. [Google Scholar]
- Yosef, N; Regev, A. Impulse control: Temporal dynamics in gene transcription. Cell 2011, 144, 886–896. [Google Scholar]
- Chechik, G; Koller, D. Timing of gene expression responses to environmental changes. J. Comput. Biol 2009, 16, 279–290. [Google Scholar]
- Rougvie, AE; Lis, JT. The RNA polymerase II molecule at the 5’ end of the uninduced hsp70 gene of D. melanogaster is transcriptionally engaged. Cell 1988, 54, 795–804. [Google Scholar]
- Schwabish, MA; Struhl, K. Asf1 mediates histone eviction and deposition during elongation by RNA polymerase II. Mol. Cell 2006, 22, 415–422. [Google Scholar]
- Eberharter, A; Becker, PB. ATP-dependent nucleosome remodelling: Factors and functions. J. Cell Sci 2004, 117, 3707–3711. [Google Scholar]
- Mizuguchi, G; Shen, X; Landry, J; Wu, WH; Sen, S; Wu, C. ATP-driven exchange of histone H2A.Z variant catalyzed by SWR1 chromatin remodeling complex. Science 2004, 303, 343–348. [Google Scholar]
- Papamichos-Chronakis, M; Watanabe, S; Rando, OJ; Peterson, CL. Global regulation of H2A.Z localization by the INO80 chromatin-remodeling enzyme is essential for genome integrity. Cell 2011, 144, 200–213. [Google Scholar]
- Guillemette, B; Bataille, AR; Gevry, N; Adam, M; Blanchette, M; Robert, F; Gaudreau, L. Variant histone H2A.Z is globally localized to the promoters of inactive yeast genes and regulates nucleosome positioning. PLoS Biol 2005, 3, e384. [Google Scholar]
- Zhang, H; Roberts, DN; Cairns, BR. Genome-wide dynamics of Htz1, a histone H2A variant that poises repressed/basal promoters for activation through histone loss. Cell 2005, 123, 219–231. [Google Scholar]
- Raisner, RM; Hartley, PD; Meneghini, MD; Bao, MZ; Liu, CL; Schreiber, SL; Rando, OJ; Madhani, HD. Histone variant H2A.Z marks the 5’ ends of both active and inactive genes in euchromatin. Cell 2005, 123, 233–248. [Google Scholar]
- Altaf, M; Auger, A; Monnet-Saksouk, J; Brodeur, J; Piquet, S; Cramet, M; Bouchard, N; Lacoste, N; Utley, RT; Gaudreau, L; et al. NuA4-dependent acetylation of nucleosomal histones H4 and H2A directly stimulates incorporation of H2A.Z by the SWR1 complex. J. Biol. Chem 2010, 285, 15966–15977. [Google Scholar]
- Keogh, MC; Mennella, TA; Sawa, C; Berthelet, S; Krogan, NJ; Wolek, A; Podolny, V; Carpenter, LR; Greenblatt, JF; Baetz, K; et al. The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4. Genes Dev 2006, 20, 660–665. [Google Scholar]
- Santisteban, MS; Hang, M; Smith, MM. Histone variant H2A.Z and RNA polymerase II transcription elongation. Mol. Cell. Biol 2011, 31, 1848–1860. [Google Scholar]
- Traven, A; Jelicic, B; Sopta, M. Yeast Gal4: A transcriptional paradigm revisited. EMBO Rep 2006, 7, 496–499. [Google Scholar]
- Cairns, BR; Erdjument-Bromage, H; Tempst, P; Winston, F; Kornberg, RD. Two actin-related proteins are shared functional components of the chromatin-remodeling complexes RSC and SWI/SNF. Mol. Cell 1998, 2, 639–651. [Google Scholar]
- Hassan, AH; Prochasson, P; Neely, KE; Galasinski, SC; Chandy, M; Carrozza, MJ; Workman, JL. Function and selectivity of bromodomains in anchoring chromatin-modifying complexes to promoter nucleosomes. Cell 2002, 111, 369–379. [Google Scholar]
- Floer, M; Wang, X; Prabhu, V; Berrozpe, G; Narayan, S; Spagna, D; Alvarez, D; Kendall, J; Krasnitz, A; Stepansky, A; et al. A RSC/nucleosome complex determines chromatin architecture and facilitates activator binding. Cell 2010, 141, 407–418. [Google Scholar]
- Larschan, E; Winston, F. The S. cerevisiae SAGA complex functions in vivo as a coactivator for transcriptional activation by Gal4. Genes Dev 2001, 15, 1946–1956. [Google Scholar]
- Kuo, MH; Brownell, JE; Sobel, RE; Ranalli, TA; Cook, RG; Edmondson, DG; Roth, SY; Allis, CD. Transcription-linked acetylation by Gcn5p of histones H3 and H4 at specific lysines. Nature 1996, 383, 269–272. [Google Scholar]
- Bryant, GO; Ptashne, M. Independent recruitment in vivo by Gal4 of two complexes required for transcription. Mol. Cell 2003, 11, 1301–1309. [Google Scholar]
