

Main Chemical Components, Activity and Mechanism of Repellence of Cyperus esculentus Essential Oil Against Tribolium confusum

 , ,
, ,
Abstract
1. Introduction
2. Results
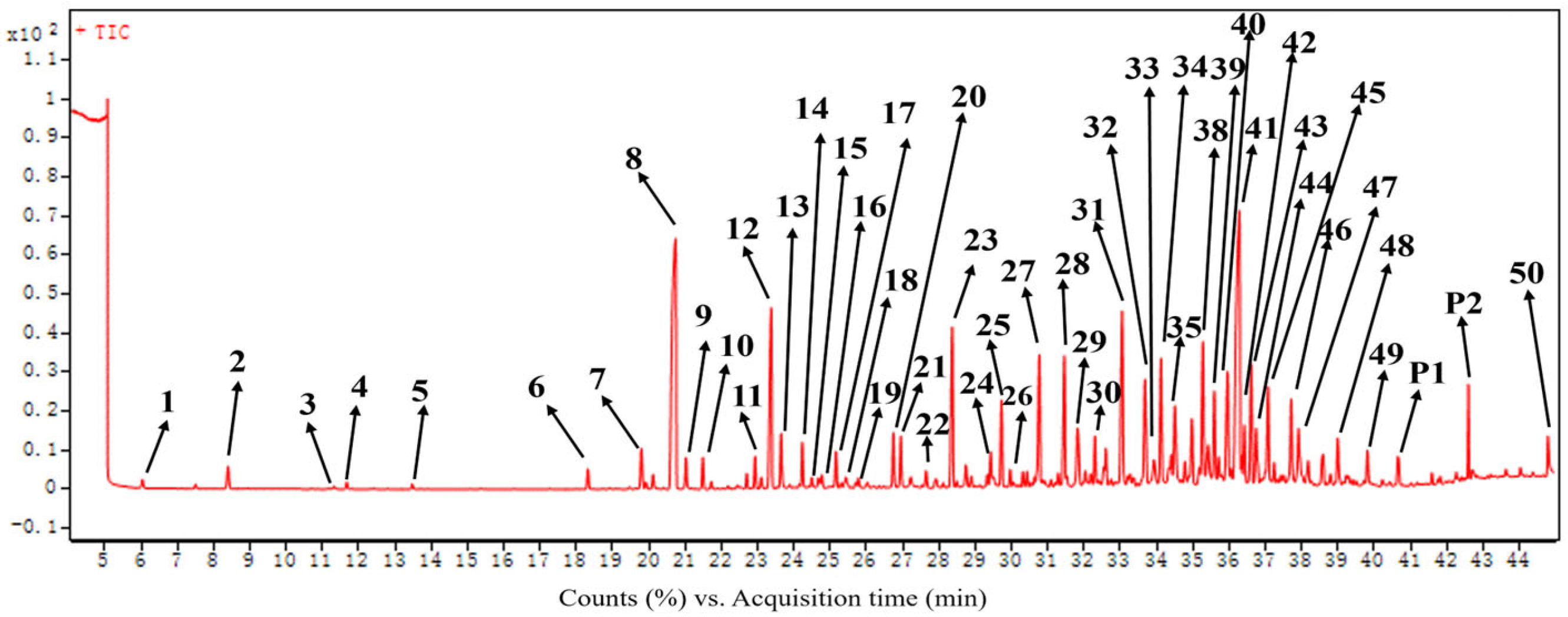
2.1. Components of the Essential Oil of C. esculentus Roots
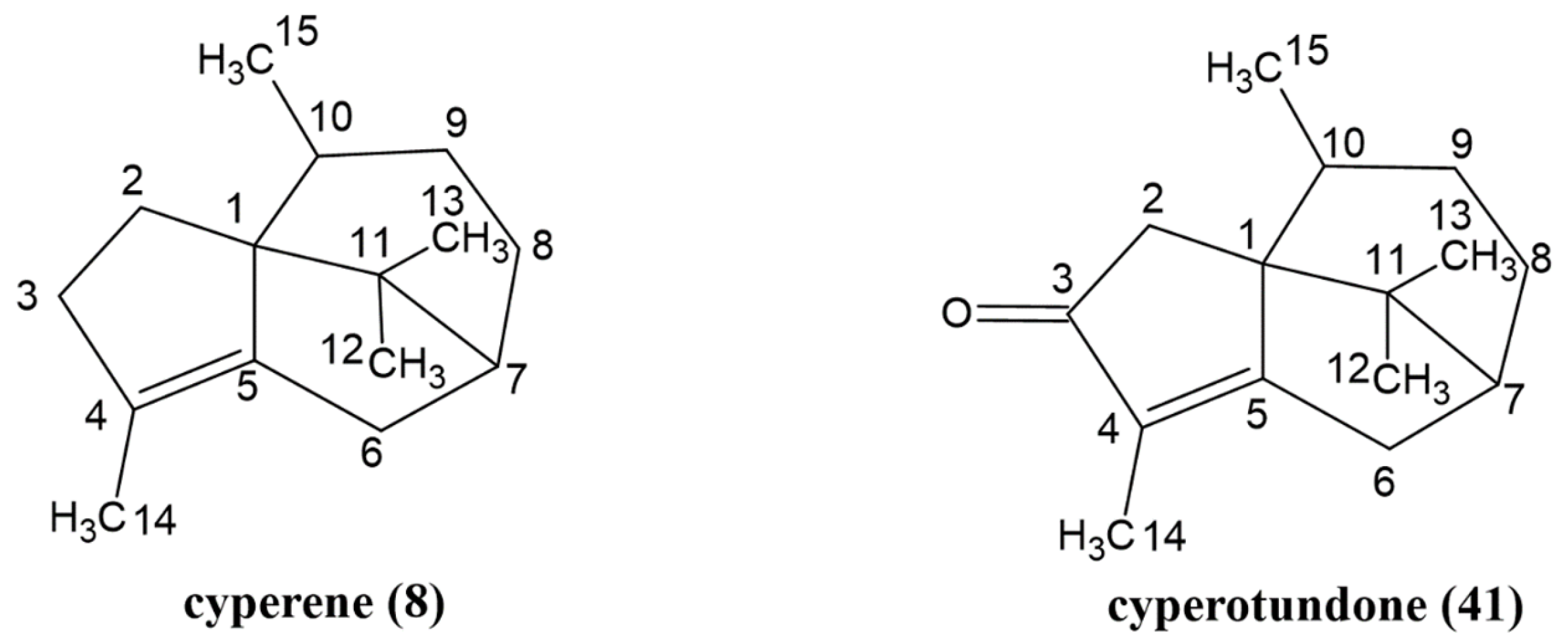
2.2. Isolation and Structural Confirmation of the Two Main Compounds in the Essential Oil
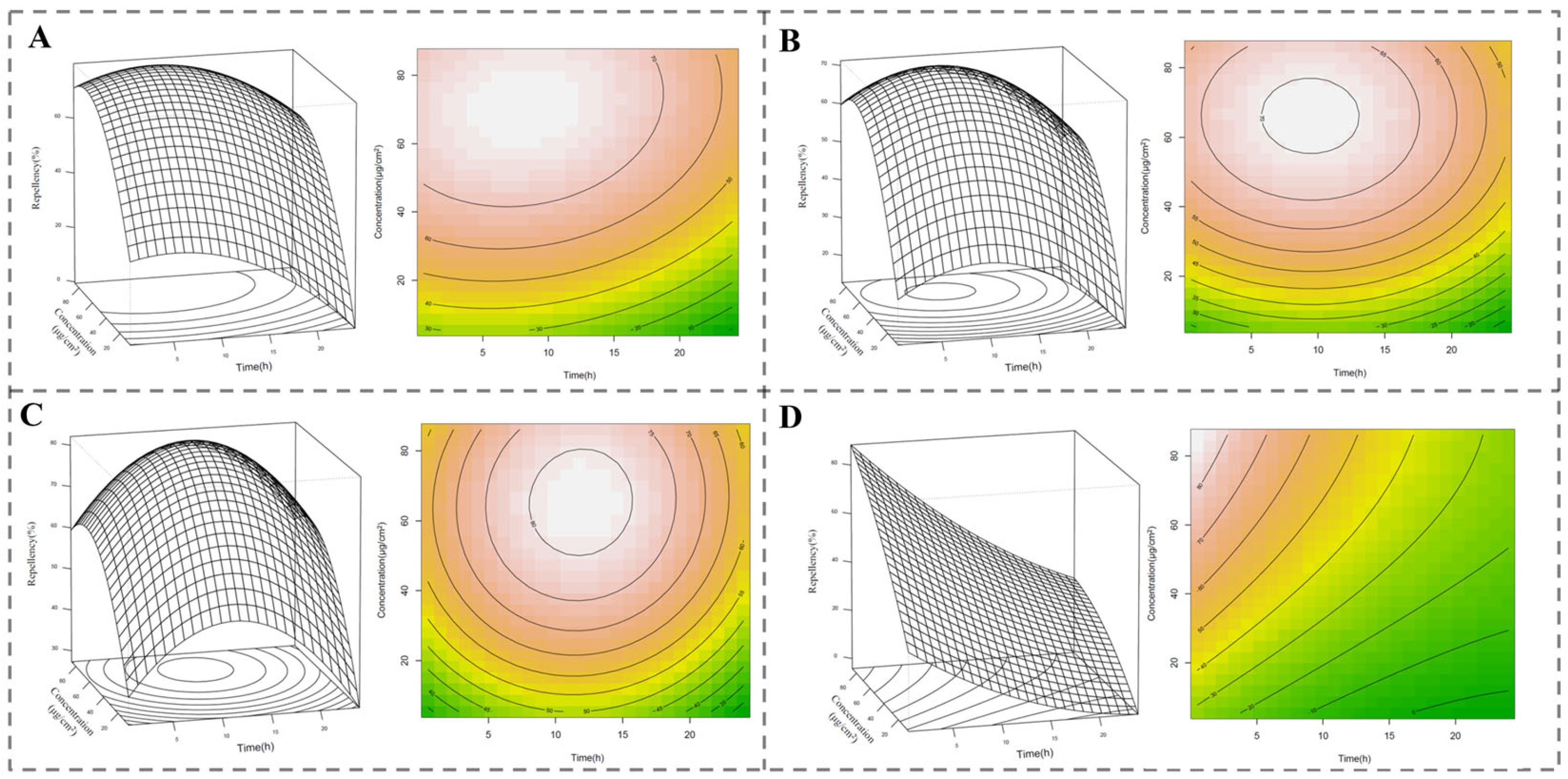
2.3. Pest-Repellent Activities of C. esculentus Root Essential Oil and the Main Compounds
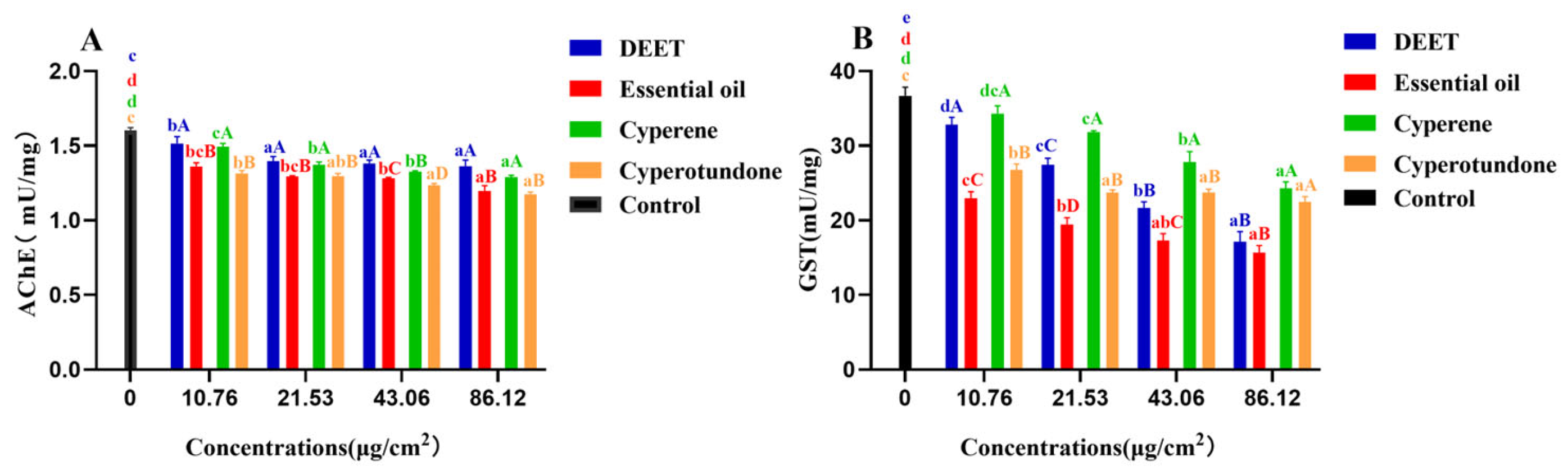
2.4. Inhibitory Activity on the Pest Enzymes
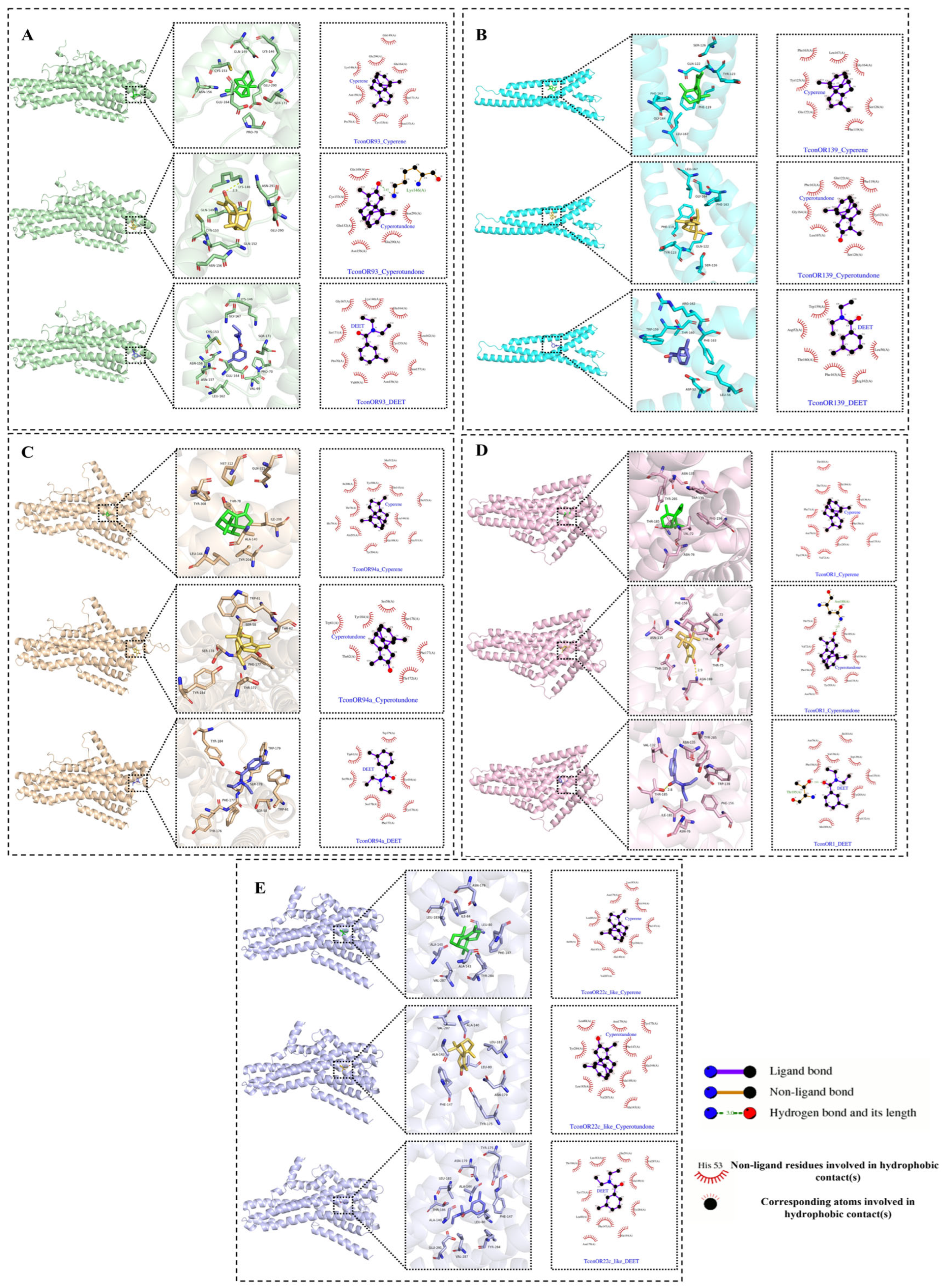
2.5. Docking Results of Main Compounds of C. esculentus Root EO with Odorant Receptor Proteins in the Pest
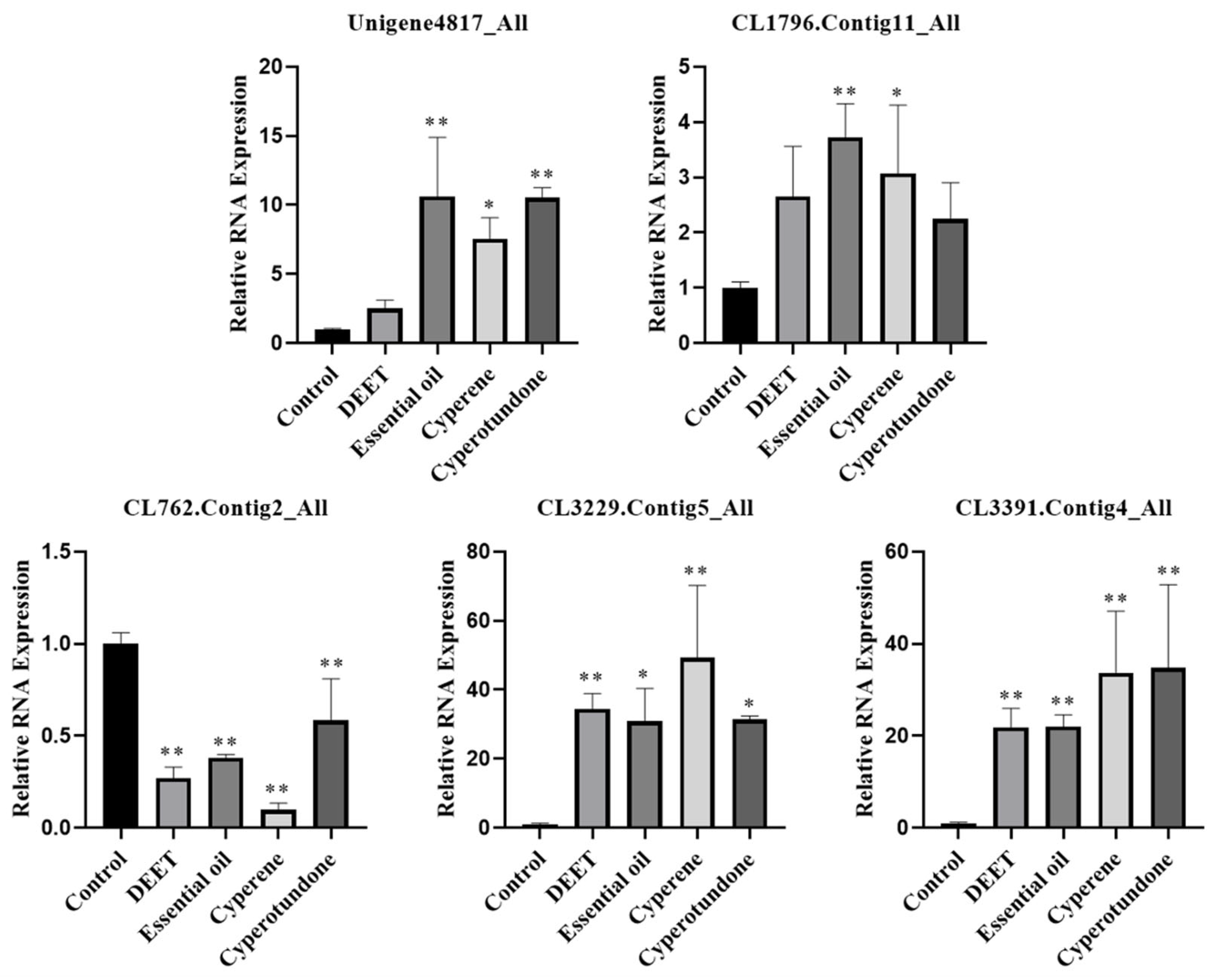
2.6. Effects on the Expression of Odorant Receptor Genes in the Pest
3. Discussion
4. Materials and Methods
4.1. Plant, Pest, Reagents, and Instruments
4.2. Extraction of the Essential Oil and Analyses of the Constituent Compounds
4.3. Isolation of the Major Chemical Constituents from the EO of C. esculentus Roots
4.4. Assessment of Pest Repellent Activity
4.5. Assessment of the Inhibitory Effects on the Pest Enzymes, Acetylcholinesterase (AChE) and Glutathione S-transferase (GST)
4.6. Molecular Docking Experiment
4.7. qRT-PCR Experiments
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Mendoza, J.; Flinn, P.W.; Campbell, J.F.; Hagstrum, D.W.; Throne, J.E. Detection of stored-grain insect infestation in wheat transported in railroad hopper-cars. J. Econ. Entomol. 2004, 97, 1474–1483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, Y.J.; Guo, W.; Luo, D.; Wu, Y.; Kučerová, Z.; Stejskal, V.; Opit, G.; Cao, Y.; Li, F.J.; et al. DNA barcoding, species-specific PCR and real-time PCR techniques for the identification of six Tribolium pests of stored products. Sci. Rep. 2016, 6, 28494. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.F.; Mullen, M.A.; Dowdy, A.K. Monitoring stored-product pests in food processing plants with pheromone trapping, contour mapping, and mark-recapture. J. Econ. Entomol. 2002, 95, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Athanassiou, C.G.; Hasan, M.M.; Phillips, T.W.; Aikins, M.J.; Throne, J.E. Efficacy of methyl bromide for control of different life stages of stored-product psocids. J. Econ. Entomol. 2015, 108, 1422–1428. [Google Scholar] [CrossRef]
