
Evidence of Time-Dependent Hepatic Cytotoxicity and Mitochondrial Remodelling Induced by Palmitoyl Epigallocatechin Gallate vs. Its Native (Poly)Phenol


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
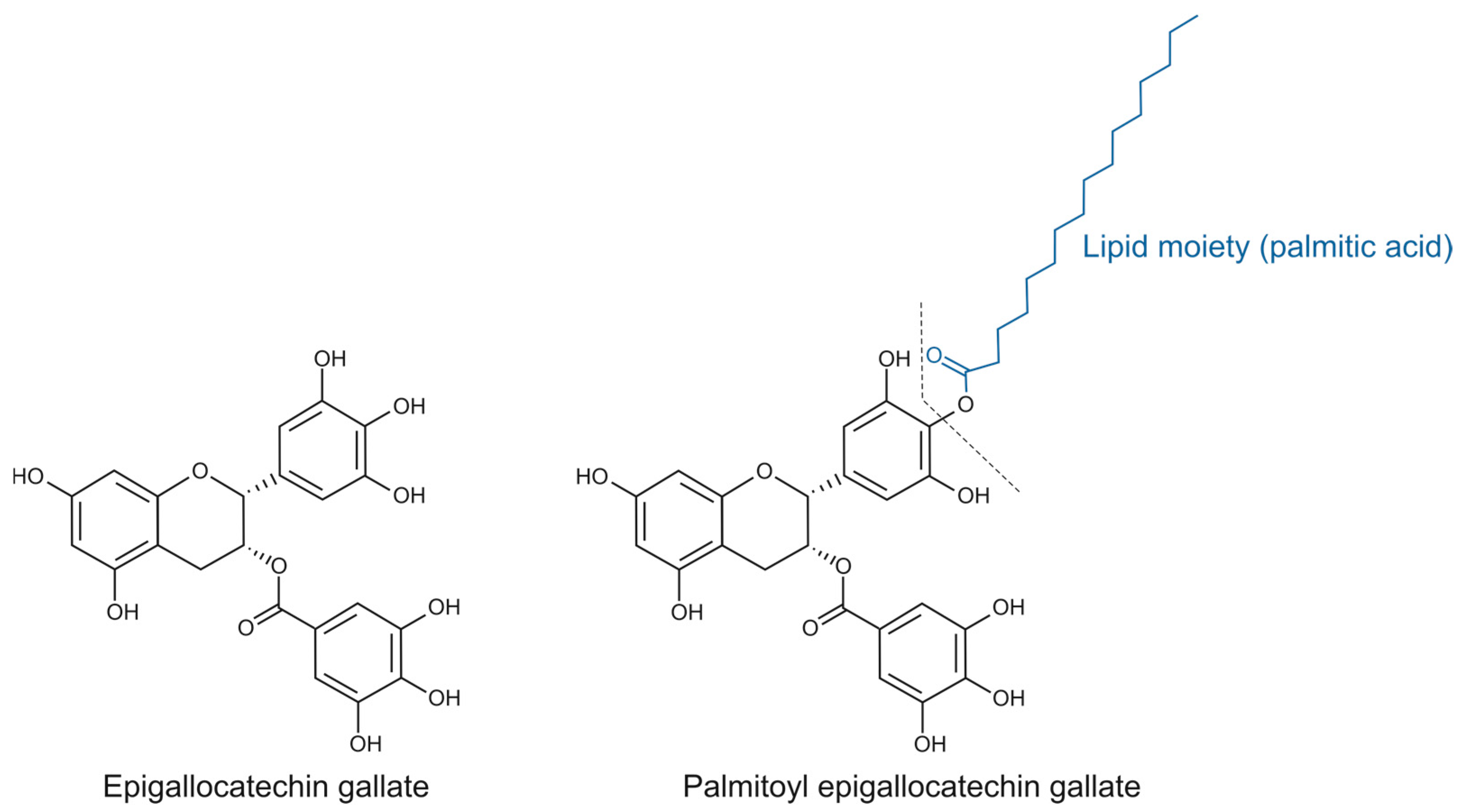
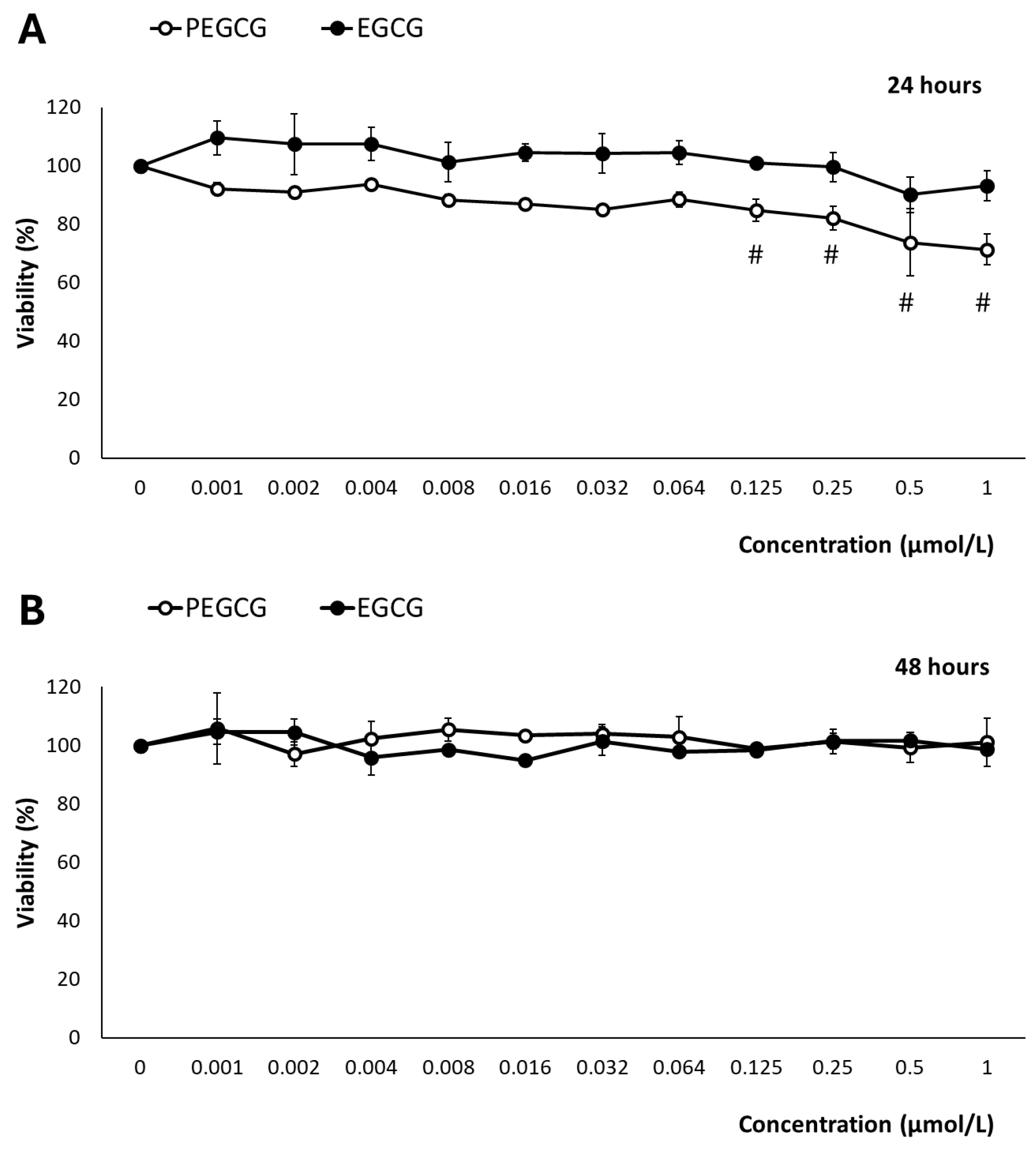
2.1. Impact of Palmitoyl Epigallocatechin Gallate and Epigallocatechin Gallate on Cell Viability
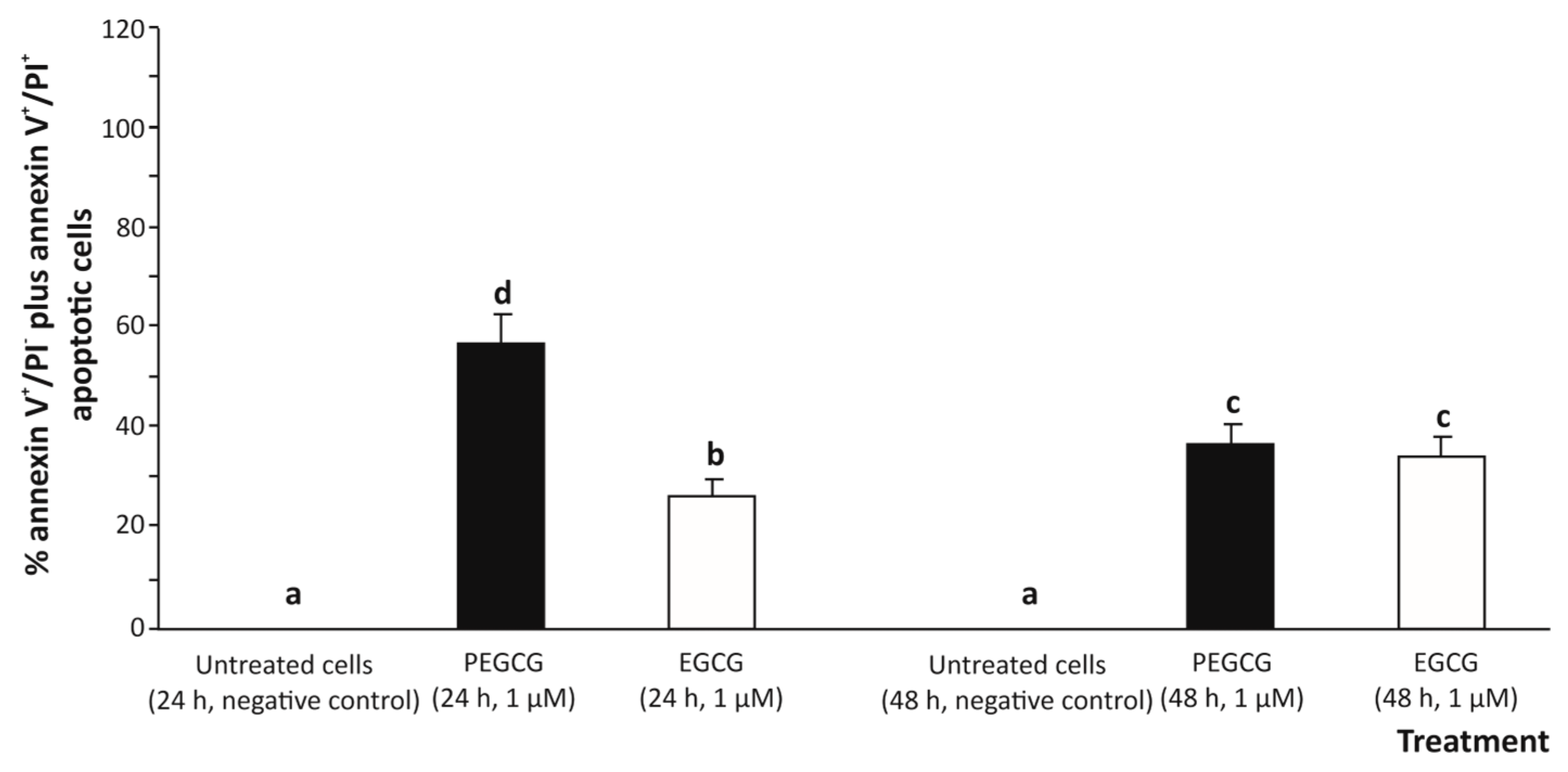
2.2. Cell Death Pathways Analysis: Apoptosis vs. Necrosis
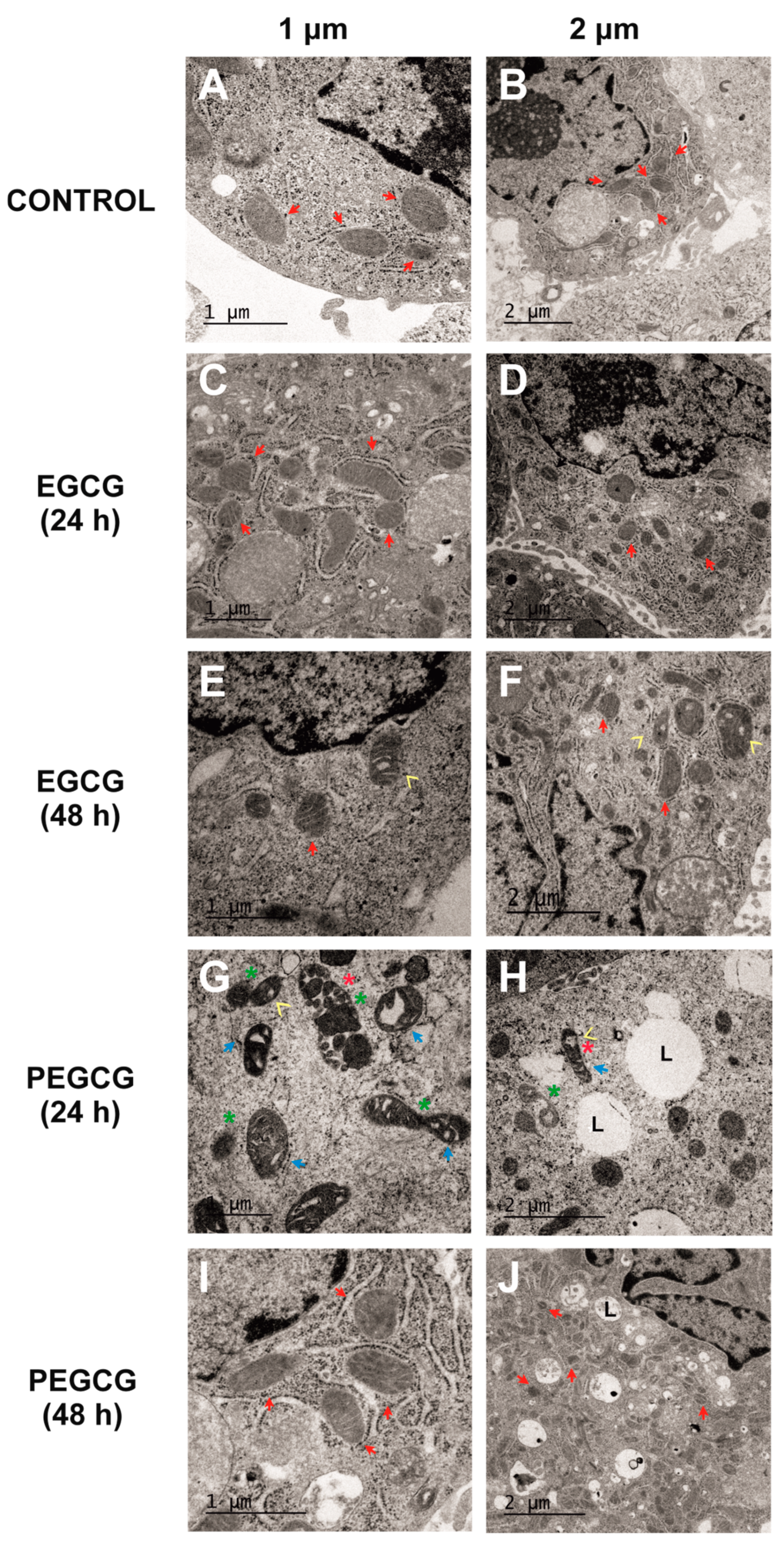
2.3. Ultrastructural Changes in Mitochondria
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Cell Line and Culture Conditions
3.3. Cytotoxicity Assay
3.4. Double-Colour Flow Cytometry Analysis of Cell Apoptosis: Evaluation of Annexin-v-FITC/Propidium Iodide
3.5. Transmission Electron Microscopic Ultrastructure Analysis of EGCG and PEGCG-Treated HepG2 Cells
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| COX-2 | Cyclooxygenase-2 |
| EGCG | Epigallocatechin gallate |
| EMEM | Eagle’s Minimum Essential Medium |
| EDTA | Ethylenediaminetetraacetic acid |
| FBS | Fetal bovine serum |
| FITC | Fluoresceine isothiocyanate |
| HepG2 | Liver hepatocarcinoma cell line |
| MTS | ((3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium salt) |
| PEGCG | Palmitoyl-epigallocatechin gallate |
| PI | Propidium iodide |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| TEM | Transmission electron microscopy |
References
- Crauste, C.; Rosell, M.; Durand, T.; Vercauteren, J. Omega-3 Polyunsaturated Lipophenols, How and Why? Biochimie 2016, 120, 62–74. [Google Scholar] [CrossRef]
