

Pinitol Improves Lipopolysaccharide-Induced Cellular Damage in Human Dermal Microvascular Endothelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
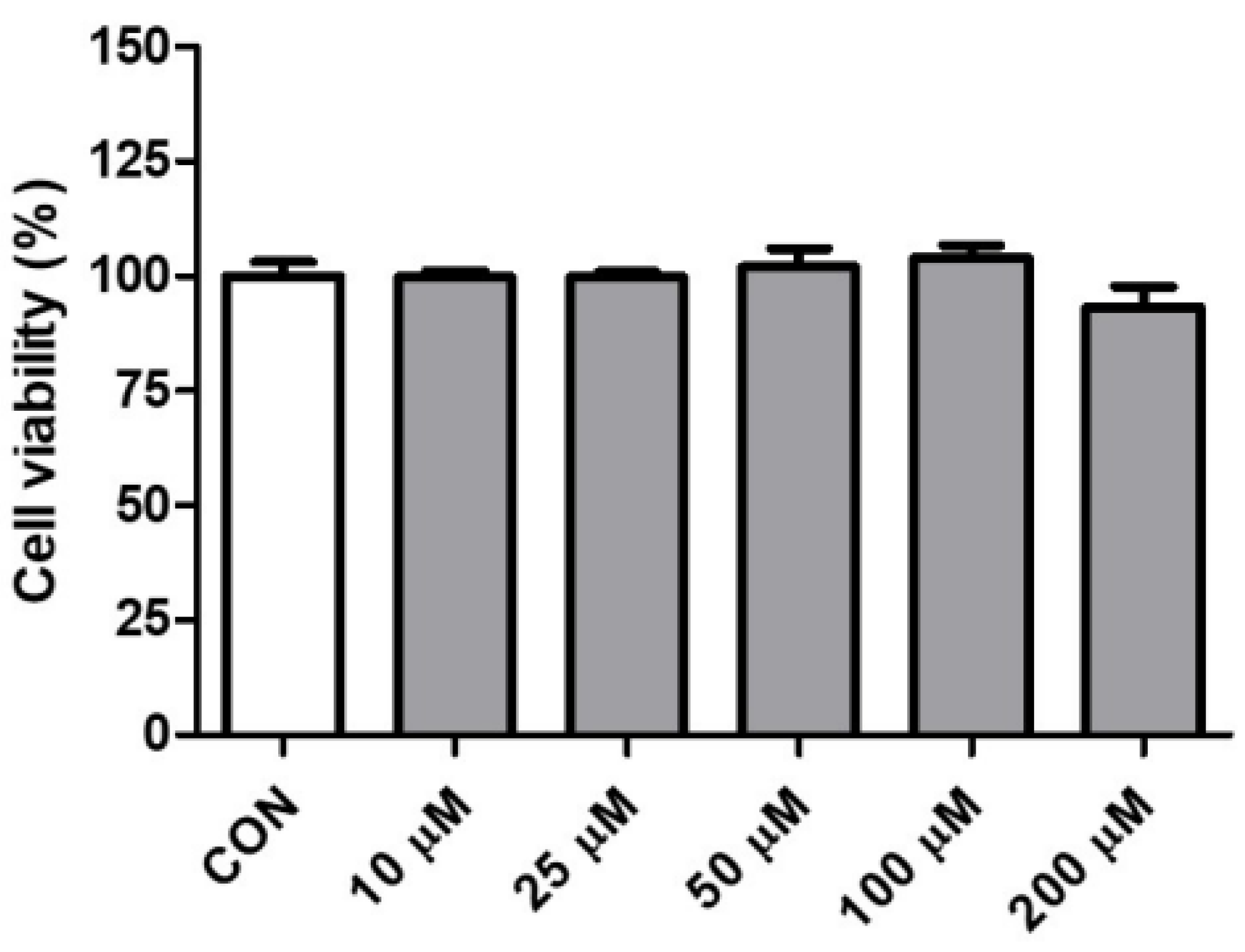
2.1. Effect of Pinitol on the Cell Viability in HDMECs
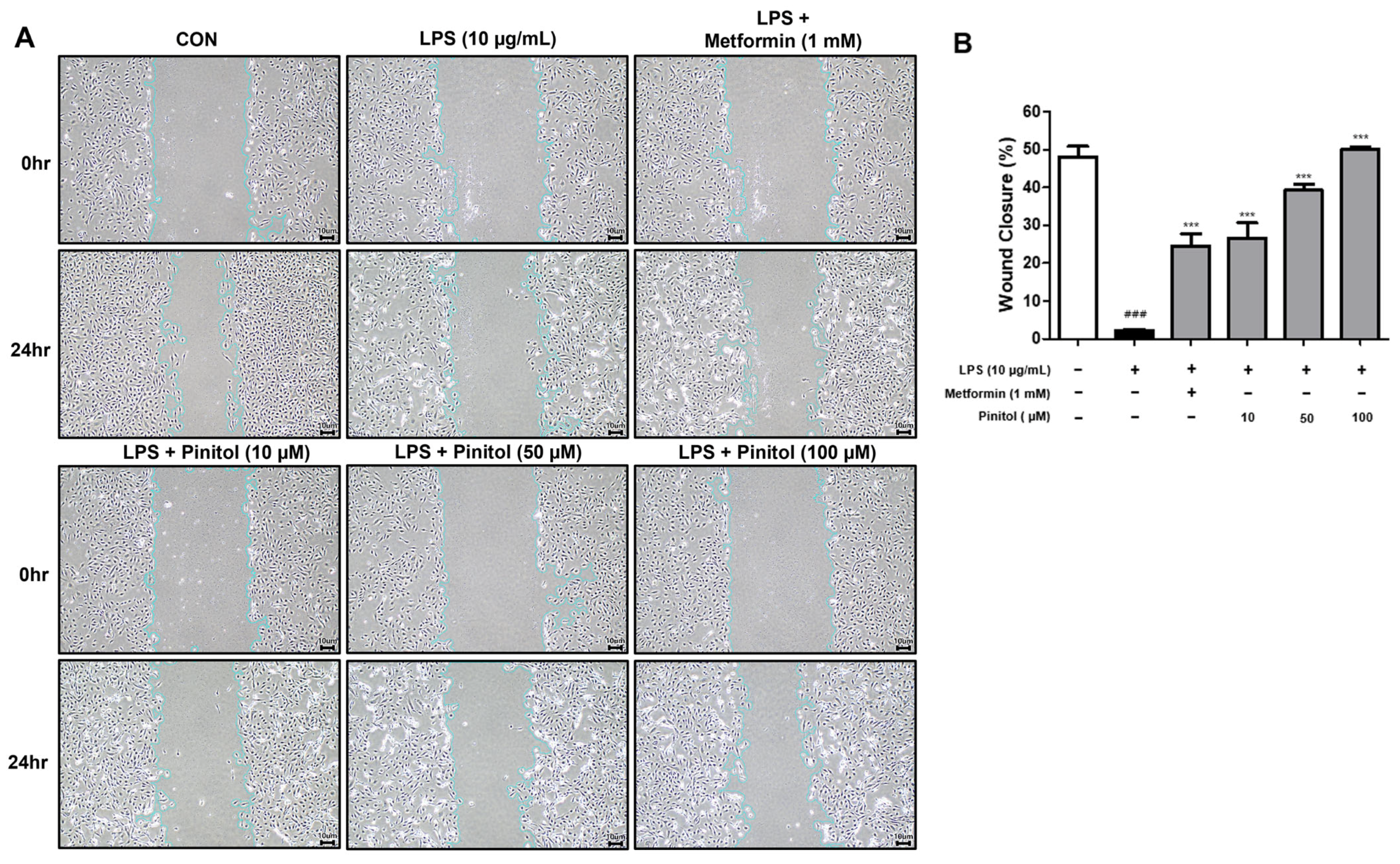
2.2. Pinitol Improved Wound Healing in LPS-Damaged HDMECs
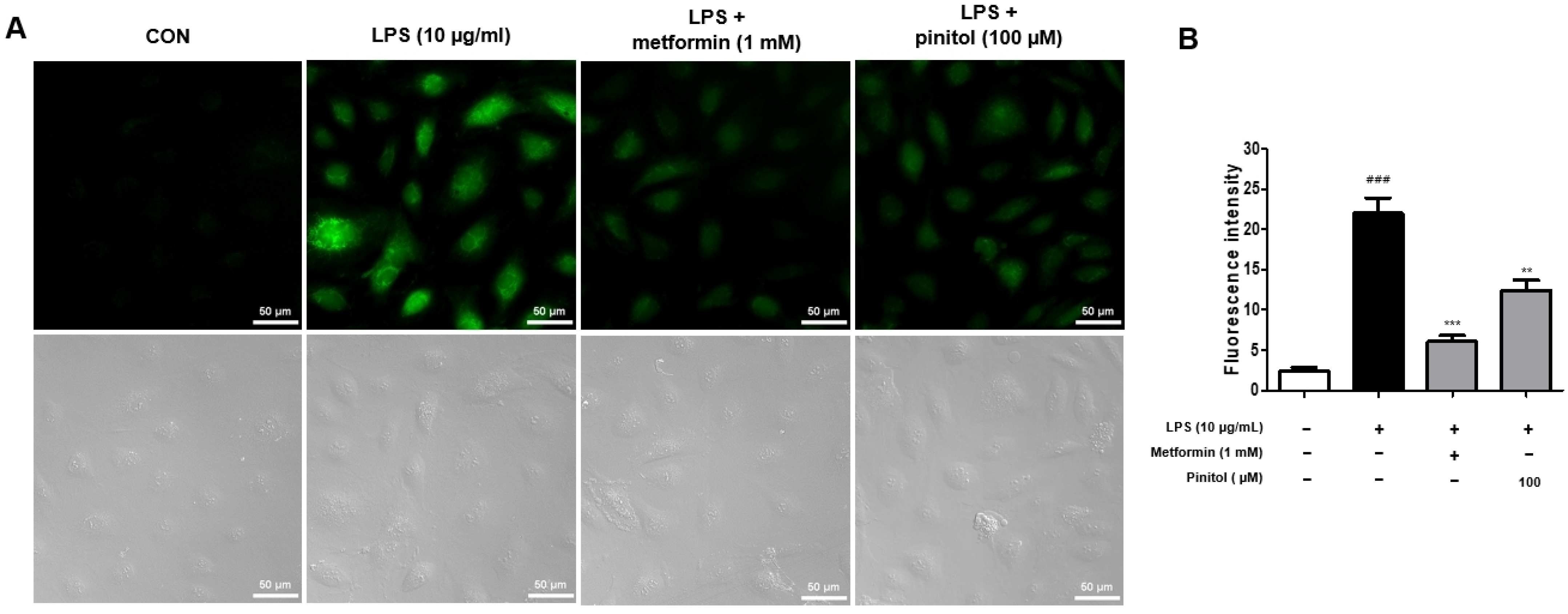
2.3. Pinitol Recovered ROS Levels in LPS-Damaged HDMECs
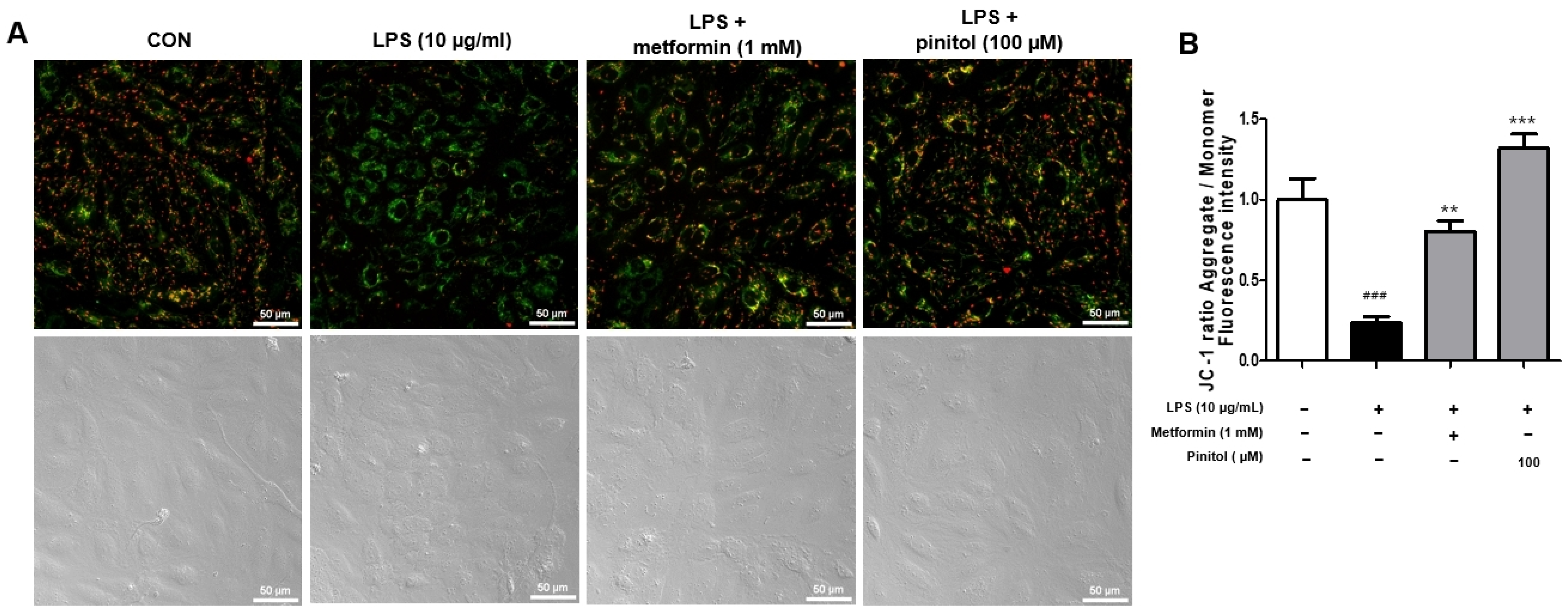
2.4. Pinitol Ameliorated the Membrane Potential of Mitochondria in LPS-Damaged HDMECs
2.5. Pinitol Inhibited NF-κB Pathway in LPS-Damaged HDMECs
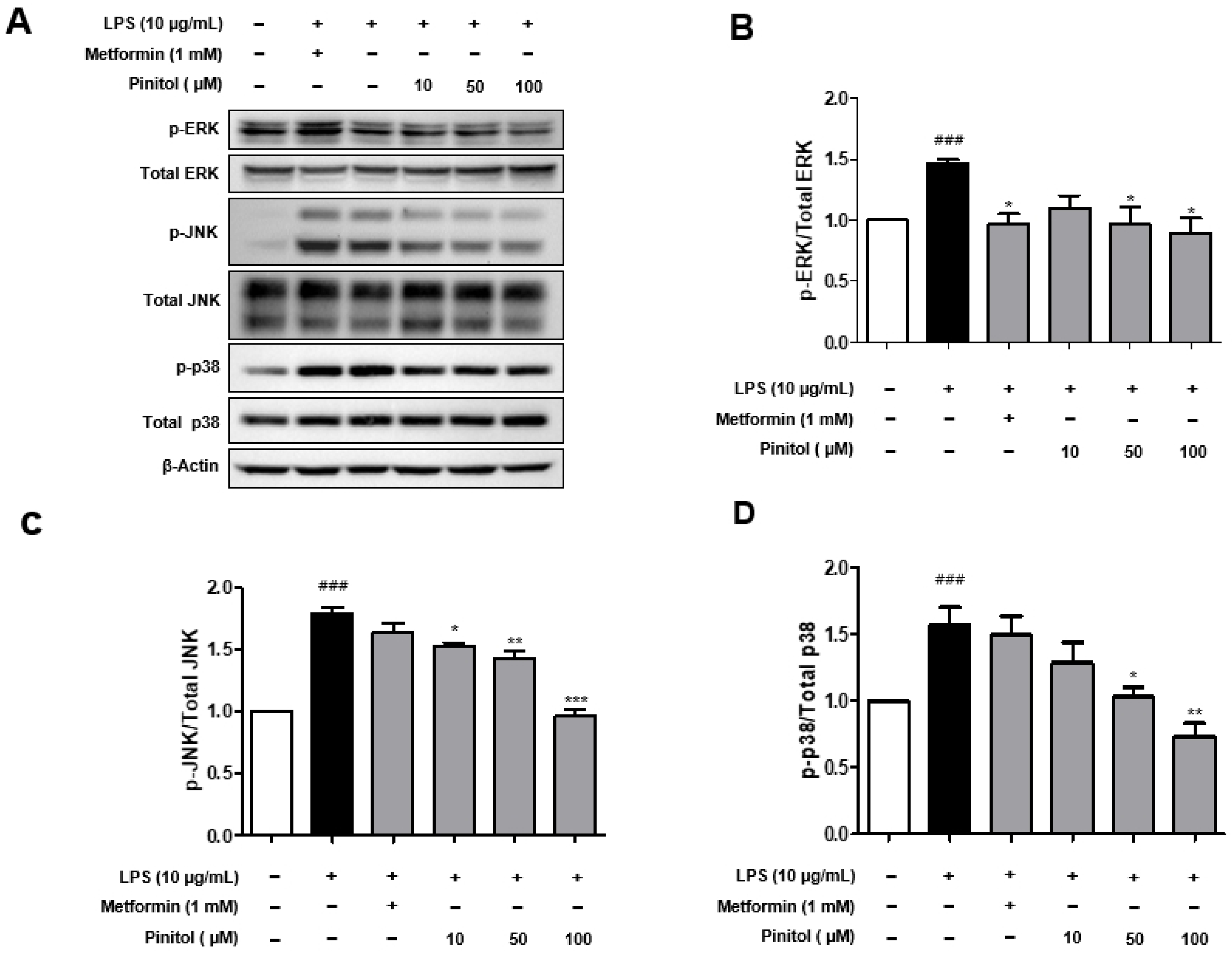
2.6. Pinitol Significantly Decreased the Phosphorylation Levels of MAPKs in LPS-Damaged HDMECs
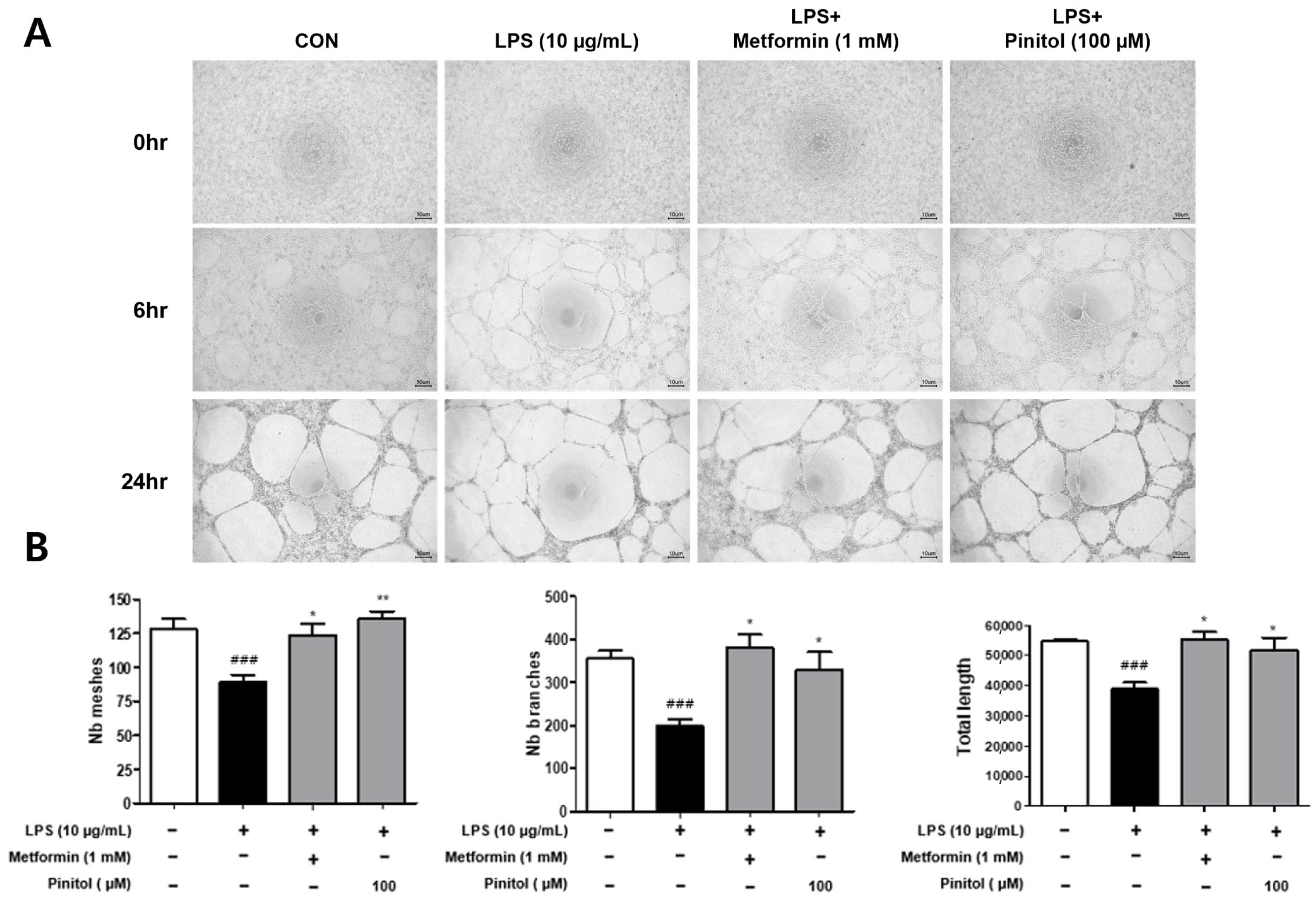
2.7. Pinitol Improved Tube Formation in LPS-Damaged HDMECs
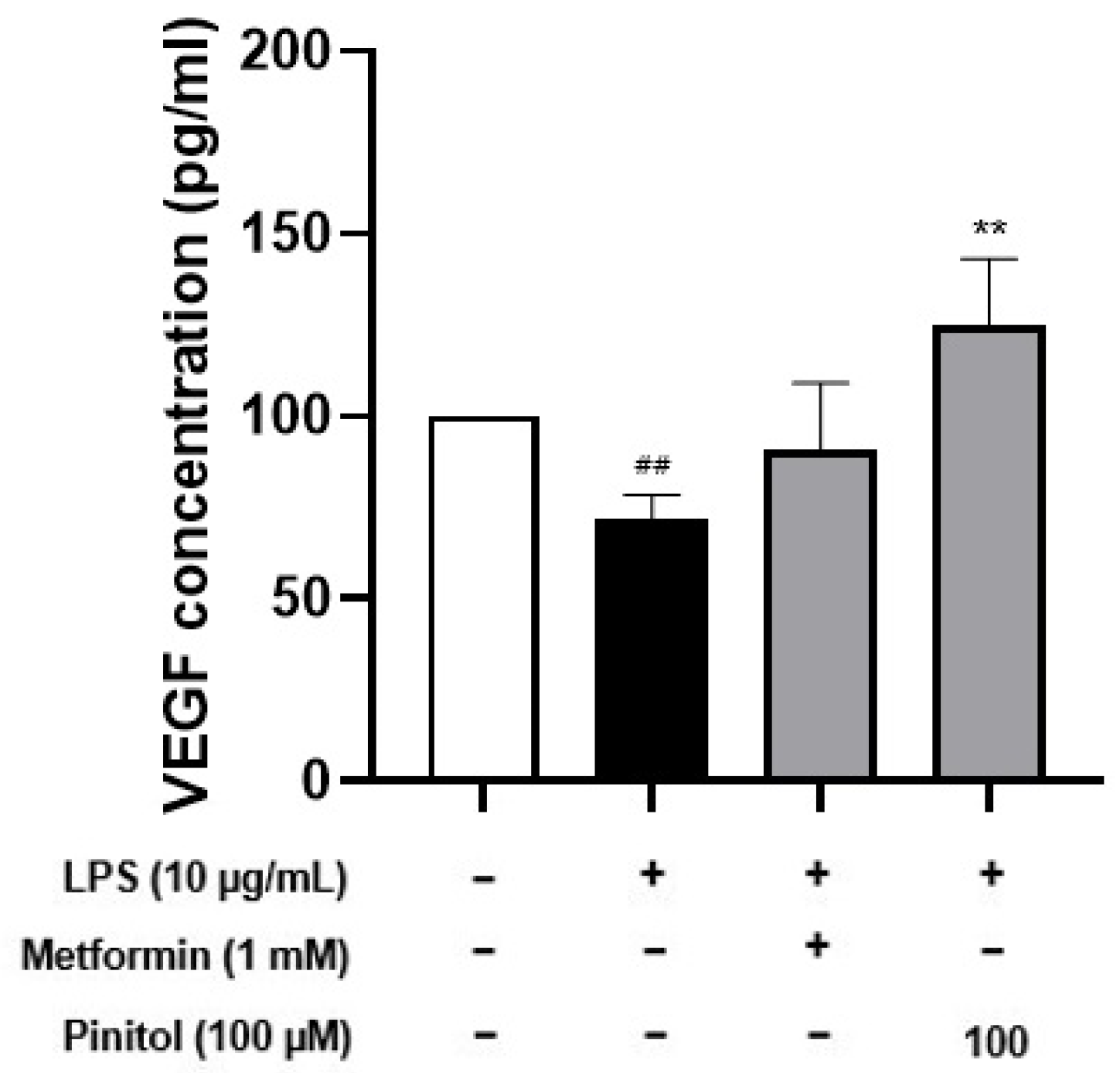
2.8. Pinitol Improved Specific Cytokine Levels in LPS-Damaged HDMECs
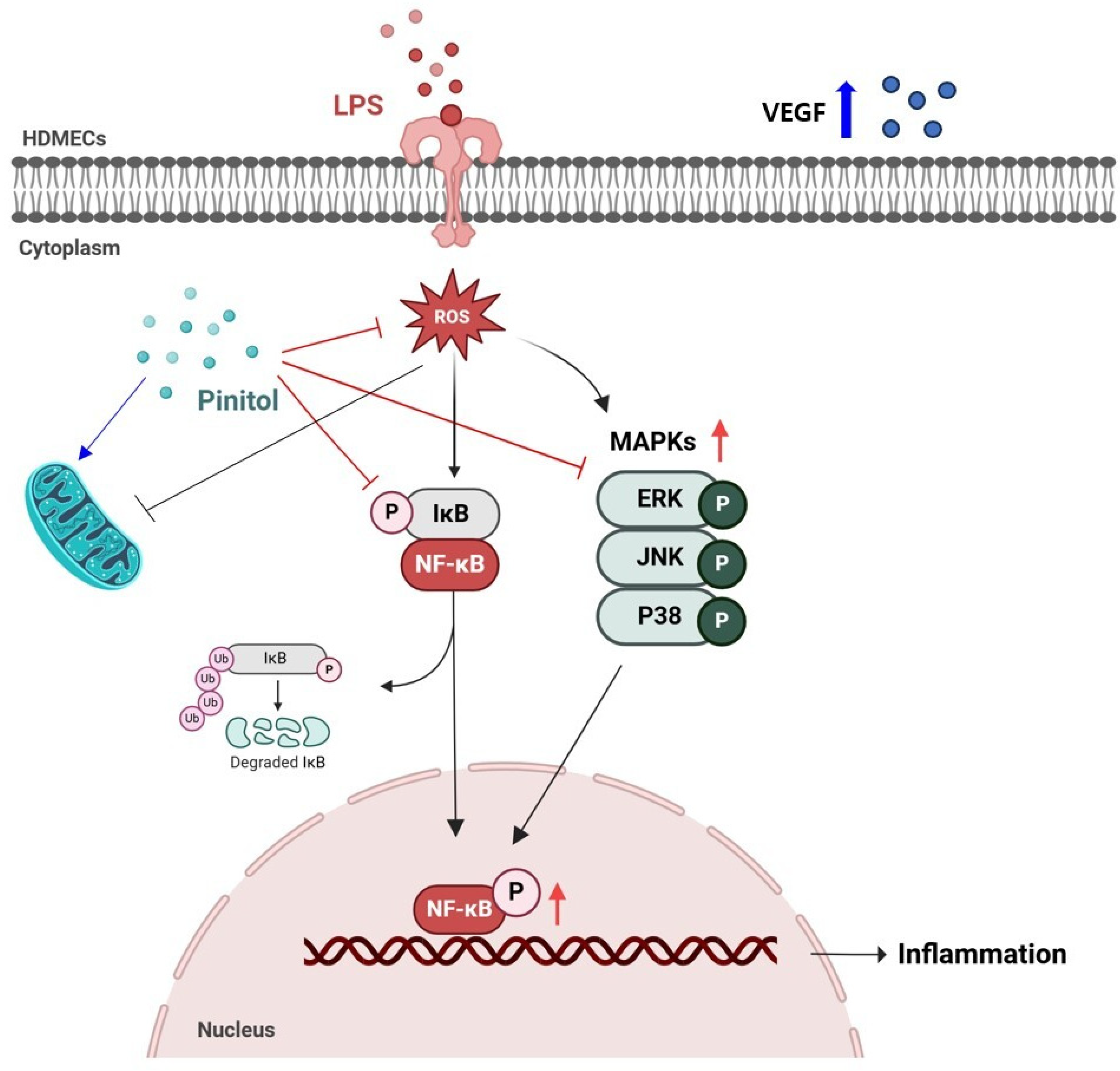
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Wound Healing Assay
4.5. Tube Formation Assay
4.6. DCF-DA ROS Assay
4.7. Mitochondrial Membrane Potential Assay
4.8. Immunofluorescence
4.9. Western Blot Analysis
4.10. ELISA Assay
4.11. Quantitative Real-Time Polymerase Chain Reaction
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prisby, R.D. Mechanical, Hormonal, and Metabolic Influences on Blood Vessels, Blood Flow and Bone. J. Endocrinol. 2017, 235, R77–R100. [Google Scholar] [CrossRef] [PubMed]
- Eelen, G.; Treps, L.; Li, X.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis Updated. Circ. Res. 2020, 127, 310–329. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Montmasson, M.; Garnier, C. Extracellular Matrix Contribution to Skin Wound Re-Epithelialization. Matrix Biol. 2019, 75–76, 12–26. [Google Scholar] [CrossRef]
- Lindley, L.E.; Stojadinovic, O.; Pastar, I.; Tomic-Canic, M. Biology and Biomarkers for Wound Healing. Plast. Reconstr. Surg. 2016, 138, 18S–28S. [Google Scholar] [CrossRef]
- Velnar, T.; Gradisnik, L. Tissue Augmentation in Wound Healing: The Role of Endothelial and Epithelial Cells. Med. Arch. 2018, 72, 444–448. [Google Scholar] [CrossRef]
- Todorova, D.; Simoncini, S.; Lacroix, R.; Sabatier, F.; Dignat-George, F. Extracellular Vesicles in Angiogenesis. Circ. Res. 2017, 120, 1658–1673. [Google Scholar] [CrossRef]
- Yuge, S.; Ishii, T.; Noishiki, C.; Fukuhara, S. Novel Regulatory Mechanisms Underlying Angiogenesis during Wound Healing Revealed by Fluorescence-Based Live-Imaging in Zebrafish. J. Biochem. 2023, 174, 5–12. [Google Scholar] [CrossRef]