- Liao, M.; Peng, Y.; Zhao, X.-P.; Yue, S.-L.; Huang, Y.; Cao, H.-Q. Identification of odorant receptors of Tribolium confusum in response to limonene repellent activity. Pestic. Biochem. Phys. 2023, 195, 105555. [Google Scholar] [CrossRef]
- Sekrane, F.Z.Y.; Righi, K.; Assia, F.; Madani, Z.; Karaca, I. Evaluation of the insecticidal potential of two medicinal plants and an entomopathogenic fungi against Tribolium confusum Jacquelin du Val. (Coleoptera: Tenebrionidae), a pest of stored foods. J. Sci. Food Agric. 2022, 6, 204–209. [Google Scholar] [CrossRef]
- Li, S.-G.; Li, M.-Y.; Huang, Y.-Z.; Hua, R.-M.; Lin, H.-F.; He, Y.-J.; Wei, L.-L.; Liu, Z.-Q. Fumigant activity of Illicium verum fruit extracts and their effects on the acetylcholinesterase and glutathione S-transferase activities in adult Sitophilus zeamais. J. Pest Sci. 2013, 86, 677–683. [Google Scholar] [CrossRef]
- Polatoglu, K.; Karakoc, O.C.; Yucel, Y.Y.; Gucel, S.; Demirci, B.; Baser, K.H.C.; Demirci, F. Insecticidal activity of edible Crithmum maritimum L. essential oil against Coleopteran and Lepidopteran insects. Ind. Crop. Prod. 2016, 89, 383–389. [Google Scholar] [CrossRef]
- Linssen, J.P.; Cozijnsen, J.L.; Pilnik, W. Chufa (Cyperus esculentus): A new source of dietary fibre. J. Sci. Food Agric. 1989, 49, 291–296. [Google Scholar] [CrossRef]
- Vega-Morales, T.; Mateos-Díaz, C.; Pérez-Machín, R.; Wiebe, J.; Gericke, N.P.; Alarcón, C.; Lopez-Romero, J.M. Chemical composition of industrially and laboratory processed Cyperus esculentus rhizomes. Food Chem. 2019, 297, 124896. [Google Scholar] [CrossRef]
- Ma, J.; Feng, X.; Meng, H.; Ma, Y.; Zhang, X.; Zhang, D.; Ma, C. Chemical constituents of Cyperus esculentus leaves and the protective effect against agricultural fungicide-induced hepatotoxicity. Chem. Biodivers. 2022, 19, e202200531. [Google Scholar] [CrossRef] [PubMed]
- Owon, M.A.; Osman, M.F.; Salama, M.A.; Esmail, A.I. Antioxidative and antimicrobial activities of tigernut tubers phenolic extract. J. Dairy Sci. 2013, 4, 333–342. [Google Scholar] [CrossRef]
