
Chemical Profiles and Biological Activities of Essential Oil from Serissa japonica


Abstract
1. Introduction
2. Results
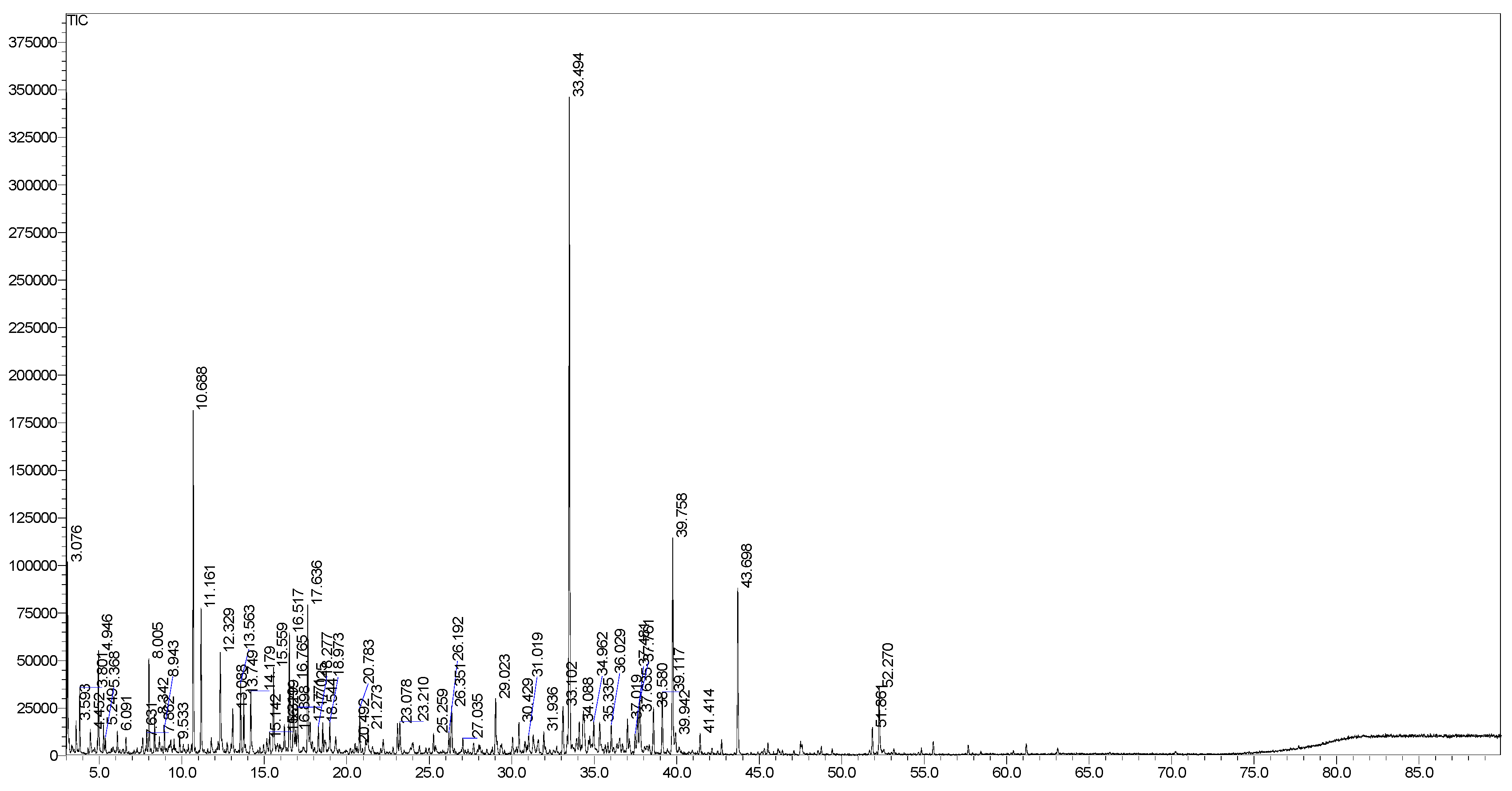
2.1. GC-MS Profiles
2.2. Biological Activities of EO
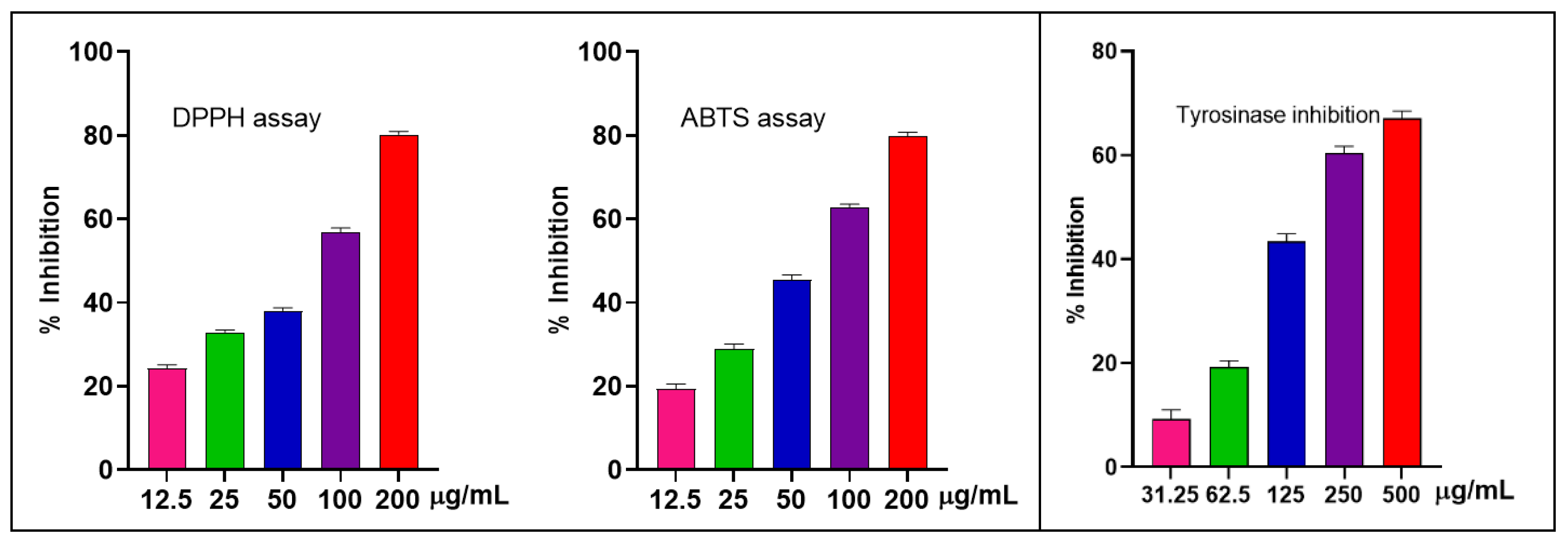
2.2.1. Antioxidant and Anti-Tyrosinase Detections
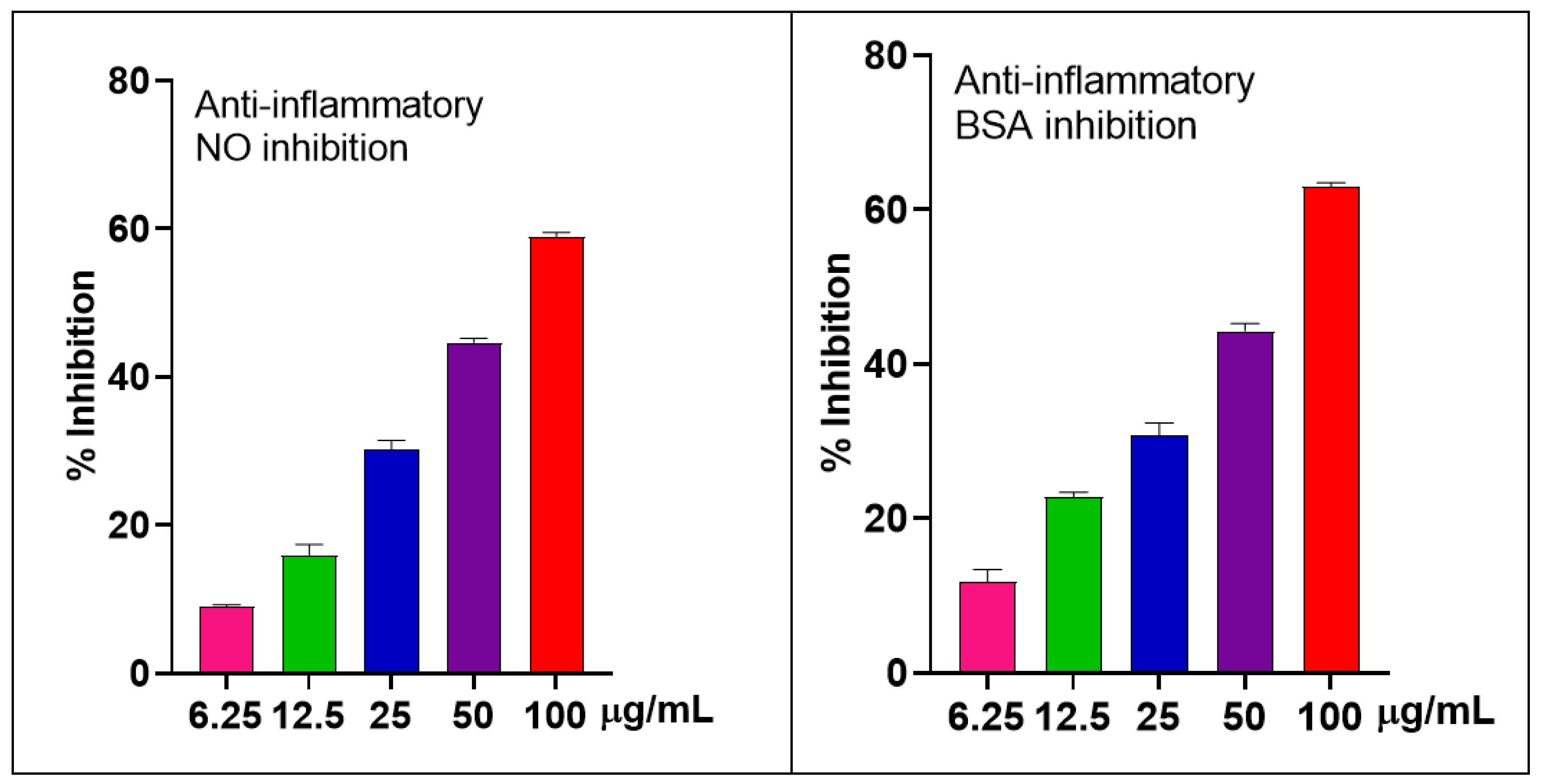
2.2.2. The Anti-Inflammatory Effects in LPS-Stimulated RAW 264.7 Macrophages and BSA
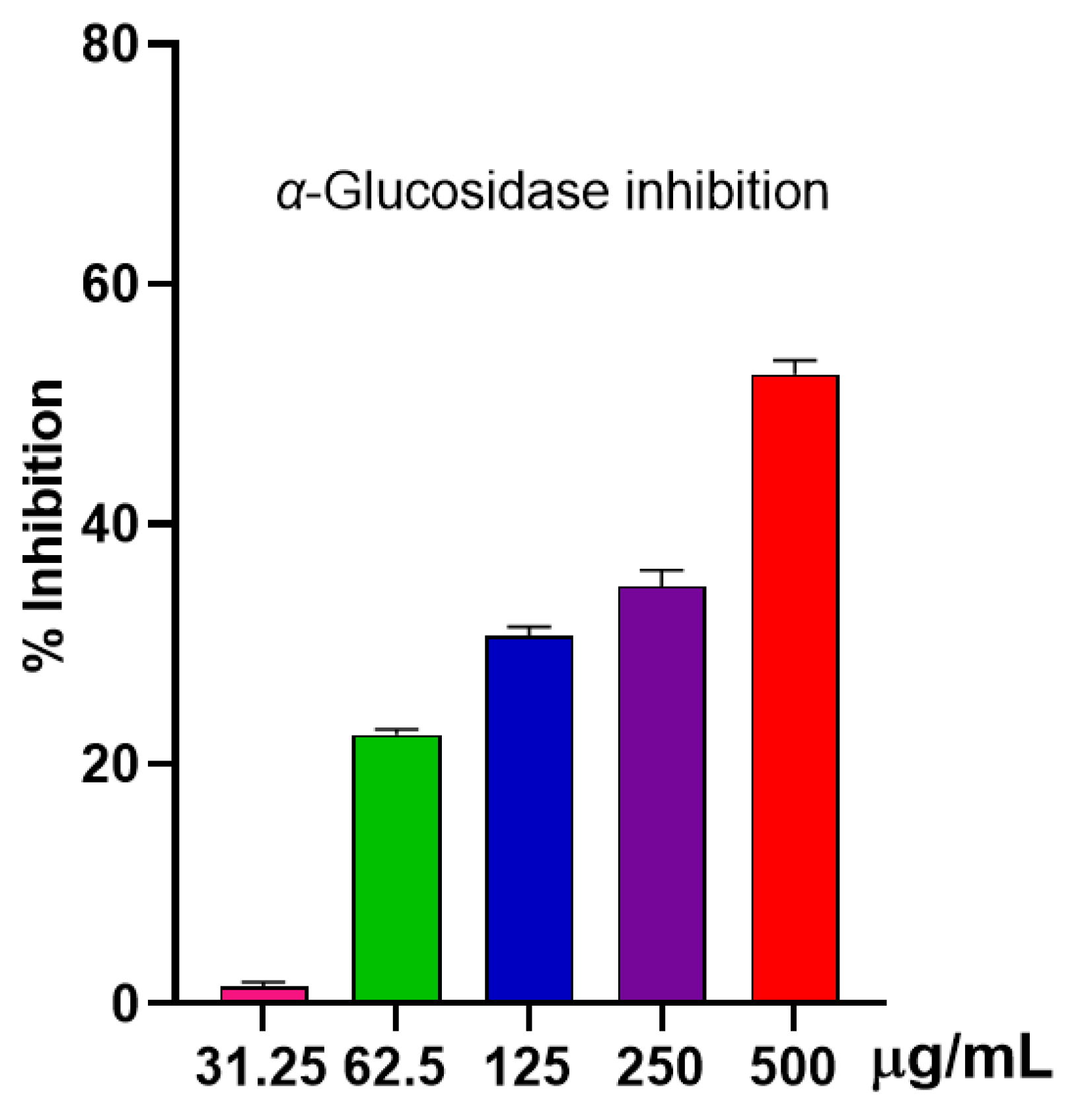
2.2.3. Anti-α-Glucosidase Activity
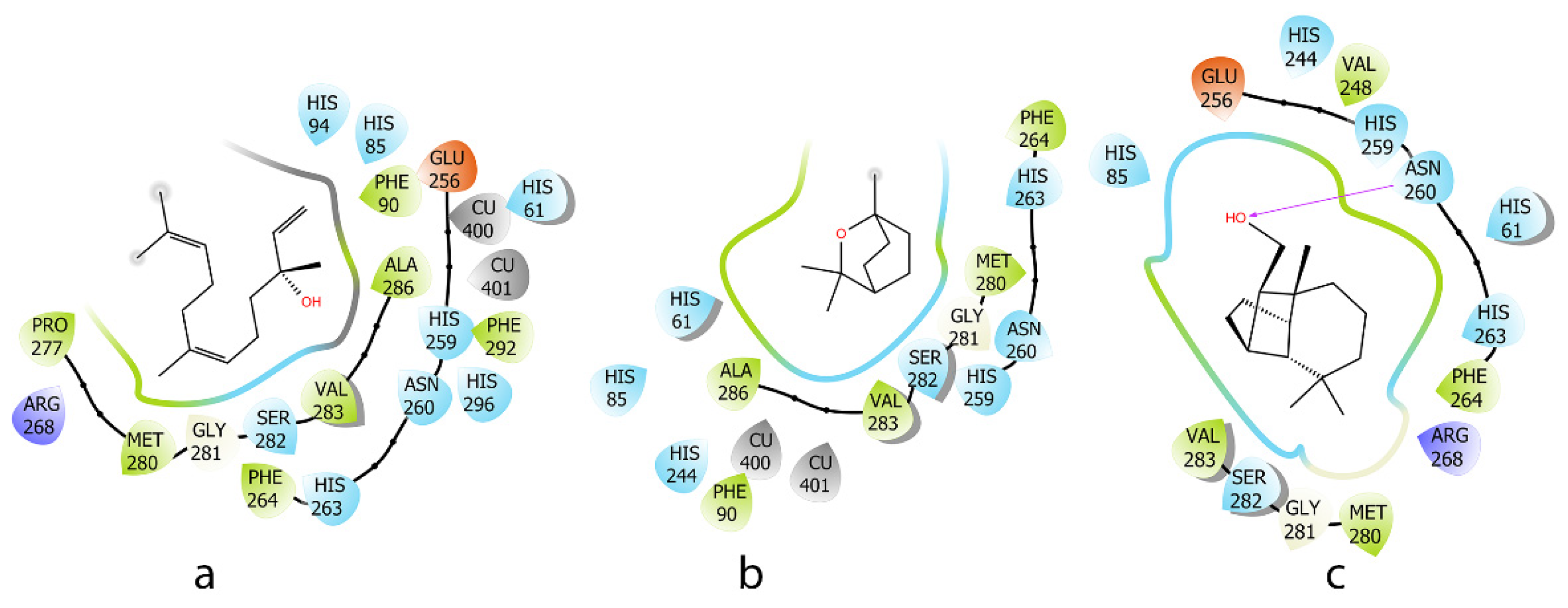
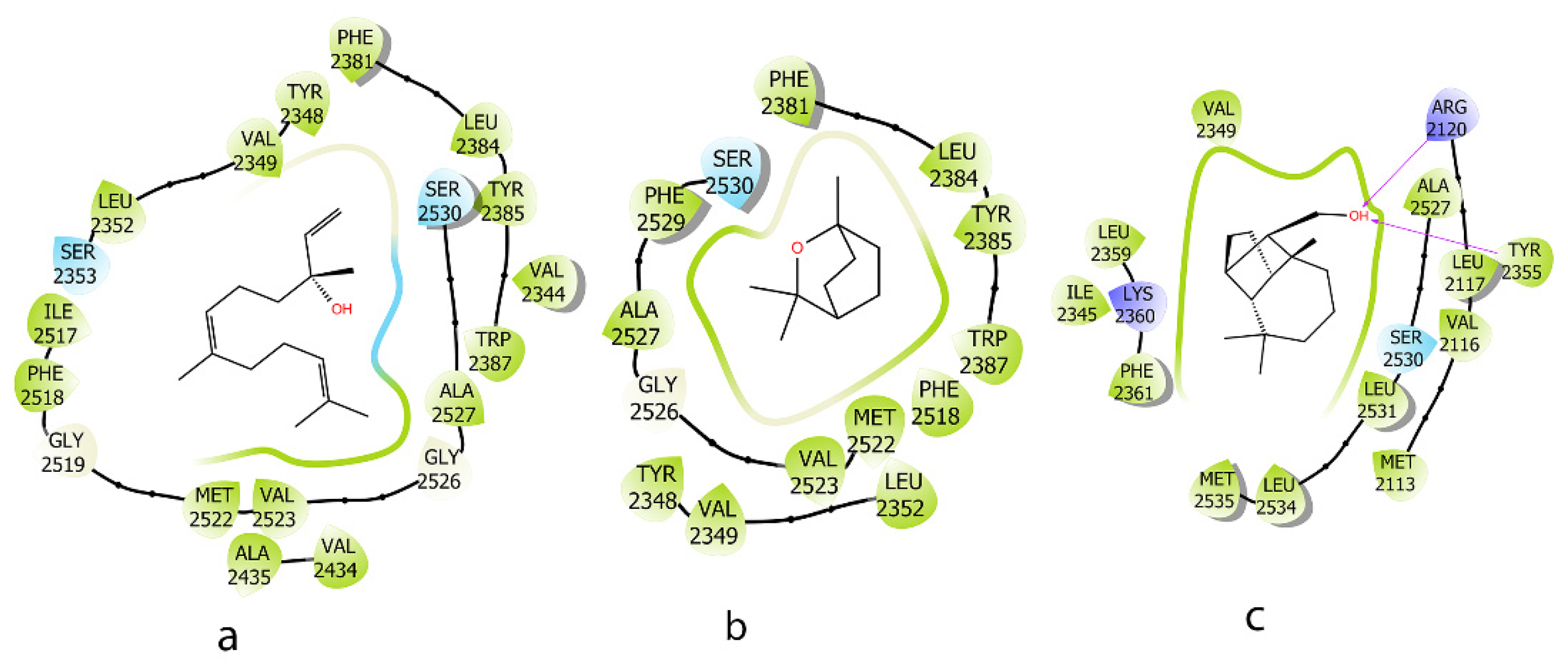
2.3. Molecular Docking Analysis
3. Discussion
Chemical Profile of EO
4. Materials and Methods
4.1. General Procedures
4.2. Plant Materials
4.3. Hydro-Distillation of EO
4.4. The GC-MS Analysis
4.5. Biological Activities
4.6. In Silico Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nguyen, H.M.; Pham, T.V.; Vo, H.Q.; Nguyen, H.T.; Nguyen, L.T.K.; Nguyen, B.C.; Chung, K.L.; Ho, D.V. Essential oil from Vietnamese Peperomia leptostachya Hook. & Arn.(Piperaceae): Chemical composition, antioxidant, anti-inflammatory, cytotoxic activities, and in silico analysis. Molecules 2024, 29, 2808. [Google Scholar] [CrossRef]
- John, R.; Manilal, A.; Varghese, L.S.; Govindarajan, S. Chemical composition, antimicrobial and antioxidant activities of Vateria indica L. oleo-gum resin (white dammar) essential oil. J. Essent. Oil Bear. Plants 2024, 27, 1516–1523. [Google Scholar] [CrossRef]
- Muscolo, A.; Mariateresa, O.; Giulio, T.; Mariateresa, R. Oxidative stress: The role of antioxidant phytochemicals in the prevention and treatment of diseases. Int. J. Mol. Sci. 2024, 25, 3264. