
Antioxidant and Anticancer Mechanisms of Unique Polyphenols in Camellia ptilophylla: Focus on Gallocatechin-3,5-di-O-gallate and 1,2,4,6-Tetra-O-galloyl-β-D-glucopyranose

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Oxidative Stress Pathways Involved in Three Major Cancers
2.2. Cellular Protection Against Oxidative Stress and Anticancer Activity of Tea Extracts
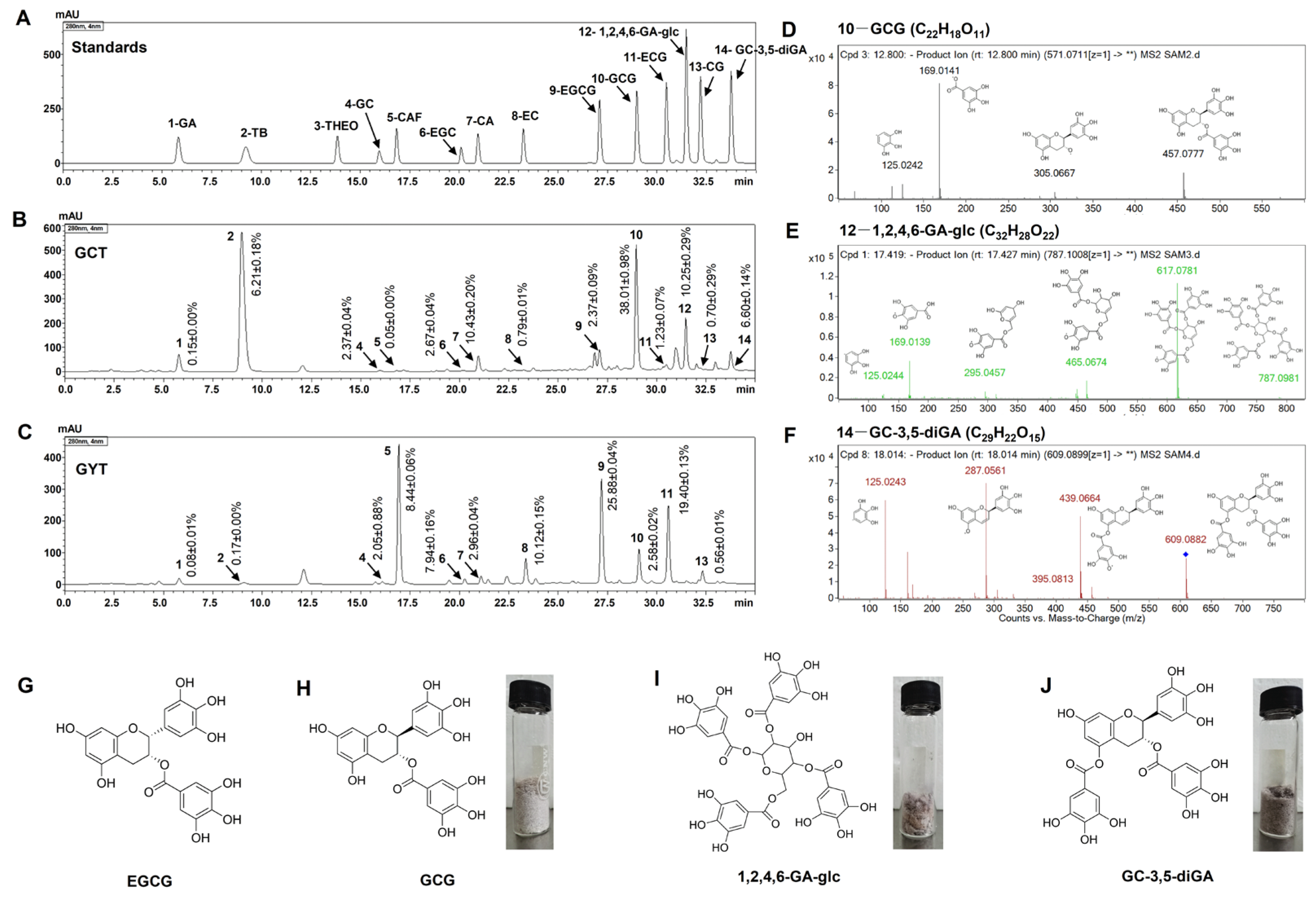
2.3. Major Chemical Compositions of GCT and GYT
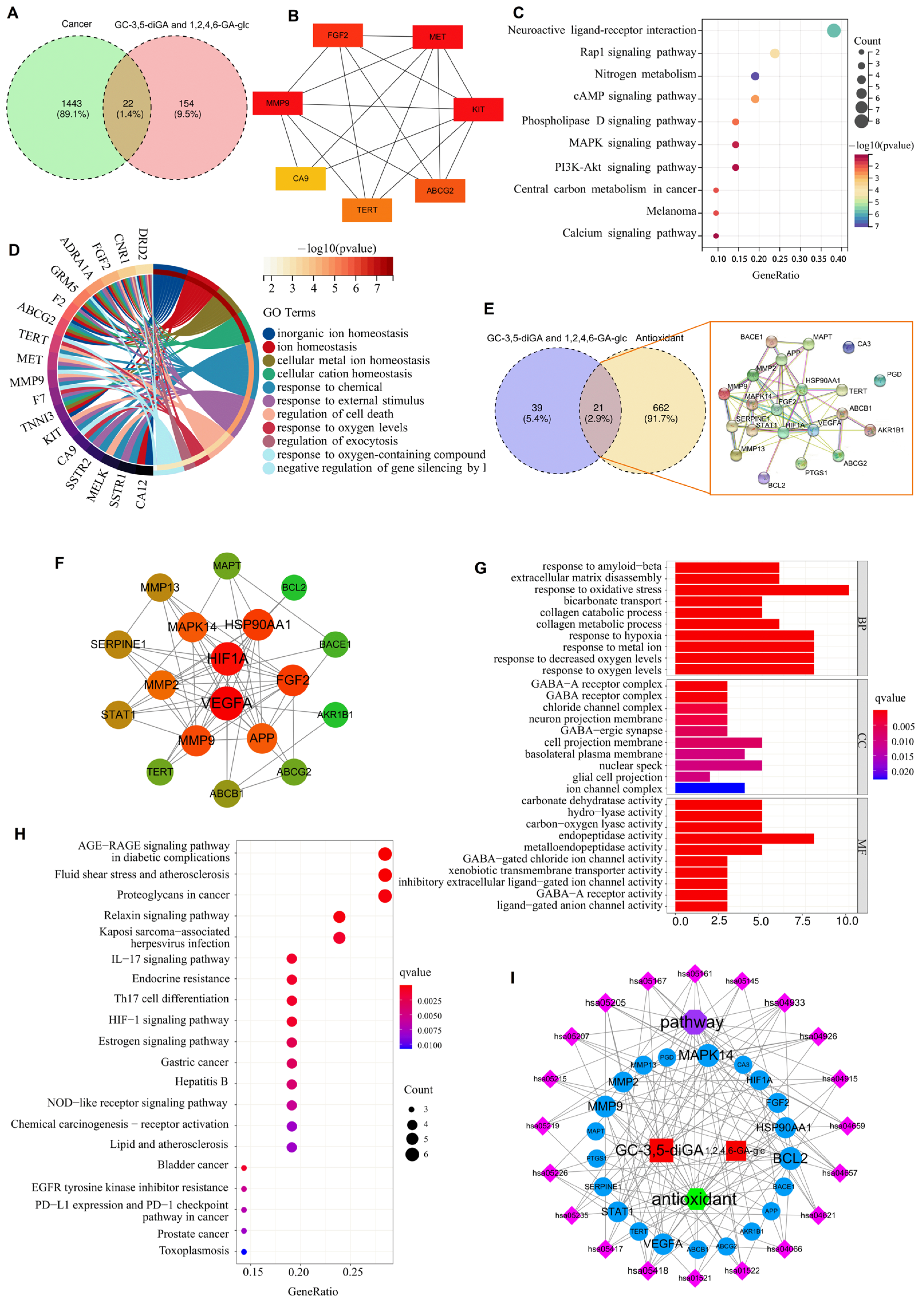
2.4. Mechanism Prediction of the Effects of GC-3,5-diGA and 1,2,4,6-GA-glc on Cancers and Oxidative Stress
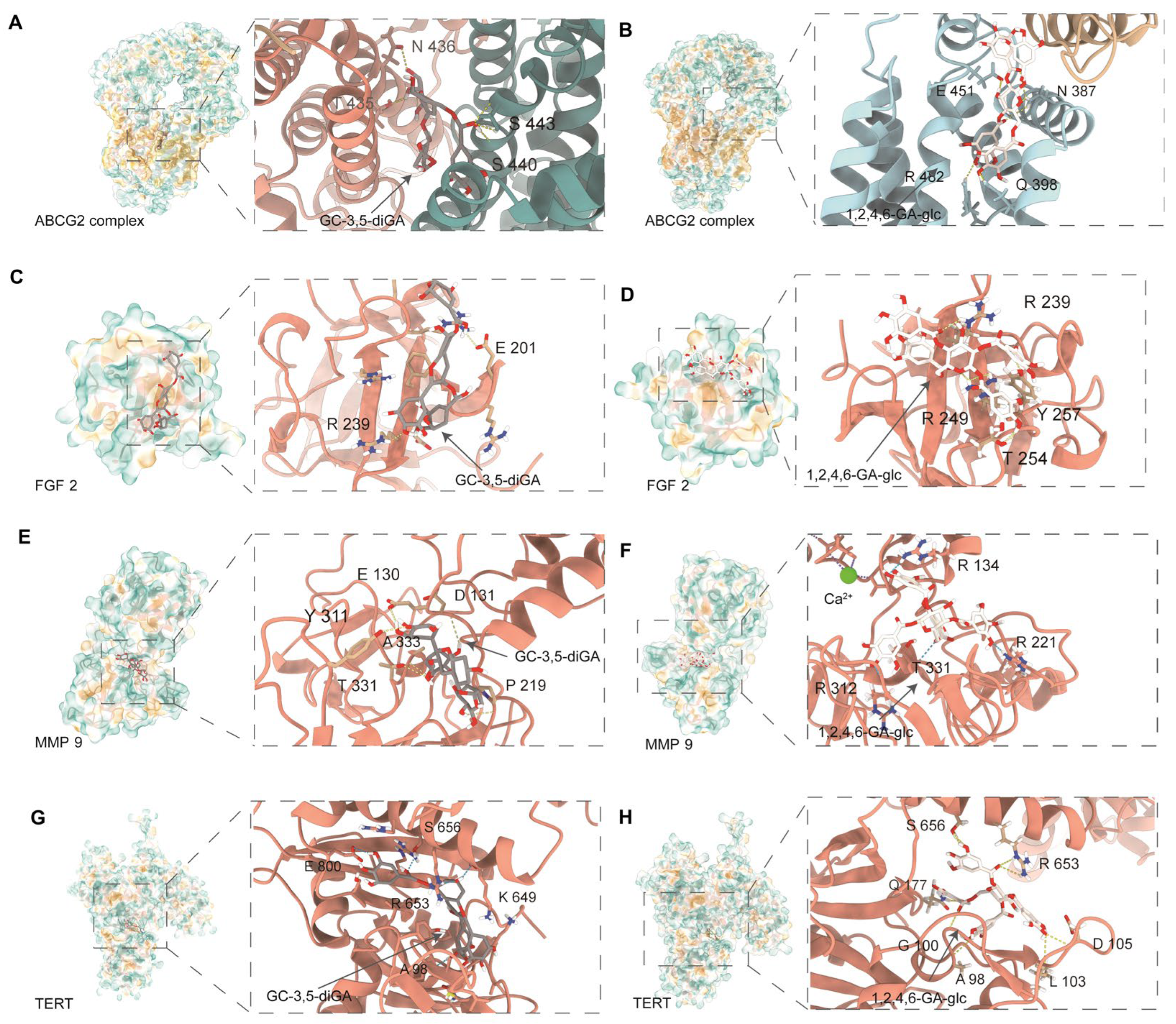
2.5. Molecular Docking of GC-3,5-diGA and 1,2,4,6-GA-glc with Shared Targets Related to Cancer and Oxidative Stress
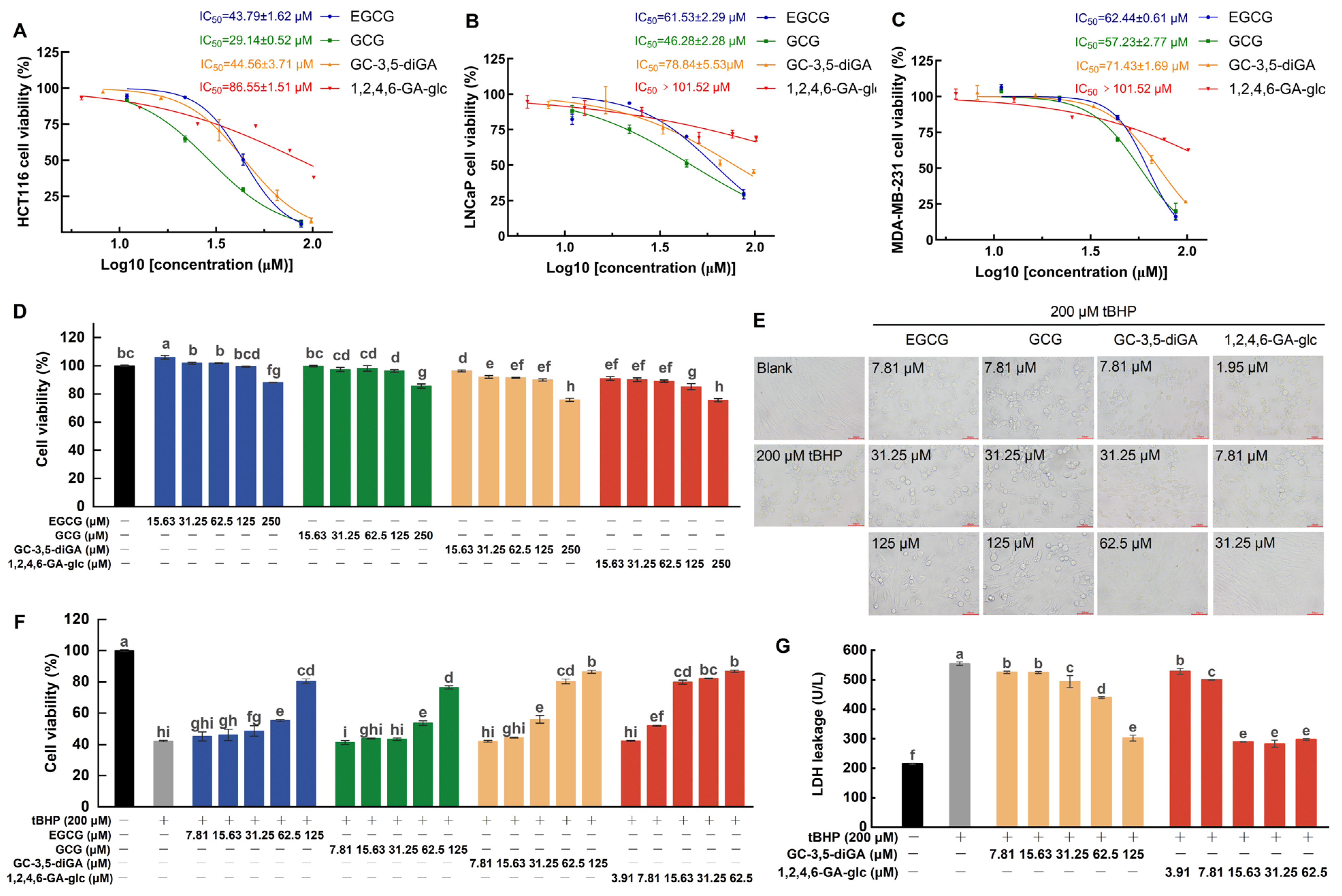
2.6. Cellular Protection Against Oxidative Stress and Anticancer Activities of GC-3,5-diGA and 1,2,4,6-GA-glc
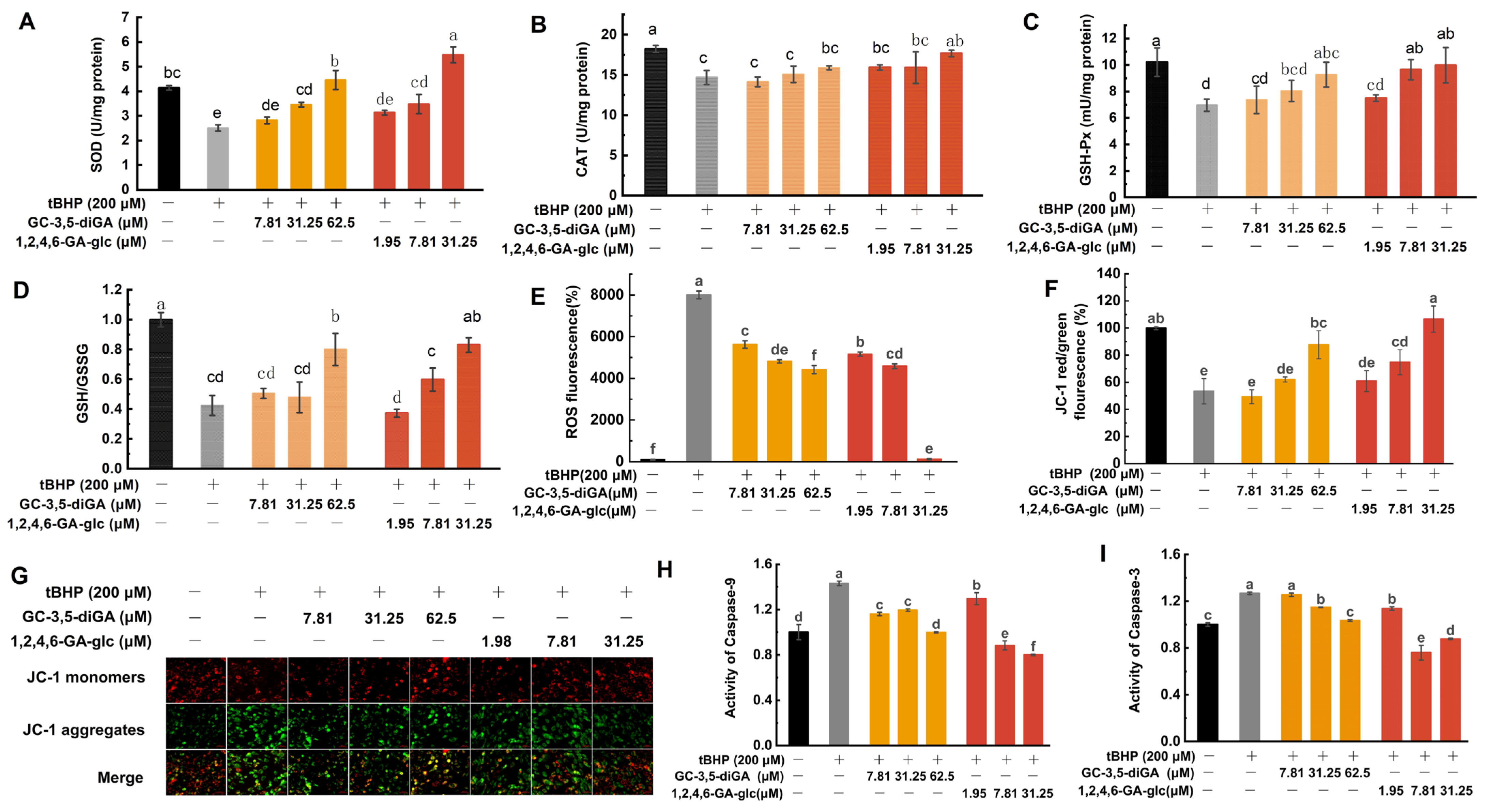
2.7. Cellular Protection Mechanisms Against Oxidative Stress Mediated by GC-3,5-diGA and 1,2,4,6-GA-glc
3. Discussion
4. Materials and Methods
4.1. Materials and Chemical Reagents
4.2. Preparation and Chemical Analysis of Tea Extracts
4.3. Isolation and Identification of GCG, GC-3,5-diGA, and 1,2,4,6-GA-glc from GCT
4.4. Difference Expression Analysis and Enrichment Analysis of Cancer-Related Genes
4.5. Analysis of GC-3,5-diGA and 1,2,4,6-GA-glc Targets with Cancer-Related Genes and Antioxidant-Related Genes
4.6. Molecular Docking
4.7. Cell Culture and Cell Viability Assay
4.8. Determination of SOD, CAT, GSH-Px, GSH, and GSSG Levels
4.9. Measurement of Intracellular ROS
4.10. MMP Assay
4.11. Caspase-3 and Caspase-9 Activation Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Li, K.; Deng, Z.; Lei, C.; Ding, X.; Li, J.; Wang, C. The Role of Oxidative Stress in Tumorigenesis and Progression. Cells 2024, 13, 441. [Google Scholar] [CrossRef]