- Nosrati, H.; Aramideh Khouy, R.; Nosrati, A.; Khodaei, M.; Banitalebi-Dehkordi, M.; Ashrafi-Dehkordi, K.; Sanami, S.; Alizadeh, Z. Nanocomposite Scaffolds for Accelerating Chronic Wound Healing by Enhancing Angiogenesis. J. Nanobiotechnol. 2021, 19, 1. [Google Scholar] [CrossRef]
- Vilas Boas, P.; Cerroni, L.; Requena, L. Intravascular Cutaneous Disorders. A Clinicopathologic Review. Am. J. Dermatopathol. 2021, 43, 119–136. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive Oxygen Species: Key Regulators in Vascular Health and Diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef]
- Cracowski, J.; Roustit, M. Human Skin Microcirculation. Compr. Physiol. 2020, 10, 1105–1154. [Google Scholar] [PubMed]
- Papi, M.; Papi, C. Vasculitic Ulcers. Int. J. Low. Extrem. Wounds 2016, 15, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Raharja, A.; Mahil, S.K.; Barker, J.N. Psoriasis: A Brief Overview. Clin. Med. 2021, 21, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Herrick, A.L.; Wigley, F.M. Raynaud’s Phenomenon. Best Pract. Res. Clin. Rheumatol. 2020, 34, 101474. [Google Scholar] [CrossRef]
- Calderon, L.M.; Pope, J.E. Scleroderma Epidemiology Update. Curr. Opin. Rheumatol. 2021, 33, 122–127. [Google Scholar] [CrossRef]
- Burgess, J.L.; Wyant, W.A.; Abdo Abujamra, B.; Kirsner, R.S.; Jozic, I. Diabetic Wound-Healing Science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Orecchia, A.; Scarponi, C.; Di Felice, F.; Cesarini, E.; Avitabile, S.; Mai, A.; Mauro, M.L.; Sirri, V.; Zambruno, G.; Albanesi, C.; et al. Sirtinol Treatment Reduces Inflammation in Human Dermal Microvascular Endothelial Cells. PLoS ONE 2011, 6, e24307. [Google Scholar] [CrossRef]
- Baselet, B.; Sonveaux, P.; Baatout, S.; Aerts, A. Pathological Effects of Ionizing Radiation: Endothelial Activation and Dysfunction. Cell Mol. Life Sci. 2019, 76, 699–728. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R.; Annese, T. The Role of Vascular Niche and Endothelial Cells in Organogenesis and Regeneration. Exp. Cell Res. 2021, 398, 112398. [Google Scholar] [CrossRef]
- Immanuel, J.; Yun, S. Vascular Inflammatory Diseases and Endothelial Phenotypes. Cells 2023, 12, 1640. [Google Scholar] [CrossRef] [PubMed]
- Lallo, V.; Bracaglia, L.G. Influencing Endothelial Cells’ Roles in Inflammation and Wound Healing Through Nucleic Acid Delivery. Tissue Eng. Part A 2024, 30, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J. Interleukin-33 (IL-33): A Critical Review of its Biology and the Mechanisms Involved in its Release as a Potent Extracellular Cytokine. Cytokine 2022, 156, 155891. [Google Scholar] [PubMed]