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Moraes, F.d.S.R.; Moraes, L.H.R.; Macedo, A.B.; Valduga, A.H.; Mizobuti, D.S.; Rocha, G.L.d.; Silva, H.N.M.d.; Salvador, M.J.; Minatel, E. Anti-inflammatory and antioxidant activities of Citrus aurantifolia (Christm.) Swingle essential oil in dystrophic muscle cells: Implication of the PGC-1α pathway. J. Essent. Oil Bear. Plants 2024, 27, 34–46. [Google Scholar] [CrossRef]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative stress in ageing and chronic degenerative pathologies: Molecular mechanisms involved in counteracting oxidative stress and chronic inflammation. Int. J. Mol. Sci. 2022, 23, 7273. [Google Scholar] [CrossRef]
- Ceriello, A. Oxidative stress and diabetes-associated complications. Endocr. Pract. 2006, 12, 60–62. [Google Scholar] [CrossRef]
- González, P.; Lozano, P.; Ros, G.; Solano, F. Hyperglycemia and oxidative stress: An integral, updated and critical overview of their metabolic interconnections. Int. J. Mol. Sci. 2023, 24, 9352. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Lin, H.-R.; Yang, C.-S.; Liaw, C.-C.; Sung, P.-J.; Kuo, Y.-H.; Cheng, M.-J.; Chen, J.-J. Antioxidant and anti-α-glucosidase activities of various solvent extracts and major bioactive components from the fruits of Crataegus pinnatifida. Antioxidants 2022, 11, 320. [Google Scholar] [CrossRef]
- González-Castelazo, F.; Soria-Jasso, L.E.; Torre-Villalvazo, I.; Cariño-Cortés, R.; Muñoz-Pérez, V.M.; Ortiz, M.I.; Fernández-Martínez, E. Plants of the Rubiaceae family with effect on metabolic syndrome: Constituents, pharmacology, and molecular targets. Plants 2023, 12, 3583. [Google Scholar] [CrossRef]
- Mamun-or-Rashid, A.; Hossain, M.S.; Hassan, N.; Dash, B.K.; Sapon, M.A.; Sen, M.K. A review on medicinal plants with antidiabetic activity. J. Pharmacogn. Phytochem. 2014, 3, 149–159. [Google Scholar] [CrossRef]
- Thunberg, C.P. Nova Genera Plantarum; Upsaliæ: London, UK, 1798; Volume 9, pp. 1743–1828. [Google Scholar]
