
Selected Polyphenols of Polish Poplar Propolis as a Key Component Shaping Its Antibacterial Properties—In Vitro and In Silico Approaches
 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Polyphenol Content and Antioxidant Properties
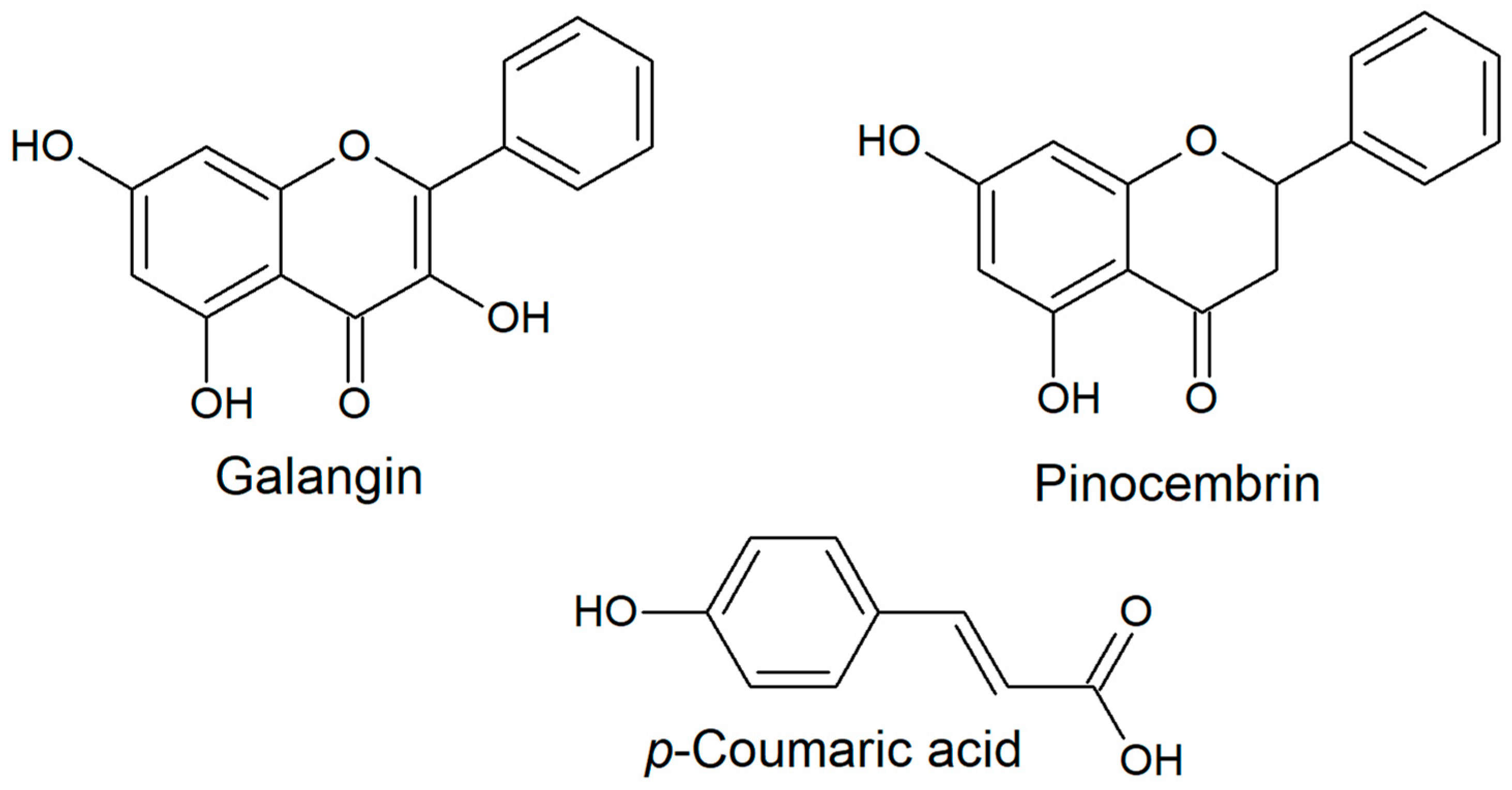
2.2. Polyphenolic Profile
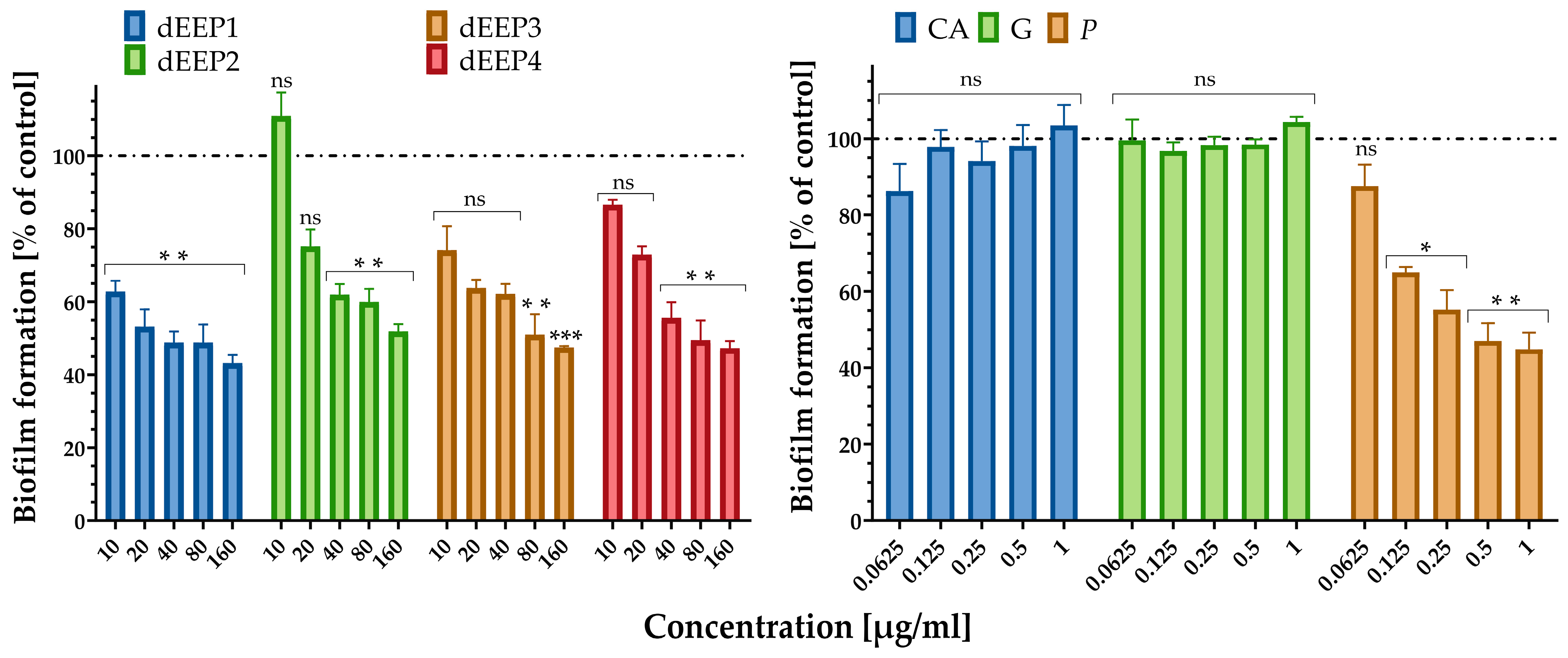
2.3. Antibacterial Properties
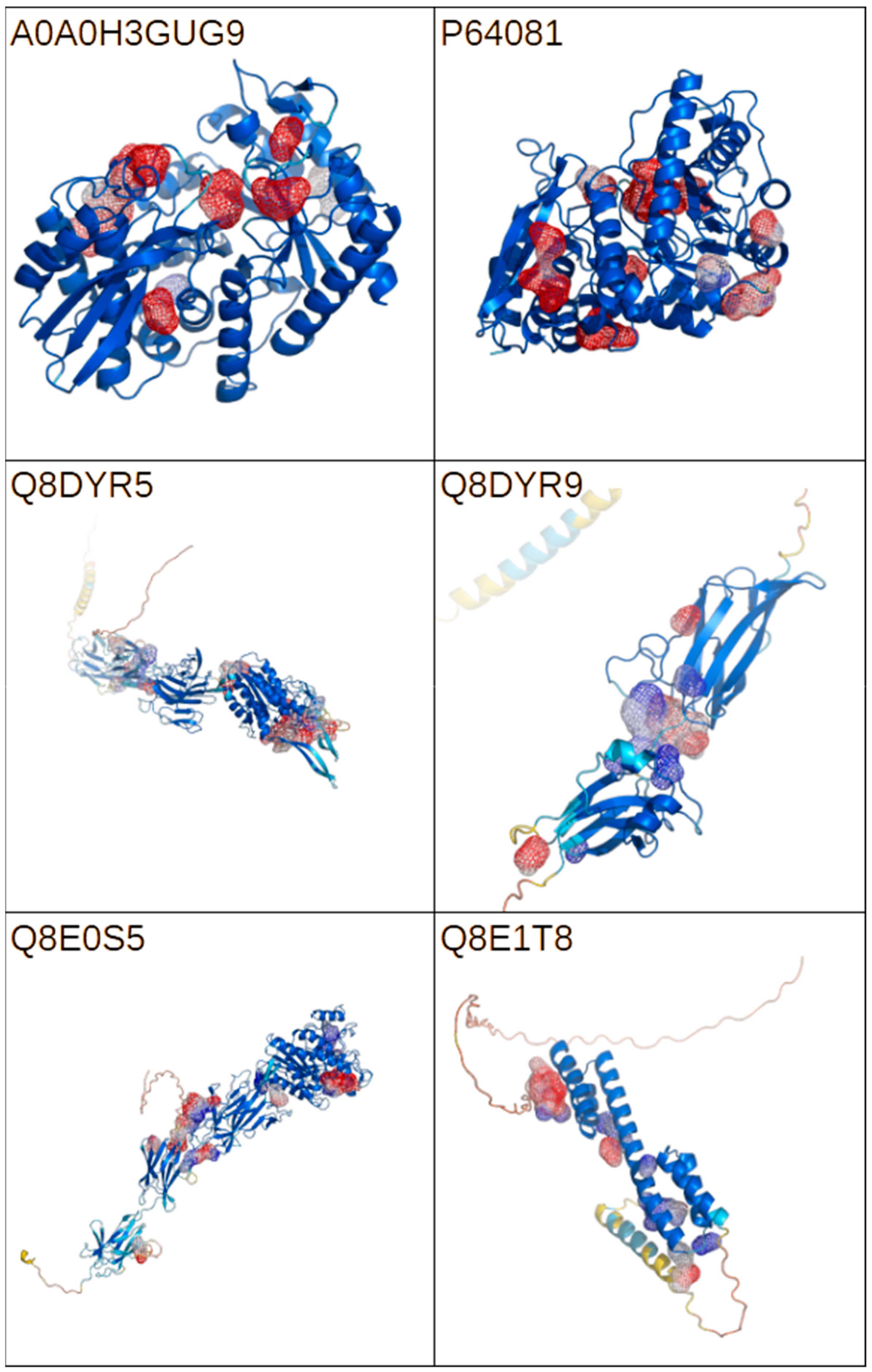
2.4. Bioinformatic Analysis of Putative Virulence Factors
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Ethanolic Extracts of Propolis Preparation
3.3. Total Phenolic and Flavonoid Contents and Antioxidant Capacity
3.4. HPLC-DAD Analysis
3.5. Antibacterial Activity
Minimum Inhibitory and Anti-Biofilm Activity
3.6. Structure Analysis and Visualization
3.7. Docking Simulations
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EEP | ethanolic extract of propolis |
| dEEP | dry ethanolic extract of propolis |
| FRAP | ferric reducing antioxidant power |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| CUPRAC | cupric reducing antioxidant activity |
| TPC | total phenolic content |
| TFC | total flavonoid content |
| MIC | minimum inhibitory concentration |
| HPLC-DAD | high-performance liquid chromatography with diode array detector |
| CAPE | caffeic acid phenetyl ester |
| QE | quercetin equivalents |
| GAE | gallic acid equivalents |
| SD | standard deviation |
| ATCC | American Type Culture Collection |
| DSM | Deutsche Sammlung von Mikroorganismen |
| TPTZ | 2,4,6-Tris(2-pyridyl)-s-triazine |
| MHB | Mueller–Hinton broth |
| MHA | Mueller–Hinton agar |
| MTT | 1-(4,5-Dimethylthiazol-2-yl)-3,5-diphenylformazan |
References
- Kuropatnicki, A.K.; Szliszka, E.; Król, W. Historical aspects of propolis research in modern times. Evid.-Based Complement. Altern. Med. 2013, 2013, 964149. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.R.; Abdelnour, S.A.; Patra, A.K.; Dhama, K.; Dawood, M.A.O.; Elnesr, S.S.; Alagawany, M. Propolis: Properties and composition, health benefits and applications in fish nutrition. Fish Shellfish Immunol. 2021, 115, 179–188. [Google Scholar] [CrossRef]
- Bhatti, N.; Hajam, Y.A.; Mushtaq, S.; Kaur, L.; Kumar, R.; Rai, S. A review on dynamic pharmacological potency and multifaceted biological activities of propolis. Discov. Sustain. 2024, 5, 185. [Google Scholar] [CrossRef]
- Bankova, V.S.; de Castro, S.L.; Marcucci, M.C. Propolis: Recent advances in chemistry and plant origin. Apidologie 2000, 31, 3–15. [Google Scholar] [CrossRef]