- Ramadan, M.F. Chemistry and Functionality of Lipo-Phenolics. In Pheno-Phospholipids Lipo-Phenolics, 1st ed.; Ramadán, M.F., Ed.; Springer: Cham, Switzerland, 2021; Volume 1, pp. 35–63. [Google Scholar]
- Zhuang, Y.; Quan, W.; Wang, X.; Cheng, Y.; Jiao, Y. Comprehensive Review of EGCG Modification: Esterification Methods and Their Impacts on Biological Activities. Foods 2024, 13, 1232. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic Effects of Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 340. [Google Scholar] [CrossRef]
- Liu, B.; Kang, Z.; Yan, W. Synthesis, Stability, and Antidiabetic Activity Evaluation of (−)-Epigallocatechin Gallate (EGCG) Palmitate Derived from Natural Tea Polyphenols. Molecules 2021, 26, 393. [Google Scholar] [CrossRef]
- Zhong, Y.; Chiou, Y.S.; Pan, M.H.; Shahidi, F. Anti-Inflammatory Activity of Lipophilic Epigallocatechin Gallate (EGCG) Derivatives in LPS-Stimulated Murine Macrophages. Food Chem. 2012, 134, 742–748. [Google Scholar] [CrossRef]
- Hu, J.; Webster, D.; Cao, J.; Shao, A. The Safety of Green Tea and Green Tea Extract Consumption in Adults—Results of a Systematic Review. Reg. Toxicol. Pharmacol. 2018, 95, 412–433. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; Nabavi, S.F.; Daglia, M.; Rastrelli, L.; Nabavi, S.M. Epigallocatechin Gallate and Mitochondria—A Story of Life and Death. Pharmacol. Res. 2016, 104, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef]