- Zhang, D.Y.; Wang, X.X.; Wang, M.; Su, B.L.; Wang, Y.X.; Li, J.Y.; Yi, J.; Chen, X.Y.; Zhuang, P.Y.; Liu, H. Chemotaxonomic significance of lignans from Serissa japonica (Thunb.) Thunb. Biochem. Syst. Ecol. 2021, 99, 104350. [Google Scholar] [CrossRef]
- Ni, S.F.; Fu, C.X.; Pan, Y.J.; Lu, Y.B.; Wu, P.; Chan, Y.S.G. Contrastive analysis of volatile oil from Serissa serissoides in different seasons. China J. Chin. Mater. Med. 2004, 28, 54–58. [Google Scholar]
- Rao, H.; Lai, P.; Gao, Y. Chemical composition, antibacterial activity, and synergistic effects with conventional antibiotics and nitric oxide production inhibitory activity of essential oil from Geophila repens (L.) IM Johnst. Molecules 2017, 22, 1561. [Google Scholar] [CrossRef]
- Tran, T.D.; Le, A.T.; Le, T.Q.; Pham, T.V. Major sesquiterpenoids in Psychotria laui leaf essential oil exhibit anti-inflammatory and cytotoxic properties. Nat. Prod. Commun. 2024, 19, 1934578X241293925. [Google Scholar] [CrossRef]
- Tava, A.; Biazzi, E.; Ronga, D.; Avato, P. Identification of the volatile components of Galium verum L. and Cruciata leavipes opiz from the western Italian Alps. Molecules 2020, 25, 2333. [Google Scholar] [CrossRef]
- Lozada-Ramírez, J.D.; Guerrero-Moras, M.C.; González-Peña, M.A.; Silva-Pereira, T.S.; Anaya de Parrodi, C.; Ortega-Regules, A.E. Stabilization of anthocyanins from coffee (Coffea arabica L.) husks and in vivo evaluation of their antioxidant activity. Molecules 2023, 28, 1353. [Google Scholar] [CrossRef]
- Syed Mohamad, S.N.A.; Khatib, A.; So’ad, S.Z.M.; Ahmed, Q.U.; Ibrahim, Z.; Nipun, T.S.; Humaryanto, H.; AlAjmi, M.F.; Khalifa, S.A.; El-Seedi, H.R. In vitro anti-diabetic, anti-inflammatory, antioxidant activities and toxicological study of optimized Psychotria malayana Jack leaves extract. Pharmaceuticals 2023, 16, 1692. [Google Scholar] [CrossRef]