- Irigoiti, Y.; Navarro, A.; Yamul, D.; Libonatti, C.; Tabera, A.; Basualdo, M. The use of propolis as a functional food ingredient: A review. Trends Food Sci. Technol. 2021, 115, 297–306. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Rzepecka-Stojko, A.; Górecki, M.; Stojko, J.; Sosada, M.; Swierczek-Zieba, G. Structure and antioxidant activity of polyphenols derived from propolis. Molecules 2014, 19, 78–101. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.I.; Ullah, A.; Khan, K.A.; Attaullah, M.; Khan, H.; Ali, H.; Bashir, M.A.; Tahir, M.; Ansari, M.J.; Ghramh, H.A.; et al. Composition and functional properties of propolis (bee glue): A review. Saudi J. Biol. Sci. 2019, 26, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Salatino, M.L.F. Scientific note: Often quoted, but not factual data about propolis composition. Apidologie 2021, 52, 312–314. [Google Scholar] [CrossRef]
- Šuran, J.; Cepanec, I.; Mašek, T.; Radi, B. Propolis Extract and Its Bioactive Compounds—From Traditional to Modern Extraction Technologies. Molecules 2021, 26, 2930. [Google Scholar] [CrossRef]
- Kaleta, J. Analiza Fizykochemiczna Propolisu i Możliwości Jego Standaryzacji. Ph.D. Thesis, Jagiellonian University in Kraków, Kraków, Poland, 2007. (In Polish). [Google Scholar]
- Kędzia, B.; Hołderna-Kędzia, E. Chemical composition of Polish propolis. Part II. New studies. Postępy Fitoter. 2009, 2, 122–128. (In Polish) [Google Scholar]
- Miłek, M.; Franke, G.; Tomczyk, M.; Górecki, M.; Cwiková, O.; Jarošová, A.; Dżugan, M. The Influence of Geographical Origin on Poplar Propolis Composition and the Impact of Human Microbiota. Pharmaceuticals 2024, 17, 768. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Keskin, Ş.; Bobis, O.; Felitti, R.; Górecki, M.; Otręba, M.; Stojko, J.; Olczyk, P.; Kolayli, S.; Rzepecka-Stojko, A. Comparison of the Antioxidant Activity of Propolis Samples from Different Geographical Regions. Plants 2022, 11, 1203. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, T.G.; dos Santos Arruda, R.E.; da Cruz Almeida, E.T.; dos Santos Oliveira, J.M.; Basílio-Júnior, I.D.; Celerino de Moraes Porto, I.C.; Rodrigues Sabino, A.; Tonholo, J.; Gray, A.; Ebel, R.A.E.; et al. Comprehensive multivariate correlations between climatic effect, metabolite-profile, antioxidant capacity and antibacterial activity of Brazilian red propolis metabolites during seasonal study. Sci. Rep. 2019, 9, 18293. [Google Scholar] [CrossRef]