- Jing, S.; Wang, S.; Li, Q.; Zheng, L.; Yue, L.; Fan, S.; Tao, G. Dynamic high pressure microfluidization-assisted extraction and bioactivities of Cyperus esculentus (C. esculentus L.) leaves flavonoids. Food Chem. 2016, 192, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Onuoha, N.O.; Ogbusua, N.O.; Okorie, A.N.; Ejike, C.E.C.C. Tigernut (Cyperus esculentus L.) “milk” as a potent “nutri-drink” for the prevention of acetaminophen-induced hepatotoxicity in a murine model. J. Intercult Ethnopharmacol. 2017, 6, 290–295. [Google Scholar] [CrossRef]
- Singh, R.; Gupta, H.; Aggarwal, G.; Bhattacharyya, K.; Sharma, U.; Reddy, S.E. Cyperus rotundus L.: Invasive weed plant with insecticidal potential against Aphis craccivora Koch and Planococcus lilacinus (Cockerell). Pestic Biochem. Physiol. 2024, 198, 105720. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, W.J.; Huang, D.Y.; Wang, Y.; Wei, J.Y.; Li, Z.H.; Sun, J.S.; Bai, J.F.; Tian, Z.F.; Wang, P.J.; et al. Chemical compositions and insecticidal activities of Alpinia kwangsiensis essential oil against Lasioderma serricorne. Molecules 2015, 20, 21939–21945. [Google Scholar] [CrossRef]
- Joseph-Nathan, P.; Martińez, E.; Santillan, R.L.; Wesener, J.R.; Günther, H. Two-dimensional NMR studies of cyperene. Org. Magn. Reson. 1984, 22, 308–311. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, H.W.; Wan, X.C.; Zou, Z.M. Complete assignments of (1)H and (13)C NMR data for two new sesquiterpenes from Cyperus rotundus L. Magn. Reson. Chem. 2009, 47, 527–531. [Google Scholar] [CrossRef]
- Lenth, R.V. Response-surface methods in R, using rsm. J. Stat. Softw. 2009, 32, 1–17. [Google Scholar] [CrossRef]
- Cagáň, Ľ.; Apacsová Fusková, M.; Hlávková, D.; Skoková Habuštová, O. Essential oils: Useful tools in storage-pest management. Plants 2022, 11, 3077. [Google Scholar] [CrossRef]
- Pang, X.; Almaz, B.; Qi, X.J.; Wang, Y.; Feng, Y.X.; Geng, Z.F.; Xi, C.; Du, S.S. Bioactivity of Essential Oil from Atalantia buxifolia Leaves and its Major Sesquiterpenes against Three Stored-Product Insects. J. Essent. Oil Bear. Plants 2020, 23, 38–50. [Google Scholar] [CrossRef]