- Du, H.; Li, S.; Lu, J.; Tang, L.; Jiang, X.; He, X.; Liang, J.; Liao, X.; Cui, T.; Huang, Y.; et al. Single-Cell RNA-Seq and Bulk-Seq Identify RAB17 as a Potential Regulator of Angiogenesis by Human Dermal Microvascular Endothelial Cells in Diabetic Foot Ulcers. Burn. Trauma 2023, 11, tkad020. [Google Scholar] [CrossRef]
- Gonzalez-Mauraza, N.H.; Leon-Gonzalez, A.J.; Espartero, J.L.; Gallego-Fernandez, J.B.; Sanchez-Hidalgo, M.; Martin-Cordero, C. Isolation and Quantification of Pinitol, a Bioactive Cyclitol, in Retama spp. Nat. Prod. Commun. 2016, 11, 405–406. [Google Scholar] [CrossRef]
- Azab, A. D-Pinitol-Active Natural Product from Carob with Notable Insulin Regulation. Nutrients 2022, 14, 1453. [Google Scholar] [CrossRef]
- Pandi, A.; Sattu, K.; Kalappan, V.M.; Lal, V.; Varikasuvu, S.R.; Ganguly, A.; Prasad, J. Pharmacological Effects of D-Pinitol—A Comprehensive Review. J. Food Biochem. 2022, 46, e14282. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, M.; Wu, T.; Xu, M.; Cai, H.; Zhang, Z. Effects of D-Pinitol on Insulin Resistance through the PI3K/Akt Signaling Pathway in Type 2 Diabetes Mellitus Rats. J. Agric. Food Chem. 2015, 63, 6019–6026. [Google Scholar]
- Sivakumar, S.; Palsamy, P.; Subramanian, S.P. Impact of D-Pinitol on the Attenuation of Proinflammatory Cytokines, Hyperglycemia-Mediated Oxidative Stress and Protection of Kidney Tissue Ultrastructure in Streptozotocin-Induced Diabetic Rats. Chem.-Biol. Interact. 2010, 188, 237–245. [Google Scholar] [CrossRef]
- Haque, M.F.; El-Nashar, H.A.S.; Akbor, M.S.; Alfaifi, M.; Bappi, M.H.; Chowdhury, A.K.; Hossain, M.K.; El-Shazly, M.; Albayouk, T.; Saleh, N.; et al. Anti-Inflammatory Activity of D-Pinitol Possibly through Inhibiting COX-2 Enzyme: In Vivo and in Silico Studies. Front. Chem. 2024, 12, 1366844. [Google Scholar]
- Jung, J.; Shim, J.H.; Cho, S.H.; Bae, I.; Yang, S.H.; Kim, J.; Lim, H.W.; Shin, D.W. The Anti-Diabetic Pinitol Improves Damaged Fibroblasts. Biomol. Ther. 2024, 32, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Go, M.Y.; Jeon, C.Y.; Shin, J.U.; Kim, M.; Lim, H.W.; Shin, D.W. Pinitol Improves Diabetic Foot Ulcers in Streptozotocin-Induced Diabetes Rats Through Upregulation of Nrf2/HO-1 Signaling. Antioxidants 2024, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Hawthorne, B.; Simmons, J.K.; Stuart, B.; Tung, R.; Zamierowski, D.S.; Mellott, A.J. Enhancing Wound Healing Dressing Development through Interdisciplinary Collaboration. J. Biomed. Mater. Res. B. Appl. Biomater. 2021, 109, 1967–1985. [Google Scholar] [PubMed]