- Vera, N.; Solorzano, E.; Ordoñez, R.; Maldonado, L.; Bedascarrasbure, E.; Isla, M.I. Chemical composition of Argentinean propolis collected in extreme regions and its relation with antimicrobial and antioxidant activities. Nat. Prod. Commun. 2011, 6, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Ristivojević, P.; Trifković, J.; Andrić, F.; Milojković-Opsenica, D. Poplar-type propolis: Chemical composition, botanical origin and biological activity. Nat. Prod. Commun. 2015, 10, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Contieri, L.S.; de Souza Mesquita, L.M.; Sanches, V.L.; Chaves, J.; Pizani, R.S.; da Silva, L.C.; Viganó, J.; Ventura, S.P.M.; Rostagno, M.A. Recent progress on the recovery of bioactive compounds obtained from propolis as a natural resource: Processes, and applications. Sep. Purif. Technol. 2022, 298, 121640. [Google Scholar] [CrossRef]
- Galeotti, F.; Maccari, F.; Fachini, A.; Volpi, N. Chemical composition and antioxidant activity of propolis prepared in different forms and in different solvents useful for finished products. Foods 2018, 7, 41. [Google Scholar] [CrossRef]
- Miłek, M.; Bonikowski, R.; Dżugan, M. The effect of extraction conditions on the chemical profile of obtained raw poplar propolis extract. Chem. Pap. 2024, 78, 6709–6720. [Google Scholar] [CrossRef]
- Bankova, V. Chemical diversity of propolis and the problem of standardization. J. Ethnopharmacol. 2005, 100, 114–117. [Google Scholar] [CrossRef]
- Marquiafável, F.S.; Nascimento, A.P.; Barud, H.d.S.; Marquele-Oliveira, F.; de-Freitas, L.A.P.; Bastos, J.K.; Berretta, A.A. Development and characterization of a novel standardized propolis dry extract obtained by factorial design with high artepillin C content. J. Pharm. Technol. Drug Res. 2015, 4, 1. [Google Scholar] [CrossRef]
- Paul, R.K.; Dutta, D.; Chakraborty, D.; Nayak, A.; Dutta, P.K.; Nag, M. Antimicrobial agents from natural sources: An overview. Adv. Pharm. J. 2019, 4, 41–51. [Google Scholar] [CrossRef]
- Parolia, A.; Thomas, M.S.; Kundabala, M.; Mohan, M. Propolis and its potential uses in oral health. Int. J. Med. Med. Sci. 2010, 2, 210–215. [Google Scholar]
- Nichitoi, M.M.; Josceanu, A.M.; Isopescu, R.D.; Isopencu, G.O.; Geana, E.I.; Ciucure, C.T.; Lavric, V. Polyphenolics profile effects upon the antioxidant and antimicrobial activity of propolis extracts. Sci. Rep. 2021, 11, 20113. [Google Scholar] [CrossRef]
- Lotfy, M. Biological activity of bee propolis in health and disease. Asian Pac. J. Cancer Prev. 2006, 7, 22–31. [Google Scholar]
- Machado, B.; Pulcino, T.; Silva, A.; Melo, D.; Silva, R.; Mendonca, I. Propolis as an alternative in prevention and control of dental cavity. J. Apitherapy 2016, 1, 47. [Google Scholar] [CrossRef]
- Miłek, M.; Ciszkowicz, E.; Tomczyk, M.; Sidor, E.; Zaguła, G.; Lecka-Szlachta, K.; Pasternakiewicz, A.; Dżugan, M. Poplar-Type Polish Propolis Considering Local Flora Diversity in Relation to Antibacterial and Anticancer Activities in Human Breast Cancer Cells. Molecules 2022, 27, 725. [Google Scholar] [CrossRef]