- Chan, W.-K.; Tan, L.T.-H.; Chan, K.-G.; Lee, L.-H.; Goh, B.-H. Nerolidol: A sesquiterpene alcohol with multi-faceted pharmacological and biological activities. Molecules 2016, 21, 529. [Google Scholar] [CrossRef]
- Fonsêca, D.V.; Salgado, P.R.; de Carvalho, F.L.; Salvadori, M.G.S.; Penha, A.R.S.; Leite, F.C.; Borges, C.J.S.; Piuvezam, M.R.; Pordeus, L.C.d.M.; Sousa, D.P. Nerolidol exhibits antinociceptive and anti-inflammatory activity: Involvement of the GABA ergic system and proinflammatory cytokines. Fundam. Clin. Pharmacol. 2016, 30, 14–22. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Fang, H.; Yang, H.; Wu, T.; Shi, X.; Pang, C. Isolongifolene alleviates liver ischemia/reperfusion injury by regulating AMPK-PGC1α signaling pathway-mediated inflammation, apoptosis, and oxidative stress. Int. Immunopharmacol. 2022, 113, 109185. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.O.B.P.B.; Rodrigues, L.B.; Cesário, F.R.A.S.; de Oliveira, M.R.C.; Tintino, C.D.M.; e Castro, F.F.; Alcantara, I.S.; Fernandes, M.N.M.; de Albuquerque, T.R.; da Silva, M.S.A. Anti-edematogenic and anti-inflammatory activity of the essential oil from Croton rhamnifolioides leaves and its major constituent 1,8-cineole (eucalyptol). Biomed. Pharmacother. 2017, 96, 384–395. [Google Scholar] [CrossRef]
- Barone, M.; Pannuzzo, G.; Santagati, A.; Catalfo, A.; De Guidi, G.; Cardile, V. Molecular docking and fluorescence characterization of benzothieno [3,2-d] pyrimidin-4-one sulphonamide thio-derivatives, a novel class of selective cyclooxygenase-2 Inhibitors. Molecules 2014, 19, 6106–6122. [Google Scholar] [CrossRef]
- Ministry of Health. Vietnamese Pharmacopoeia; Medical Publishing House: Hanoi, Vietnam, 2017; p. 89. [Google Scholar]
- Pham, T.V.; Le, A.T.; Nguyen, N.H.; Nguyen, H.H.; Tran, G.-B.; Do, B.H. Phytochemical analysis and investigation of the anti-inflammatory and anticancerous activity of Walsura robusta leaf volatile compounds. J. Essent. Oil Bear. Plants 2025, 28, 83–94. [Google Scholar] [CrossRef]