- Shi, W.; Li, L.; Ding, Y.; Yang, K.; Chen, Z.; Fan, X.; Jiang, S.; Guan, Y.; Liu, Z.; Xu, D.; et al. The Critical Role of Epigallocatechin Gallate in Regulating Mitochondrial Metabolism. Future Med. Chem. 2018, 10, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, R.; Ito, K.; Ishida, T.; Hamanoue, M.; Adachi, S.; Watanabe, T.; Sato, Y. Catechin, Green Tea Component, Causes Caspase-Independent Necrosis-like Cell Death in Chronic Myelogenous Leukemia. Cancer Sci. 2009, 100, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Bimonte, S.; Albino, V.; Piccirillo, M.; Nasto, A.; Molino, C.; Palaia, R.; Cascella, M. Epigallocatechin-3-Gallate in the Prevention and Treatment of Hepatocellular Carcinoma: Experimental Findings and Translational Perspectives. Drug Des. Dev. Ther. 2019, 13, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Salker, M.S.; Lang, F. Green Tea Polyphenol-Sensitive Calcium Signaling in Immune T Cell Function. Front. Nutr. 2021, 7, 616934. [Google Scholar] [CrossRef]
- Landis-Piwowar, K.R.; Huo, C.; Chen, D.; Milacic, V.; Shi, G.; Tak, H.C.; Dou, Q.P. A Novel Prodrug of the Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate as a Potential Anticancer Agent. Cancer Res. 2007, 67, 4303–4310. [Google Scholar] [CrossRef] [PubMed]
- Kucera, O.; Mezera, V.; Moravcova, A.; Endlicher, R.; Lotkova, H.; Drahota, Z.; Cervinkova, Z. In Vitro Toxicity of Epigallocatechin Gallate in Rat Liver Mitochondria and Hepatocytes. Oxidative Med. Cell. Longev. 2015, 2015, 476180. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.N.; Khosravi, M.; Rahman, H.S.; Adili, A.; Kamali, N.; Soloshenkov, P.P.; Thangavelu, L.; Saeedi, H.; Shomali, N.; Tamjidifar, R.; et al. Anastasis: Cell Recovery Mechanisms and Potential Role in Cancer. Cell Commun. Signal. 2022, 20, 81. [Google Scholar] [CrossRef]