- Miłek, M.; Ciszkowicz, E.; Zaguła, G.; Grabek-Lejko, D.; Pasternakiewicz, A.; Lecka-Szlachta, K.; Dzugan, M. Quality of Propolis Commercially Available on Podkarpacki Beekeeping Market. J. Apic. Sci. 2024, 68, 35–49. [Google Scholar] [CrossRef]
- Marquele, F.D.; Stracieri, K.M.; Fonseca, M.J.V.; Freitas, L.A.P. Spray-dried propolis extract. I: Physicochemical and antioxidant properties. Pharmazie 2006, 61, 325–330. [Google Scholar]
- Khawaja, G.; El-Orfali, Y.; Shoujaa, A.; Abou Najem, S. Galangin: A Promising Flavonoid for the Treatment of Rheumatoid Arthritis—Mechanisms, Evidence, and Therapeutic Potential. Pharmaceuticals 2024, 17, 963. [Google Scholar] [CrossRef]
- Spiegel, M. Unveiling the Antioxidative Potential of Galangin: Complete and Detailed Mechanistic Insights through Density Functional Theory Studies. J. Org. Chem. 2024, 89, 8676–8690. [Google Scholar] [CrossRef]
- Spiegel, M. Theoretical Insights into the Oxidative Stress-Relieving Properties of Pinocembrin─An Isolated Flavonoid from Honey and Propolis. J. Phys. Chem. B 2023, 127, 8769–8779. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Song, X.; Li, L.; Sun, J.; Jaiswal, Y.; Huang, J.; Liu, C.; Yang, W.; Williams, L.; Zhang, H.; et al. Protective effects of p-coumaric acid against oxidant and hyperlipidemia-an in vitro and in vivo evaluation. Biomed. Pharmacother. 2019, 111, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Mrówczyńska, L.; Kwaśniewska-Sip, P.; Waśkiewicz, A.; Nowak, P.; Ratajczak, I. Effect of the Solvent on Propolis Phenolic Profile and its Antifungal, Antioxidant, and In Vitro Cytoprotective Activity in Human Erythrocytes Under Oxidative Stress. Molecules 2020, 25, 4266. [Google Scholar] [CrossRef]
- Özkök, A.; Keskin, M.; Tanuğur Samancı, A.E.; Yorulmaz Önder, E.; Takma, Ç. Determination of antioxidant activity and phenolic compounds for basic standardization of Turkish propolis. Appl. Biol. Chem. 2021, 64, 37. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xia, W.; Shao, P.; Wu, W.; Chen, H.; Fang, X.; Mu, H.; Xiao, J.; Gao, H. Impact of thermal processing on dietary flavonoids. Curr. Opin. Food Sci. 2022, 48, 100915. [Google Scholar] [CrossRef]
- Cheng, Y.; Xu, Q.; Liu, J.; Zhao, C.; Xue, F.; Zhao, Y. Decomposition of five phenolic compounds in high temperature water. J. Braz. Chem. Soc. 2014, 25, 2102–2107. [Google Scholar] [CrossRef]
- Abascal, K.; Ganora, L.; Yarnell, E. The effect of freeze-drying and its implications for botanical medicine: A review. Phyther. Res. 2005, 19, 655–660. [Google Scholar] [CrossRef]