- Pham, T.V.; Hoang, T.X.; Do, B.H.; Nguyen, K.Q.T.; Nguyen, N.H.; Tran, G.B. Chemical compositions, molecular docking, anti-inflammatory, and anti-cancer effects of the leaf essential oils isolated from three species of the Rutaceae family in Vietnam. J Chem. Biodiver. 2025, 22, e202401466. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Texensis Publishing: Gruver, TX, USA, 2007; pp. 46–52. [Google Scholar]
- Nguyen, H.M.; Nguyen, K.P.; Le, A.T.P.; Nguyen, N.H.T.; Vu-Huynh, L.K.; Le, C.K.T.; Delpe Acharige, A.; Hull, K.; Romo, D. Antioxidant, anti-tyrosinase, hepatoprotective, and anti-inflammatory potential in flowers and seeds of Ochna integerrima (Lour.) Merr. J Nat. Prod. Res. 2024, 17, 1–11. [Google Scholar] [CrossRef]
- Sakat, S.; Juvekar, A.R.; Gambhire, M.N. In vitro antioxidant and anti-inflammatory activity of methanol extract of Oxalis corniculata Linn. J Int. Pharm. Pharm. Sci. 2010, 2, 146–155. [Google Scholar]
- Shulgau, Z.; Palamarchuk, I.V.; Sergazy, S.; Urazbayeva, A.; Ramankulov, Y.; Kulakov, I.V. Synthesis, computational study, and in vitro α-Glucosidase inhibitory action of 1, 3, 4-Thiadiazole derivatives of 3-Aminopyridin-2 (1 H)-ones. Pharmaceuticals 2024, 17, 377. [Google Scholar] [CrossRef]
- Kiefer, J.R.; Pawlitz, J.L.; Moreland, K.T.; Stegeman, R.A.; Hood, W.F.; Gierse, J.K.; Stevens, A.M.; Goodwin, D.C.; Rowlinson, S.W.; Marnett, L.J. Structural insights into the stereochemistry of the cyclooxygenase reaction. Nature 2000, 405, 97–101. [Google Scholar] [CrossRef]