- Rault, L.C.; Morrison, W.R., III; Gerken, A.R.; Bingham, G.V. Challenges in assessing repellency via the behavioral response by the global pest Tribolium castaneum to protect stored grains. Insects 2024, 15, 626. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.G.; Wei, W.; Cheng, K.; Zheng, L.L.; Ma, C.M.; Wang, Y.C. Insecticidal activity of triterpenoids and volatile oil from the stems of Tetraena mongolica. Pestic. Biochem. Physiol. 2020, 166, 104551. [Google Scholar] [CrossRef] [PubMed]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Benton, R. On the origin of smell: Odorant receptors in insects. Cell. Mol. Life Sci. 2006, 63, 1579–1585. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Suh, E.; Bohbot, J.D.; Zwiebel, L.J. Peripheral olfactory signaling in insects. Curr. Opin. Insect Sci. 2014, 6, 86–92. [Google Scholar] [CrossRef]
- Basile, S.; Badalamenti, N.; Riccobono, O.; Guarino, S.; Ilardi, V.; Bruno, M.; Peri, E. Chemical composition and evaluation of insecticidal activity of Calendula incana subsp. maritima and Laserpitium siler subsp. siculum essential oils against stored products pests. Molecules 2022, 27, 588. [Google Scholar] [CrossRef]
- Nararak, J.; Sanguanpong, U.; Sukkanon, C.; Manguin, S.; Chareonviriyaphap, T. Synergistic repellent and irritant effects of a mixture of β-caryophyllene oxide and vetiver oil against mosquito vectors. Insects 2023, 14, 773. [Google Scholar] [CrossRef]
- Janaki, S.; Zandi-Sohani, N.; Ramezani, L.; Szumny, A. Chemical composition and insecticidal efficacy of Cyperus rotundus essential oil against three stored product pests. Int. Biodeter. Biodegr. 2018, 133, 93–98. [Google Scholar] [CrossRef]
- You, C.-X.; Guo, S.-S.; Zhang, W.-J.; Geng, Z.-F.; Liang, J.-Y.; Lei, N.; Du, S.-S.; Deng, Z.-W. Chemical constituents of Murraya tetramera Huang and their repellent activity against Tribolium castaneum. Molecules 2017, 22, 1379. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Gallardo, K.; Olivero-Verbel, J.; Stashenko, E.E. Repellent activity of essential oils and some of their individual constituents against Tribolium castaneum Herbst. J. Agric. Food Chem. 2011, 59, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Zablotowicz, R.M.; Hoagland, R.E.; Locke, M.A.; Hickey, W.J. Glutathione-S-transferase activity and metabolism of glutathione conjugates by rhizosphere bacteria. Appl. Environ. Microbiol. 1995, 61, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirdita, M.; Steinegger, M.; Söding, J. MMseqs2 desktop and local web server app for fast, interactive sequence searches. Bioinformatics 2019, 35, 2856–2858. [Google Scholar] [CrossRef]