- DiPietro, L.A. Angiogenesis and Wound Repair: When enough is Enough. J. Leukoc. Biol. 2016, 100, 979–984. [Google Scholar]
- Wang, P.; Huang, B.; Horng, H.; Yeh, C.; Chen, Y. Wound Healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar]
- Crompton, R.; Williams, H.; Ansell, D.; Campbell, L.; Holden, K.; Cruickshank, S.; Hardman, M.J. Oestrogen Promotes Healing in a Bacterial LPS Model of Delayed Cutaneous Wound Repair. Lab. Investig. 2016, 96, 439–449. [Google Scholar] [CrossRef]
- Farhana, A.; Khan, Y.S. Biochemistry, Lipopolysaccharide. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2024. [Google Scholar]
- Chan, Y.H.; Harith, H.H.; Israf, D.A.; Tham, C.L. Differential Regulation of LPS-Mediated VE-Cadherin Disruption in Human Endothelial Cells and the Underlying Signaling Pathways: A Mini Review. Front. Cell Dev. Biol. 2020, 7, 280. [Google Scholar]
- Murphy, M.P.; Holmgren, A.; Larsson, N.; Halliwell, B.; Chang, C.J.; Kalyanaraman, B.; Rhee, S.G.; Thornalley, P.J.; Partridge, L.; Gems, D.; et al. Unraveling the Biological Roles of Reactive Oxygen Species. Cell Metab. 2011, 13, 361–366. [Google Scholar]
- Panieri, E.; Santoro, M.M. ROS Signaling and Redox Biology in Endothelial Cells. Cell. Mol. Life Sci. 2015, 72, 3281–3303. [Google Scholar]
- Tian, M.; Ma, Y.; Lin, W. Fluorescent Probes for the Visualization of Cell Viability. Acc. Chem. Res. 2019, 52, 2147–2157. [Google Scholar]
- Natarajan, V.; Chawla, R.; Mah, T.; Vivekanandan, R.; Tan, S.Y.; Sato, P.Y.; Mallilankaraman, K. Mitochondrial Dysfunction in Age-Related Metabolic Disorders. Proteomics 2020, 20, e1800404. [Google Scholar] [PubMed]
- Rastogi, S.; Haldar, C. Comparative Effect of Melatonin and Quercetin in Counteracting LPS Induced Oxidative Stress in Bone Marrow Mononuclear Cells and Spleen of Funambulus Pennanti. Food Chem. Toxicol. 2018, 120, 243–252. [Google Scholar] [PubMed]
- Fock, E.M.; Parnova, R.G. Protective Effect of Mitochondria-Targeted Antioxidants Against Inflammatory Response to Lipopolysaccharide Challenge: A Review. Pharmaceutics 2021, 13, 144. [Google Scholar] [CrossRef] [PubMed]
- Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.; Tzur, A. JC-1: Alternative Excitation Wavelengths Facilitate Mitochondrial Membrane Potential Cytometry. Cell Death Dis. 2012, 3, e430. [Google Scholar]
- Yang, R.; Gao, W.; Wang, Z.; Jian, H.; Peng, L.; Yu, X.; Xue, P.; Peng, W.; Li, K.; Zeng, P. Polyphyllin I Induced Ferroptosis to Suppress the Progression of Hepatocellular Carcinoma through Activation of the Mitochondrial Dysfunction Via Nrf2/HO-1/GPX4 Axis. Phytomedicine 2024, 122, 155135. [Google Scholar]