- İzol, E.; Bursal, E.; Yapıcı, İ.; Abdullah Yilmaz, M.; Yilmaz, İ.; Gülçin, İ. Chemical Content by LC–MS/MS, Antiglaucoma, and Antioxidant Activity of Propolis Samples from Different Regions of Türkiye. J. Food Biochem. 2024, 2024, 7488590. [Google Scholar] [CrossRef]
- Miraldi, E.; Cappellucci, G.; Baini, G.; Pistone, E.S.; Allodi, M.; Costantino, G.; Spaggiari, C.; Biagi, M. Chemical Markers in Italian Propolis: Chrysin, Galangin and CAPE as Indicators of Geographic Origin. Plants 2024, 13, 2734. [Google Scholar] [CrossRef]
- Bankova, V.; Bertelli, D.; Borba, R.; Conti, B.J.; da Silva Cunha, I.B.; Danert, C.; Eberlin, M.N.; Falcão, S.I.; Isla, M.I.; Moreno, M.I.N.; et al. Standard methods for Apis mellifera propolis research. J. Apic. Res. 2019, 58, 1–49. [Google Scholar] [CrossRef]
- Bocian, A.; Ciszkowicz, E.; Hus, K.K.; Buczkowicz, J.; Lecka-Szlachta, K.; Pietrowska, M.; Petrilla, V.; Petrillova, M.; Legáth, Ľ.; Legáth, J. Antimicrobial activity of protein fraction from Naja ashei venom against Staphylococcus epidermidis. Molecules 2020, 25, 293. [Google Scholar] [CrossRef] [PubMed]
- Murínová, I.; Švidrnoch, M.; Gucký, T.; Hlaváč, J.; Michálek, P.; Slanař, O.; Šíma, M. Population Pharmacokinetic Analysis Proves Superiority of Continuous Infusion in PK/PD Target Attainment with Oxacillin in Staphylococcal Infections. Antibiotics 2022, 11, 1736. [Google Scholar] [CrossRef] [PubMed]
- Wouters, C.L.; Heydarian, N.; Pusavat, J.; Panlilio, H.; Lam, A.K.; Moen, E.L.; Brennan, R.E.; Rice, C.V. Breaking membrane barriers to neutralize E. coli and K. pneumoniae virulence with PEGylated branched polyethylenimine. Biochim. Biophys. Acta Biomembr. 2023, 1865, 184172. [Google Scholar] [CrossRef] [PubMed]
- Nasralddin, N.A.; Haeili, M.; Karimzadeh, S.; Alsahlani, F. Tetracycline and chloramphenicol exposure induce decreased susceptibility to tigecycline and genetic alterations in AcrAB-TolC efflux pump regulators in Escherichia coli and Klebsiella pneumoniae. PLoS ONE 2025, 20, e0315847. [Google Scholar] [CrossRef]
- Hanna, M.; Noor, A. Streptococcus Group B. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025; Available online: https://www.ncbi.nlm.nih.gov/books/NBK553143/ (accessed on 21 March 2025).
- Elbatreek, M.H.; Mahdi, I.; Ouchari, W.; Mahmoud, M.F.; Sobeh, M. Current advances on the therapeutic potential of pinocembrin: An updated review. Biomed. Pharmacother. 2023, 157, 114032. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, J.; Wei, W.; Miao, Y.; Liang, C.; Wu, J.; Huang, X.; Yin, L.; Geng, Y.; Chen, D.; et al. A study of the antibacterial mechanism of pinocembrin against multidrug-resistant Aeromonas hydrophila. Int. Microbiol. 2022, 25, 605–613. [Google Scholar] [CrossRef]