- Steinegger, M.; Meier, M.; Mirdita, M.; Vöhringer, H.; Haunsberger, S.J.; Söding, J. HH-suite3 for fast remote homology detection and deep protein annotation. BMC Bioinform. 2019, 20, 1–15. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Constituents | Structural Type | RIcalc a | RIdb b | RA c |
|---|---|---|---|---|---|
| 1 | α-Pinene | Monoterpenoid | 1023 | 1016 | 0.18 ± 0.03 |
| 2 | β-Pinene | Monoterpenoid | 1099 | 1096 | 0.56 ± 0.06 |
| 3 | Limonene | Monoterpenoid | 1188 | 1189 | 0.06 ± 0.01 |
| 4 | 1,8-Cineole | Monoterpenoid | 1199 | 1199 | 0.14 ± 0.02 |
| 5 | p-Cymene | Monoterpenoid | 1258 | 1269 | 0.10 ± 0.01 |
| 6 | Cyprotene | Sesquiterpene analog | 1428 | - | 0.42 ± 0.06 |
| 7 | α-Copaene | Sesquiterpenoid | 1485 | 1488 | 0.98 ± 0.14 |
| 8 | Cyperene | Sesquiterpenoid | 1522 | 1527 | 13.54 ± 0.17 |
| 9 | Cypera-2,4-diene | Sesquiterpenoid | 1533 | - | 0.65 ± 0.11 |
| 10 | Pinocarvone | Monoterpenoid | 1552 | - | 0.61 ± 0.03 |
| 11 | Myrtenal | Monoterpenoid | 1611 | 1626 | 0.61 ± 0.01 |
| 12 | Rotundene | Sesquiterpenoid | 1630 | 1629 | 5.30 ± 0.05 |
| 38 | Pinocarveol | Monoterpenoid | 1642 | 1651 | 1.18 ± 0.03 |
| 14 | trans-Verbenol | Monoterpenoid | 1667 | 1673 | 0.86 ± 0.04 |
| 15 | γ-Muurolene | Sesquiterpenoid | 1678 | 1681 | 0.20 ± 0.02 |
| 16 | Verbenone | Monoterpenoid | 1690 | 1701 | 0.27 ± 0.02 |
| 17 | δ-Guaiene | Sesquiterpenoid | 1707 | 1642 | 0.72 ± 0.07 |
| 18 | Muurolene | Sesquiterpenoid | 1714 | 1720 | 0.10 ± 0.04 |
| 19 | Carvone | Monoterpenoid | 1718 | 1740 | 0.35 ± 0.04 |
| 20 | Myrtenol | Monoterpenoid | 1779 | 1792 | 1.12 ± 0.07 |
| 21 | Ar-himachalen-2-ol | Sesquiterpenoid | 1788 | - | 1.09 ± 0.14 |
| 22 | Calamenene | Sesquiterpenoid | 1820 | 1832 | 0.37 ± 0.13 |
| 23 | Cyperene epoxide | Sesquiterpenoid | 1854 | - | 3.99 ± 0.14 |
| 24 | α-Calacorene | Sesquiterpenoid | 1900 | 1916 | 0.28 ± 0.08 |
| 25 | Aristolene epoxide | Sesquiterpenoid | 1920 | - | 1.76 ± 0.02 |