- Liu, S.; Chen, Q.; Liu, J.; Yang, X.; Zhang, Y.; Huang, F. Sinomenine Protects Against E.Coli-Induced Acute Lung Injury in Mice through Nrf2-NF-kappaB Pathway. Biomed. Pharmacother. 2018, 107, 696–702. [Google Scholar]
- Zhang, P.; Yang, M.; Chen, C.; Liu, L.; Wei, X.; Zeng, S. Toll-Like Receptor 4 (TLR4)/Opioid Receptor Pathway Crosstalk and Impact on Opioid Analgesia, Immune Function, and Gastrointestinal Motility. Front. Immunol. 2020, 11, 1455. [Google Scholar]
- Zhao, Z.; Sun, Y.; Ruan, X. Bornyl Acetate: A Promising Agent in Phytomedicine for Inflammation and Immune Modulation. Phytomedicine 2023, 114, 154781. [Google Scholar]
- Tang, J.; Xu, L.; Zeng, Y.; Gong, F. Effect of Gut Microbiota on LPS-Induced Acute Lung Injury by Regulating the TLR4/NF-kB Signaling Pathway. Int. Immunopharmacol. 2021, 91, 107272. [Google Scholar]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar]
- Yu, J.; Ming, H.; Li, H.Y.; Yu, B.; Chu, M.; Zhu, H.; Zhu, X. IMM-H007, a Novel Small Molecule Inhibitor for Atherosclerosis, Represses Endothelium Inflammation by Regulating the Activity of NF-kappaB and JNK/AP1 Signaling. Toxicol. Appl. Pharmacol. 2019, 381, 114732. [Google Scholar] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [PubMed]
- Wang, Z.; Fang, C.; Yao, M.; Wu, D.; Chen, M.; Guo, T.; Mo, J. Research Progress of NF-kappaB Signaling Pathway and Thrombosis. Front. Immunol. 2023, 14, 1257988. [Google Scholar]
- DiPietro, L.A. Angiogenesis and Scar Formation in Healing Wounds. Curr. Opin. Rheumatol. 2013, 25, 87–91. [Google Scholar]
- Arnaoutova, I.; Kleinman, H.K. In Vitro Angiogenesis: Endothelial Cell Tube Formation on Gelled Basement Membrane Extract. Nat. Protoc. 2010, 5, 628–635. [Google Scholar]
- Kelley, M.; Fierstein, S.; Purkey, L.; DeCicco-Skinner, K. Endothelial Cell Tube Formation Assay: An In Vitro Model for Angiogenesis. Methods Mol. Biol. 2022, 2475, 187–196. [Google Scholar]
- Kubota, Y.; Kleinman, H.K.; Martin, G.R.; Lawley, T.J. Role of Laminin and Basement Membrane in the Morphological Differentiation of Human Endothelial Cells into Capillary-Like Structures. J. Cell Biol. 1988, 107, 1589–1598. [Google Scholar]
- DeCicco-Skinner, K.L.; Henry, G.H.; Cataisson, C.; Tabib, T.; Gwilliam, J.C.; Watson, N.J.; Bullwinkle, E.M.; Falkenburg, L.; O’Neill, R.C.; Morin, A.; et al. Endothelial Cell Tube Formation Assay for the in Vitro Study of Angiogenesis. J. Vis. Exp. 2014, 91, e51312. [Google Scholar]
