
Characterization and In Vitro Prebiotic Activity of Pterostilbene/β-Cyclodextrin Inclusion Complexes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Phase Solubility Method
2.2. Characterization of PTS/β-CD Inclusion Complexes
2.2.1. SEM Analysis
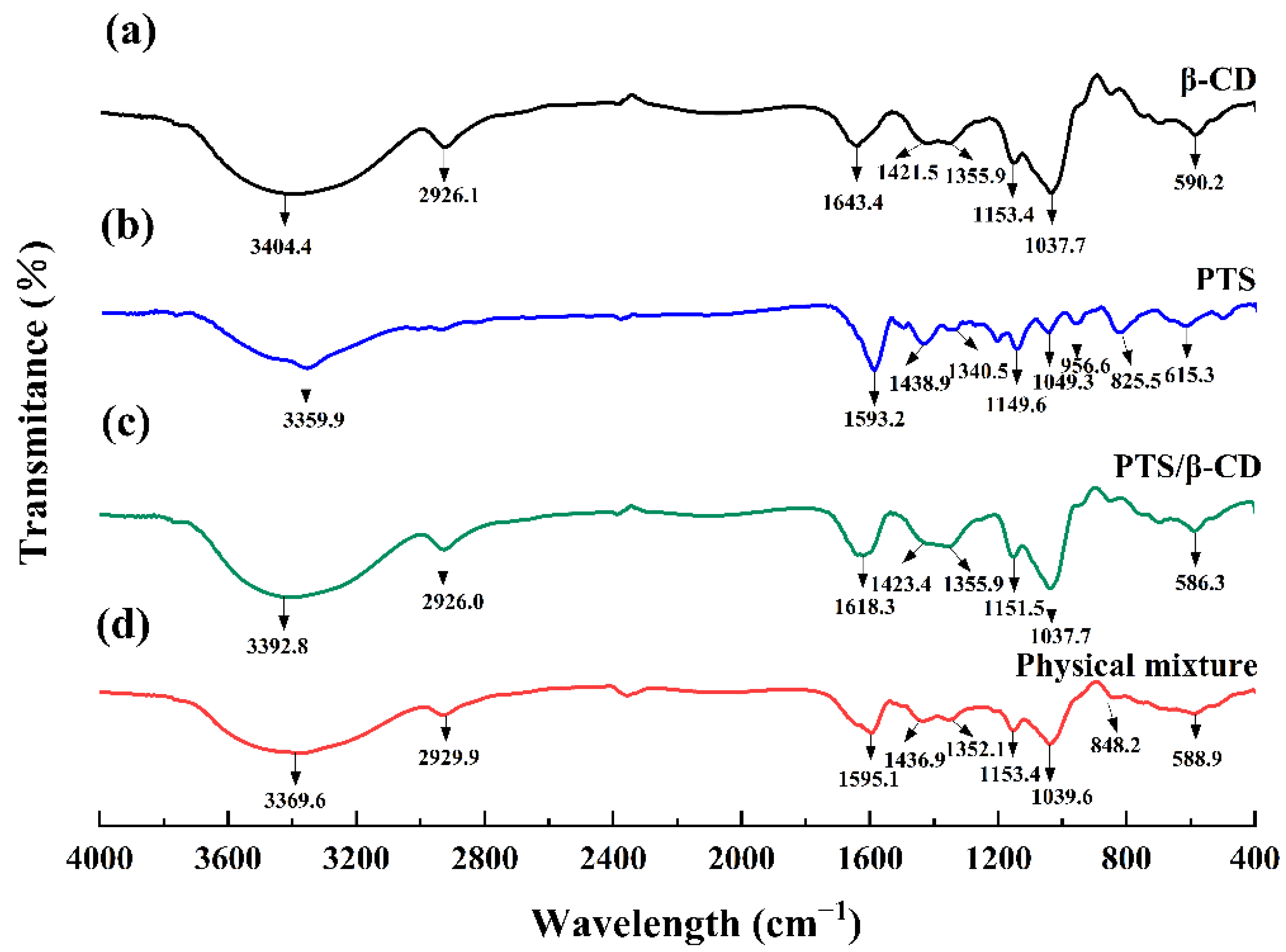
2.2.2. FT-IR Analysis
2.2.3. DSC Analysis
2.2.4. TG Analysis
2.2.5. XRD Analysis
2.2.6. NMR Analysis
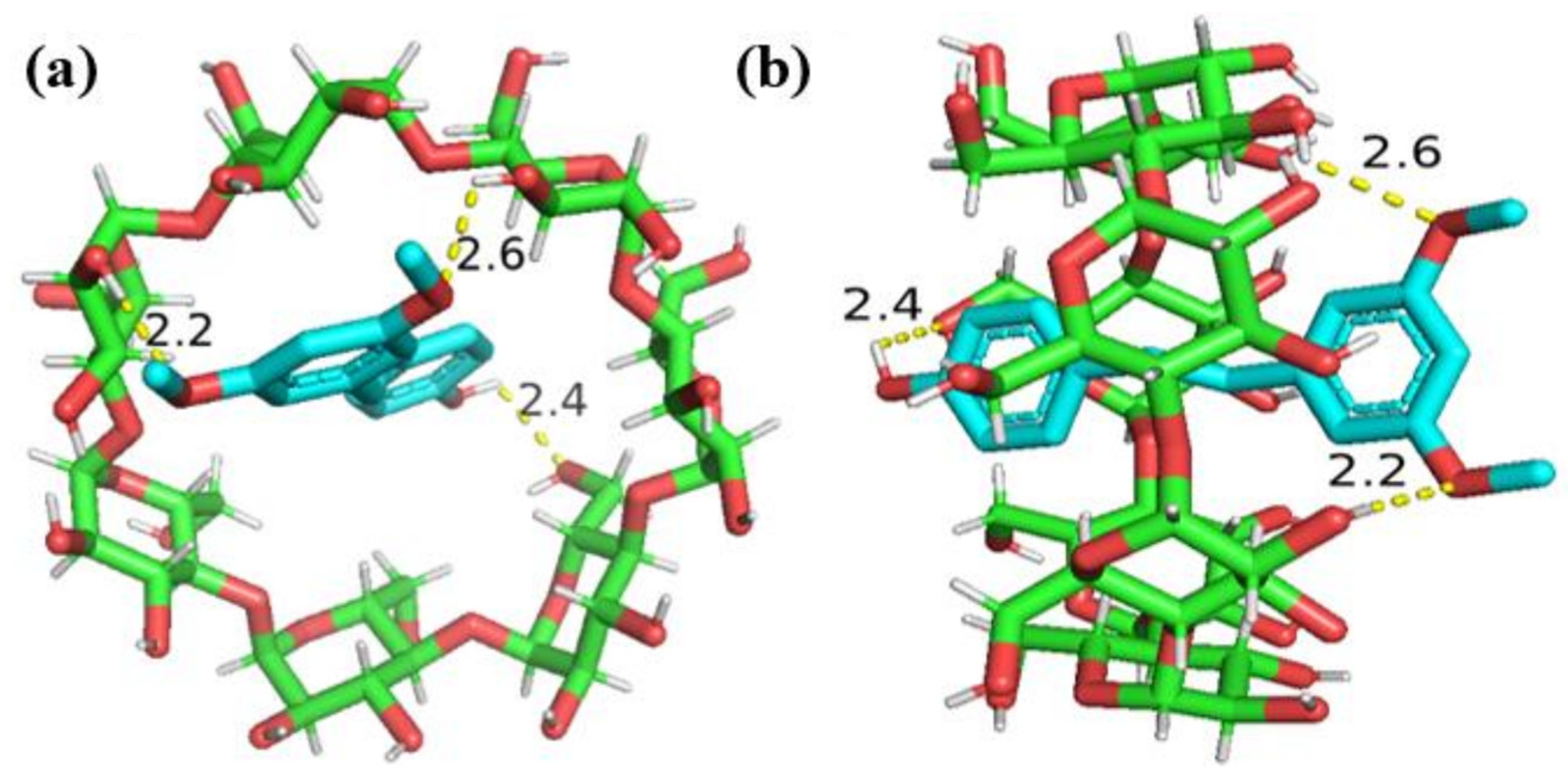
2.3. Molecular Docking Analysis
2.4. Antioxidant Analysis of PTS/β-CD Inclusion Complexes Concentrations
2.5. Stability of PTS/β-CD Inclusion Complexes
2.6. Simulated Digestion of PTS/β-CD Inclusion Complexes In Vitro
2.7. The Antioxidant Properties of PTS/β-CD Inclusion Complexes During Digestion in Vitro
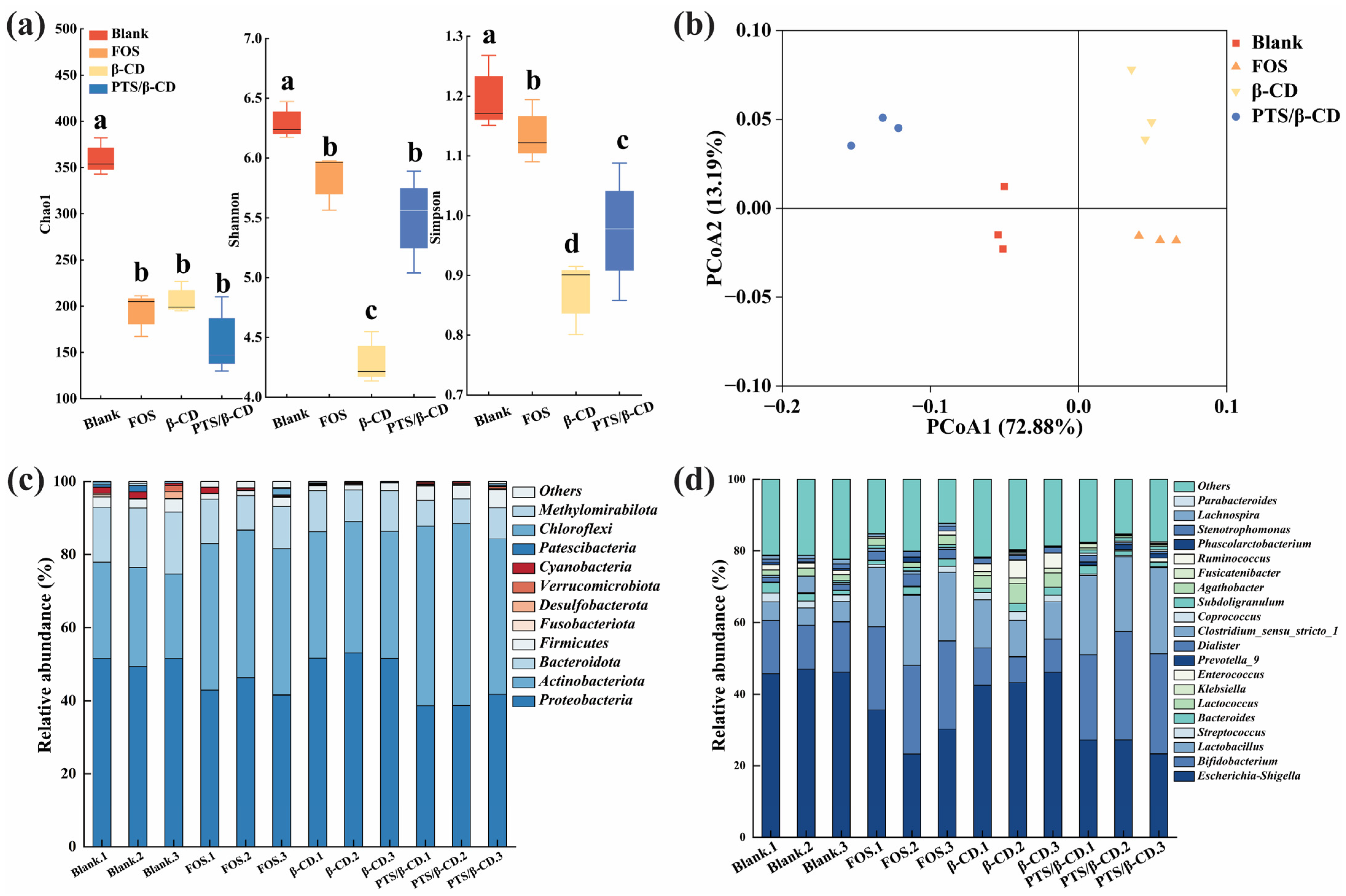
2.8. Influences of PTS/β-CD Inclusion Complexes on Intestinal Flora
2.8.1. Influences of PTS/β-CD Inclusion Complexes on pH and Biomass of Intestinal Flora Fermentation
2.8.2. Influences of PTS/β-CD Inclusion Complexes on SCFAs of Intestinal Flora Fermentation
2.8.3. Effects of PTS/β-CD Inclusion Complexes on Intestinal Flora
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Experimental Method
3.2.1. Phase Solubility Curve
3.2.2. Synthesis of PTS/β-CD Inclusion Complexes
3.2.3. Preparation of PTS/β-CD Physical Mixtures
3.2.4. Analysis of PTS/β-CD Inclusion Complexes
3.2.5. Molecular Docking
3.2.6. Different Concentration of PTS/β-CD Inclusion Complexes Antioxidant Activity
3.2.7. Stability Experiment of PTS/β-CD Inclusion Complexes
3.2.8. Simulated Digestion of PTS/β-CD Inclusion Complexes in Saliva, Gastric Juice, and Small Intestine In Vitro
3.2.9. Fecal Flora Fermentation in Healthy Human In Vitro
3.2.10. Measurement of SCFAs and Lactic Acid Concentrations
3.2.11. Detection of Fecal Flora Composition