- McDonald, P.C.; Nagel, J.M.; Dedhar, S. Anastasis, Recovery from the Brink of Death as a Mechanism of Drug Resistance. In Biological Mechanisms and the Advancing Approaches to Overcoming Cancer Drug Resistance; Academic Press: Cambridge, MA, USA, 2021; pp. 251–260. [Google Scholar]
- Patil, A.A.; Bhor, S.A.; Rhee, W.J. Cell Death in Culture: Molecular Mechanisms, Detections, and Inhibition Strategies. J. Ind. Eng. Chem. 2020, 91, 37–53. [Google Scholar] [CrossRef]
- Brauchle, E.; Thude, S.; Brucker, S.Y.; Schenke-Layland, K. Cell Death Stages in Single Apoptotic and Necrotic Cells Monitored by Raman Microspectroscopy. Sci. Rep. 2014, 4, 4698. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green Tea Catechin, Epigallocatechin-3-Gallate (EGCG): Mechanisms, Perspectives and Clinical Applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Tang, H.M.; Talbot, C.C.; Fung, M.C.; Tang, H.L. Molecular Signature of Anastasis for Reversal of Apoptosis. F1000Research 2017, 6, 43. [Google Scholar] [CrossRef]
- Plecitá-Hlavatá, L.; Engstová, H.; Alán, L.M.; Špaček, T.; Dlasková, A.; Smolková, K.; Špačková, J.; Tauber, J.; Strádalová, V.; Malínský, J.; et al. Hypoxic HepG2 Cell Adaptation Decreases ATP Synthase Dimers and ATP Production in Inflated Cristae by Mitofilin Down-Regulation Concomitant to MICOS Clustering. FASEB J. 2016, 30, 1941–1957. [Google Scholar] [CrossRef]