- Klaunig, J. Oxidative Stress and Cancer. Curr. Pharm. Des. 2019, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.; Dinkova-Kostova, A.; Tew, K. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Az-Zahra, F.; Wongso, H.; Setyawati, L.; Novitasari, D.; Ikram, E. Molecular Mechanism of Natural Food Antioxidants to Regulate ROS in Treating Cancer: A Review. Antioxidants 2024, 13, 207. [Google Scholar] [CrossRef]
- Mileo, A.; Miccadei, S. Polyphenols as Modulator of Oxidative Stress in Cancer Disease: New Therapeutic Strategies. Oxidative Med. Cell. Longev. 2015, 2016, 6475624. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Panda, C. Molecular aspects of cancer chemopreventive and therapeutic efficacies of tea and tea polyphenols. Nutrition 2017, 43–44, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Gupta, P.; Ghosh, S.; Choudhury, S.; Das, A.; Ahir, M.; Adhikary, A.; Chattopadhyay, S. Targeted tumor killing by pomegranate polyphenols: Pro-oxidant role of a classical antioxidant. J. Nutr. Biochem. 2023, 115, 109283. [Google Scholar] [CrossRef]
- Sandur, S.K.; Ichikawa, H.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Sethi, G.; Aggarwal, B.B. Role of pro-oxidants and antioxidants in the anti-inflammatory and apoptotic effects of curcumin (diferuloylmethane). Free Radic. Bio. Med. 2007, 43, 568–580. [Google Scholar] [CrossRef]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K. ROS-Mediated Cancer cell killing through dietary phytochemicals. Oxidative Med. Cell. Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Trachootham, D.; Zhou, Y.; Zhang, H.; Demizu, Y.; Chen, Z.; Chiao, P.J.; Achanta, G.; Arlinghaus, R.B.; Liu, J.; Pelicano, H. Selective killing of oncogenically transformed cells through a ROS-mediated mechanism by β-phenylethyl isothiocyanate. Cancer Cell. 2006, 10, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Ye, C.; Zhang, R.; Ma, Y.; Zhang, P. A discovery of new tea resource—cocoa tea tree containing theobromine from China. Acta Sci. Nat. Univ. Sunyatseni 1988, 27, 131–133. [Google Scholar]