3.2.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pralhad, T.; Rajendrakumar, K. Study of freeze-dried quercetin-cyclodextrin binary systems by DSC, FT-IR, X-ray diffraction and SEM analysis. J. Pharm. Biomed. Anal. 2004, 34, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Jia, F.; Han, Y.; Meng, X.; Xiao, Y.; Bai, S. Development and characterization of zein edible films incorporated with catechin/β-cyclodextrin inclusion complex nanoparticles. Carbohydr. Polym. 2021, 261, 117877–117884. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, S.; Costa Duarte, F.; Heimfarth, L.; Siqueira Quintans, J.; Quintans-Júnior, L.; Veiga Júnior, V.; Neves De Lima, Á. Cyclodextrin–Drug Inclusion Complexes: In Vivo and In Vitro Approaches. Int. J. Mol. Sci. 2019, 20, 642. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.W.; Yuan, X.F.; Li, H.; Zhu, Y.; Zhao, B. Preparation, characterization, and physicochemical property of the inclusion complexes of Cannabisin A with β-cyclodextrin and hydroxypropyl-β-cyclodextrin. J. Mol. Struct. 2023, 1272, 134168–134178. [Google Scholar] [CrossRef]
- Kumar, P.; Bhardwaj, V.K.; Purohit, R. Highly robust quantum mechanics and umbrella sampling studies on inclusion complexes of curcumin and β-cyclodextrin. Carbohydr. Polym. 2024, 323, 121432–121445. [Google Scholar] [CrossRef]
- Dutra, F.V.A.; Francisco, C.S.; Carneiro Pires, B.; Borges, M.M.C.; Torres, A.L.H.; Resende, V.A.; Mateus, M.F.M.; Cipriano, D.F.; Miguez, F.B.; Freitas, J.C.C.D.; et al. Coumarin/β-Cyclodextrin Inclusion Complexes Promote Acceleration and Improvement of Wound Healing. ACS Appl. Mater. Interfaces 2024, 16, 30900–30914. [Google Scholar] [CrossRef]
- Liang, X.; Chen, L.; Zhang, Z.; Zhang, R.; Mcclements, D.J.; Xu, Z.; Meng, M.; Jin, Z. Pterostilbene binding to whey protein: Multi-spectroscopy and the antioxidant activity. Food Hydrocoll. 2024, 150, 109681–109693. [Google Scholar] [CrossRef]
- Lin, W.; Leland, J.; Ho, C.; Pan, M. Occurrence, Bioavailability, Anti-inflammatory, and Anticancer Effects of Pterostilbene. J. Agric. Food Chem. 2020, 68, 12788–12799. [Google Scholar] [CrossRef]
- Koh, Y.C.; Lee, P.S.; Kuo, Y.L.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Dietary Pterostilbene and Resveratrol Modulate the Gut Microbiota Influenced by Circadian Rhythm Dysregulation. Mol. Nutr. Food Res. 2021, 65, 2100434. [Google Scholar] [CrossRef]