- Murakami, A. Dose-Dependent Functionality and Toxicity of Green Tea Polyphenols in Experimental Rodents. Arch. Biochem. Biophys. 2014, 557, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, M.; Scorrano, L. The Changing Shape of Mitochondrial Apoptosis. Trend Endocrin. Met. 2009, 20, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Long, L.H.; Kirkland, D.; Whitwell, J.; Halliwell, B. Different Cytotoxic and Clastogenic Effects of Epigallocatechin Gallate in Various Cell-Culture Media Due to Variable Rates of Its Oxidation in the Culture Medium. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2007, 634, 177–183. [Google Scholar] [CrossRef]
- Ramírez-Sánchez, I.; Rodríguez, A.; Moreno-Ulloa, A.; Ceballos, G.; Villarreal, F. (−)-Epicatechin-Induced Recovery of Mitochondria from Simulated Diabetes: Potential Role of Endothelial Nitric Oxide Synthase. Diab. Vasc. Dis. Res. 2016, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Medrano-Padial, C.; Puerto, M.; del Mar Merchan-Gragero, M.; Moreno, F.J.; Richard, T.; Cantos-Villar, E.; Pichardo, S. Cytotoxicity Studies of a Stilbene Extract and Its Main Components Intended to Be Used as Preservative in the Wine Industry. Food Res. Int. 2020, 137, 109738. [Google Scholar] [CrossRef]
- Domínguez-Perles, R.; Guedes, A.; Queiroz, M.; Silva, A.M.; Barros, A.I.R.N.A. Oxidative Stress Prevention and Anti-Apoptosis Activity of Grape (Vitis vinifera L.) Stems in Human Keratinocytes. Food Res. Int. 2016, 87, 92–102. [Google Scholar] [CrossRef]