- Peng, L.; Wang, X.; Shi, X.; Li, C.; Ye, C.; Song, X. Characterization of the constituents and antioxidative activity of cocoa tea (Camellia ptilophylla). Food Chem. 2011, 129, 1475–1482. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, L.; Xuan, J.; Yang, L.; Zhao, T.; Peng, W. Tea for histamine anti-allergy: Component analysis of tea extracts and potential mechanism for treating histamine anti-allergy. Front. Pharmacol. 2024, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Jia, Y.; Hu, T.; Du, J.; Wang, Y.; Cheng, B.; Li, K. GC-(4→8)-GCG, A proanthocyanidin dimer from Camellia ptilophylla, modulates obesity and adipose tissue inflammation in high-fat diet induced obese mice. Mol. Nutr. Food Res. 2019, 63, 1900082. [Google Scholar] [CrossRef]
- Gao, X.; Li, X.; Ho, C.; Lin, X.; Zhang, Y.; Li, B.; Chen, Z. Cocoa tea (Camellia ptilophylla) induces mitochondria-dependent apoptosis in HCT116 cells via ROS generation and PI3K/Akt signaling pathway. Food Res Int. 2020, 129, 108854. [Google Scholar] [CrossRef]
- Peng, L.; Khan, N.; Afaq, F.; Ye, C.; Mukhtar, H. In vitro and in vivo effects of water extract of white cocoa tea (Camellia ptilophylla) against human prostate cancer. Pharm. Res. 2010, 27, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, C.; Tam, J.; Kwok, H.; Lau, C.; Leung, P.; Ko, C.; Ye, C. In vitro and in vivo mechanistic study of a novel proanthocyanidin, GC-(4→8)-GCG from cocoa tea (Camellia ptilophylla) in antiangiogenesis. J. Nutr. Biochem. 2014, 25, 19–28. [Google Scholar] [CrossRef]
- Kurihara, H.; Shibata, H.; Fukui, Y.; Kiso, Y.; Xu, J.K.; Yao, X.S.; Fukami, H. Evaluation of the hypolipemic property of Camellia sinensis var. ptilophylla on postprandial hypertriglyceridemia. J. Agr. Food Chem. 2006, 54, 4977–4981. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef]