- Klančnik, A.; Šimunović, K.; Kovac, J.; Sahin, O.; Wu, Z.; Vučković, D.; Abram, M.; Zhang, Q.; Možina, S.S. The anti-campylobacter activity and mechanisms of pinocembrin action. Microorganisms 2019, 7, 675. [Google Scholar] [CrossRef]
- Ayamuang, I.O.; Teethaisong, Y.; Sirichaiwetchakoon, K.; Suknasang, S.; Watthana, S.; Chaiseha, Y.; Eumkeb, G. Galangin synergistically revives the antibacterial activity of vancomycin against vancomycin-resistant Enterococcus faecium. J. Appl. Microbiol. 2025, 136, lxaf011. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Lamb, A.J. Detection of galangin-induced cytoplasmic membrane damage in Staphylococcus aureus by measuring potassium loss. J. Ethnopharmacol. 2005, 101, 243–248. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. P-Coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Li, J.; Zhao, N.; Xu, R.; Li, G.; Dong, H.; Wang, B.; Li, Z.; Fan, M.; Wei, X. Deciphering the antibacterial activity and mechanism of p-coumaric acid against Alicyclobacillus acidoterrestris and its application in apple juice. Int. J. Food Microbiol. 2022, 378, 109822. [Google Scholar] [CrossRef]
- Zapała, L.; Ciszkowicz, E.; Kosińska-Pezda, M.; Maciołek, U.; Kozioł, A.E.; Miłoś, A.; Woźnicka, E.; Bocian, A.; Zapała, W.; Rydel-Ciszek, K.; et al. Novel silver(I) complexes with fenamates: Insights into synthesis, spectral characterization, and bioactivity. J. Inorg. Biochem. 2025, 266, 112846. [Google Scholar] [CrossRef]
- Dżugan, M.; Ciszkowicz, E.; Tomczyk, M.; Miłek, M.; Lecka-Szlachta, K. Coniferous Honeydew Honey: Antibacterial Activity and Anti-Migration Properties against Breast Cancer Cell Line (MCF-7). Appl. Sci. 2024, 14, 710. [Google Scholar] [CrossRef]
- Woźnicka, E.; Zapała, L.; Miłoś, A.; Ciszkowicz, E.; Lecka-Szlachta, K.; Woźnicki, P.; Przygórzewska, A.; Kosińska-Pezda, M.; Byczyński, Ł. Synthesis, spectroscopic characterization and biological activities of complexes of light lanthanide ions with 3-hydroxyflavone. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2024, 322, 124870. [Google Scholar] [CrossRef]
- Varadi, M.; Bertoni, D.; Magana, P.; Paramval, U.; Pidruchna, I.; Radhakrishnan, M.; Tsenkov, M.; Nair, S.; Mirdita, M.; Yeo, J.; et al. Alpha Fold Protein Structure Database in 2024. Nucleic Acids Res. 2024, 52, D368–D375. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System, Version 3.0; Schrödinger, LLC.: New York, NY, USA, 2024.