- Li, Y.; Zhu, H.; Wei, X.; Li, H.; Yu, Z.; Zhang, H.; Liu, W. LPS Induces HUVEC Angiogenesis in Vitro through miR-146a-Mediated TGF-Beta1 Inhibition. Am. J. Transl. Res. 2017, 9, 591–600. [Google Scholar]
- Kim, Y.; West, X.Z.; Byzova, T.V. Inflammation and Oxidative Stress in Angiogenesis and Vascular Disease. J. Mol. Med. 2013, 91, 323–328. [Google Scholar]
- Karl, E.; Zhang, Z.; Dong, Z.; Neiva, K.G.; Soengas, M.S.; Koch, A.E.; Polverini, P.J.; Núñez, G.; Nör, J.E. Unidirectional crosstalk between Bcl-xL and Bcl-2 enhances the angiogenic phenotype of endothelial cells. Cell Death Differ. 2007, 14, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Tao, R.; Chen, L.; Xiong, Y.; Xue, H.; Hu, L.; Yan, C.; Xie, X.; Lin, Z.; Panayi, A.C.; et al. Exosomes derived from pioglitazone-pretreated MSCs accelerate diabetic wound healing through enhancing angiogenesis. Nanobiotechnology 2021, 19, 150. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Goodison, S.; Urquidi, V.; Gomes Giacoia, E.; Rosser, C.J. Expression of CXCL1 in human endothelial cells induces angiogenesis through the CXCR2 receptor and the ERK1/2 and EGF pathways. Lab. Investig. 2013, 93, 768–778. [Google Scholar] [CrossRef]
- O’Leary, A.P.; Fox, J.M.; Pullar, C.E. Beta-Adrenoceptor Activation Reduces Both Dermal Microvascular Endothelial Cell Migration via a cAMP-Dependent Mechanism and Wound Angiogenesis. J. Cell. Physiol. 2015, 230, 356–365. [Google Scholar] [CrossRef]
- Li, Y.; Xu, T.; Tu, Z.; Dai, W.; Xue, Y.; Tang, C.; Gao, W.; Mao, C.; Lei, B.; Lin, C. Bioactive antibacterial silica-based nanocomposites hydrogel scaffolds with high angiogenesis for promoting diabetic wound healing and skin repair. Theranostics 2020, 10, 4929–4943. [Google Scholar] [CrossRef]
- Laiva, A.L.; O’Brien, F.J.; Keogh, M.B. Innovations in gene and growth factor delivery systems for diabetic wound healing. J. Tissue Eng. Regen. Med. 2018, 12, e296–e312. [Google Scholar] [CrossRef]
- Shi, Y.; Vanhoutte, P.M. Macro- and Microvascular Endothelial Dysfunction in Diabetes. J. Diabetes 2017, 9, 434–449. [Google Scholar] [CrossRef]
- Fung, T.H.; Patel, B.; Wilmot, E.G.; Amoaku, W.M. Diabetic Retinopathy for the Non-Ophthalmologist. Clin. Med. 2022, 22, 112–116. [Google Scholar] [CrossRef]
- Guo, W.; Song, Y.; Sun, Y.; Du, H.; Cai, Y.; You, Q.; Fu, H.; Shao, L. Systemic Immune-Inflammation Index is Associated with Diabetic Kidney Disease in Type 2 Diabetes Mellitus Patients: Evidence from NHANES 2011–2018. Front. Endocrinol. 2022, 13, 1071465. [Google Scholar] [CrossRef]
- Armstrong, D.G.; Tan, T.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers: A Review. JAMA 2023, 330, 62–75. [Google Scholar] [CrossRef]