- Piao, M.J.; Kang, K.A.; Zhang, R.; Ko, D.O.; Wang, Z.H.; Lee, K.H.; Chang, W.Y.; Chae, S.; Jee, Y.; Shin, T.; et al. Antioxidant properties of 1,2,3,4,6-penta-O-galloyl-b-D-glucose from Elaeocarpus sylvestris var. Ellipticus. Food Chem. 2009, 115, 412–418. [Google Scholar] [CrossRef]
- Kuang, X.; Gao, X.; Lin, X.; Chen, Z.; Li, B.; Zhang, Y. Isolation, purification and antioxidant activities of three kinds of polyphenol monomers from Camellia ptilophylla Chang. Sci. Technol. Food Ind. 2020, 41, 31–39. [Google Scholar]
- Chen, D.; Wu, Z.; Wu, L.; Jiang, J.; Hu, G. Theaflavin attenuates TBHP-induced endothelial cells oxidative stress by activating PI3K/AKT/Nrf2 and accelerates wound healing in rats. Front. Bioeng. Biotech. 2022, 10, 830574. [Google Scholar] [CrossRef] [PubMed]
- Asha Parveen, S.M.; Kami Reddy, K.R.; Ummanni, R. Dimethylarginine Dimethylaminohydrolase-1 expression is increased under tBHP-induced oxidative stress regulates nitric oxide production in PCa cells attenuates mitochondrial ROS-mediated apoptosis. Nitric Oxide 2023, 138–139, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wu, Y.; Jiang, G.; Feng, L.; Wang, Z.; Yuan, G.; Tong, H. Sulfated polysaccharides from Rhodiola sachalinensis reduce D-gal-induced oxidative stress in NIH3T3 cells. Int. J. Biol. Macromol. 2019, 140, 288–293. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Z.; Lin, X.; Zhang, Y.; Zhou, L.; Gao, X.; Li, B. Protective Effect of Camellia ptilophylla Chang on the tBHP-Induced NIH3T3 Cells. Sci. Technol. Food Ind. 2021, 42, 90–98. [Google Scholar]
- Yin, Z.; Ivanov, V.; Habelhah, H.; Tew, K.; Ronai, Z. Glutathione S-transferase p elicits protection against H2O2-induced cell death via coordinated regulation of stress kinases. Cancer Res. 2000, 60, 4053–4057. [Google Scholar]