- Bhaumik, SR; Green, MR. SAGA is an essential in vivo target of the yeast acidic activator Gal4p. Genes Dev 2001, 15, 1935–1945. [Google Scholar]
- Bryant, GO; Prabhu, V; Floer, M; Wang, X; Spagna, D; Schreiber, D; Ptashne, M. Activator control of nucleosome occupancy in activation and repression of transcription. PLoS Biol 2008, 6, 2928–2939. [Google Scholar]
- Hassan, AH; Neely, KE; Workman, JL. Histone acetyltransferase complexes stabilize swi/snf binding to promoter nucleosomes. Cell 2001, 104, 817–827. [Google Scholar]
- Chandy, M; Gutierrez, JL; Prochasson, P; Workman, JL. SWI/SNF displaces SAGA-acetylated nucleosomes. Eukaryotic Cell 2006, 5, 1738–1747. [Google Scholar]
- Korber, P; Luckenbach, T; Blaschke, D; Horz, W. Evidence for histone eviction in trans upon induction of the yeast PHO5 promoter. Mol. Cell. Biol 2004, 24, 10965–10974. [Google Scholar]
- Almer, A; Horz, W. Nuclease hypersensitive regions with adjacent positioned nucleosomes mark the gene boundaries of the PHO5/PHO3 locus in yeast. EMBO J 1986, 5, 2681–2687. [Google Scholar]
- Schmid, A; Fascher, KD; Horz, W. Nucleosome disruption at the yeast PHO5 promoter upon PHO5 induction occurs in the absence of DNA replication. Cell 1992, 71, 853–864. [Google Scholar]
- Gregory, PD; Schmid, A; Zavari, M; Munsterkotter, M; Horz, W. Chromatin remodelling at the PHO8 promoter requires SWI-SNF and SAGA at a step subsequent to activator binding. EMBO J 1999, 18, 6407–6414. [Google Scholar]
- Barbaric, S; Luckenbach, T; Schmid, A; Blaschke, D; Horz, W; Korber, P. Redundancy of chromatin remodeling pathways for the induction of the yeast PHO5 promoter in vivo. J. Biol. Chem 2007, 282, 27610–27621. [Google Scholar]
- Adkins, MW; Howar, SR; Tyler, JK. Chromatin disassembly mediated by the histone chaperone Asf1 is essential for transcriptional activation of the yeast PHO5 and PHO8 genes. Mol. Cell 2004, 14, 657–666. [Google Scholar]
- Adkins, MW; Williams, SK; Linger, J; Tyler, JK. Chromatin disassembly from the PHO5 promoter is essential for the recruitment of the general transcription machinery and coactivators. Mol. Cell. Biol 2007, 27, 6372–6382. [Google Scholar]
- Korber, P; Barbaric, S; Luckenbach, T; Schmid, A; Schermer, UJ; Blaschke, D; Horz, W. The histone chaperone Asf1 increases the rate of histone eviction at the yeast PHO5 and PHO8 promoters. J. Biol. Chem 2006, 281, 5539–5545. [Google Scholar]
- Hirsch, JP; Henry, SA. Expression of the Saccharomyces cerevisiae inositol-1-phosphate synthase (INO1) gene is regulated by factors that affect phospholipid synthesis. Mol. Cell. Biol 1986, 6, 3320–3328. [Google Scholar]
- Ford, J; Odeyale, O; Eskandar, A; Kouba, N; Shen, CH. A SWI/SNF- and INO80-dependent nucleosome movement at the INO1 promoter. Biochem. Biophys. Res. Commun 2007, 361, 974–979. [Google Scholar]
- Ford, J; Odeyale, O; Shen, CH. Activator-dependent recruitment of SWI/SNF and INO80 during INO1 activation. Biochem. Biophys. Res. Commun 2008, 373, 602–606. [Google Scholar]
- Alepuz, PM; de Nadal, E; Zapater, M; Ammerer, G; Posas, F. Osmostress-induced transcription by Hot1 depends on a Hog1-mediated recruitment of the RNA Pol II. EMBO J 2003, 22, 2433–2442. [Google Scholar]
- Alepuz, PM; Jovanovic, A; Reiser, V; Ammerer, G. Stress-induced map kinase Hog1 is part of transcription activation complexes. Mol. Cell 2001, 7, 767–777. [Google Scholar]
- De Nadal, E; Zapater, M; Alepuz, PM; Sumoy, L; Mas, G; Posas, F. The MAPK Hog1 recruits Rpd3 histone deacetylase to activate osmoresponsive genes. Nature 2004, 427, 370–374. [Google Scholar]
- Ruiz-Roig, C; Vieitez, C; Posas, F; de Nadal, E. The Rpd3L HDAC complex is essential for the heat stress response in yeast. Mol. Microbiol 2010, 76, 1049–1062. [Google Scholar]
- Alejandro-Osorio, AL; Huebert, DJ; Porcaro, DT; Sonntag, ME; Nillasithanukroh, S; Will, JL; Gasch, AP. The histone deacetylase Rpd3p is required for transient changes in genomic expression in response to stress. Genome Biol 2009, 10, R57. [Google Scholar]