- Shi, Y.; Zhou, S.; Fan, S.; Ma, Y.; Han, Y. Encapsulation of bioactive polyphenols by starch and their impacts on gut microbiota. Curr. Opin. Food Sci. 2021, 38, 102–111. [Google Scholar] [CrossRef]
- Kaade, W.; Rubio, S.; Bogas-Droy, A.; De Villedon, V.; Laurichesse, E.; Schmitt, V. Highly Concentrated Emulsions Containing High Loads of Pterostilbene. ACS Omega 2023, 8, 25951–25959. [Google Scholar] [CrossRef] [PubMed]
- Lui, A.C.W.; Pow, K.C.; Lin, N.; Lam, L.P.Y.; Liu, G.; Godwin, I.D.; Fan, Z.; Khoo, C.J.; Tobimatsu, Y.; Wang, L.; et al. Regioselective stilbene O-methylations in Saccharinae grasses. Nat. Commun. 2023, 14, 3462. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.T.; Hu, G.A.; Zhu, L.; Zhao, Z.C.; Zhan, X.B. In vitro digestion and fecal fermentation of highly resistant starch rice and its effect on the gut microbiota. Food Chem. 2021, 361, 130095–130103. [Google Scholar] [CrossRef] [PubMed]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Li, H.; Gao, Z.; Xu, J.; Sun, W.; Wu, J.; Zhu, L.; Gao, M.; Zhan, X. Encapsulation of polyphenols in pH-responsive micelles self-assembled from octenyl-succinylated curdlan oligosaccharide and its effect on the gut microbiota. Colloids Surf. B. 2022, 219, 112857–112864. [Google Scholar] [CrossRef]
- Higuchi, T.A.; Connors, K.A. Phase-Solubility Techniques. In Advances in Analytical Chemistry and Instrumentation; Wiley-Interscience: New York, NY, USA, 1965. [Google Scholar]
- Catenacci, L.; Vicatos, A.I.; Milena, S.; Bonferoni, M.C.; Caira, M.R. Native Cyclodextrins as Complexation Agents for Pterostilbene: Complex Preparation and Characterization in Solution and in the Solid State. Pharmaceutics 2022, 14, 8. [Google Scholar] [CrossRef]
- Pu, H.; Sun, Q.; Tang, P.; Zhao, L.; Li, Q.; Liu, Y.; Li, H. Characterization and antioxidant activity of the complexes of tertiary butylhydroquinone with β-cyclodextrin and its derivatives. Food Chem. 2018, 260, 183–192. [Google Scholar] [CrossRef]