- Ying, S.; Huang, Q.; Lu, S.; Gao, X.; Chen, Z.; Zhang, Y.; Liu, X.; Li, B.; Lin, X. Comparative quantication of 14 bioactive compounds in Camellia ptilophylla and Camellia sinensis. Res. Sq. 2022. preprint. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Ighodaro, O.; Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Sandoval-Acuña, C.; Ferreira, J.; Speisky, H. Polyphenols and mitochondria: An update on their increasingly emerging ROS-scavenging independent actions. Arch. Biochem. Biophys. 2014, 559, 75–90. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jin, P.; Guan, Y.; Luo, M.; Wang, Y.; He, B.; Li, B.; He, K.; Cao, J.; Huang, C.; et al. Exploiting polyphenol-mediated redox reorientation in cancer therapy. Pharmaceuticals 2022, 15, 1540. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Jiang, J.; Zhou, L.; Huang, Z.; Nice, E.C.; Huang, C.; Fu, L. Mitochondrial adaptation in cancer drug resistance: Prevalence, mechanisms, and management. J. Hematol. Oncol. 2022, 15, 97. [Google Scholar] [CrossRef]
- Neganova, M.; Liu, J.; Aleksandrova, Y.; Klochkov, S.; Fan, R. Therapeutic Influence on Important Targets Associated with Chronic Inflammation and Oxidative Stress in Cancer Treatment. Cancers 2021, 13, 6062. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Wang, S.; Cui, W.; Pan, D.; Jiang, M.; Chang, B.; Sang, L. Tea polyphenols and their chemopreventive and therapeutic effects on colorectal cancer. World J. Gastroenterol. 2020, 26, 562–597. [Google Scholar] [CrossRef]
- Allahmoradi, H.; Asghari, S.; Ahmadi, A.; Assareh, E.; Nazari, M. Anti-tumor and anti-metastatic activity of the FGF2 118-126 fragment dependent on the loop structure. Biochem. J. 2022, 479, 1285–1302. [Google Scholar] [CrossRef]
- Yu, Y.; Bai, F.; Liu, Y.; Yang, Y.; Yuan, Q.; Zou, D.; Qu, S.; Tian, G.; Song, L.; Zhang, T.; et al. Fibroblast growth factor (FGF21) protects mouse liver against d-galactose-induced oxidative stress and apoptosis via activating Nrf2 and PI3K/Akt pathways. Mol. Cell. Biochem. 2015, 403, 287–299. [Google Scholar] [CrossRef]