- Laurent, S. Antihypertensive Drugs. Pharmacol. Res. 2017, 124, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.; Bhattacharjee, K.; Choudhury, Y. Pleiotropic Effects of Anti-Diabetic Drugs: A Comprehensive Review. Eur. J. Pharmacol. 2020, 884, 173349. [Google Scholar] [PubMed]
- Attardo, S.; Musumeci, O.; Velardo, D.; Toscano, A. Statins Neuromuscular Adverse Effects. Int. J. Mol. Sci. 2022, 23, 8364. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.L.; Shaw, N.M.; Abbasi, B.; Hakam, N.; Breyer, B.N. Adverse Reactions of PDE5 Inhibitors: An Analysis of the World Health Organization Pharmacovigilance Database. Andrology 2023, 11, 1408–1417. [Google Scholar]
- Dzidic-Krivic, A.; Sher, E.K.; Kusturica, J.; Farhat, E.K.; Nawaz, A.; Sher, F. Unveiling Drug Induced Nephrotoxicity using Novel Biomarkers and Cutting-Edge Preventive Strategies. Chem. Biol. Interact. 2024, 388, 110838. [Google Scholar]
- LaMoia, T.E.; Shulman, G.I. Cellular and Molecular Mechanisms of Metformin Action. Endocr. Rev. 2021, 42, 77–96. [Google Scholar]
- Triggle, C.R.; Marei, I.; Ye, K.; Ding, H.; Anderson, T.J.; Hollenberg, M.D.; Hill, M.A. Repurposing Metformin for Vascular Disease. Curr. Med. Chem. 2023, 30, 3955–3978. [Google Scholar] [CrossRef]
- Feng, J.; Wang, X.; Ye, X.; Ares, I.; Lopez-Torres, B.; Martinez, M.; Martinez-Larranaga, M.; Wang, X.; Anadon, A.; Martinez, M. Mitochondria as an Important Target of Metformin: The Mechanism of Action, Toxic and Side Effects, and New Therapeutic Applications. Pharmacol. Res. 2022, 177, 106114. [Google Scholar]
- Salvatore, T.; Pafundi, P.C.; Galiero, R.; Rinaldi, L.; Caturano, A.; Vetrano, E.; Aprea, C.; Albanese, G.; Di Martino, A.; Ricozzi, C.; et al. Can Metformin Exert as an Active Drug on Endothelial Dysfunction in Diabetic Subjects? Biomedicines 2020, 9, 3. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, Y.; Ling, P.; Feng, X.; Luo, S.; Zheng, X.; Little, P.J.; Xu, S.; Weng, J. Metformin in Cardiovascular Diabetology: A Focused Review of its Impact on Endothelial Function. Theranostics 2021, 11, 9376–9396. [Google Scholar] [CrossRef]
- Dutta, S.; Shah, R.B.; Singhal, S.; Dutta, S.B.; Bansal, S.; Sinha, S.; Haque, M. Metformin: A Review of Potential Mechanism and Therapeutic Utility Beyond Diabetes. Drug Des. Devel. Ther. 2023, 17, 1907–1932. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ji, J.Y. Progerin-Induced Impairment in Wound Healing and Proliferation in Vascular Endothelial Cells. Front. Aging 2022, 3, 844885. [Google Scholar]