- Ferreiro, I; Barragan, M; Gubern, A; Ballestar, E; Joaquin, M; Posas, F. The p38 SAPK is recruited to chromatin via its interaction with transcription factors. J. Biol. Chem 2010, 285, 31819–31828. [Google Scholar]
- Erkina, TY; Zou, Y; Freeling, S; Vorobyev, VI; Erkine, AM. Functional interplay between chromatin remodeling complexes RSC, SWI/SNF and ISWI in regulation of yeast heat shock genes. Nucleic Acids Res 2010, 38, 1441–1449. [Google Scholar]
- Mason, PB; Struhl, K. The FACT complex travels with elongating RNA polymerase II and is important for the fidelity of transcriptional initiation in vivo. Mol. Cell. Biol 2003, 23, 8323–8333. [Google Scholar]
- Kaplan, CD; Laprade, L; Winston, F. Transcription elongation factors repress transcription initiation from cryptic sites. Science 2003, 301, 1096–1099. [Google Scholar]
- Lindstrom, KC; Vary, JC, Jr; Parthun, MR; Delrow, J; Tsukiyama, T. Isw1 functions in parallel with the NuA4 and Swr1 complexes in stress-induced gene repression. Mol. Cell. Biol 2006, 26, 6117–6129. [Google Scholar]
- Whitehouse, I; Tsukiyama, T. Antagonistic forces that position nucleosomes in vivo. Nat. Struct. Mol. Biol 2006, 13, 633–640. [Google Scholar]
- Rundlett, SE; Carmen, AA; Suka, N; Turner, BM; Grunstein, M. Transcriptional repression by UME6 involves deacetylation of lysine 5 of histone H4 by RPD3. Nature 1998, 392, 831–835. [Google Scholar]
- Carrozza, MJ; Li, B; Florens, L; Suganuma, T; Swanson, SK; Lee, KK; Shia, WJ; Anderson, S; Yates, J; Washburn, MP; et al. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell 2005, 123, 581–592. [Google Scholar]
- Keogh, MC; Kurdistani, SK; Morris, SA; Ahn, SH; Podolny, V; Collins, SR; Schuldiner, M; Chin, K; Punna, T; Thompson, NJ; et al. Cotranscriptional set2 methylation of histone H3 lysine 36 recruits a repressive Rpd3 complex. Cell 2005, 123, 593–605. [Google Scholar]
- Klopf, E; Paskova, L; Sole, C; Mas, G; Petryshyn, A; Posas, F; Wintersberger, U; Ammerer, G; Schuller, C. Cooperation between the INO80 complex and histone chaperones determines adaptation of stress gene transcription in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol 2009, 29, 4994–5007. [Google Scholar]
- Yoh, SM; Cho, H; Pickle, L; Evans, RM; Jones, KA. The Spt6 SH2 domain binds Ser2-P RNAPII to direct Iws1-dependent mRNA splicing and export. Genes Dev 2007, 21, 160–174. [Google Scholar]
- Ivanovska, I; Jacques, PE; Rando, OJ; Robert, F; Winston, F. Control of chromatin structure by spt6: Different consequences in coding and regulatory regions. Mol. Cell. Biol 2011, 31, 531–541. [Google Scholar]
- Morrison, AJ; Highland, J; Krogan, NJ; Arbel-Eden, A; Greenblatt, JF; Haber, JE; Shen, X. INO80 and gamma-H2AX interaction links ATP-dependent chromatin remodeling to DNA damage repair. Cell 2004, 119, 767–775. [Google Scholar]
- Udugama, M; Sabri, A; Bartholomew, B. The INO80 ATP-dependent chromatin remodeling complex is a nucleosome spacing factor. Mol. Cell. Biol 2011, 31, 662–673. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Niederacher, G.; Klopf, E.; Schüller, C. Interplay of Dynamic Transcription and Chromatin Remodeling: Lessons from Yeast. Int. J. Mol. Sci. 2011, 12, 4758-4769. https://doi.org/10.3390/ijms12084758
Niederacher G, Klopf E, Schüller C. Interplay of Dynamic Transcription and Chromatin Remodeling: Lessons from Yeast. International Journal of Molecular Sciences. 2011; 12(8):4758-4769. https://doi.org/10.3390/ijms12084758
Chicago/Turabian StyleNiederacher, Gerhard, Eva Klopf, and Christoph Schüller. 2011. "Interplay of Dynamic Transcription and Chromatin Remodeling: Lessons from Yeast" International Journal of Molecular Sciences 12, no. 8: 4758-4769. https://doi.org/10.3390/ijms12084758
APA StyleNiederacher, G., Klopf, E., & Schüller, C. (2011). Interplay of Dynamic Transcription and Chromatin Remodeling: Lessons from Yeast. International Journal of Molecular Sciences, 12(8), 4758-4769. https://doi.org/10.3390/ijms12084758