| 26 | Cubebol | Sesquiterpenoid | 1931 | 1914 | 0.33 ± 0.01 |
| 27 | Caryophyllene oxide | Sesquiterpenoid | 1971 | 1974 | 3.35 ± 0.18 |
| 28 | Longifolenaldehyde | Sesquiterpenoid | 2005 | - | 3.38 ± 0.04 |
| 29 | Spathulenol | Sesquiterpenoid | 2025 | 2088 | 1.77 ± 0.03 |
| 30 | 3,4,8,8-Tetramethyl-4,5,6,7,8,8a- hexahydro-1H-3a,7-methanoazulen-4-ol | Sesquiterpenoid | 2049 | - | 0.89 ± 0.06 |
| 31 | Aristolone | Sesquiterpenoid | 2087 | 2284 | 4.84 ± 0.08 |
| 32 | 15-Hydroxy-3-copaene | Sesquiterpenoid | 2121 | - | 3.06 ± 0.45 |
| 33 | Lemnalol | Sesquiterpenoid | 2136 | 2135 | 0.91 ± 0.06 |
| 34 | 6-Isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-naphthalen-2-ol | Sesquiterpenoid | 2145 | - | 2.86 ± 0.04 |
| 35 | Cyperadione | Sesquiterpenoid | 2165 | - | 2.18 ± 0.11 |
| 36 | 2H-Cycloprop [c]indene-2,3 (3ah)-dione, hexahydro-3a,7,7-trimethyl- | Sesquiterpene analog | 2190 | - | 1.64 ± 0.19 |
| 37 | Cadalene | Sesquiterpenoid | 2202 | 2200 | 0.38 ± 0.07 |
| 38 | 2,5-Di-tert-butyl-1,4-benzoquinone | Aromatic | 2207 | - | 3.54 ± 0.54 |
| 39 | Globulol | Sesquiterpenoid | 2225 | - | 1.94 ± 0.15 |
| 40 | Longiverbenone | Sesquiterpenoid | 2245 | 2254 | 2.96 ± 0.14 |
| 41 | Cyperotundone | Sesquiterpenoid | 2263 | - | 13.50 ± 0.64 |
| 42 | α-Cyperone | Sesquiterpenoid | 2271 | - | 1.21 ± 0.37 |
| 43 | γ-Gurjunenepoxide-(2) | Sesquiterpenoid | 2282 | - | 2.80 ± 0.31 |
| 44 | Zizanal | Sesquiterpenoid | 2289 | - | 1.47 ± 0.09 |
| 45 | 8-Oxo-9H-cycloisolongifolene | Sesquiterpenoid | 2308 | - | 2.54 ± 0.04 |
| 46 | 7R,8R-8-Hydroxy-4-isopropylidene-7-methylbicyclo [5.3.1]undec-1-ene | Sesquiterpenoid | 2345 | - | 1.96 ± 0.42 |
| 47 | β-Betulenol | Sesquiterpenoid | 2357 | - | 1.88 ± 0.36 |
| 48 | Costol | Sesquiterpenoid | 2420 | 2606 | 1.16 ± 0.03 |
| 49 | Aristolochene | Sesquiterpenoid | 2470 | - | 0.84 ± 0.03 |
| P1d | Phthalic Acid Diisobutyl Ester | Aromatic | 2522 | 2526 | 0.84 ± 0.28 |
| P2d | Dibutyl phthalate | Aromatic | 2677 | 2680 | 1.30 ± 0.08 |
| 50 | Palmitic acid | Fatty acid | 2896 | 2899 | 1.00 ± 0.05 |
| Concentration (μg/cm2) | Exposure Interval (h) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0.5 | 1 | 2 | 4 | 8 | 16 | 24 | ||