- Tom, L.; Nirmal, C.R.; Dusthackeer, A.; Magizhaveni, B.; Kurup, M.R.P. Formulation and evaluation of β-cyclodextrin-mediated inclusion complexes of isoniazid scaffolds: Molecular docking and in vitro assessment of antitubercular properties. New J. Chem. 2020, 44, 4467–4477. [Google Scholar] [CrossRef]
- Deng, S.; Liu, H.; Qi, C.; Yang, A.; Li, Z. Study on preparation and inclusion behavior of inclusion complexes between β-cyclodextrin derivatives with benzophenone. J. Incl. Phenom. Macrocycl. Chem. 2018, 90, 321–329. [Google Scholar] [CrossRef]
- Olga, G.; Styliani, C.; Ioannis, R.G. Coencapsulation of Ferulic and Gallic acid in hp-b-cyclodextrin. Food Chem. 2015, 185, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wei, B.; Li, H.; Wu, C.; Bai, Y.; Xu, X.; Jin, Z.; Tian, Y. Preparation of the β-cyclodextrin-vitamin C (β-CD-Vc) inclusion complex under high hydrostatic pressure (HHP). Carbohydr. Polym. 2012, 90, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Bin Jardan, Y.A.; Ahad, A.; Raish, M.; Al-Mohizea, A.M.; Al-Jenoobi, F.I. Microwave-Assisted Formation of Ternary Inclusion Complex of Pterostilbene. Pharmaceuticals 2023, 16, 1641. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.D.; Ni, Y.; Li, D. Preparation, characterization and evaluation of an inclusion complex of steviolbioside with γ-cyclodextrin. Food Biosci. 2018, 26, 65–72. [Google Scholar] [CrossRef]
- Yang, D.; Liu, H.; Meng, H.; Li, S. Preparation and physicochemical characterization of inclusion complex of pterostilbene-hydroxypropyl-β-cyclodextrin for enhancing the application of pterostilbene in cosmetics. J. Photochem. Photobiol. A. 2024, 447, 115221–115229. [Google Scholar] [CrossRef]
- Dai, K.; Wu, J.; Liu, X.; Wang, S.; Liu, Y.; Li, H.; Wang, H. Inclusion complex of quercetin with sulfobutylether β-cyclodextrin: Preparation, characterization, antioxidant and antibacterial activities and the inclusion mechanism. RSC Adv. 2024, 14, 9472–9481. [Google Scholar] [CrossRef]