- Ismaya, W.T.; Rozeboom, H.J.; Weijn, A.; Mes, J.J.; Fusetti, F.; Wichers, H.J.; Dijkstra, B.W. Crystal structure of Agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Sherman, W.; Day, T.; Jacobson, M.P.; Friesner, R.A.; Farid, R. Novel procedure for modeling ligand/receptor induced fit effects. J. Med. Chem. 2006, 49, 534–553. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.; Honig, B.; Shaw, D.E.; Friesner, R.A. A hierarchical approach to all-atom protein loop prediction. Proteins Struct. Funct. Bioinf. 2004, 55, 351–367. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound a | RT | RI b | RI c | Concentrations (%) | Classification |
|---|---|---|---|---|---|---|
| 1 | 2-methyl-1,3-cyclohexadiene | 3.08 | 767 | 760 | 3.0 | NT |
| 2 | 1-octene | 3.59 | 790 | 788 | 0.6 | NT |
| 3 | (3E)-hexenal | 3.80 | 799 | 792 | 1.1 | NT |
| 4 | Furfural | 4.45 | 828 | 828 | 0.5 | NT |
| 5 | (3Z)-hexenol | 4.95 | 850 | 850 | 2.0 | NT |
| 6 | n-hexanol | 5.25 | 863 | 863 | 0.6 | NT |
| 7 | Isopentyl acetate | 5.37 | 868 | 869 | 0.3 | NT |
| 8 | n-Heptanal | 6.09 | 900 | 901 | 0.4 | NT |
| 9 | Camphene | 7.63 | 946 | 946 | 0.3 | MH |
| 10 | (2E)-heptenal | 7.86 | 953 | 947 | 0.4 | NT |
| 11 | Benzaldehyde | 8.01 | 957 | 952 | 2.0 | NT |
| 12 | n-heptanol | 8.34 | 967 | 959 | 0.8 | NT |
| 13 | 6-methyl-5-hepten-2-one | 8.94 | 985 | 981 | 0.5 | OM |
| 14 | n-Octanal | 9.53 | 1002 | 998 | 0.2 | NT |
| 15 | 1,8-Cineole | 10.69 | 1030 | 1026 | 7.3 | OM |
| 16 | Benzeneacetaldehyde | 11.16 | 1041 | 1036 | 3.3 | NT |
| 17 | n-octanol | 12.33 | 1069 | 1063 | 2.3 | NT |
| 18 | Fenchone | 13.09 | 1088 | 1083 | 1.2 | OM |
| 19 | Linalool | 13.56 | 1099 | 1095 | 1.5 | OM |
| 20 | n-nonanal | 13.75 | 1103 | 1100 | 1.3 | NT |
| 21 | endo-fenchol | 14.18 | 1113 | 1114 | 1.8 | OM |
| 22 | cis-limonene oxide | 15.15 | 1135 | 1132 | 0.4 | OM |
| 23 | trans-limonene oxide | 15.32 | 1138 | 1137 | 0.6 | OM |
| 24 | Camphor | 15.56 | 1144 | 1141 | 2.0 | OM |
| 25 | (2E)-nonen-1-al | 16.22 | 1159 | 1157 | 0.7 | NT |
| 26 | Borneol | 16.52 | 1165 | 1165 | 3.0 | OM |