- Steinkellner, G.; Rader, R.; Thallinger, G.G.; Kratky, C.; Gruber, K. VASCo: Computation and visualization of annotated protein surface contacts. BMC Bioinform. 2009, 10, 32. [Google Scholar] [CrossRef]
- Hendlich, M.; Rippmann, F.; Barnickel, G. LIGSITE: Automatic and efficient detection of potential small molecule-binding sites in proteins. J. Mol. Graph. Model. 1997, 15, 359–363. [Google Scholar] [CrossRef]
- Guedes, I.A.; Pereira da Silva, M.M.; Galheigo, M.; Krempser, E.; de Magalhães, C.S.; Correa Barbosa, H.J.; Dardenne, L.E. DockThor-VS: A Free Platform for Receptor-Ligand Virtual Screening. J. Mol. Biol. 2024, 436, 168548. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TPC [mg GAE/g] | TFC [mg QE/g] | TFC/TPC [%] |
|---|---|---|---|
| dEEP1 | 171.13 ± 4.97 bc | 86.41 ± 3.23 a | 50.55 ± 3.18 a |
| dEEP2 | 163.10 ± 8.81 c | 62.60 ± 2.44 b | 38.42 ± 1.68 b |
| dEEP3 | 179.56 ± 7.82 ab | 82.44 ± 3.90 a | 45.91 ± 0.63 a |
| dEEP4 | 188.10 ± 6.15 a | 84.48 ± 5.3 a | 44.91 ± 2.38 a |
| Sample | FRAP [μmol TE/g] | DPPH [μmol TE/g] | CUPRAC [μmol TE/g] |
|---|---|---|---|
| dEEP1 | 587.72 ± 3.31 a | 331.43 ± 20.94 a | 4325.64 ± 174.45 a |
| dEEP2 | 590.35 ± 16.24 a | 288.77 ± 4.10 b | 4146.15 ± 160.62 a |
| dEEP3 | 668.86 ± 31.10 b | 343.94 ± 16.66 a | 4412.82 ± 158.08 a |
| dEEP4 | 627.19 ± 31.31 ab | 329.38 ± 23.16 a | 4169.23 ± 104.06 a |
| Phenolic standards | |||
| Galangin (500 μg/mL) | 738.16 ± 8.00 | 338.20 ± 13.87 | 5887.18 ± 242.81 |
| Pinocembrin (500 μg/mL) | 6.80 ± 1.00 | 21.94 ± 6.16 | 1492.31 ± 149.75 |
| p-Coumaric acid (500 μg/mL) | 331.58 ± 11.24 | 38.15 ± 4.26 | 6838.46 ± 247.95 |
| Sample | Caffeic Acid | p-Coumaric Acid | Ferulic Acid | Benzoic Acid | CAPE | Vanillin | Pinobanksin | Pinocembrin | Galangin |
|---|---|---|---|---|---|---|---|---|---|
| dEEP1 | 1.08 ± 0.05 a | 24.86 ± 1.24 a | 7.66 ± 0.38 a | 10.85 ± 0.54 a | 6.42 ± 0.32 d | 1.61 ± 0.08 a | 4.60 ± 0.23 b | 4.21 ± 0.21 d | 4.17 ± 0.21 c |
| dEEP2 | 2.13 ± 0.11 c | 45.55 ± 2.28 c | 13.16 ± 0.66 c | 14.05 ± 0.70 c | 3.05 ± 0.15 b | 2.59 ± 0.13 d | 4.14 ± 0.21 b | 1.81 ± 0.09 b | 1.10 ± 0.06 a |
| dEEP3 | 1.86 ± 0.09 b | 39.83 ± 1.99 b | 11.33 ± 0.57 b | 10.58 ± 0.53 b | 0.90 ± 0.04 a | 1.92 ± 0.10 b | 2.92 ± 0.15 a | 0.27 ± 0.01 a | 0.92 ± 0.05 a |
| dEEP4 | 2.58 ± 0.13 d | 46.70 ± 2.33 c | 13.52 ± 0.68 c | 12.20 ± 0.61 b | 4.76 ± 0.24 c | 2.28 ± 0.11 c | 3.06 ± 0.15 a | 2.94 ± 0.15 c | 2.79 ± 0.14 b |
| Klebsiella pneumoniae | Streptococcus agalactiae | |
|---|---|---|
| MIC [µg/mL] | ||
| dEEP1 | 625.00 | 78.13 |
| dEEP2 | 625.00 | 156.25 |
| dEEP3 | NAA | 39.06 |
| dEEP4 | NAA | 39.06 |
| p-Coumaric acid | NAA | 125.00 |
| Galangin | NAA | NAA |
| Pinocembrin | 31.25 | 15.62 |
| Chloramphenicol | 7.81 | 1.95 |
| Oxacillin | NAA | 31.25 |
| Streptomycin | 1.95 | 0.49 |
| UniProt ID | Function | Link | AlphaFold Model |
|---|---|---|---|
| P64081 | Enolase OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/P64081 | Yes |