- Zhang, S.; Zhou, Y.; Jin, S.; Meng, X.; Yang, L.; Wang, H. Preparation and structural characterization of corn starch–aroma compound inclusion complexes. J. Sci. Food Agric. 2016, 97, 182–190. [Google Scholar] [CrossRef]
- Michalska, P.; Wojnicz, A.; Ruiz-Nuo, A.; Abril, S.; León, R. Inclusion complex of ITH12674 with 2-hydroxypropyl-β-cyclodextrin: Preparation, physical characterization and pharmacological effect. Carbohydr. Polym. 2017, 157, 94–104. [Google Scholar] [CrossRef]
- Deng, C.; Cao, C.; Zhang, Y.; Hu, J.; Gong, Y.; Zheng, M.; Zhou, Y. Formation and stabilization mechanism of β-cyclodextrin inclusion complex with C10 aroma molecules. Food Hydrocoll. 2022, 123, 107013–107024. [Google Scholar] [CrossRef]
- Fuendjiep, V.; Wandji, J.; Tillequin, F.; Mulholland, D.A.; Budzikiewicz, H.; Fomum, Z.T.; Nyemba, A.M.; Koch, M. Chalconoid and stilbenoid glycosides from Guibourtia tessmanii. Phytochemistry 2002, 60, 803–806. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, J.Q.; Jiang, K.M.; Li, K.; Cong, Y.W.; Pu, S.P.; Jin, Y.; Lin, J. Preparation, characterisation and antitumour activity of β-, γ- and HP-β-cyclodextrin inclusion complexes of oxaliplatin. Spectrochim. Acta Part A 2016, 152, 501–508. [Google Scholar] [CrossRef]
- Jiang, Y.; Fang, Z.; Leonard, W.; Zhang, P. Phenolic compounds in Lycium berry: Composition, health benefits and industrial applications. J. Funct. Foods 2020, 77, 104340. [Google Scholar] [CrossRef]
- Strazisar, M.; Andrensek, S.; Smidovnik, A. Effect of β-cyclodextrin on antioxidant activity of coumaric acids. Food Chem. 2008, 110, 636–642. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Xu, N.; Yang, X.; Zhang, G.; Zhang, Y.; Liu, Q. Study of the protein, antioxidant activity, and starch duringin vitrosimulated digestion of green wheat and wheat cooked flours. Int. J. Food Prop. 2020, 23, 722–735. [Google Scholar] [CrossRef]
- Seraglio, S.K.T.; Valese, A.C.; Daguer, H.; Bergamo, G.; Azevedo, M.S.; Nehring, P.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Effect of in vitro gastrointestinal digestion on the bioaccessibility of phenolic compounds, minerals, and antioxidant capacity of Mimosa scabrella Bentham honeydew honeys. Food Res. Int. 2017, 99, 670–678. [Google Scholar] [CrossRef]