- Medrano-Padial, C.; Puerto, M.; Moreno, F.J.; Richard, T.; Cantos-Villar, E.; Pichardo, S. In Vitro Toxicity Assessment of Stilbene Extract for Its Potential Use as Antioxidant in the Wine Industry. Antioxidants 2019, 8, 467. [Google Scholar] [CrossRef]
- Graham, L.; Orenstein, J.M. Processing Tissue and Cells for Transmission Electron Microscopy in Diagnostic Pathology and Research. Nat. Protoc. 2007, 2, 2439. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medrano-Padial, C.; García-Viguera, C.; Domínguez-Perles, R.; Medina, S. Evidence of Time-Dependent Hepatic Cytotoxicity and Mitochondrial Remodelling Induced by Palmitoyl Epigallocatechin Gallate vs. Its Native (Poly)Phenol. Molecules 2025, 30, 2889. https://doi.org/10.3390/molecules30132889
Medrano-Padial C, García-Viguera C, Domínguez-Perles R, Medina S. Evidence of Time-Dependent Hepatic Cytotoxicity and Mitochondrial Remodelling Induced by Palmitoyl Epigallocatechin Gallate vs. Its Native (Poly)Phenol. Molecules. 2025; 30(13):2889. https://doi.org/10.3390/molecules30132889
Chicago/Turabian StyleMedrano-Padial, Concepción, Cristina García-Viguera, Raúl Domínguez-Perles, and Sonia Medina. 2025. "Evidence of Time-Dependent Hepatic Cytotoxicity and Mitochondrial Remodelling Induced by Palmitoyl Epigallocatechin Gallate vs. Its Native (Poly)Phenol" Molecules 30, no. 13: 2889. https://doi.org/10.3390/molecules30132889
APA StyleMedrano-Padial, C., García-Viguera, C., Domínguez-Perles, R., & Medina, S. (2025). Evidence of Time-Dependent Hepatic Cytotoxicity and Mitochondrial Remodelling Induced by Palmitoyl Epigallocatechin Gallate vs. Its Native (Poly)Phenol. Molecules, 30(13), 2889. https://doi.org/10.3390/molecules30132889