| Q8DX06 | Serine protease, subtilase family OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/Q8DX06/ | No |
| Q8DYR5 | Cell wall surface anchor family protein OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/Q8DYR5/ | Yes |
| Q8E1T8 | Peptidase OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/Q8E1T8/ | Yes |
| Q8DYR9 | Cell wall surface anchor family protein OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/Q8DYR9/ | Yes |
| Q8E0S5 | Cell wall surface anchor family protein, putative OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | https://www.uniprot.org/uniprotkb/Q8E0S5/ | Yes |
| A0A0H3GUG9 | Enolase OS = Klebsiella pneumoniae subsp. pneumoniae | https://www.uniprot.org/uniprotkb/A0A0H3GUG9/ | Yes |
| UniProt ID | Score | Energy | |||
|---|---|---|---|---|---|
| Ligand | Total | Potential | vdW | Electrostatic | |
| A0A0H3GUG9 | Enolase OS = Klebsiella pneumoniae subsp. Pneumoniae | ||||
| galangin | −7.194 | 23.906 | −26.592 | −16.127 | −10.465 |
| pinocembrin | −7.174 | 27.963 | −25.888 | −14.225 | −11.663 |
| p-coumaric acid | −6.567 | −14.429 | −27.390 | −5.051 | −22.339 |
| P64081 | Enolase OS = Streptococcus agalactiae serotype V (strain ATCC BAA-611/2603 V/R) | ||||
| galangin | −7.314 | 24.453 | −28.751 | −7.803 | −20.948 |
| pinocembrin | −7.291 | 29.160 | −27.769 | −10.261 | −17.508 |
| p-coumaric acid | −6.638 | −14.091 | −27.657 | −4.885 | −22.772 |
| Q8E1T8 | Peptidase OS = Streptococcus agalactiae serotype V (strain ATCC BAA-/2603 V/R) | ||||
| galangin | −7.606 | 32.579 | −21.420 | −16.787 | −4.633 |
| pinocembrin | −7.176 | 28.490 | −21.470 | −3.997 | −17.473 |
| p-coumaric acid | −7.105 | −11.272 | −23.826 | −10.08 | −13.746 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dżugan, M.; Miłek, M.; Ciszkowicz, E.; Łyskowski, A.; Tomczyk, M. Selected Polyphenols of Polish Poplar Propolis as a Key Component Shaping Its Antibacterial Properties—In Vitro and In Silico Approaches. Molecules 2025, 30, 2036. https://doi.org/10.3390/molecules30092036
Dżugan M, Miłek M, Ciszkowicz E, Łyskowski A, Tomczyk M. Selected Polyphenols of Polish Poplar Propolis as a Key Component Shaping Its Antibacterial Properties—In Vitro and In Silico Approaches. Molecules. 2025; 30(9):2036. https://doi.org/10.3390/molecules30092036
Chicago/Turabian StyleDżugan, Małgorzata, Michał Miłek, Ewa Ciszkowicz, Andrzej Łyskowski, and Monika Tomczyk. 2025. "Selected Polyphenols of Polish Poplar Propolis as a Key Component Shaping Its Antibacterial Properties—In Vitro and In Silico Approaches" Molecules 30, no. 9: 2036. https://doi.org/10.3390/molecules30092036
APA StyleDżugan, M., Miłek, M., Ciszkowicz, E., Łyskowski, A., & Tomczyk, M. (2025). Selected Polyphenols of Polish Poplar Propolis as a Key Component Shaping Its Antibacterial Properties—In Vitro and In Silico Approaches. Molecules, 30(9), 2036. https://doi.org/10.3390/molecules30092036