- Wu, C.; Gao, Z.; Liang, R.; Zhu, L.; Zhang, H.; Gao, M.; Zhan, X. Characterization and in vitro prebiotic activity analysis of succinoglycan produced by Rhizobium radiobacter. Process Biochem. 2023, 129, 293–305. [Google Scholar] [CrossRef]
- Hagihara, M.; Kuroki, Y.; Ariyoshi, T.; Higashi, S.; Fukuda, K.; Yamashita, R.; Matsymoto, A.; Mori, T.; Mimura, K.; Yamaguchi, N.; et al. Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier. iScience 2019, 23, 100772. [Google Scholar] [CrossRef]
- Sun, W.; Xu, J.; Yin, Z.; Li, H.; Li, J.; Zhu, L.; Li, Z.; Zhan, X. Fractionation, preliminary structural characterization and prebiotic activity of polysaccharide from the thin stillage of distilled alcoholic beverage. Process Biochem. 2022, 118, 52–64. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects from Hypertensive Cardiovascular. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Gao, W.; Fang, Z.; Lei, L.; Ju, L.; Jin, B.; Loor, J.J.; Liang, Y.; Shi, Z.; Shen, T.; Yu, H. Propionate alleviates palmitic acid-induced endoplasmic reticulum stress by enhancing autophagy in calf hepatic cells. J. Dairy Sci. 2021, 104, 9316–9326. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Q.; Goff, H.D.; Lapointe, G. Oligosaccharides: Structure, Function and Application. Encycl. Food Chem. 2019, 1, 202–207. [Google Scholar] [CrossRef]
- Brooks, A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, T.; Zhang, Y.; Chen, Y.; Ge, X.; Sui, W.; Zhu, Q.; Geng, J.; Zhang, M. Release of bound polyphenols from wheat bran soluble dietary fiber during simulated gastrointestinal digestion and colonic fermentation in vitro. Food Chem. 2023, 402, 134111–134119. [Google Scholar] [CrossRef]
- Shabbir, U.; Rubab, M.; Banan-Mwine, E.; Daliri, D.H. Curcumin, Quercetin, Catechins and Metabolic Diseases: The Role of Gut Microbiota. Nutrients 2021, 13, 206. [Google Scholar] [CrossRef]