| Essential oil | 86.12 | 60.0 ± 7.1 a | 70.0 ± 12.3 abc | 84.0 ± 11.4 a | 80.0 ± 10.0 a | 76.0 ± 8.9 a | 68.0 ± 8.4 a | 62.0 ± 11.0 a |
| Cyperene | 86.12 | 48.0 ± 19.2 a | 56.0 ± 5.5 ac | 74.0 ± 8.9 a | 76.0 ± 11.4 a | 69.0 ± 8.9 a | 58.0 ± 13.0 ab | 48.0 ± 13.0 a |
| Cyperotundone | 86.12 | 52.0 ± 8.4 a | 60.0 ± 7.1 ac | 70.0 ± 14.1 a | 82.0 ± 14.8 a | 74.0 ± 11.4 a | 68.0 ± 16.4 a | 64.0 ± 15.2 a |
| DEET | 86.12 | 82.0 ± 8.4 b | 88.0 ± 13.0 ab | 90.0 ± 10.0 a | 78.0 ± 11.0 a | 52.0 ± 27.8 a | 38.0 ± 23.9 b | 32.0 ± 40.9 a |
| Essential oil | 43.06 | 56.0 ± 13.4 a | 64.0 ± 15.2 a | 74.0 ± 5.8 a | 78.0 ± 13.0 a | 66.0 ± 8.9 a | 54.0 ± 11.4 ab | 44.0 ± 15.2 a |
| Cyperene | 43.06 | 44.0 ± 8.9 a | 52.0 ± 11.0 a | 68.0 ± 13.0 a | 64.0 ± 16.7 a | 56.0 ± 15.2 a | 52.0 ± 8.4 ab | 44.0 ± 16.7 a |
| Cyperotundone | 43.06 | 48.0 ± 21.7 a | 56.0 ± 11.4 a | 62.0 ± 11.0 a | 80.0 ± 15.8 a | 60.0 ± 12.3 a | 64.0 ± 15.2 a | 54.0 ± 15.2 a |
| DEET | 43.06 | 60.0 ± 7.1 a | 68.0 ± 8.4 a | 72.0 ± 8.4 a | 64.0 ± 11.4 a | 32.0 ± 44.4 a | 26.0 ± 30.5 b | 22.0 ± 59.3 a |
| Essential oil | 21.53 | 50.0 ± 18.7 a | 56.0 ± 16.7 a | 62.0 ± 13.0 a | 66.0 ± 11.4 ab | 56.0 ± 11.4 ab | 40.0 ± 31.3 ab | 26.0 ± 30.5 a |
| Cyperene | 21.53 | 36.0 ± 5.48 a | 48.0 ± 13.0 a | 44.0 ± 16.7 a | 64.0 ± 11.4 ab | 56.0 ± 13.4 ab | 46.0 ± 8.9 ab | 42.0 ± 13.0 a |
| Cyperotundone | 21.53 | 42.0 ± 11.0 a | 58.0 ± 11.0 a | 60.0 ± 18.7 a | 70.0 ± 12.3 a | 76.0 ± 16.7 a | 66.0 ± 15.2 a | 48.0 ± 16.4 a |
| DEET | 21.53 | 24.0 ± 46.7 a | 36.0 ± 35.1 a | 44.0 ± 16.7 a | 34.0 ± 32.1 b | 26.0 ± 25.1 b | 4.0 ± 56.8 b | 2.0 ± 48.7 a |
| Essential oil | 10.76 | 24.0 ± 16.7 a | 30.0 ± 15.8 a | 38.0 ± 30.3 a | 42.0 ± 19.2 a | 32.0 ± 31.1 a | 26.0 ± 30.5 ab | 2.0 ± 51.7 a |
| Cyperene | 10.76 | 30.0 ± 14.1 a | 36.0 ± 11.4 a | 56.0 ± 13.4 a | 46.0 ± 15.2 a | 44.0 ± 16.7 a | 36.0 ± 15.1 ab | 30.0 ± 18.7 a |
| Cyperotundone | 10.76 | 32.0 ± 16.4 a | 46.0 ± 15.2 a | 48.0 ± 13.0 a | 60.0 ± 14.1 a | 68.0 ± 8.4 a | 56.0 ± 11.4 a | 44.0 ± 16.7 a |
| DEET | 10.76 | 22.0 ± 23.9 a | 30.0 ± 25.5 a | 26.0 ± 37.8 a | 38.0 ± 44.9 a | −4.0 ± 78.3 a | −2.0 ± 49.7 b | −2.0 ± 73.3 a |
| Essential oil | 5.38 | 18.0 ± 34.9 a | 30.0 ± 48.5 a | 34.0 ± 28.8 a | 24.0 ± 51.8 a | 36.0 ± 30.5 a | 22.0 ± 43.2 a | 6.0 ± 56.8 a |