| 27 | n-nonanol | 16.77 | 1171 | 1165 | 1.8 | NT |
| 28 | cis-pinocamphone | 16.90 | 1174 | 1172 | 0.6 | OM |
| 29 | Terpinen-4-ol | 17.03 | 1177 | 1174 | 1.3 | OM |
| 30 | α-terpineol | 17.64 | 1190 | 1186 | 3.6 | OM |
| 31 | Methyl salicylate | 17.77 | 1194 | 1190 | 0.9 | NT |
| 32 | n-decanal | 18.28 | 1205 | 1201 | 0.6 | NT |
| 33 | β-cyclocitral | 18.97 | 1221 | 1217 | 0.7 | OM |
| 34 | Geraniol | 20.50 | 1255 | 1249 | 0.2 | OM |
| 35 | (2E)-decenal | 20.78 | 1261 | 1260 | 1.1 | NT |
| 36 | n-decanol | 21.27 | 1272 | 1266 | 0.6 | NT |
| 37 | ρ-vinyl-guaiacol | 23.08 | 1313 | 1309 | 0.8 | NT |
| 38 | (2E,4E)-decadienal | 23.21 | 1316 | 1315 | 0.9 | NT |
| 39 | (2E)-undecenal | 25.26 | 1364 | 1357 | 0.5 | NT |
| 40 | (E)-β-damascenone | 26.19 | 1385 | 1383 | 0.5 | OS |
| 41 | Methyl decyl ketone | 26.35 | 1389 | 1388 | 1.4 | NT |
| 42 | (E)-Jasmonyl acetate | 27.04 | 1405 | 1398 | 0.4 | OS |
| 43 | Geranyl acetone | 29.02 | 1453 | 1453 | 1.4 | OM |
| 44 | (E)-β-ionone | 30.43 | 1487 | 1487 | 0.7 | OS |
| 45 | α-muurolene | 31.02 | 1502 | 1500 | 0.4 | SH |
| 46 | δ-cadinene | 31.94 | 1525 | 1522 | 0.7 | SH |
| 47 | Norpatchoulenol | 33.10 | 1555 | 1553 | 1.3 | OS |
| 48 | (E)-nerolidol | 33.49 | 1565 | 1561 | 16.1 | OS |
| 49 | trans-sesquisabinene hydrate | 34.09 | 1580 | 1577 | 0.8 | OS |
| 50 | n-Hexadecane | 34.96 | 1602 | 1600 | 0.8 | NT |
| 51 | Tetradecanal | 35.34 | 1612 | 1611 | 0.8 | NT |
| 52 | 1-epi-cubenol | 36.03 | 1631 | 1627 | 0.7 | OS |
| 53 | Valerianol | 37.02 | 1657 | 1656 | 1.0 | OS |
| 54 | Citronellyl tiglate | 37.48 | 1670 | 1666 | 0.4 | OS |
| 55 | Bulnesol | 37.64 | 1674 | 1670 | 1.3 | OS |
| 56 | n-Tetradecanol | 37.76 | 1677 | 1671 | 1.9 | NT |
| 57 | n-Heptadecane | 38.58 | 1700 | 1700 | 1.3 | NT |
| 58 | Cedroxyde | 39.12 | 1714 | 1713 | 1.6 | OS |
| 59 | iso-longifolol | 39.76 | 1733 | 1728 | 5.8 | OS |