- Wu, C.C.; Zhang, H.T.; Gao, Z.X.; Qu, J.J.; Zhu, L.; Zhan, X.B. Enhanced solubility of curcumin by complexation with fermented cyclic β-1, 2-glucans. J. Pharm. Biomed. Anal. 2022, 211, 114613–114620. [Google Scholar] [CrossRef]
- Catenacci, L.; Vicatos, A.I.; Sorrenti, M.; Edmonds-Smith, C.; Bonferoni, M.C.; Caira, M.R. Complexation between the Antioxidant Pterostilbene and Derivatized Cyclodextrins in the Solid State and in Aqueous Solution. Pharmaceuticals 2023, 16, 247. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Liu, H.; Ma, Z.; Guo, P.; Chen, H.; Gao, D. Extraction, structure analysis and antioxidant activity of Sibiraea laevigata (L.) Maxim polysaccharide. Int. J. Food Prop. 2022, 25, 2267–2285. [Google Scholar] [CrossRef]
- Wu, C.C.; Qu, J.J.; Zhang, H.T.; Gao, M.J.; Zhu, L.; Zhan, X.B. New two-stage pH combined with dissolved oxygen control strategy for cyclic β-1,2 glucans synthesis. Appl. Microbiol. Biotechnol. 2023, 107, 2235–2247. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-C.; Qian, L.; Rong, Z.; Li, Y.-Q.; Zhang, H.-M.; He, R.-Y.; Zhang, G.-Q. Characterization and In Vitro Prebiotic Activity of Pterostilbene/β-Cyclodextrin Inclusion Complexes. Molecules 2025, 30, 1363. https://doi.org/10.3390/molecules30061363
Wu C-C, Qian L, Rong Z, Li Y-Q, Zhang H-M, He R-Y, Zhang G-Q. Characterization and In Vitro Prebiotic Activity of Pterostilbene/β-Cyclodextrin Inclusion Complexes. Molecules. 2025; 30(6):1363. https://doi.org/10.3390/molecules30061363
Chicago/Turabian StyleWu, Chuan-Chao, Long Qian, Zhen Rong, Yu-Qi Li, Hui-Min Zhang, Rui-Yu He, and Guo-Qiang Zhang. 2025. "Characterization and In Vitro Prebiotic Activity of Pterostilbene/β-Cyclodextrin Inclusion Complexes" Molecules 30, no. 6: 1363. https://doi.org/10.3390/molecules30061363
APA StyleWu, C.-C., Qian, L., Rong, Z., Li, Y.-Q., Zhang, H.-M., He, R.-Y., & Zhang, G.-Q. (2025). Characterization and In Vitro Prebiotic Activity of Pterostilbene/β-Cyclodextrin Inclusion Complexes. Molecules, 30(6), 1363. https://doi.org/10.3390/molecules30061363