- Marinaccio, J.; Micheli, E.; Udroiu, I.; Di Nottia, M.; Carrozzo, R.; Baranzini, N.; Grimaldi, A.; Leone, S.; Moreno, S.; Muzzi, M.; et al. TERT Extra-Telomeric Roles: Antioxidant Activity and Mitochondrial Protection. Int. J. Mol. Sci. 2023, 24, 4450. [Google Scholar] [CrossRef] [PubMed]
- Augoff, K.; Hryniewicz-Jankowska, A.; Tabola, R.; Stach, K. MMP9: A Tough Target for Targeted Therapy for Cancer. Cancers 2022, 14, 1847. [Google Scholar] [CrossRef]
- Walter, L.; Pujada, A.; Bhatnagar, N.; Bialkowska, A.; Yang, V.; Laroui, H.; Garg, P. Epithelial derived-matrix metalloproteinase (MMP9) exhibits a novel defensive role of tumor suppressor in colitis associated cancer by activating MMP9-Notch1-ARF-p53 axis. Oncotarget 2017, 8, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Fan, Y.; Cai, C.; Wang, J.; Teng, Q.; Lei, Z.; Zeng, L.; Gupta, P.; Chen, Z. Olmutinib (BI1482694/HM61713), a Novel Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor, Reverses ABCG2-Mediated Multidrug Resistance in Cancer Cells. Front. Pharmacol. 2018, 9, 1097. [Google Scholar] [CrossRef]
- Xu, J.; Yeung, S.; Chang, Q.; Huang, Y.; Chen, Z. Comparison of antioxidant activity and bioavailability of tea epicatechins with their epimers. Brit. J. Nutr. 2004, 91, 873–881. [Google Scholar] [PubMed]
- Wright, J.; Johnson, E.; DiLabio, G. Predicting the activity of phenolic antioxidants: Theoretical method, analysis of substituent effects, and application to major families of antioxidants. J. Am. Chem. Soc. 2001, 6, 1173–1183. [Google Scholar] [CrossRef]
- Li, X.; Xie, H.; Zhan, R.; Chen, D. Effect of Double Bond Position on 2-Phenyl-benzofuran Antioxidants: A Comparative Study of Moracin C and Iso-Moracin C. Molecules 2018, 23, 754. [Google Scholar] [CrossRef]
- Lin, J.; Li, X.; Chen, B.; Wei, G.; Chen, D. E-Configuration Improves Antioxidant and Cytoprotective Capacities of Resveratrols. Molecules 2018, 23, 1790. [Google Scholar] [CrossRef]