| 60 | (E)-β-santalol | 39.93 | 1738 | 1738 | 0.8 | OS |
| 61 | n-pentadecanol | 41.41 | 1779 | 1773 | 0.5 | NT |
| 62 | Isoamyl dodecanoate | 43.70 | 1846 | 1844 | 4.3 | NT |
| 63 | n-Heneicosane | 51.86 | 2100 | 2100 | 0.8 | NT |
| 64 | cis-phytol | 52.27 | 2114 | 2113 | 2.0 | OD |
| Total | 99.4 | |||||
| Monoterpene hydrocarbons (MHs) | 0.3 | |||||
| Oxygenated monoterpenes (OMs) | 25.6 | |||||
| Sesquiterpene hydrocarbons (SHs) | 1.1 | |||||
| Oxygenated sesquiterpenes (OS) | 31.4 | |||||
| Oxygenated diterpenenes (ODs) | 2.0 | |||||
| Non-terpenic compounds (NTs) | 39.0 | |||||
| Compound | Induce Fit Docking XP (Kcal/mol) | MM-GBSA (Kcal/mol) | Interaction |
|---|---|---|---|
| a | −4.0 | −55.2 | - |
| b | −3.3 | −36.3 | - |
| c | −4.6 | −55.7 | H-Bond: ASN 260 |
| Compound | Induce Fit Docking XP (Kcal/mol) | MM-GBSA (Kcal/mol) | Interaction |
|---|---|---|---|
| a | −6.541 | −75.31 | - |
| b | −6.457 | −55.10 | - |
| c | −7.245 | −93.96 | H-Bond: ARG 2120, TYR 2355 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, T.V.; Vu, T.-Y.; Nguyen, H.M. Chemical Profiles and Biological Activities of Essential Oil from Serissa japonica. Molecules 2025, 30, 2485. https://doi.org/10.3390/molecules30122485
Pham TV, Vu T-Y, Nguyen HM. Chemical Profiles and Biological Activities of Essential Oil from Serissa japonica. Molecules. 2025; 30(12):2485. https://doi.org/10.3390/molecules30122485
Chicago/Turabian StylePham, Ty Viet, Thien-Y Vu, and Hien Minh Nguyen. 2025. "Chemical Profiles and Biological Activities of Essential Oil from Serissa japonica" Molecules 30, no. 12: 2485. https://doi.org/10.3390/molecules30122485
APA StylePham, T. V., Vu, T.-Y., & Nguyen, H. M. (2025). Chemical Profiles and Biological Activities of Essential Oil from Serissa japonica. Molecules, 30(12), 2485. https://doi.org/10.3390/molecules30122485