| Cyperene | 5.38 | 10.0 ± 57.0 a | 14.0 ± 8.9 a | 28.0 ± 21.7 a | 28.0 ± 21.7 a | 26.0 ± 19.5 a | 12.0 ± 63.4 a | 4.0 ± 47.2 a |
| Cyperotundone | 5.38 | 20.0 ± 20.0 a | 30.0 ± 35.4 a | 38.0 ± 21.7 a | 44.0 ± 20.7 a | 38.0 ± 25.9 a | 28.0 ± 27.8 a | 20.0 ± 37.4 a |
| DEET | 5.38 | 22.0 ± 55.4 a | 30.0 ± 55.2 a | 20.0 ± 74.8 a | 50.0 ± 52.0 a | 10.0 ± 48.5 a | 22.0 ± 47.6 a | −8.0 ± 57.6 a |
| Protein | Compound | Binding Energy (kcal/mol) | Hydrogen Bonding Residue (Bond length) |
|---|---|---|---|
| TconOR93 | Cyperene | −7.9 | |
| TconOR93 | Cyperotundone | −7.9 | LYS-146 (2.87 Å) |
| TconOR93 | DEET | −7.0 | |
| TconOR139 | Cyperene | −7.5 | |
| TconOR139 | Cyperotundone | −8.2 | |
| TconOR139 | DEET | −6.8 | |
| TconOR94a | Cyperene | −6.9 | |
| TconOR94a | Cyperotundone | −7.2 | |
| TconOR94a | DEET | −6.0 | |
| TconOR1 | Cyperene | −7.7 | |
| TconOR1 | Cyperotundone | −7.2 | ASN-188 (2.92 Å) |
| TconOR1 | DEET | −5.7 | THR (2.81 Å) |
| TconOR22c-like | Cyperene | −6.7 | |
| TconOR22c-like | Cyperotundone | −6.6 | |
| TconOR22c-like | DEET | −5.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Shan, C.-B.; Ma, J.-N.; Ma, Y.; Li, N.; Zhang, D.-J.; Lu, Z.-Y.; Ma, C.-M. Main Chemical Components, Activity and Mechanism of Repellence of Cyperus esculentus Essential Oil Against Tribolium confusum. Molecules 2025, 30, 631. https://doi.org/10.3390/molecules30030631
Feng X, Shan C-B, Ma J-N, Ma Y, Li N, Zhang D-J, Lu Z-Y, Ma C-M. Main Chemical Components, Activity and Mechanism of Repellence of Cyperus esculentus Essential Oil Against Tribolium confusum. Molecules. 2025; 30(3):631. https://doi.org/10.3390/molecules30030631
Chicago/Turabian StyleFeng, Xu, Cheng-Bin Shan, Jian-Nan Ma, Yue Ma, Na Li, De-Jian Zhang, Zhan-Yuan Lu, and Chao-Mei Ma. 2025. "Main Chemical Components, Activity and Mechanism of Repellence of Cyperus esculentus Essential Oil Against Tribolium confusum" Molecules 30, no. 3: 631. https://doi.org/10.3390/molecules30030631
APA StyleFeng, X., Shan, C.-B., Ma, J.-N., Ma, Y., Li, N., Zhang, D.-J., Lu, Z.-Y., & Ma, C.-M. (2025). Main Chemical Components, Activity and Mechanism of Repellence of Cyperus esculentus Essential Oil Against Tribolium confusum. Molecules, 30(3), 631. https://doi.org/10.3390/molecules30030631