- Minnelli, C.; Galeazzi, R.; Laudadio, E.; Amici, A.; Rusciano, D.; Armeni, T.; Cantarini, M.; Stipa, P.; Mobbili, G. Monoalkylated Epigallocatechin-3-gallate (C18-EGCG) as Novel Lipophilic EGCG Derivative: Characterization and Antioxidant Evaluation. Antioxidants 2020, 9, 208. [Google Scholar] [CrossRef]
- Li, X.; Xie, Y.; Xie, H.; Yang, J.; Chen, D. π-π Conjugation Enhances Oligostilbene’s Antioxidant Capacity: Evidence from α-Viniferin and Caraphenol A. Molecules 2018, 23, 694. [Google Scholar] [CrossRef]
- Cotelle, N.; Hapiot, P.; Pinson, J.; Rolando, C.; Vezin, H. Polyphenols deriving from chalcones: Investigations of redox activities. J. Phys. Chem. B 2005, 49, 23720–23729. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Li, M.; Wang, F.; Mackenzie, G.; Oteiza, P. The inhibitory effect of ECG and EGCG dimeric procyanidins on colorectal cancer cells growth is associated with their actions at lipid rafts and the inhibition of the epidermal growth factor receptor signaling. Biochem. Pharmacol. 2020, 175, 113923. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhao, B.; Shen, S.; Hou, J.; Hu, J.; Xin, W. ESR study on the structure-antioxidant activity relationship of tea catechins and their epimers. Biochim. Biophys. Acta 1999, 1427, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Sayin, V.; Ibrahim, M.; Larsson, E.; Nilsson, J.; Lindahl, P.; Bergo, M. Antioxidants Accelerate Lung Cancer Progression in Mice. Sci. Transl. Med. 2014, 6, 221ra15. [Google Scholar] [CrossRef] [PubMed]
- Min, N.; Kim, J.; Choi, J.; Liang, W.; Ko, Y.; Rhee, S.; Bang, H.; Ham, S.; Park, A.; Lee, K. Selective death of cancer cells by preferential induction of reactive oxygen species in response to (-)-epigallocatechin-3-gallate. Biochem. Biophys. Res. Commun. 2012, 421, 91–97. [Google Scholar] [CrossRef]
- Qanungo, S.; Das, M.; Haldar, S.; Basu, A. Epigallocatechin-3-gallate induces mitochondrial membrane depolarization and caspase-dependent apoptosis in pancreatic cancer cells. Carcinogenesis 2005, 26, 958–967. [Google Scholar] [CrossRef]