- Meng, M.; Jiang, Y.; Wang, Y.; Huo, R.; Ma, N.; Shen, X.; Chang, G. Beta-Carotene Targets IP3R/GRP75/VDAC1-MCU Axis to Renovate LPS-Induced Mitochondrial Oxidative Damage by Regulating STIM1. Free Radic. Biol. Med. 2023, 205, 25–46. [Google Scholar] [PubMed]
- Zheng, D.; Liu, J.; Piao, H.; Zhu, Z.; Wei, R.; Liu, K. ROS-Triggered Endothelial Cell Death Mechanisms: Focus on Pyroptosis, Parthanatos, and Ferroptosis. Front. Immunol. 2022, 13, 1039241. [Google Scholar]
- Chen, H.; Chong, I.; Lee, Y.; Tsai, J.; Yuan, S.F.; Wang, H.; Liu, W.; Liu, P. Kruppel-Like Factor 5 Mediates Proinflammatory Cytokine Expression in Lipopolysaccharide-Induced Acute Lung Injury through Upregulation of Nuclear Factor-kappaB Phosphorylation in Vitro and in Vivo. Mediat. Inflamm. 2014, 2014, 281984. [Google Scholar]
- Hu, T.; Ju, J.; Mo, L.; Ma, L.; Hu, W.; You, R.; Chen, X.; Chen, Y.; Liu, Z.; Qiu, S.; et al. Anti-Inflammation Action of Xanthones from Swertia Chirayita by Regulating COX-2/NF-kappaB/MAPKs/Akt Signaling Pathways in RAW 264.7 Macrophage Cells. Phytomedicine 2019, 55, 214–221. [Google Scholar]
- Medina-Vera, D.; Navarro, J.A.; Tovar, R.; Rosell-Valle, C.; Gutiérrez-Adan, A.; Ledesma, J.C.; Sanjuan, C.; Pavón, F.J.; Baixeras, E.; Rodríguez de Fonseca, F.; et al. Activation of PI3K/Akt Signaling Pathway in Rat Hypothalamus Induced by an Acute Oral Administration of D-Pinitol. Nutrients 2021, 13, 2268. [Google Scholar] [CrossRef]
- Li, X.L.; Xu, M.; Yu, F.; Fu, C.L.; Yu, X.; Cheng, M.; Gao, H.Q. Effects of D-pinitol on myocardial apoptosis and fibrosis in streptozocin-induced aging-accelerated mice. J. Food Biochem. 2021, 45, e13669. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, M.Y.; Kim, J.; Jeon, C.Y.; Kim, M.; Shin, D.W. Pinitol Improves Lipopolysaccharide-Induced Cellular Damage in Human Dermal Microvascular Endothelial Cells. Molecules 2025, 30, 1513. https://doi.org/10.3390/molecules30071513
Go MY, Kim J, Jeon CY, Kim M, Shin DW. Pinitol Improves Lipopolysaccharide-Induced Cellular Damage in Human Dermal Microvascular Endothelial Cells. Molecules. 2025; 30(7):1513. https://doi.org/10.3390/molecules30071513
Chicago/Turabian StyleGo, Min Young, Jinsick Kim, Chae Young Jeon, Mujun Kim, and Dong Wook Shin. 2025. "Pinitol Improves Lipopolysaccharide-Induced Cellular Damage in Human Dermal Microvascular Endothelial Cells" Molecules 30, no. 7: 1513. https://doi.org/10.3390/molecules30071513
APA StyleGo, M. Y., Kim, J., Jeon, C. Y., Kim, M., & Shin, D. W. (2025). Pinitol Improves Lipopolysaccharide-Induced Cellular Damage in Human Dermal Microvascular Endothelial Cells. Molecules, 30(7), 1513. https://doi.org/10.3390/molecules30071513