- Tomczak, K.; Czerwińska, P.; Wiznerowicz, M. The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge. Contemp. Oncol. 2015, 19, A68–A77. [Google Scholar] [CrossRef]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.; Morris, J.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Li, J.; Shen, S.; Xu, H. AlphaFold3 versus experimental structures: Assessment of the accuracy in ligand-bound G protein-coupled receptors. Acta. Pharmacol Sin. 2024, 46, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Lu, S.; Gao, X.; Chen, Z.; Zhang, Y.; Zhong, W.; Zhu, F.; Li, B.; Lin, X. Antioxidant and Anticancer Mechanisms of Unique Polyphenols in Camellia ptilophylla: Focus on Gallocatechin-3,5-di-O-gallate and 1,2,4,6-Tetra-O-galloyl-β-D-glucopyranose. Molecules 2025, 30, 1919. https://doi.org/10.3390/molecules30091919
Zhou L, Lu S, Gao X, Chen Z, Zhang Y, Zhong W, Zhu F, Li B, Lin X. Antioxidant and Anticancer Mechanisms of Unique Polyphenols in Camellia ptilophylla: Focus on Gallocatechin-3,5-di-O-gallate and 1,2,4,6-Tetra-O-galloyl-β-D-glucopyranose. Molecules. 2025; 30(9):1919. https://doi.org/10.3390/molecules30091919
Chicago/Turabian StyleZhou, Langhua, Sen Lu, Xiong Gao, Zhongzheng Chen, Yuanyuan Zhang, Weixia Zhong, Fuming Zhu, Bin Li, and Xiaorong Lin. 2025. "Antioxidant and Anticancer Mechanisms of Unique Polyphenols in Camellia ptilophylla: Focus on Gallocatechin-3,5-di-O-gallate and 1,2,4,6-Tetra-O-galloyl-β-D-glucopyranose" Molecules 30, no. 9: 1919. https://doi.org/10.3390/molecules30091919
APA StyleZhou, L., Lu, S., Gao, X., Chen, Z., Zhang, Y., Zhong, W., Zhu, F., Li, B., & Lin, X. (2025). Antioxidant and Anticancer Mechanisms of Unique Polyphenols in Camellia ptilophylla: Focus on Gallocatechin-3,5-di-O-gallate and 1,2,4,6-Tetra-O-galloyl-β-D-glucopyranose. Molecules, 30(9), 1919. https://doi.org/10.3390/molecules30091919